

Urban Heat Island and Reduced Habitat Complexity Explain Spider Community Composition by Excluding Large and Heat-Sensitive Species

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
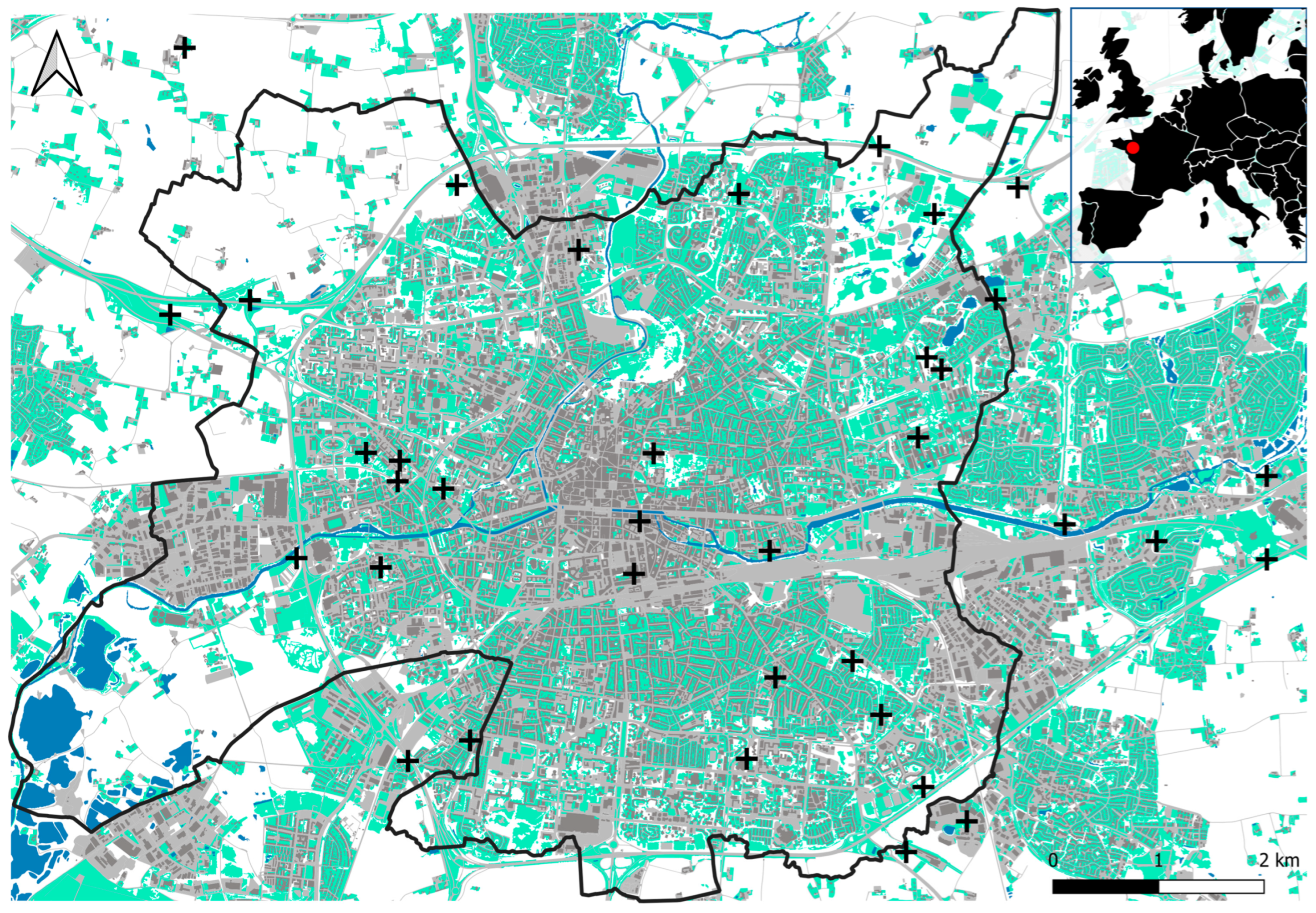
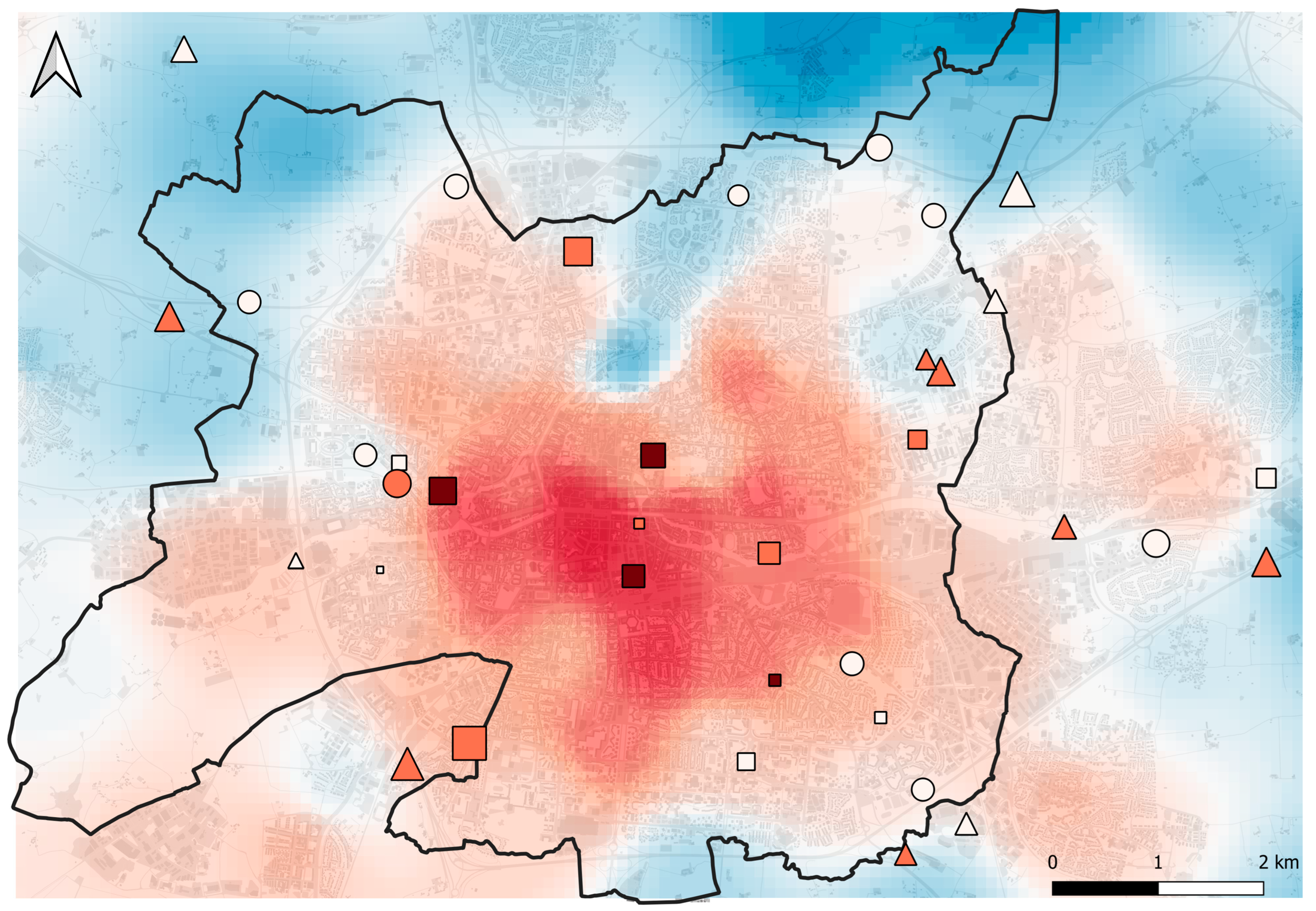
2.1. Study Area and Sampling Sites
2.2. Spider Sampling
2.3. Functional Trait Selection
2.4. Environmental Variables
2.5. Data Analyses
2.5.1. Characterization of the Urbanization Gradient
2.5.2. Spider Community Composition along an Urbanization Gradient
2.5.3. Relationships between Functional Traits and Environmental Conditions
2.5.4. Characterization of Changes in Functional Trait Community Indices
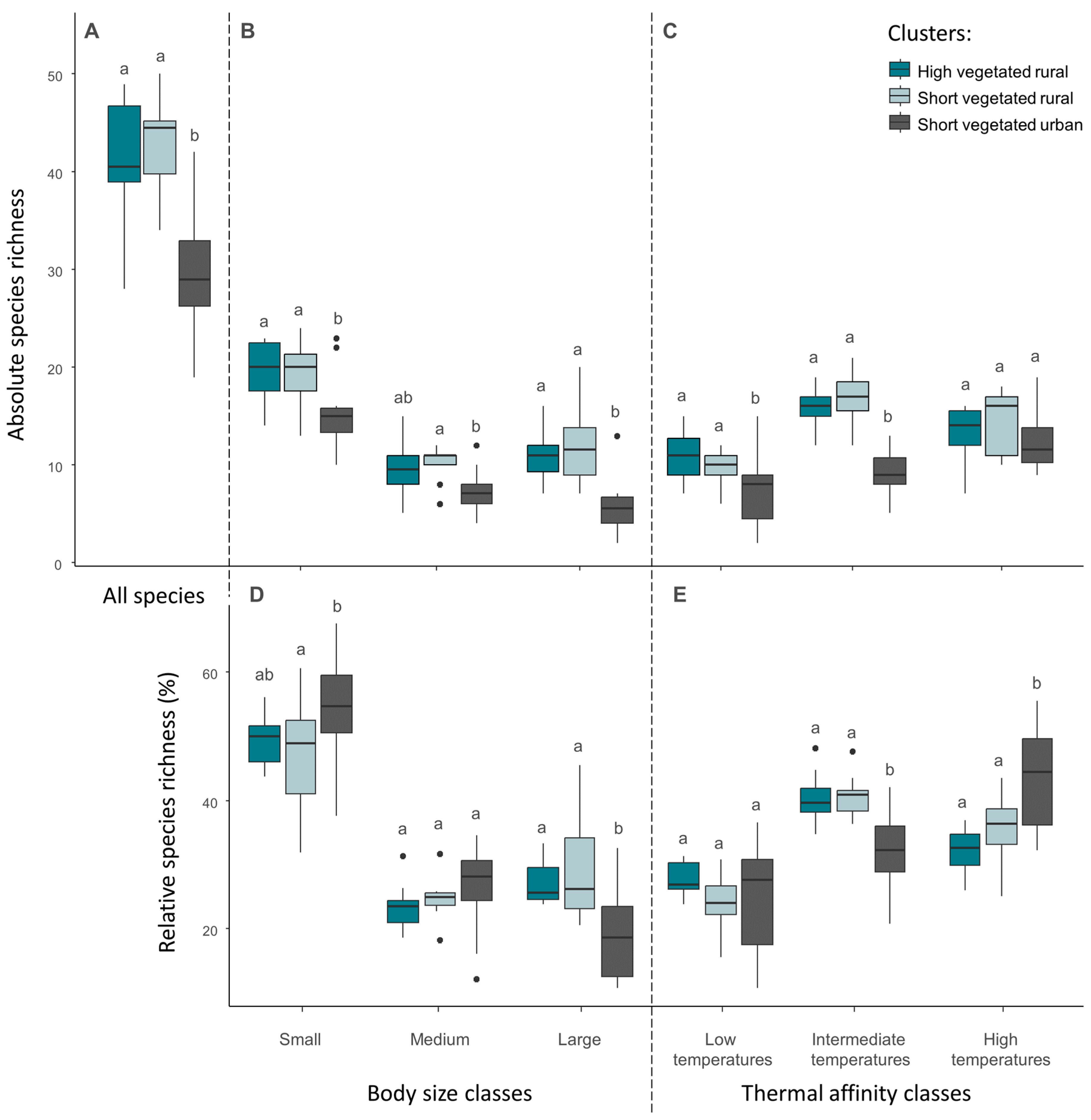
3. Results
3.1. Sampling Results and Urbanization Gradient
3.2. Spider Community Composition and Indicator Species along an Urbanization Gradient
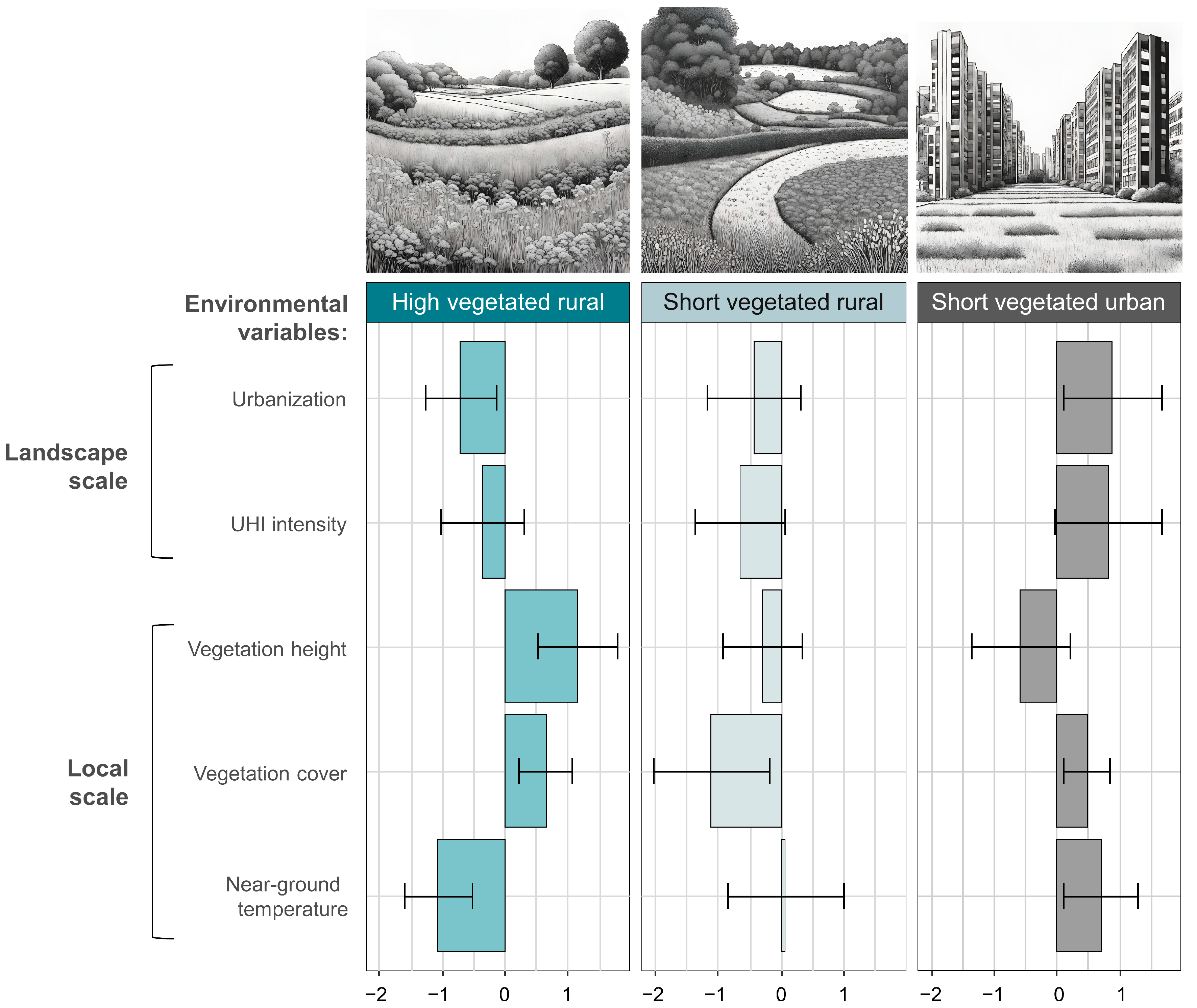
3.3. Relationships between Functional Traits and Environmental Conditions
