

The Effect of Multi-Use Landscapes on Mammal Assemblages and Its Implication for Conservation

Abstract
1. Introduction
2. Materials and Methods
2.1. Land-Uses
2.2. Camera Trapping
2.3. Single-Season Multi-Region Community Occurrence Model
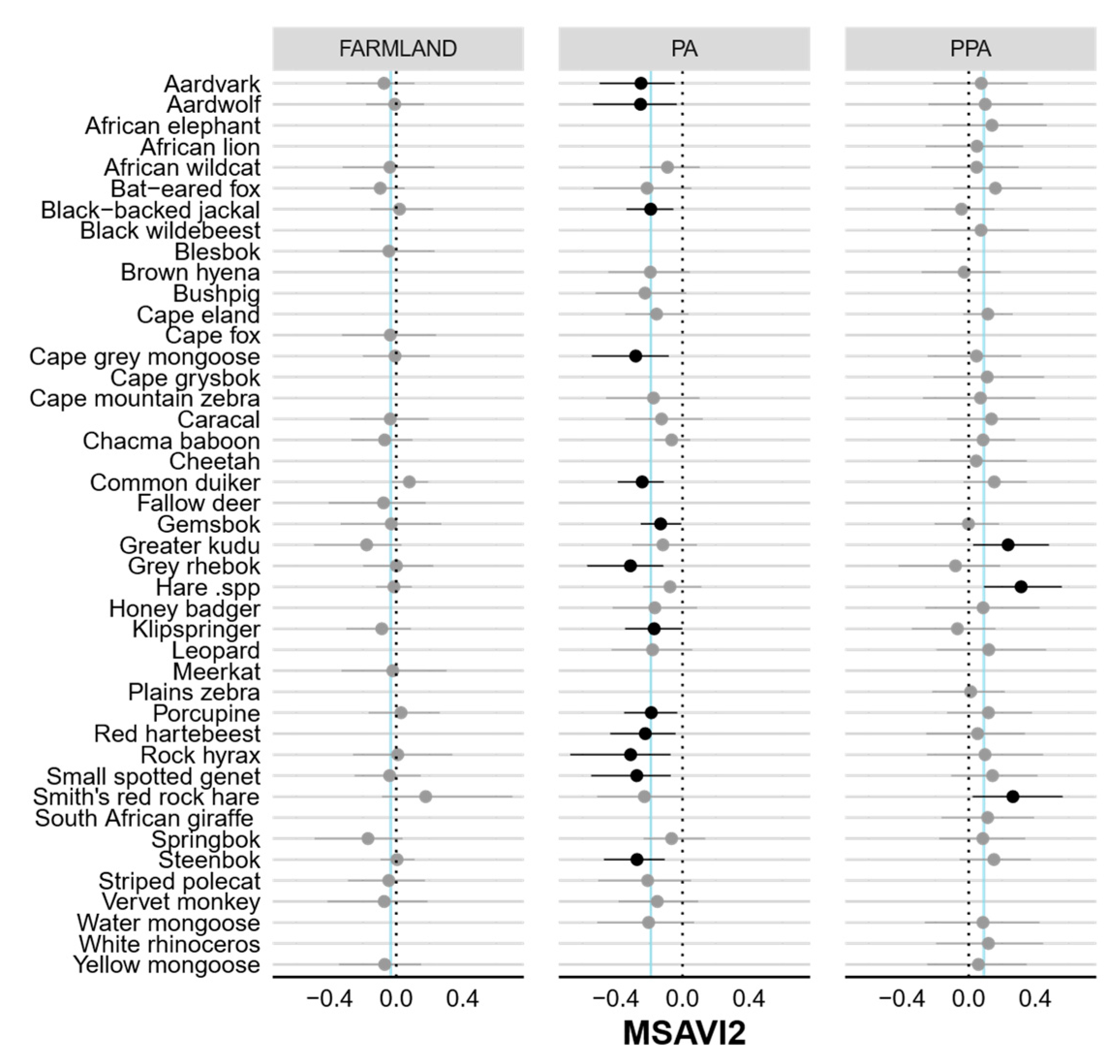
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land-se | Area (km2) | Sampling Period | Total No. Sites | Effort | Total Independent Detections | Overall Trapping Rate | Total Detected Species | Mean No. Species per Site |
|---|---|---|---|---|---|---|---|---|
| Farmland | 754 | September–March 2012/2013 | 156 | 10,842 | 3480 | 0.27 | 27 1 | 10.8 |
| PA | 802 | October–June 2013/2014 | 176 | 9538 | 2966 | 0.28 | 30 | 10.8 |
| PPA | 540 | August–November 2015 | 119 | 5951 | 2484 | 0.38 | 36 | 11.4 |
| Total | 2096 | 451 | 15,489 | 8930 |
| Family Common Name Species | Total Independent Detections | Total Camera Trap Sites | Naïve Occupancy | Detection Frequency |
|---|---|---|---|---|
| Bovidae | ||||
| Black Wildebeest Connochaetes gnou | 0 | 0 | 0.00 | 0.00 |
| Blesbok Damaliscus pygargus | 27 | 10 | 0.06 | 0.25 |
| Cape eland Taurotragus oryx | 0 | 0 | 0.00 | 0.00 |
| Cape grysbok Raphicerus melanotis | 0 | 0 | 0.00 | 0.00 |
| Common duiker Sylvicapra grimmia | 957 | 107 | 0.69 | 8.83 |
| Gemsbok Oryx gazella | 20 | 3 | 0.02 | 0.18 |
| Greater kudu Tragelaphus strepsiceros | 124 | 27 | 0.17 | 1.24 |
| Grey rhebuck Pelea capreolus | 69 | 20 | 0.13 | 0.64 |
| Klipspringer Oreotragus oreotragus | 50 | 14 | 0.09 | 0.46 |
| Red hartebeest Alcelaphus buselaphus caama | 2 | 1 | * | * |
| Springbok Antidorcas marsupialis | 86 | 14 | 0.09 | 0.79 |
| Steenbok Raphicerus campestris | 700 | 116 | 0.74 | 6.46 |
| Canidae | ||||
| Bat-eared fox Otocyon megalotis | 218 | 41 | 0.26 | 2.01 |
| Black-backed jackal Canis mesomelas | 107 | 34 | 0.22 | 0.99 |
| Cape fox Vulpes chama | 16 | 11 | 0.07 | 0.15 |
| Cercopithecidae | ||||
| Chacma baboon Papio ursinus | 95 | 36 | 0.23 | 0.88 |
| Vervet monkey Chlorocebus pygerythrus | 8 | 6 | 0.04 | 0.07 |
| Cervidae | ||||
| Fallow deer Dama dama | 18 | 4 | 0.03 | 0.16 |
| Elephantidae | 0.00 | |||
| African elephant Loxodonta africana | 0 | 0 | 0.00 | 0.00 |
| Equidae | ||||
| Cape mountain zebra Equus zebra zebra | 0 | 0 | 0.00 | 0.00 |
| Plains zebra Equus quagga | 0 | 0 | 0.00 | 0.00 |
| Felidae | ||||
| African Lion Panthera leo | 0 | 0 | 0.00 | 0.00 |
| African wildcat Felis silvestris | 10 | 10 | 0.06 | 0.09 |
| Caracal Caracal caracal | 23 | 19 | 0.12 | 0.21 |
| Cheetah Acinonyx jubatus | 0 | 0 | 0.00 | 0.00 |
| Leopard Panthera pardus | 0 | 0 | 0.00 | 0.00 |
| Giraffidae | ||||
| South African giraffe Giraffa camelopardalis giraffa | 0 | 0 | 0.00 | 0.00 |
| Herpestidae | ||||
| Cape grey mongoose Herpestes pulverulentus | 37 | 27 | 0.17 | 0.34 |
| Meerkat Suricata suricatta | 3 | 3 | 0.02 | 0.02 |
| Water mongoose Atilax paludinosus | 0 | 0 | 0.00 | 0.00 |
| Yellow mongoose Cynictis penicillata | 16 | 12 | 0.08 | 0.15 |
| Hyaenidae | ||||
| Aardwolf Proteles cristata | 100 | 49 | 0.31 | 0.92 |
| Brown hyena Parahyaena brunnea | 0 | 0 | 0.00 | 0.00 |
| Hystricidae | ||||
| Porcupine Hystrix africaeaustralis | 65 | 36 | 0.23 | 0.60 |
| Leporiade | ||||
| Hare spp. Lepus saxatilis and Lepus capensis | 556 | 114 | 0.73 | 5.12 |
| Smith’s red rock rabbit Pronolagus rupestris | 30 | 2 | 0.01 | 0.28 |
| Mustelidae | ||||
| Honey badger Mellivora capensis | 0 | 0 | 0.00 | 0.00 |
| Striped polecat Ictonyx striatus | 36 | 25 | 0.16 | 0.33 |
| Orycteropodidae | ||||
| Aardvark Orycteropus afer | 43 | 24 | 0.15 | 0.40 |
| Procaviidae | ||||
| Rock hyrax Procavia capensis | 14 | 5 | 0.03 | 0.13 |
| Rhinocerotidae | ||||
| White rhinoceros Ceratotherium simum | 0 | 0 | 0.00 | 0.00 |
| Suidae | ||||
| Bushpig Potamochoerus larvatus | 0 | 0 | 0.00 | 0.00 |
| Viverridae | ||||
| Small spotted genet Genetta genetta | 42 | 24 | 0.15 | 0.39 |
| Family Common Name Species | Total Independent Detections | Total Camera Trap Sites | Naïve Occupancy | Detection Frequency |
|---|---|---|---|---|
| Bovidae | ||||