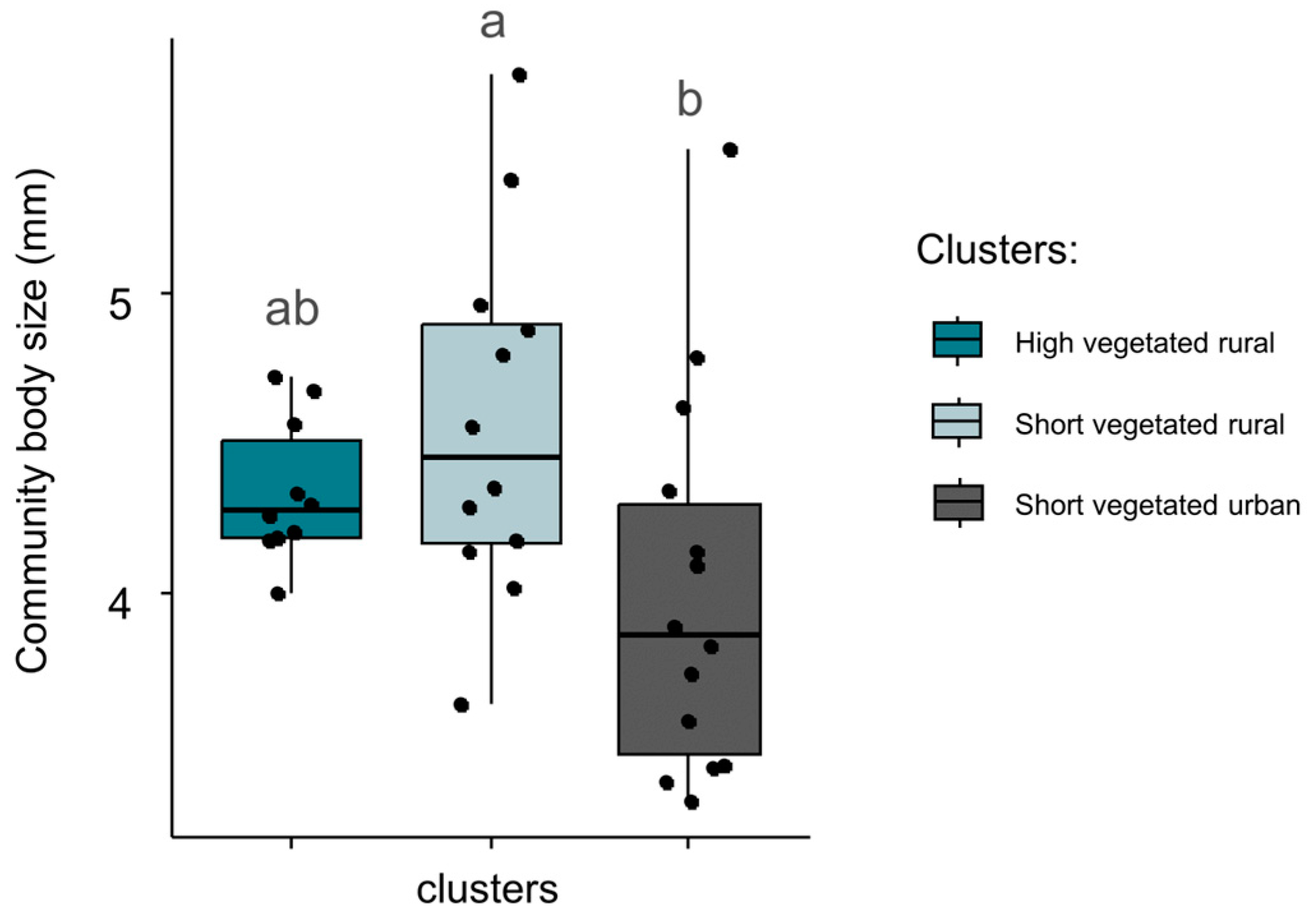
3.4. Characterization of Changes in Functional Trait Community Indices
4. Discussion
4.1. Spider Community Composition and Indicator Species along an Urbanization Gradient
4.2. Relationships between Functional Traits and Environmental Conditions
4.3. Changes in Functional Trait Community Indices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
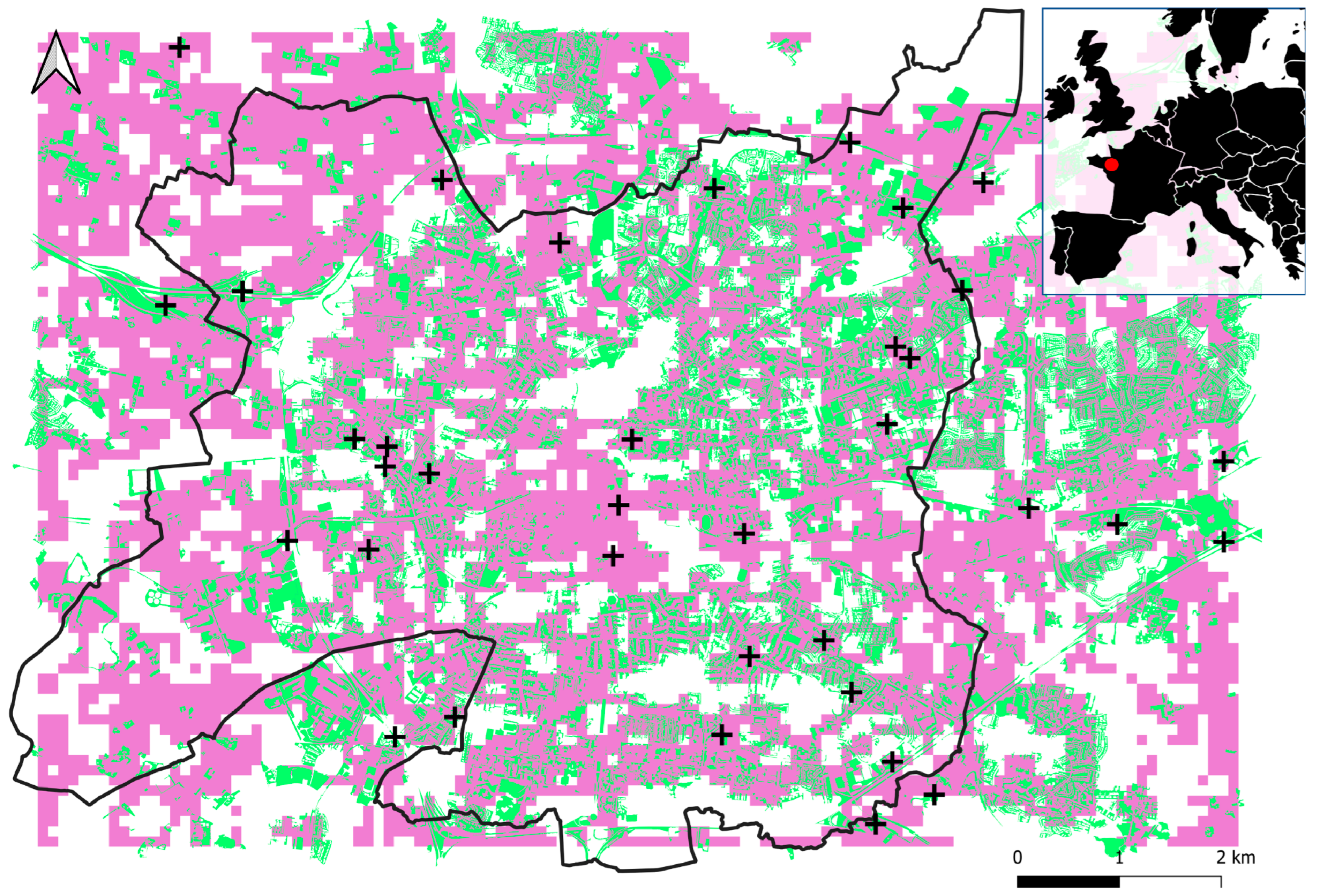
Appendix A. Sampling Site Selection Procedure
- (1)
- We performed a Spearman spatial correlation analysis to identify all pixels showing a non-significant covariation between atmospheric UHI intensity and the proportion of built-up areas. This analysis was performed using the ‘rasterCorrelation’ function from the ‘spatialEco’ package (version 1.3.7). Correlations were calculated within a 500 m square sliding window (i.e., 25 pixels), centered on each map pixel. This window size corresponds to a trade-off between a number of pixels large enough to calculate a reliable correlation (N = 25) and an area small enough (25 ha) to identify uncorrelated areas;
- (2)
- We extracted all pixels corresponding to non-significant correlations (p-value > 0.05), resulting in a raster map indicating suitable (i.e., uncorrelated) areas for sampling;
- (3)
- We selected all patches of herbaceous vegetation (excluding agricultural fields) from the Rennes land cover map that overlapped the suitable sampling area (Figure A1);
- (4)
- We identified 39 accessible sampling sites encompassing a broad gradient of built-up cover proportions and UHI intensities;
- (5)
- Once fieldwork was completed in 2022, we repeated the first four steps with updated atmospheric UHI data from 2022 corresponding to the arthropod sampling period, to check that all sampling sites were still located in suitable (i.e., uncorrelated) areas;
- (6)
- We removed three sites located in unsuitable areas, resulting in 36 remaining sampling sites.