| Black Wildebeest Connochaetes gnou | 0 | 0 | 0.00 | 0.00 |
| Blesbok Damaliscus pygargus | 0 | 0 | 0.00 | 0.00 |
| Cape eland Taurotragus oryx | 64 | 25 | 0.14 | 0.67 |
| Cape grysbok Raphicerus melanotis | 0 | 0 | 0.00 | 0.00 |
| Common duiker Sylvicapra grimmia | 320 | 63 | 0.36 | 3.36 |
| Gemsbok Oryx gazella | 441 | 68 | 0.39 | 4.62 |
| Greater kudu Tragelaphus strepsiceros | 80 | 18 | 0.10 | 0.84 |
| Grey rhebuck Pelea capreolus | 73 | 19 | 0.11 | 0.77 |
| Klipspringer Oreotragus oreotragus | 193 | 49 | 0.28 | 2.02 |
| Red hartebeest Alcelaphus buselaphus caama | 115 | 19 | 0.11 | 1.21 |
| Springbok Antidorcas marsupialis | 91 | 22 | 0.13 | 0.95 |
| Steenbok Raphicerus campestris | 103 | 37 | 0.21 | 1.08 |
| Canidae | ||||
| Bat-eared fox Otocyon megalotis | 7 | 1 | 0.01 | 0.07 |
| Black-backed jackal Canis mesomelas | 237 | 51 | 0.29 | 2.48 |
| Cape fox Vulpes chama | 0 | 0 | 0.00 | 0.00 |
| Cercopithecidae | ||||
| Chacma baboon Papio ursinus | 536 | 112 | 0.64 | 5.62 |
| Vervet monkey Chlorocebus pygerythrus | 16 | 5 | 0.03 | 0.17 |
| Cervidae | ||||
| Fallow deer Dama dama | 0 | 0 | 0.00 | 0.00 |
| Elephantidae | ||||
| African elephant Loxodonta africana | 0 | 0 | 0.00 | 0.00 |
| Equidae | ||||
| Cape mountain zebra Equus zebra zebra | 20 | 10 | 0.06 | 0.21 |
| Plains zebra Equus quagga | 0 | 0 | 0.00 | 0.00 |
| Felidae | ||||
| African Lion Panthera leo | 0 | 0 | 0.00 | 0.00 |
| African wildcat Felis silvestris | 97 | 44 | 0.25 | 1.02 |
| Caracal Caracal caracal | 9 | 5 | 0.03 | 0.09 |
| Cheetah Acinonyx jubatus | 0 | 0 | 0.00 | 0.00 |
| Leopard Panthera pardus | 23 | 15 | 0.09 | 0.24 |
| Giraffidae | ||||
| South African giraffe Giraffa camelopardalis giraffa | 0 | 0 | 0.00 | 0.00 |
| Herpestidae | ||||
| Cape grey mongoose Herpestes pulverulentus | 49 | 21 | 0.12 | 0.51 |
| Meerkat Suricata suricatta | 0 | 0 | 0.00 | 0.00 |
| Water mongoose Atilax paludinosus | 1 | 1 | 0.01 | 0.01 |
| Yellow mongoose Cynictis penicillata | 0 | 0 | 0.00 | 0.00 |
| Hyaenidae | ||||
| Aardwolf Proteles cristata | 23 | 16 | 0.09 | 0.24 |
| Brown hyena Parahyaena brunnea | 15 | 9 | 0.05 | 0.16 |
| Hystricidae | ||||
| Porcupine Hystrix africaeaustralis | 112 | 36 | 0.20 | 1.17 |
| Leporidae | ||||
| Hare spp. Lepus saxatilis and Lepus capensis | 137 | 25 | 0.14 | 1.44 |
| Smith’s red rock rabbit Pronolagus rupestris | 99 | 28 | 0.16 | 1.04 |
| Mustelidae | ||||
| Honey badger Mellivora capensis | 17 | 9 | 0.05 | 0.18 |
| Striped polecat Ictonyx striatus | 1 | 1 | 0.01 | 0.01 |
| Orycteropodidae | ||||
| Aardvark Orycteropus afer | 28 | 23 | 0.13 | 0.29 |
| Procaviidae | ||||
| Rock hyrax Procavia capensis | 37 | 4 | 0.02 | 0.39 |
| Rhinocerotidae | ||||
| White rhinoceros Ceratotherium simum | 0 | 0 | 0.00 | 0.00 |
| Suidae | ||||
| Bushpig Potamochoerus larvatus | 4 | 4 | 0.02 | 0.04 |
| Viverridae | ||||
| Small spotted genet Genetta genetta | 18 | 13 | 0.07 | 0.19 |
| Family Common Name Species | Total Independent Detections | Total Camera Trap Sites | Naïve Occupancy | Detection Frequency |
|---|---|---|---|---|
| Bovidae | ||||
| Black Wildebeest Connochaetes gnou | 24 | 4 | 0.03 | 0.51 |
| Blesbok Damaliscus pygargus | 0 | 0 | 0.00 | 0.00 |
| Cape eland Taurotragus oryx | 460 | 69 | 0.58 | 7.10 |
| Cape grysbok Raphicerus melanotis | 2 | 1 | 0.01 | 0.03 |
| Common duiker Sylvicapra grimmia | 159 | 44 | 0.37 | 2.45 |
| Gemsbok Oryx gazella | 231 | 55 | 0.46 | 3.56 |
| Greater kudu Tragelaphus strepsiceros | 121 | 30 | 0.25 | 1.87 |
| Grey rhebuck Pelea capreolus | 33 | 9 | 0.08 | 0.51 |
| Klipspringer Oreotragus oreotragus | 81 | 19 | 0.16 | 1.25 |
| Red hartebeest Alcelaphus buselaphus caama | 25 | 6 | 0.05 | 0.39 |
| Springbok Antidorcas marsupialis | 68 | 11 | 0.09 | 1.05 |
| Steenbok Raphicerus campestris | 89 | 31 | 0.26 | 1.37 |
| Canidae | ||||
| Bat-eared fox Otocyon megalotis | 7 | 4 | 0.03 | 0.11 |
| Black-backed jackal Canis mesomelas | 134 | 51 | 0.43 | 2.07 |
| Cape fox Vulpes chama | 0 | 0 | 0.00 | 0.00 |
| Cercopithecidae | ||||
| Chacma baboon Papio ursinus | 228 | 54 | 1.92 | 3.52 |
| Vervet monkey Chlorocebus pygerythrus | 0 | 0 | 0.00 | 0.00 |
| Cervidae | ||||
| Fallow deer Dama dama | 0 | 0 | 0.00 | 0.00 |
| Elephantidae | ||||
| African elephant Loxodonta africana | 24 | 6 | 0.05 | 0.37 |
| Equidae | ||||
| Cape mountain zebra Equus zebra zebra | 1 | 1 | 0.01 | 0.02 |
| Plains zebra Equus quagga | 165 | 32 | 0.27 | 2.55 |
| Felidae | ||||
| African Lion Panthera leo | 42 | 10 | 0.08 | 0.65 |
| African wildcat Felis silvestris | 32 | 20 | 0.17 | 0.49 |
| Family Common name Species | Total independent detections | Total camera traps | Naïve occupancy | Detection frequency (RAI) |
| Caracal Caracal caracal | 21 | 12 | 0.10 | 0.32 |
| Cheetah Acinonyx jubatus | 10 | 5 | 0.04 | 0.15 |
| Leopard Panthera pardus | 6 | 4 | 0.03 | 0.09 |
| Giraffidae | ||||
| South African giraffe Giraffa camelopardalis giraffa | 42 | 12 | 0.10 | 0.65 |
| Herpestidae | ||||
| Cape grey mongoose Herpestes pulverulentus | 14 | 9 | 0.08 | 0.22 |
| Meerkat Suricata suricatta | 0 | 0 | 0.00 | 0.00 |
| Water mongoose Atilax paludinosus | 2 | 2 | 0.02 | 0.03 |
| Yellow mongoose Cynictis penicillata | 3 | 3 | 0.03 | 0.05 |
| Hyaenidae | ||||
| Aardwolf Proteles cristata | 2 | 1 | 0.01 | 0.03 |
| Brown hyena Parahyaena brunnea | 49 | 27 | 0.23 | 0.76 |
| Hystricidae | ||||
| Porcupine Hystrix africaeaustralis | 55 | 24 | 0.20 | 0.85 |
| Leporidae | ||||
| Hare spp. Lepus saxatilis and Lepus capensis | 244 | 28 | 0.24 | 3.77 |
| Smith’s red rock rabbit Pronolagus rupestris | 41 | 14 | 0.12 | 0.63 |
| Mustelidae | ||||
| Honey badger Mellivora capensis | 3 | 3 | 0.03 | 0.05 |
| Striped polecat Ictonyx striatus | 0 | 0 | 0.00 | 0.00 |
| Orycteropodidae | ||||
| Aardvark Orycteropus afer | 19 | 11 | 0.09 | 0.29 |
| Procaviidae | ||||
| Rock hyrax Procavia capensis | 5 | 2 | 0.02 | 0.08 |
| Rhinocerotidae | ||||
| White rhinoceros Ceratotherium simum | 12 | 4 | 0.03 | 0.19 |
| Suidae | ||||
| Bushpig Potamochoerus larvatus | 0 | 0 | 0.00 | 0.00 |
| Viverridae | ||||
| Small spotted genet Genetta genetta | 30 | 18 | 0.15 | 0.46 |






References
- O’Bryan, C.J.; Garnett, S.T.; Fa, J.E.; Leiper, I.; Rehbein, J.A.; Fernández-Llamazares, Á.; Jackson, M.V.; Jonas, H.D.; Brondizio, E.S.; Burgess, N.D.; et al. The Importance of Indigenous Peoples’ Lands for the Conservation of Terrestrial Mammals. Conserv. Biol. 2021, 35, 1002–1008. [Google Scholar] [CrossRef] [PubMed]
- Zungu, M.M.; Maseko, M.S.T.; Kalle, R.; Ramesh, T.; Downs, C.T. Factors Affecting the Occupancy of Forest Mammals in an Urban-Forest Mosaic in EThekwini Municipality, Durban, South Africa. Urban For. Urban Green. 2020, 48, 126562. [Google Scholar] [CrossRef]
- Hoffmann, S.; Beierkuhnlein, C. Climate Change Exposure and Vulnerability of the Global Protected Area Estate from an International Perspective. Divers. Distrib. 2020, 26, 1496–1509. [Google Scholar] [CrossRef]
- Jenkins, C.N.; Joppa, L. Expansion of the Global Terrestrial Protected Area System. Biol. Conserv. 2009, 142, 2166–2174. [Google Scholar] [CrossRef]
- Venter, O.; Magrach, A.; Outram, N.; Klein, C.J.; Possingham, H.P.; Di Marco, M.; Watson, J.E.M. Bias in Protected-Area Location and Its Effects on Long-Term Aspirations of Biodiversity Conventions. Conserv. Biol. 2018, 32, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Hoveka, L.N.; van der Bank, M.; Davies, T.J. Evaluating the Performance of a Protected Area Network in South Africa and Its Implications for Megadiverse Countries. Biol. Conserv. 2020, 248, 108577. [Google Scholar] [CrossRef]
- Sarkar, S.; Pressey, R.L.; Faith, D.P.; Margules, C.R.; Fuller, T.; Stoms, D.M.; Moffett, A.; Wilson, K.A.; Williams, K.J.; Williams, P.H.; et al. Biodiversity Conservation Planning Tools: Present Status and Challenges for the Future. Annu. Rev. Environ. Resour. 2006, 31, 123–159. [Google Scholar] [CrossRef]
- Clements, H.S.; Kerley, G.I.H.; Cumming, G.S.; De Vos, A.; Cook, C.N. Privately Protected Areas Provide Key Opportunities for the Regional Persistence of Large- and Medium-Sized Mammals. J. Appl. Ecol. 2019, 56, 537–546. [Google Scholar] [CrossRef]