References
- Elmqvist, T.; Fragkias, M.; Goodness, J.; Güneralp, B.; Marcotullio, P.J.; McDonald, R.I.; Parnell, S.; Schewenius, M.; Sendstad, M.; Seto, K.C. Urbanization, Biodiversity and Ecosystem Services: Challenges and Opportunities: A global Assessment; Springer Nature: Berlin/Heidelberg, Germany, 2013; ISBN 978-94-007-7087-4. [Google Scholar]
- Shwartz, A.; Turbé, A.; Julliard, R.; Simon, L.; Prévot, A.-C. Outstanding Challenges for Urban Conservation Research and Action. Glob. Environ. Chang. 2014, 28, 39–49. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in Cities Needs Space: A Meta-Analysis of Factors Determining Intra-Urban Biodiversity Variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- MacGregor-Fors, I.; Escobar, F.; Rueda-Hernández, R.; Avendaño-Reyes, S.; Baena, M.L.; Bandala, V.M.; Chacón-Zapata, S.; Guillén-Servent, A.; González-García, F.; Lorea-Hernández, F.; et al. City “Green” Contributions: The Role of Urban Greenspaces as Reservoirs for Biodiversity. Forests 2016, 7, 146. [Google Scholar] [CrossRef]
- Buchholz, S.; Hannig, K.; Moller, M.; Schirmel, J. Reducing Management Intensity and Isolation as Promising Tools to Enhance Ground-Dwelling Arthropod Diversity in Urban Grasslands. Urban Ecosyst. 2018, 21, 1139–1149. [Google Scholar] [CrossRef]
- McIntyre, N.E. Wildlife Responses to Urbanization: Patterns of Diversity and Community Structure in Built Environments. In Urban Wildlife Conservation: Theory and Practice; McCleery, R.A., Moorman, C.E., Peterson, M.N., Eds.; Springer: Boston, MA, USA, 2014; pp. 103–115. ISBN 978-1-4899-7500-3. [Google Scholar]
- Piano, E.; De Wolf, K.; Bona, F.; Bonte, D.; Bowler, D.E.; Isaia, M.; Lens, L.; Merckx, T.; Mertens, D.; Van Kerckvoorde, M.; et al. Urbanization Drives Community Shifts towards Thermophilic and Dispersive Species at Local and Landscape Scales. Glob. Chang. Biol. 2017, 23, 2554–2564. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, S.; Gathof, A.K.; Grossmann, A.J.; Kowarik, I.; Fischer, L.K. Wild Bees in Urban Grasslands: Urbanisation, Functional Diversity and Species Traits. Landsc. Urban Plan. 2020, 196, 103731. [Google Scholar] [CrossRef]
- Sattler, T.; Borcard, D.; Arlettaz, R.; Bontadina, F.; Legendre, P.; Obrist, M.; Moretti, M. Spider, Bee, and Bird Communities in Cities Are Shaped by Environmental Control and High Stochasticity. Ecology 2010, 91, 3343–3353. [Google Scholar] [CrossRef] [PubMed]
- Philpott, S.; Cotton, J.; Bichier, P.; Friedrich, R.; Moorhead, L.; Uno, S.; Valdez, M. Local and landscape drivers of arthropod abundance, richness, and trophic composition in urban habitats. Urban Ecosyst. 2014, 17, 513–532. [Google Scholar] [CrossRef]
- Piano, E.; Bona, F.; Isaia, M. Urbanization drivers differentially affect ground arthropod assemblages in the city of Turin (NW-Italy). Urban Ecosyst. 2020, 23, 617–629. [Google Scholar] [CrossRef]
- Fenoglio, M.S.; Rossetti, M.R.; Videla, M. Negative effects of urbanization on terrestrial arthropod communities: A meta-analysis. Glob. Ecol. Biogeogr. 2020, 29, 1412–1429. [Google Scholar] [CrossRef]
- Maher, G.M.; Johnson, G.A.; Burdine, J.D. Impervious surface and local abiotic conditions influence arthropod communities within urban greenspaces. PeerJ 2022, 10, e12818. [Google Scholar] [CrossRef]
- Bennett, A.B.; Gratton, C. Local and landscape scale variables impact parasitoid assemblages across an urbanization gradient. Landsc. Urban Plan. 2012, 104, 26–33. [Google Scholar] [CrossRef]
- Delgado de la Flor, Y.A.; Burkman, C.E.; Eldredge, T.K.; Gardiner, M.M. Patch and landscape-scale variables influence the taxonomic and functional composition of beetles in urban greenspaces. Ecosphere 2017, 8, e02007. [Google Scholar] [CrossRef]
- Peng, M.-H.; Hung, Y.-C.; Liu, K.-L.; Neoh, K.-B. Landscape configuration and habitat complexity shape arthropod assemblage in urban parks. Sci. Rep. 2020, 10, 16043. [Google Scholar] [CrossRef]
- McCary, M.A.; Minor, E.; Wise, D.H. Covariation between local and landscape factors influences the structure of ground-active arthropod communities in fragmented metropolitan woodlands. Landsc. Ecol. 2018, 33, 225–239. [Google Scholar] [CrossRef]
- Meineke, E.K.; Holmquist, A.J.; Wimp, G.M.; Frank, S.D. Changes in spider community composition are associated with urban temperature, not herbivore abundance. J. Urban Ecol. 2017, 3, juw010. [Google Scholar] [CrossRef]
- Angilletta, M.J. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: New York, NY, USA, 2009; ISBN 978-0-19-857087-5. [Google Scholar]
- Huey, R.B.; Kearney, M.R.; Krockenberger, A.; Holtum, J.A.M.; Jess, M.; Williams, S.E. Predicting organismal vulnerability to climate warming: Roles of behaviour, physiology and adaptation. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1665–1679. [Google Scholar] [CrossRef] [PubMed]
- Oke, T.R.; Mills, G.; Christen, A.; Voogt, J.A. Urban Climates; Cambridge University Press: Cambridge, UK, 2017; ISBN 978-0-521-84950-0. [Google Scholar]
- Foissard, X.; Dubreuil, V.; Quenol, H. Defining scales of the land use effect to map the urban heat island in a mid-size European city: Rennes (France). Urban Clim. 2019, 29, 100490. [Google Scholar] [CrossRef]
- Dubreuil, V.; Foissard, X.; Nabucet, J.; Thomas, A.; Quénol, H. Fréquence et intensité des îlots de chaleur à rennes: Bilan de 16 années d’observations (2004–2019). Climatologie 2020, 17, 6. [Google Scholar] [CrossRef]
- McGlynn, T.P.; Meineke, E.K.; Bahlai, C.A.; Li, E.J.; Hartop, E.A.; Adams, B.J.; Brown, B.V. Temperature accounts for the biodiversity of a hyperdiverse group of insects in urban Los Angeles. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191818. [Google Scholar] [CrossRef]
- Daly, C.; Conklin, D.R.; Unsworth, M.H. Local atmospheric decoupling in complex topography alters climate change impacts. Int. J. Climatol. 2010, 30, 1857–1864. [Google Scholar] [CrossRef]