- Lindsey, P.A.; Romañach, S.S.; Davies-Mostert, H.T. The Importance of Conservancies for Enhancing the Value of Game Ranch Land for Large Mammal Conservation in Southern Africa. J. Zool. 2009, 277, 99–105. [Google Scholar] [CrossRef]
- Taylor, W.A.; Child, M.F.; Lindsey, P.A.; Nicholson, S.K.; Relton, C.; Davies-Mostert, H.T. South Africa’s Private Wildlife Ranches Protect Globally Significant Populations of Wild Ungulates. Biodivers. Conserv. 2021, 30, 4111–4135. [Google Scholar] [CrossRef]
- Spierenburg, M.; Brooks, S. Private Game Farming and Its Social Consequences in Post-Apartheid South Africa: Contestations over Wildlife, Property and Agrarian Futures. J. Contemp. African Stud. 2014, 32, 151–172. [Google Scholar] [CrossRef]
- Faure, J.P.B.; Swanepoel, L.H.; Cilliers, D.; Venter, J.A.; Hill, R.A. Estimates of Carnivore Densities in a Human-Dominated Agricultural Matrix in South Africa. Oryx 2021, 56, 774–781. [Google Scholar] [CrossRef]
- Taylor, W.A.; Lindsey, P.A.; Nicholson, S.K.; Relton, C.; Davies-Mostert, H.T. Jobs, Game Meat and Profits: The Benefits of Wildlife Ranching on Marginal Lands in South Africa. Biol. Conserv. 2020, 245, 108561. [Google Scholar] [CrossRef]
- Gordon, I.J.; Hester, A.J.; Festa-Bianchet, M. REVIEW: The Management of Wild Large Herbivores to Meet Economic, Conservation and Environmental Objectives. J. Appl. Ecol. 2004, 41, 1021–1031. [Google Scholar] [CrossRef]
- Boronyak, L.; Jacobs, B.; Wallach, A.; McManus, J.; Stone, S.; Stevenson, S.; Smuts, B.; Zaranek, H. Pathways towards Coexistence with Large Carnivores in Production Systems. Agric. Hum. Values 2022, 39, 47–64. [Google Scholar] [CrossRef]
- Carter, E.; Adams, W.M.; Hutton, J. Private Protected Areas: Management Regimes, Tenure Arrangements and Protected Area Categorization in East Africa. Oryx 2008, 42, 177–186. [Google Scholar] [CrossRef]
- Cousins, J.A.; Sadler, J.P.; Evans, J. Exploring the Role of Private Wildlife Ranching as a Conservation Tool in South Africa: Stakeholder Perspectives. Ecol. Soc. 2008, 13, 43. [Google Scholar] [CrossRef]
- Maciejewski, K.; Kerley, G.I.H.H. Understanding Tourists’ Preference for Mammal Species in Private Protected Areas: Is There a Case for Extralimital Species for Ecotourism? PLoS ONE 2014, 9, e88192. [Google Scholar] [CrossRef]
- Palfrey, R.; Oldekop, J.; Holmes, G. Conservation and Social Outcomes of Private Protected Areas. Conserv. Biol. 2021, 35, 1098–1110. [Google Scholar] [CrossRef]
- Barany, M.E.; Hammett, A.L.; Shillington, L.J.; Murphy, B.R. The Role of Private Wildlife Reserves in Nicaragua’s Emerging Ecotourism Industry. J. Sustain. Tour. 2001, 9, 95–110. [Google Scholar] [CrossRef]
- de Santo, E.M. From Paper Parks to Private Conservation: The Role of NGOs in Adapting Marine Protected Area Strategies to Climate Change. J. Int. Wildl. Law Policy 2012, 15, 25–40. [Google Scholar] [CrossRef]
- Gooden, J.; ‘t Sas-Rolfes, M. A Review of Critical Perspectives on Private Land Conservation in Academic Literature. Ambio 2020, 49, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Niemeijer, D.; Puigdefabregas, J.; White, R.; Lal, R.; Winslow, M.; Ziedler, J.; Prince, S.; Archer, E.; King, C.; Shaprio, B.; et al. Dryland Systems. In Ecosystems and Human Well-Being: Current State and Trends; Hassan, R., Scholes, R., Neville, A., Eds.; Island Press: Washington, DC, USA, 2005; pp. 623–662. [Google Scholar]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Ochoa, V.; Delgado-Baquerizo, M.; García-Gómez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; et al. Plant Species Richness and Ecosystem Multifunctionality in Global Drylands. Science 2012, 335, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Kiffner, C.; Wenner, C.; LaViolet, A.; Yeh, K.; Kioko, J. From Savannah to Farmland: Effects of Land-Use on Mammal Communities in the Tarangire-Manyara Ecosystem, Tanzania. Afr. J. Ecol. 2015, 53, 156–166. [Google Scholar] [CrossRef]
- Woodgate, Z.; Distiller, G.; O’Riain, J. Variation in Mammal Species Richness and Relative Abundance in the Karoo. Afr. J. Range Forage Sci. 2018, 35, 325–334. [Google Scholar] [CrossRef]
- Fabricius, C.; Burger, M.; Hockey, P.A.R. Comparing Biodiversity between Protected Areas and Adjacent Rangeland in Xeric Succulent Thicket, South Africa: Arthropods and Reptiles. J. Appl. Ecol. 2003, 40, 392–403. [Google Scholar] [CrossRef]
- Greve, M.; Chown, S.L.; van Rensburg, B.J.; Dallimer, M.; Gaston, K.J. The Ecological Effectiveness of Protected Areas: A Case Study for South African Birds. Anim. Conserv. 2011, 14, 295–305. [Google Scholar] [CrossRef]
- Shumba, T.; De Vos, A.; Biggs, R.; Esler, K.J.; Ament, J.M.; Clements, H.S. Effectiveness of Private Land Conservation Areas in Maintaining Natural Land Cover and Biodiversity Intactness. Glob. Ecol. Conserv. 2020, 22, e00935. [Google Scholar] [CrossRef]
- Ament, J.M.; Cumming, G.S. Scale Dependency in Effectiveness, Isolation, and Social-Ecological Spillover of Protected Areas. Conserv. Biol. 2016, 30, 846–855. [Google Scholar] [CrossRef]
- Curveira-Santos, G.; Sutherland, C.; Santos-Reis, M.; Swanepoel, L.H. Responses of Carnivore Assemblages to Decentralized Conservation Approaches in a South African Landscape. J. Appl. Ecol. 2021, 58, 92–103. [Google Scholar] [CrossRef]
- Drouilly, M.; Clark, A.; O’Riain, M.J. Multi-Species Occupancy Modelling of Mammal and Ground Bird Communities in Rangeland in the Karoo: A Case for Dryland Systems Globally. Biol. Conserv. 2018, 224, 16–25. [Google Scholar] [CrossRef]
- Wells, H.B.M.; Kimuyu, D.M.; Odadi, W.O.; Dougill, A.J.; Stringer, L.C.; Young, T.P. Wild and Domestic Savanna Herbivores Increase Smaller Vertebrate Diversity, but Less than Additively. J. Appl. Ecol. 2021, 58, 953–963. [Google Scholar] [CrossRef]
- Dean, W.R.J.; Milton, S.L. The Karoo: Ecological Patterns and Processes; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Milton, S.J. Studies of Herbivory and Vegetation Change in Karoo Shrublands. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 1993; p. 146. [Google Scholar]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland. In Strelitzia 19; South African National Biodiversity Institute: Pretoria, South Africa, 2006; pp. 1–30. ISBN 978-1-919976-21-1. [Google Scholar]
- Timm Hoffman, M.; Skowno, A.; Bell, W.; Mashele, S. Long-Term Changes in Land Use, Land Cover and Vegetation in the Karoo Drylands of South Africa: Implications for Degradation Monitoring. Afr. J. Range Forage Sci. 2018, 35, 209–221. [Google Scholar] [CrossRef]
- Allsopp, N.; Gaika, L.; Knight, R.; Monakisi, C.; Hoffman, M.T. The Impact of Heavy Grazing on an Ephemeral River System in the Succulent Karoo, South Africa. J. Arid Environ. 2007, 71, 82–96. [Google Scholar] [CrossRef]
- Brand, M.; Schutte-Vlok, A.; Huisamen, J. Anysberg Nature Reserve and World Heritage Site. Protected Area Management Plan 2018–2028. Cape Nature: Cape Town, South Africa, 2018; 170, Unpublished Internal Report. [Google Scholar]
- Cowling, R.M.; Pressey, R.L.; Lombard, A.T.; Desmet, P.G.; Ellis, A.G. From Representation to Persistence: Requirements for a Sustainable System of Conservation Areas in the Species-Rich Mediterranean-Climate Desert of Southern Africa. Divers. Distrib. 1999, 5, 51–71. [Google Scholar] [CrossRef]
- Hoffman, M.T.; Madden, C.F.; Erasmus, K.; Saayman, N.; Botha, J.C. The Impact of Indigenous Ungulate Herbivory over Five Years (2004–2008) on the Vegetation of the Little Karoo, South Africa. Afr. J. Range Forage Sci. 2009, 26, 169–179. [Google Scholar] [CrossRef]
- Mann, G.K.H.; Lagesse, J.V.; O’Riain, M.J.; Parker, D.M. Beefing Up Species Richness? The Effect of Land-Use on Mammal Diversity in an Arid Biodiversity Hotspot. Afr. J. Wildl. Res. 2015, 45, 321–331. [Google Scholar] [CrossRef]
- Cusack, J.J.; Dickman, A.J.; Rowcliffe, J.M.; Carbone, C.; Macdonald, D.W.; Coulson, T. Random versus Game Trail-Based Camera Trap Placement Strategy for Monitoring Terrestrial Mammal Communities. PLoS ONE 2015, 10, e0126379. [Google Scholar] [CrossRef]
- Rovero, F.; Ahumada, J. The Tropical Ecology, Assessment and Monitoring (TEAM) Network: An Early Warning System for Tropical Rain Forests. Sci. Total Environ. 2017, 574, 914–923. [Google Scholar] [CrossRef]