- Pincebourde, S.; Salle, A. On the importance of getting fine-scale temperature records near any surface. Glob. Chang. Biol. 2020, 26, 6025–6027. [Google Scholar] [CrossRef] [PubMed]
- Lembrechts, J.J.; Aalto, J.; Ashcroft, M.B.; De Frenne, P.; Kopecký, M.; Lenoir, J.; Luoto, M.; Maclean, I.M.D.; Roupsard, O.; Fuentes-Lillo, E.; et al. SoilTemp: A global database of near-surface temperature. Glob. Chang. Biol. 2020, 26, 6616–6629. [Google Scholar] [CrossRef]
- Ziter, C.D.; Pedersen, E.J.; Kucharik, C.J.; Turner, M.G. Scale-dependent interactions between tree canopy cover and impervious surfaces reduce daytime urban heat during summer. Proc. Natl. Acad. Sci. USA 2019, 116, 7575–7580. [Google Scholar] [CrossRef] [PubMed]
- Buyantuyev, A.; Wu, J. Urban heat islands and landscape heterogeneity: Linking spatiotemporal variations in surface temperatures to land-cover and socioeconomic patterns. Landsc. Ecol. 2010, 25, 17–33. [Google Scholar] [CrossRef]
- Li, J.; Song, C.; Cao, L.; Zhu, F.; Meng, X.; Wu, J. Impacts of landscape structure on surface urban heat islands: A case study of Shanghai, China. Remote Sens. Environ. 2011, 115, 3249–3263. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Zhu, F.; Song, C.; Wu, J. Spatiotemporal pattern of urbanization in Shanghai, China between 1989 and 2005. Landsc. Ecol. 2013, 28, 1545–1565. [Google Scholar] [CrossRef]
- Pincebourde, S.; Woods, H.A. There is plenty of room at the bottom: Microclimates drive insect vulnerability to climate change. Curr. Opin. Insect Sci. 2020, 41, 63–70. [Google Scholar] [CrossRef]
- Woods, H.A.; Dillon, M.E.; Pincebourde, S. The roles of microclimatic diversity and of behavior in mediating the responses of ectotherms to climate change. J. Therm. Biol. 2015, 54, 86–97. [Google Scholar] [CrossRef]
- Merckx, T.; Van Dyck, H. Urbanization-driven homogenization is more pronounced and happens at wider spatial scales in nocturnal and mobile flying insects. Glob. Ecol. Biogeogr. 2019, 28, 1440–1455. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; Chapin, F.S.; Tecco, P.A.; Gurvich, D.E.; Grigulis, K. Functional Diversity—At the Crossroads between Ecosystem Functioning and Environmental Filters. In Terrestrial Ecosystems in a Changing World; Canadell, J.G., Pataki, D.E., Pitelka, L.F., Eds.; Global Change—The IGBP Series; Springer: Berlin/Heidelberg, Germany, 2007; pp. 81–91. ISBN 978-3-540-32730-1. [Google Scholar]
- Moretti, M.; Dias, A.T.C.; de Bello, F.; Altermatt, F.; Chown, S.L.; Azcárate, F.M.; Bell, J.R.; Fournier, B.; Hedde, M.; Hortal, J.; et al. Handbook of protocols for standardized measurement of terrestrial invertebrate functional traits. Funct. Ecol. 2017, 31, 558–567. [Google Scholar] [CrossRef]
- Kingsolver, J.; Huey, R. Size, temperature, and fitness: Three rules. Evol. Ecol. Res. 2008, 10, 251–268. [Google Scholar]
- Koivula, M.; Punttila, P.; Haila, Y.; Niemelä, J. Leaf Litter and the Small-Scale Distribution of Carabid Beetles (Coleoptera, Carabidae) in the Boreal Forest. Ecography 1999, 22, 424–435. [Google Scholar] [CrossRef]
- Small, E.C.; Sadler, J.P.; Telfer, M.G. Carabid beetle assemblages on urban derelict sites in Birmingham, UK. J. Insect Conserv. 2002, 6, 233–246. [Google Scholar] [CrossRef]
- Tyler, G. Differences in abundance, species richness, and body size of ground beetles (Coleoptera: Carabidae) between beech (Fagus sylvatica L.) forests on Podzol and Cambisol. For. Ecol. Manag. 2008, 256, 2154–2159. [Google Scholar] [CrossRef]
- Philpott, S.M.; Albuquerque, S.; Bichier, P.; Cohen, H.; Egerer, M.H.; Kirk, C.; Will, K.W. Local and Landscape Drivers of Carabid Activity, Species Richness, and Traits in Urban Gardens in Coastal California. Insects 2019, 10, 112. [Google Scholar] [CrossRef]
- Delgado de la Flor, Y.A.; Perry, K.I.; Turo, K.J.; Parker, D.M.; Thompson, J.L.; Gardiner, M.M. Local and landscape-scale environmental filters drive the functional diversity and taxonomic composition of spiders across urban greenspaces. J. Appl. Ecol. 2020, 57, 1570–1580. [Google Scholar] [CrossRef]
- Piano, E.; Giuliano, D.; Isaia, M. Islands in cities: Urbanization and fragmentation drive taxonomic and functional variation in ground arthropods. Basic Appl. Ecol. 2020, 43, 86–98. [Google Scholar] [CrossRef]
- Merckx, T.; Kaiser, A.; Van Dyck, H. Increased body size along urbanization gradients at both community and intraspecific level in macro-moths. Glob. Chang. Biol. 2018, 24, 3837–3848. [Google Scholar] [CrossRef]
- Merckx, T.; Souffreau, C.; Kaiser, A.; Baardsen, L.F.; Backeljau, T.; Bonte, D.; Brans, K.I.; Cours, M.; Dahirel, M.; Debortoli, N.; et al. Body-size shifts in aquatic and terrestrial urban communities. Nature 2018, 558, 113–116. [Google Scholar] [CrossRef]
- Atkinson, D. Temperature and organism size: A biological law for ectotherms? Adv. Ecol. Res. 1994, 25, 1–58. [Google Scholar]
- Angilletta, M.J.; Wilson, R.S.; Niehaus, A.C.; Sears, M.W.; Navas, C.A.; Ribeiro, P.L. Urban physiology: City ants possess high heat tolerance. PLoS ONE 2007, 2, e258. [Google Scholar] [CrossRef] [PubMed]
- Brans, K.I.; Jansen, M.; Vanoverbeke, J.; Tüzün, N.; Stoks, R.; De Meester, L. The heat is on: Genetic adaptation to urbanization mediated by thermal tolerance and body size. Glob. Chang. Biol. 2017, 23, 5218–5227. [Google Scholar] [CrossRef]
- Diamond, S.E.; Chick, L.D.; Perez, A.; Strickler, S.A.; Zhao, C. Evolution of plasticity in the city: Urban acorn ants can better tolerate more rapid increases in environmental temperature. Conserv. Physiol. 2018, 6, coy030. [Google Scholar] [CrossRef]
- Diamond, S.E.; Chick, L.D.; Perez, A.; Strickler, S.A.; Martin, R.A. Evolution of thermal tolerance and its fitness consequences: Parallel and non-parallel responses to urban heat islands across three cities. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180036. [Google Scholar] [CrossRef]
- Sato, A.; Takahashi, Y. Responses in thermal tolerance and daily activity rhythm to urban stress in Drosophila suzukii. Ecol. Evol. 2022, 12, e9616. [Google Scholar] [CrossRef]
- Cabon, V.; Pincebourde, S.; Colinet, H.; Dubreuil, V.; Georges, R.; Launoy, M.; Pétillon, J.; Quénol, H.; Bergerot, B. Preferred temperature in the warmth of cities: Body size, sex and development stage matter more than urban climate in a ground-dwelling spider. J. Therm. Biol. 2023, 117, 103706. [Google Scholar] [CrossRef] [PubMed]
- Menke, S.B.; Guenard, B.; Sexton, J.O.; Weiser, M.D.; Dunn, R.R.; Silverman, J. Urban areas may serve as habitat and corridors for dry-adapted, heat tolerant species; an example from ants. Urban Ecosyst. 2011, 14, 135–163. [Google Scholar] [CrossRef]
- Varet, M.; Pétillon, J.; Burel, F. Comparative responses of spider and carabid beetle assemblages along an urban-rural boundary gradient. J. Arachnol. 2011, 39, 236–243. [Google Scholar] [CrossRef]
- Vergnes, A.; Pellissier, V.; Lemperiere, G.; Rollard, C.; Clergeau, P. Urban densification causes the decline of ground-dwelling arthropods. Biodivers. Conserv. 2014, 23, 1859–1877. [Google Scholar] [CrossRef]
- Michalko, R.; Pekár, S.; Entling, M.H. An updated perspective on spiders as generalist predators in biological control. Oecologia 2019, 189, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Foelix, R. Biology of Spiders; Oxford University Press: New York, NY, USA, 2011; ISBN 978-0-19-973482-5. [Google Scholar]
- Malmos, K.G.; Lüdeking, A.H.; Vosegaard, T.; Aagaard, A.; Bechsgaard, J.; Sørensen, J.G.; Bilde, T. Behavioural and physiological responses to thermal stress in a social spider. Funct. Ecol. 2021, 35, 2728–2742. [Google Scholar] [CrossRef]
- Concepción, E.D.; Moretti, M.; Altermatt, F.; Nobis, M.P.; Obrist, M.K. Impacts of urbanisation on biodiversity: The role of species mobility, degree of specialisation and spatial scale. Oikos 2015, 124, 1571–1582. [Google Scholar] [CrossRef]
- Cheptou, P.-O.; Hargreaves, A.L.; Bonte, D.; Jacquemyn, H. Adaptation to fragmentation: Evolutionary dynamics driven by human influences. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160037. [Google Scholar] [CrossRef] [PubMed]
- Chollet, S.; Brabant, C.; Tessier, S.; Jung, V. From urban lawns to urban meadows: Reduction of mowing frequency increases plant taxonomic, functional and phylogenetic diversity. Landsc. Urban Plan. 2018, 180, 121–124. [Google Scholar] [CrossRef]
- Proske, A.; Lokatis, S.; Rolff, J. Impact of mowing frequency on arthropod abundance and diversity in urban habitats: A meta-analysis. Urban For. Urban Green. 2022, 76, 127714. [Google Scholar] [CrossRef]
- Ward, D.F.; New, T.R.; Yen, A.L. Effects of Pitfall Trap Spacing on the Abundance, Richness and Composition of Invertebrate Catches. J. Insect Conserv. 2001, 5, 47–53. [Google Scholar] [CrossRef]
- Roberts, M.J. The Spiders of Great Britain and Ireland, Volume 1: Atypidae to Theridiosomatidae; Harley Books: Colchester, UK, 1985; ISBN 978-0-946589-18-0. [Google Scholar]
- Roberts, M.J. The Spiders of Great Britain and Ireland, Volume 2: Linyphiidae and Check List; Harley Books: Colchester, UK, 1987; ISBN 978-90-04-61178-8. [Google Scholar]
- Macías-Hernández, N.; Ramos, C.; Domènech, M.; Febles, S.; Santos, I.; Arnedo, M.; Borges, P.; Emerson, B.; Cardoso, P. A database of functional traits for spiders from native forests of the Iberian Peninsula and Macaronesia. Biodivers. Data J. 2020, 8, e49159. [Google Scholar] [CrossRef]
- Nentwig, W.; Blick, T.; Bosmans, R.; Gloor, D.; Hänggi, A.; Christian, K. Spiders of Europe-Version 09.2023. Available online: http://www.araneae.unibe.ch/ (accessed on 1 September 2023).
- Blandenier, G.; Fürst, P. Ballooning spiders caught by a suction trap in an agricultural landscape in Switzerland. In Proceedings of the 17th European Colloquium of Arachnology, Edinburgh, UK, 14–18 July 1997; British Arachnological Society: Buckinghamshore, UK, 1998; Volume 1997, pp. 178–186. [Google Scholar]
- Bell, J.; Bohan, D.; Shaw, E.; Weyman, G. Ballooning dispersal using silk: World fauna, phylogenies, genetics and models. Bull. Entomol. Res. 2005, 95, 69–114. [Google Scholar] [CrossRef]
- Magura, T.; Lövei, G.L. Consequences of Urban Living: Urbanization and Ground Beetles. Curr. Landsc. Ecol. Rep. 2021, 6, 9–21. [Google Scholar] [CrossRef]
- Bonte, D.; Dahirel, M. Dispersal: A central and independent trait in life history. Oikos 2017, 126, 472–479. [Google Scholar] [CrossRef]
- Bowler, D.E.; Haase, P.; Kröncke, I.; Tackenberg, O.; Bauer, H.G.; Brendel, C.; Brooker, R.W.; Gerisch, M.; Henle, K.; Hickler, T.; et al. A cross-taxon analysis of the impact of climate change on abundance trends in central Europe. Biol. Conserv. 2015, 187, 41–50. [Google Scholar] [CrossRef]
- Cornes, R.C.; van der Schrier, G.; van den Besselaar, E.J.M.; Jones, P.D. An Ensemble Version of the E-OBS Temperature and Precipitation Data Sets. J. Geophys. Res. Atmos. 2018, 123, 9391–9409. [Google Scholar] [CrossRef]
- Moll, R.J.; Cepek, J.D.; Lorch, P.D.; Dennis, P.M.; Tans, E.; Robison, T.; Millspaugh, J.J.; Montgomery, R.A. What does urbanization actually mean? A framework for urban metrics in wildlife research. J. Appl. Ecol. 2019, 56, 1289–1300. [Google Scholar] [CrossRef]
- Chatelain, M.; Rüdisser, J.; Traugott, M. Urban-driven decrease in arthropod richness and diversity associated with group-specific changes in arthropod abundance. Front. Ecol. Evol. 2023, 11, 980387. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K.; Studer, M.; Roudier, P.; Gonzalez, J. Package ‘Cluster’. 2013. Available online: https://cran.r-project.org/web/packages/cluster/ (accessed on 1 September 2023).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 December 2020).
- Herve, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics 2023. Available online: https://cran.r-project.org/web/packages/RVAideMemoire/ (accessed on 1 September 2023).
- Roberts, D.W. labdsv: Ordination and Multivariate Analysis for Ecology 2016. Available online: https://cran.r-project.org/web/packages/labdsv/ (accessed on 1 September 2023).