- Rich, L.N.; Miller, D.A.W.; Muñoz, D.J.; Robinson, H.S.; McNutt, J.W.; Kelly, M.J. Sampling Design and Analytical Advances Allow for Simultaneous Density Estimation of Seven Sympatric Carnivore Species from Camera Trap Data. Biol. Conserv. 2019, 233, 12–20. [Google Scholar] [CrossRef]
- Hofmeester, T.R.; Cromsigt, J.P.G.M.; Odden, J.; Andrén, H.; Kindberg, J.; Linnell, J.D.C. Framing Pictures: A Conceptual Framework to Identify and Correct for Biases in Detection Probability of Camera Traps Enabling Multi-Species Comparison. Ecol. Evol. 2019, 9, 2320–2336. [Google Scholar] [CrossRef] [PubMed]
- Larrucea, E.S.; Brussard, P.F.; Jaeger, M.M.; Barrett, H.R. Cameras, Coyotes, and the Assumption of Equal Detectability. J. Wildl. Manag. 2007, 71, 1682–1689. [Google Scholar] [CrossRef]
- Meek, P.D.; Ballard, G.; Claridge, A.; Kays, R.; Moseby, K.; O’Brien, T.; O’Connell, A.; Sanderson, J.; Swann, D.E.; Tobler, M.; et al. Recommended Guiding Principles for Reporting on Camera Trapping Research. Biodivers. Conserv. 2014, 23, 2321–2343. [Google Scholar] [CrossRef]
- O’Brien, T.G. Wildlife Picture Index and Biodiversity Monitoring: Issues and Future Directions. Anim. Conserv. 2010, 13, 350–352. [Google Scholar] [CrossRef]
- Tenan, S.; Brambilla, M.; Pedrini, P.; Sutherland, C. Quantifying Spatial Variation in the Size and Structure of Ecologically Stratified Communities. Methods Ecol. Evol. 2017, 8, 976–984. [Google Scholar] [CrossRef]
- Oberosler, V.; Tenan, S.; Zipkin, E.F.; Rovero, F. Poor Management in Protected Areas Is Associated with Lowered Tropical Mammal Diversity. Anim. Conserv. 2020, 23, 171–181. [Google Scholar] [CrossRef]
- Sutherland, C.; Brambilla, M.; Pedrini, P.; Tenan, S. A Multiregion Community Model for Inference about Geographic Variation in Species Richness. Methods Ecol. Evol. 2016, 7, 783–791. [Google Scholar] [CrossRef]
- Mackenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Andrew, J.; Langtimm, C.A. Estimating Site Occupancy Rates When Detection Probabilities Are Less Than One. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Dorazio, R.M.; Royle, J.A.; Söderström, B.; Glimskär, A. Estimating Species Richness and Accumulation by Modeling Species Occurrence and Detectability. Ecology 2006, 87, 842–854. [Google Scholar] [CrossRef]
- Kéry, M.; Royle, J.A. Hierarchical Bayes Estimation of Species Richness and Occupancy in Spatially Replicated Surveys. J. Appl. Ecol. 2008, 45, 589–598. [Google Scholar] [CrossRef]
- Tingley, M.W.; Nadeau, C.P.; Sandor, M.E. Multi-Species Occupancy Models as Robust Estimators of Community Richness. Methods Ecol. Evol. 2020, 11, 633–642. [Google Scholar] [CrossRef]
- Dorazio, R.M.; Royle, J.A. Estimating Size and Composition of Biological Communities by Modeling the Occurrence of Species. J. Am. Stat. Assoc. 2005, 100, 389–398. [Google Scholar] [CrossRef]
- Lehner, B.; Verdin, K.; Jarvis, A. New Global Hydrography Derived From Spaceborne Elevation Data. Eos Trans. Am. Geophys. Union 2008, 89, 93. [Google Scholar] [CrossRef]
- Petracca, L.S.; Funston, P.J.; Henschel, P.; Cohen, J.B.; Maclennan, S.; Frair, J.L. Modeling Community Occupancy from Line Transect Data: A Case Study with Large Mammals in Post-War Angola. Anim. Conserv. 2020, 23, 420–433. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, RG2004. [Google Scholar] [CrossRef]
- Sappington, J.M.; Longshore, K.M.; Thompson, D.B. Quantifying Landscape Ruggedness for Animal Habitat Analysis: A Case Study Using Bighorn Sheep in the Mojave Desert. J. Wildl. Manag. 2007, 71, 1419–1426. [Google Scholar] [CrossRef]
- Plummer, M. JAGS: A Program for Analysis of Bayesian Graphical Models Using Gibbs Sampling JAGS: Just Another Gibbs Sampler. In Proceedings of the 3rd International Workshop on Distributed Statistical Computing, Vienna, Austria, 20–22 March 2003; pp. 1–10. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing 2022. Available online: https://www.bibsonomy.org/bibtex/7469ffee3b07f9167cf47e7555041ee7 (accessed on 28 February 2023).
- Kellner, K. JagsUI: A Wrapper around Rjags to Streamline JAGS Analyses. R Package Version 2015, 1. [Google Scholar]
- Geweke, J. Comment: Inference and Prediction in the Presence of Uncertainty and Determinism. Stat. Sci. 1992, 7, 94–101. [Google Scholar] [CrossRef]
- Gelman, A.; Hwang, J.; Vehtari, A. Understanding Predictive Information Criteria for Bayesian Models. Stat. Comput. 2014, 24, 997–1016. [Google Scholar] [CrossRef]
- Davies-Mostert, H.T. Overcoming Barriers to Understanding the Biodiversity Contribution of Private Ranchlands. Anim. Conserv. 2014, 17, 399–400. [Google Scholar] [CrossRef]
- Arbieu, U.; Grünewald, C.; Martín-López, B.; Schleuning, M.; Böhning-Gaese, K. Large Mammal Diversity Matters for Wildlife Tourism in Southern African Protected Areas: Insights for Management. Ecosyst. Serv. 2018, 31, 481–490. [Google Scholar] [CrossRef]
- Lynch, K.; Vorster, L.; Vorster, P. Sanbona Wildlife Reserve: Environmental Management Plan. Unpublished Internal Report. 2015; 27, Unpublished. [Google Scholar]
- Lindsey, P.A.; Masterson, C.L.; Beck, A.L.; Romañach, S. Ecological, Social and Financial Issues Related to Fencing as a Conservation Tool in Africa. In Fencing for Conservation; Somers, M.J., Hayward, M., Eds.; Springer: New York, NY, USA, 2012; pp. 215–234. ISBN 9781461409021. [Google Scholar]
- Milton, S.J.; Dean, W.R.J. Anthropogenic Impacts and Implications for Ecological Restoration in the Karoo, South Africa. Anthropocene 2021, 36, 100307. [Google Scholar] [CrossRef]
- Ramesh, T.; Kalle, R.; Rosenlund, H.; Downs, C.T. Native Habitat and Protected Area Size Matters: Preserving Mammalian Assemblages in the Maputaland Conservation Unit of South Africa. For. Ecol. Manag. 2016, 360, 20–29. [Google Scholar] [CrossRef]
- Van der Weyde, L.K.; Tobler, M.W.; Gielen, M.C.; Cozzi, G.; Weise, F.J.; Adams, T.; Bauer, D.; Bennitt, E.; Bowles, M.; Brassine, A.; et al. Collaboration for Conservation: Assessing Countrywide Carnivore Occupancy Dynamics from Sparse Data. Divers. Distrib. 2022, 28, 917–929. [Google Scholar] [CrossRef]
- Tyrrell, P.; Toit, J.T.; Macdonald, D.W. Conservation beyond Protected Areas: Using Vertebrate Species Ranges and Biodiversity Importance Scores to Inform Policy for an East African Country in Transition. Conserv. Sci. Pract. 2020, 2, e136. [Google Scholar] [CrossRef]
- Verschueren, S.; Briers-Louw, W.D.; Monterroso, P.; Marker, L. Local-Scale Variation in Land Use Practice Supports a Diverse Carnivore Guild on Namibian Multiple-Use Rangeland. Rangel. Ecol. Manag. 2021, 79, 64–76. [Google Scholar] [CrossRef]
- Harris, N.C.; Mills, K.L.; Harissou, Y.; Hema, E.M.; Gnoumou, I.T.; VanZoeren, J.; Abdel-Nasser, Y.I.; Doamba, B. First Camera Survey in Burkina Faso and Niger Reveals Human Pressures on Mammal Communities within the Largest Protected Area Complex in West Africa. Conserv. Lett. 2019, 12, e12667. [Google Scholar] [CrossRef]
- Thapa, A.; Pradhan, P.K.; Joshi, B.D.; Mukherjee, T.; Thakur, M.; Chandra, K.; Sharma, L.K. Non-Protected Areas Demanding Equitable Conservation Strategies as of Protected Areas in the Central Himalayan Region. PLoS ONE 2021, 16, e0255082. [Google Scholar] [CrossRef]
- Bellón, B.; Henry, D.; Renaud, P.C.; Roque, F.d.O.; Santos, C.C.; Melo, I.; Arvor, D.; de Vos, A. Landscape Drivers of Mammal Habitat Use and Richness in a Protected Area and Its Surrounding Agricultural Lands. Agric. Ecosyst. Environ. 2022, 334, 107989. [Google Scholar] [CrossRef]
- Owen-Smith, N.; Cromsigt, J.P.G.M.; Le Roux, E. Smaller Ungulates Are First to Incur Imminent Extirpation from an African Protected Area. Biol. Conserv. 2017, 216, 108–114. [Google Scholar] [CrossRef]
- Rottstock, T.; Göttert, T.; Zeller, U. Relatively Undisturbed African Savannas—An Important Reference for Assessing Wildlife Responses to Livestock Grazing Systems in European Rangelands. Glob. Ecol. Conserv. 2020, 23, e01124. [Google Scholar] [CrossRef]
- Holechek, J.; Valdez, R. Wildlife Conservation on the Rangelands of Eastern and Southern Africa: Past, Present, and Future. Rangel. Ecol. Manag. 2018, 71, 245–258. [Google Scholar] [CrossRef]
- Cox, R.L.; Underwood, E.C. The Importance of Conserving Biodiversity Outside of Protected Areas in Mediterranean Ecosystems. PLoS ONE 2011, 6, e14508. [Google Scholar] [CrossRef] [PubMed]
- TOIT, J.T. DU The Feeding Ecology of a Very Small Ruminant, the Steenbok (Raphicerus Campestris). Afr. J. Ecol. 1993, 31, 35–48. [Google Scholar] [CrossRef]
- Lima, S.L.; Dill, L.M. Behavioral Decisions Made under the Risk of Predation: A Review and Prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Mayer, M.; Fog Bjerre, D.H.; Sunde, P. Better Safe than Sorry: The Response to a Simulated Predator and Unfamiliar Scent by the European Hare. Ethology 2020, 126, 704–715. [Google Scholar] [CrossRef]
- Young, T.P.; Porensky, L.M.; Riginos, C.; Veblen, K.E.; Odadi, W.O.; Kimuyu, D.M.; Charles, G.K.; Young, H.S. Relationships Between Cattle and Biodiversity in Multiuse Landscape Revealed by Kenya Long-Term Exclosure Experiment. Rangel. Ecol. Manag. 2018, 71, 281–291. [Google Scholar] [CrossRef]