- Zuur, A.; Leno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Zhang, D.B. A Coefficient of Determination for Generalized Linear Models. Am. Stat. 2017, 71, 310–316. [Google Scholar] [CrossRef]
- Hollander, J.D.; Lof, H. Differential use of the habitat by Pardosa pullata (Clerck) and Pardosa prativaga (L. Koch) in a mixed population (Araneae, Lycosidae). Tijdschr. Voor Entomol. 1972, 115, 205–215. [Google Scholar]
- de Keer, R.; Maelfait, J.-P. Observations on the life cycle of Erigone atra (Araneae, Erigoninae) in a heavily grazed pasture. Pedobiologia 1988, 32, 201–212. [Google Scholar] [CrossRef]
- Downie, I.S.; Ribera, I.; McCracken, D.I.; Wilson, W.L.; Foster, G.N.; Waterhouse, A.; Abernethy, V.J.; Murphy, K.J. Modelling populations of Erigone atra and E. dentipalpis (Araneae: Linyphiidae) across an agricultural gradient in Scotland. Agric. Ecosyst. Environ. 2000, 80, 15–28. [Google Scholar] [CrossRef]
- Sührig, A. Cryptachaea blattea, eine weitere nach Deutschland eingeschleppte Spinnenart (Araneae: Theridiidae). Arachnol. Mitteilungen 2010, 39, 1–4. [Google Scholar] [CrossRef]
- Marriott, D. Cryptachaea blattea (Urquhart, 1886) a theridiid new to Great Britain. Newsl. Br. Arachnol. Soc. 2012, 123, 9–10. [Google Scholar]
- Hänggi, A.; Straub, S. Storage buildings and greenhouses as stepping stones for non-native potentially invasive spiders (Araneae)—A baseline study in Basel, Switzerland. Arachnol. Mitteilungen 2016, 51, 1–8. [Google Scholar] [CrossRef]
- Le Divelec, R.; Courtial, C.; Rollard, C. Cryptachaea blattea (Urquart, 1886) et Pseudomaro aenigmaticus (Denis, 1966): Deux araignées nouvelles pour la France (Araneae: Theridiidae, Linyphiidae). Rev. Arachnol. 2018, 2, 2–6. [Google Scholar]
- Bink, J. Cryptachaea blattea (Urquhart, 1886) (Araneae, Theridiidae) voor het eerst waargenomen in Nederland. Nieuwsbr. SPINED 2014, 34, 11. [Google Scholar]
- Nolan, M. Notes on ‘irish’ spiders (Arachnida): Atypus affinis EICHWALD (Atypidae) and Entelecara errata O.P.-CAMBRIDGE (Linyphiidae) removed from the Irish list; Cryptachaea blattea (Urquhart)(Theridiidae) new to Ireland. Bull. Ir. Biogeogr. Soc. 2020, 44, 189–204. [Google Scholar]
- Hacala, A.; Le Roy, M.; Sawtschuk, J.; Pétillon, J. Comparative responses of spiders and plants to maritime heathland restoration. Biodivers. Conserv. 2020, 29, 229–249. [Google Scholar] [CrossRef]
- Magura, T.; Tóthmérész, B.; Molnár, T. Changes in carabid beetle assemblages along an urbanisation gradient in the city of Debrecen, Hungary. Landsc. Ecol. 2004, 19, 747–759. [Google Scholar] [CrossRef]
- Gaublomme, E.; Dhuyvetter, H.; Verdyck, P.; Desender, K. Effects of urbanisation on carabid beetles in old beech forests. DIAS Rep. 2005, 114, 111–123. [Google Scholar]
- Sadler, J.; Small, E.; Fiszpan, H.; Telfer, M.; Niemelä, J. Investigating environmental variation and landscape characteristics of an urban–rural gradient using woodland carabid assemblages. J. Biogeogr. 2006, 33, 1126–1138. [Google Scholar] [CrossRef]
- Alberti, M.; Correa, C.; Marzluff, J.M.; Hendry, A.P.; Palkovacs, E.P.; Gotanda, K.M.; Hunt, V.M.; Apgar, T.M.; Zhou, Y. Global urban signatures of phenotypic change in animal and plant populations. Proc. Natl. Acad. Sci. USA 2017, 114, 8951–8956. [Google Scholar] [CrossRef]
- Johnson, J.C.; Urcuyo, J.; Moen, C.; Stevens, D.R., II. Urban heat island conditions experienced by the Western black widow spider (Latrodectus hesperus): Extreme heat slows development but results in behavioral accommodations. PLoS ONE 2019, 14, e0220153. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, A.L.; Youngsteadt, E.; Lopez-Uribe, M.M.; Frank, S.D. Physiological thermal limits predict differential responses of bees to urban heat-island effects. Biol. Lett. 2017, 13, 20170125. [Google Scholar] [CrossRef] [PubMed]
- Blanckenhorn, W.U. The evolution of body size: What keeps organisms small? Q. Rev. Biol. 2000, 75, 385–407. [Google Scholar] [CrossRef]
- Nielsen, M.E.; Papaj, D.R. Effects of developmental change in body size on ectotherm body temperature and behavioral thermoregulation: Caterpillars in a heat-stressed environment. Oecologia 2015, 177, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Klockmann, M.; Günter, F.; Fischer, K. Heat resistance throughout ontogeny: Body size constrains thermal tolerance. Glob. Chang. Biol. 2016, 23, 686–696. [Google Scholar] [CrossRef]
- Niemelä, J.; Kotze, D.J. Carabid beetle assemblages along urban to rural gradients: A review. Landsc. Urban Plan. 2009, 92, 65–71. [Google Scholar] [CrossRef]
- Scheffers, B.R.; De Meester, L.; Bridge, T.C.L.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.M.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The broad footprint of climate change from genes to biomes to people. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef]
- Blanckenhorn, W.U.; Demont, M. Bergmann and Converse Bergmann Latitudinal Clines in Arthropods: Two Ends of a Continuum? Integr. Comp. Biol. 2004, 44, 413–424. [Google Scholar] [CrossRef]
- Meineke, E.K.; Dunn, R.R.; Sexton, J.O.; Frank, S.D. Urban Warming Drives Insect Pest Abundance on Street Trees. PLoS ONE 2013, 8, e59687. [Google Scholar] [CrossRef]
- Hamblin, A.L.; Youngsteadt, E.; Frank, S.D. Wild bee abundance declines with urban warming, regardless of floral density. Urban Ecosyst. 2018, 21, 419–428. [Google Scholar] [CrossRef]
- Francoeur, X.W.; Dagenais, D.; Paquette, A.; Dupras, J.; Messier, C. Complexifying the urban lawn improves heat mitigation and arthropod biodiversity. Urban For. Urban Green. 2021, 60, 127007. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Mean + SD | Unit | Details |
|---|---|---|---|
| Landscape scale | |||
| Urbanization | 36.80 ± 21.94 | % | Percentage of impervious surface within a 100 m radius buffer around sampling sites. Land cover data were obtained at a resolution of 5 m from the French portal for geographical data (dataset OCSGE 2017; https://geo.data.gouv.fr, accessed on 1 December 2021). |
| Urban heat island | 1.71 ± 0.56 | °C | Average difference between the minimum daily temperature at a reference cold station located outside the urban area and the minimum daily temperature at each other sensor, over the sampling period. Atmospheric temperature data used to calculate the UHI were retrieved from the long-term Rennes Climate Urban Network (see https://run.letg.cnrs.fr for details). Based on 30 DAVIS Vantage-Pro-2 automated weather stations (Davis Instruments Corporation, Hayward, CA 94545, USA) and 93 RisingHF RHF1SOO1 connected temperature sensors (LoRaWAN Solutions RisingHF, Shenzhen 518057, China), UHI was interpolated by inverse distance weighting to spatialize the estimated UHI values over the city extent at a resolution of 100 m [22]. From this map, we finally extracted the mean pixel values within circular buffers of 100 m around the sampling sites. |
| Local scale | |||
| Vegetation height | 19.73 ± 11.54 | cm | Mean vegetation height measured within the triangle materialized by the three pitfall traps during each of the seven sampling sessions. All seven measurements were averaged to obtain one final value per site. |
| Vegetation cover | 90.50 ± 9.38 | % | Percentage of herbaceous vegetation cover estimated in 10% increments within the triangle materialized by the three pitfall traps during each of the seven sampling sessions. All seven measurements were averaged to obtain one final value per site. A low vegetation cover indicates the presence of litter or bare soil. |
| Near-ground temperature | 2.58 ± 1.39 | °C | Average difference between the minimum daily temperature at a reference cold station and the minimum daily temperature at each other sensor, over the sampling period. Temperatures were recorded every 15 min during each trapping session using a dedicated network of temperature loggers composed of 21 Lascar EL-USB-2+ (Lascar Electronics Ltd., Whiteparish, Wiltshire SP5 2SJ, UK) and 15 Tinytag Talk 2 TK-4023 (Gemini Data Loggers Ltd., Chichester, West Sussex PO19 8UJ, UK). The sensors were placed 5 cm above the ground surface at each sampling site, in close proximity to the north trap. |
| High Vegetated Rural | N | % | Short Vegetated Rural | N | % | Short Vegetated Urban | N | % |
|---|---|---|---|---|---|---|---|---|
| Total | 7535 | 100 | Total | 6529 | 100 | Total | 5983 | 100 |
| Pachygnatha degeeri | 2159 | 28.6 | Pachygnatha degeeri | 1650 | 25.3 | Pachygnatha degeeri | 1128 | 18.9 |
| Pardosa cf. tenuipes | 691 | 9.2 | Pardosa cf. tenuipes | 1464 | 22.4 | Erigone dentipalpis | 1080 | 18.1 |
| Oedothorax fuscus | 657 | 8.7 | Tenuiphantes tenuis | 319 | 4.9 | Tenuiphantes tenuis | 522 | 8.7 |
| Oedothorax retusus | 551 | 7.3 | Alopecosa pulverulenta | 227 | 3.5 | Pardosa cf. tenuipes | 442 | 7.4 |
| Tenuiphantes tenuis | 368 | 4.9 | Oedothorax fuscus | 195 | 3.0 | Erigone atra | 393 | 6.6 |
| Pardosa pullata | 357 | 4.7 | Pardosa pullata | 195 | 3.0 | Bathyphantes gracilis | 327 | 5.5 |
| Agyneta affinis | 240 | 3.2 | Diplostyla concolor | 189 | 2.9 | Oedothorax fuscus | 222 | 3.7 |
| Bathyphantes gracilis | 214 | 2.8 | Hahnia nava | 188 | 2.9 | Hahnia nava | 213 | 3.6 |
| Aulonia albimana | 207 | 2.7 | Aulonia albimana | 182 | 2.8 | Oedothorax retusus | 169 | 2.8 |
| Diplostyla concolor | 194 | 2.6 | Bathyphantes gracilis | 169 | 2.6 | Tiso vagans | 161 | 2.7 |
| Pardosa prativaga | 177 | 2.3 | Oedothorax retusus | 153 | 2.3 | Diplostyla concolor | 115 | 1.9 |
| Hahnia nava | 174 | 2.3 | Tiso vagans | 121 | 1.9 | Ozyptila simplex | 115 | 1.9 |
| Alopecosa pulverulenta | 151 | 2.0 | Agyneta affinis | 115 | 1.8 | Agyneta mollis | 104 | 1.7 |
| Agyneta mollis | 144 | 1.9 | Erigone dentipalpis | 107 | 1.6 | Agyneta affinis | 92 | 1.5 |
| Panamomops sulcifrons | 138 | 1.8 | Erigone atra | 104 | 1.6 | Pardosa pullata | 88 | 1.5 |
| Erigone dentipalpis | 112 | 1.5 | Pardosa prativaga | 103 | 1.6 | Aulonia albimana | 82 | 1.4 |
| Trochosa terricola | 91 | 1.2 | Trochosa terricola | 101 | 1.5 | Cryptachaea blattea | 82 | 1.4 |
| Tiso vagans | 86 | 1.1 | Phrurolithus festivus | 78 | 1.2 | Panamomops sulcifrons | 80 | 1.3 |
| Erigone atra | 61 | 0.8 | Agyneta mollis | 76 | 1.2 | Phrurolithus festivus | 72 | 1.2 |
| Species | N | % | Environmental Variables | Adj-R2 | ||||
|---|---|---|---|---|---|---|---|---|
| Urbanization | UHI | Near-Ground Temperature | Vegetation Height | Vegetation Cover | ||||
| Pachygnatha degeeri | 4937 | 24.62 | ↓ (0.020) | 0.27 | ||||
| Pardosa cf. tenuipes | 2597 | 12.95 | 0.00 | |||||
| Erigone dentipalpis | 1299 | 6.48 | ↑ (0.002) | ↓ (0.023) | ↓ (0.033) | 0.51 | ||
| Tenuiphantes tenuis | 1209 | 6.03 | 0.02 | |||||
| Oedothorax fuscus | 1074 | 5.36 | ↑ (<0.001) | ↓ (0.002) | 0.47 | |||
| Oedothorax retusus | 873 | 4.35 | ↑ (0.003) | ↓ (0.020) | 0.33 | |||
| Bathyphantes gracilis | 710 | 3.54 | 0.02 | |||||
| Pardosa pullata | 640 | 3.19 | ↑ (0.010) | ↓ (0.020) | ↓ (<0.001) | ↑ (0.010) | 0.55 | |
| Hahnia nava | 575 | 2.87 | 0.06 | |||||
| Erigone atra | 558 | 2.78 | ↓ (0.030) | ↑ (0.010) | 0.34 | |||
| Diplostyla concolor | 498 | 2.48 | 0.06 | |||||
| Aulonia albimana | 471 | 2.35 | ↓ (0.040) | ↑ (0.030) | 0.11 | |||
| Agyneta affinis | 447 | 2.23 | 0.06 | |||||
| Alopecosa pulverulenta | 403 | 2.01 | ↓ (0.005) | ↓ (0.030) | 0.14 | |||
| Tiso vagans | 368 | 1.84 | 0.00 | |||||
| Agyneta mollis | 324 | 1.62 | ↑ (0.010) | 0.16 | ||||
| Pardosa prativaga | 304 | 1.52 | 0.10 | |||||
| Panamomops sulcifrons | 288 | 1.44 | 0.13 | |||||
| Trochosa terricola | 213 | 1.06 | ↓ (0.020) | 0.26 | ||||
| Phrurolithus festivus | 207 | 1.03 | 0.00 | |||||
| Ozyptila simplex | 203 | 1.01 | 0.01 | |||||
| Mermessus trilobatus | 142 | 0.71 | 0.00 | |||||
| Pachygnatha clercki | 100 | 0.50 | ↓ (0.050) | 0.06 | ||||
| Pelecopsis parallela | 99 | 0.49 | 0.00 | |||||
| Palliduphantes pallidus | 94 | 0.47 | ↑ (0.050) | 0.15 | ||||
| Dicymbium nigrum | 84 | 0.42 | 0.07 | |||||
| Monocephalus fuscipes | 84 | 0.42 | 0.00 | |||||
| Cryptachaea blattea | 82 | 0.41 | ↑ (0.001) | ↓ (0.002) | 0.85 | |||
| Pardosa palustris | 76 | 0.38 | 0.58 | |||||
| Environmental Variables | Adj-R2 | ||||
|---|---|---|---|---|---|
| Urbanization | UHI | Vegetation Height | Vegetation Cover | ||
| Body size classes | |||||
| Small | ↓ (0.04) | - | - | - | 0.36 |
| Medium | - | ↓ (0.02) | - | - | 0.29 |
| Large | - | ↓ (<0.001) | ↑ (<0.001) | ↓ (0.01) | 0.60 |
| Thermal affinity classes | |||||
| Low temperatures | - | ↓ (0.03) | - | - | 0.45 |
| Intermediate temperatures | - | ↓ (0.002) | ↑ (0.003) | ↓ (<0.001) | 0.75 |
| High temperatures | - | - | - | - | 0.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabon, V.; Quénol, H.; Dubreuil, V.; Ridel, A.; Bergerot, B. Urban Heat Island and Reduced Habitat Complexity Explain Spider Community Composition by Excluding Large and Heat-Sensitive Species. Land 2024, 13, 83. https://doi.org/10.3390/land13010083
Cabon V, Quénol H, Dubreuil V, Ridel A, Bergerot B. Urban Heat Island and Reduced Habitat Complexity Explain Spider Community Composition by Excluding Large and Heat-Sensitive Species. Land. 2024; 13(1):83. https://doi.org/10.3390/land13010083
Chicago/Turabian StyleCabon, Valentin, Hervé Quénol, Vincent Dubreuil, Aurélien Ridel, and Benjamin Bergerot. 2024. "Urban Heat Island and Reduced Habitat Complexity Explain Spider Community Composition by Excluding Large and Heat-Sensitive Species" Land 13, no. 1: 83. https://doi.org/10.3390/land13010083
APA StyleCabon, V., Quénol, H., Dubreuil, V., Ridel, A., & Bergerot, B. (2024). Urban Heat Island and Reduced Habitat Complexity Explain Spider Community Composition by Excluding Large and Heat-Sensitive Species. Land, 13(1), 83. https://doi.org/10.3390/land13010083