- Drouilly, M.; Nattrass, N.; O’Riain, M.J. Dietary Niche Relationships among Predators on Farmland and a Protected Area. J. Wildl. Manag. 2018, 82, 507–518. [Google Scholar] [CrossRef]
- Beinart, W. The Rise of Conservation in South Africa: Settlers, Livestock, and the Environment; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- McCain, C.M.; Beck, J. Species Turnover in Vertebrate Communities along Elevational Gradients Is Idiosyncratic and Unrelated to Species Richness. Glob. Ecol. Biogeogr. 2016, 25, 299–310. [Google Scholar] [CrossRef]
- Berryman, E.M.; Barnard, H.R.; Adams, H.R.; Burns, M.A.; Gallo, E.; Brooks, P.D. Complex Terrain Alters Temperature and Moisture Limitations of Forest Soil Respiration across a Semiarid to Subalpine Gradient. J. Geophys. Res. Biogeosciences 2015, 120, 707–723. [Google Scholar] [CrossRef]
- Fritz, S.A.; Eronen, J.T.; Schnitzler, J.; Hof, C.; Janis, C.M.; Mulch, A.; Böhning-Gaese, K.; Graham, C.H. Twenty-Million-Year Relationship between Mammalian Diversity and Primary Productivity. Proc. Natl. Acad. Sci. USA 2016, 113, 10908–10913. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global Consequences of Land Use. Science 2005, 309, 570. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, E.W.; Jaiteh, M.; Levy, M.A.; RedfordD, K.H.; Wannebo, A.V.; Woolmer, G. The Human Footprint and the Last of the Wild. Bioscience 2002, 52, 891–904. [Google Scholar] [CrossRef]
- Gebert, F.; Njovu, H.K.; Treydte, A.C.; Steffan-Dewenter, I.; Peters, M.K. Primary Productivity and Habitat Protection Predict Elevational Species Richness and Community Biomass of Large Mammals on Mt. Kilimanjaro. J. Anim. Ecol. 2019, 88, 1860–1872. [Google Scholar] [CrossRef]
- Ferreira, G.B.; Collen, B.; Newbold, T.; Oliveira, M.J.R.; Pinheiro, M.S.; de Pinho, F.F.; Rowcliffe, M.; Carbone, C. Strict Protected Areas Are Essential for the Conservation of Larger and Threatened Mammals in a Priority Region of the Brazilian Cerrado. Biol. Conserv. 2020, 251, 108762. [Google Scholar] [CrossRef]
- Chen, C.; Brodie, J.F.; Kays, R.; Davies, T.J.; Liu, R.; Fisher, J.T.; Ahumada, J.; McShea, W.; Sheil, D.; Agwanda, B.; et al. Global Camera Trap Synthesis Highlights the Importance of Protected Areas in Maintaining Mammal Diversity. Conserv. Lett. 2022, 15, e12865. [Google Scholar] [CrossRef]
- Parker, K.; De Vos, A.; Clements, H.S.; Biggs, D.; Biggs, R. Impacts of a Trophy Hunting Ban on Private Land Conservation in South African Biodiversity Hotspots. Conserv. Sci. Pract. 2020, 2, e214. [Google Scholar] [CrossRef]
- Gallo, J.A.; Pasquini, L.; Reyers, B.; Cowling, R.M. The Role of Private Conservation Areas in Biodiversity Representation and Target Achievement within the Little Karoo Region, South Africa. Biol. Conserv. 2009, 142, 446–454. [Google Scholar] [CrossRef]
- Torres-Romero, E.J.; Giordano, A.J.; Ceballos, G.; López-Bao, J.V. Reducing the Sixth Mass Extinction: Understanding the Value of Human-Altered Landscapes to the Conservation of the World’s Largest Terrestrial Mammals. ABiol. Conserv. 2020, 249, 108706. [Google Scholar] [CrossRef]
- Clements, H.S.; Baum, J.; Cumming, G.S. Money and Motives: An Organizational Ecology Perspective on Private Land Conservation. Biol. Conserv. 2016, 197, 108–115. [Google Scholar] [CrossRef]




| Land-Use | Parameter | Mean | Lower | Upper | |
|---|---|---|---|---|---|
| Occupancy (Ψ) | Farmland | Intercept | |||
| Distance to drainage (m) | 0.05 | −0.05 | 0.14 | ||
| Vegetation productivity (MSAVI2) | −0.11 | −0.22 | 0.00 | ||
| Terrain ruggedness (TRI) | 0.05 | −0.05 | 0.15 | ||
| PA | Intercept | ||||
| Distance to drainage (m) | −0.11 | −0.32 | 0.09 | ||
| Vegetation productivity (MSAVI2) | 0.23 | 0.05 | 0.40 | ||
| Terrain ruggedness (TRI) | −0.67 | −1.11 | −0.27 | ||
| PPA | Intercept | ||||
| Distance to drainage (m) | −0.15 | −0.26 | −0.03 | ||
| Vegetation productivity (MSAVI2) | 0.14 | 0.02 | 0.27 | ||
| Terrain ruggedness (TRI) | −0.31 | −0.56 | −0.08 | ||
| Detection (p) | Farmland PA PPA | Intercept | |||
| Vegetation productivity | −0.03 | −0.13 | 0.05 | ||
| Vegetation productivity (MSAVI2) | −1.19 | −0.28 | −0.11 | ||
| Vegetation productivity (TRI) | 0.09 | 0.00 | 0.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woodgate, Z.; Drouilly, M.; Distiller, G.; O’Riain, M.J. The Effect of Multi-Use Landscapes on Mammal Assemblages and Its Implication for Conservation. Land 2023, 12, 599. https://doi.org/10.3390/land12030599
Woodgate Z, Drouilly M, Distiller G, O’Riain MJ. The Effect of Multi-Use Landscapes on Mammal Assemblages and Its Implication for Conservation. Land. 2023; 12(3):599. https://doi.org/10.3390/land12030599
Chicago/Turabian StyleWoodgate, Zoë, Marine Drouilly, Greg Distiller, and M. Justin O’Riain. 2023. "The Effect of Multi-Use Landscapes on Mammal Assemblages and Its Implication for Conservation" Land 12, no. 3: 599. https://doi.org/10.3390/land12030599
APA StyleWoodgate, Z., Drouilly, M., Distiller, G., & O’Riain, M. J. (2023). The Effect of Multi-Use Landscapes on Mammal Assemblages and Its Implication for Conservation. Land, 12(3), 599. https://doi.org/10.3390/land12030599