
_Kazoglou.png)
Opportunities for Adaptation to Climate Change of Extensively Grazed Pastures in the Central Apennines (Italy)

 , ,
, ,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites, Experimental Layout, and Data Collection
2.2. Climate Scenarios and Models
2.3. The Grassland Model
2.4. Simulation Design

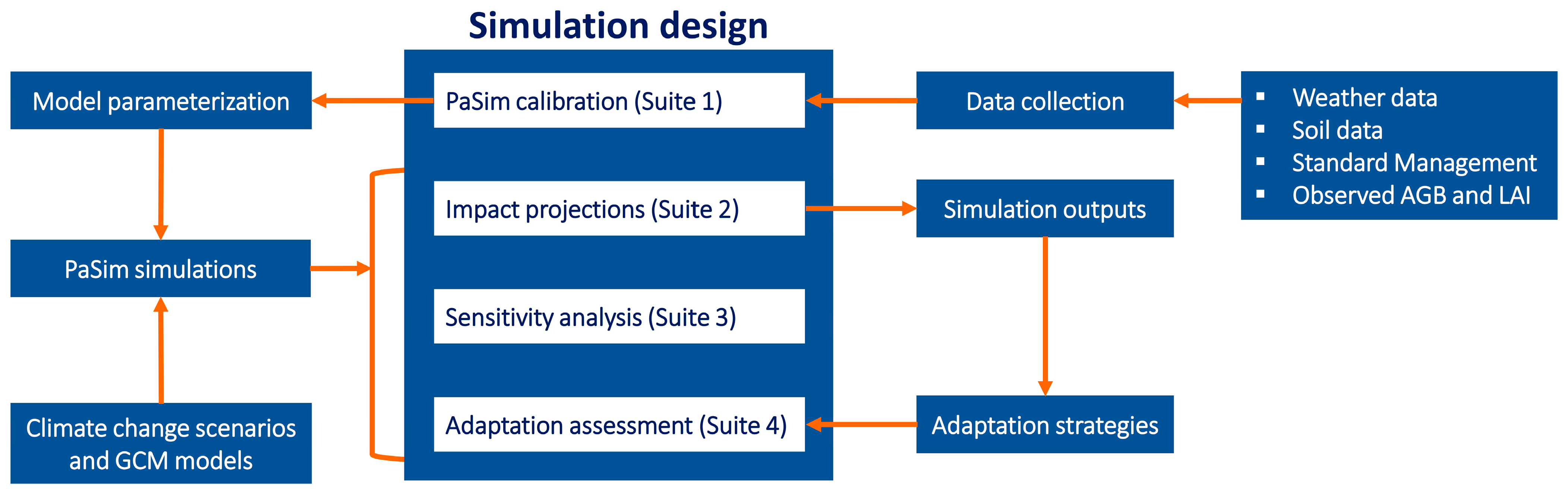{kind=link}
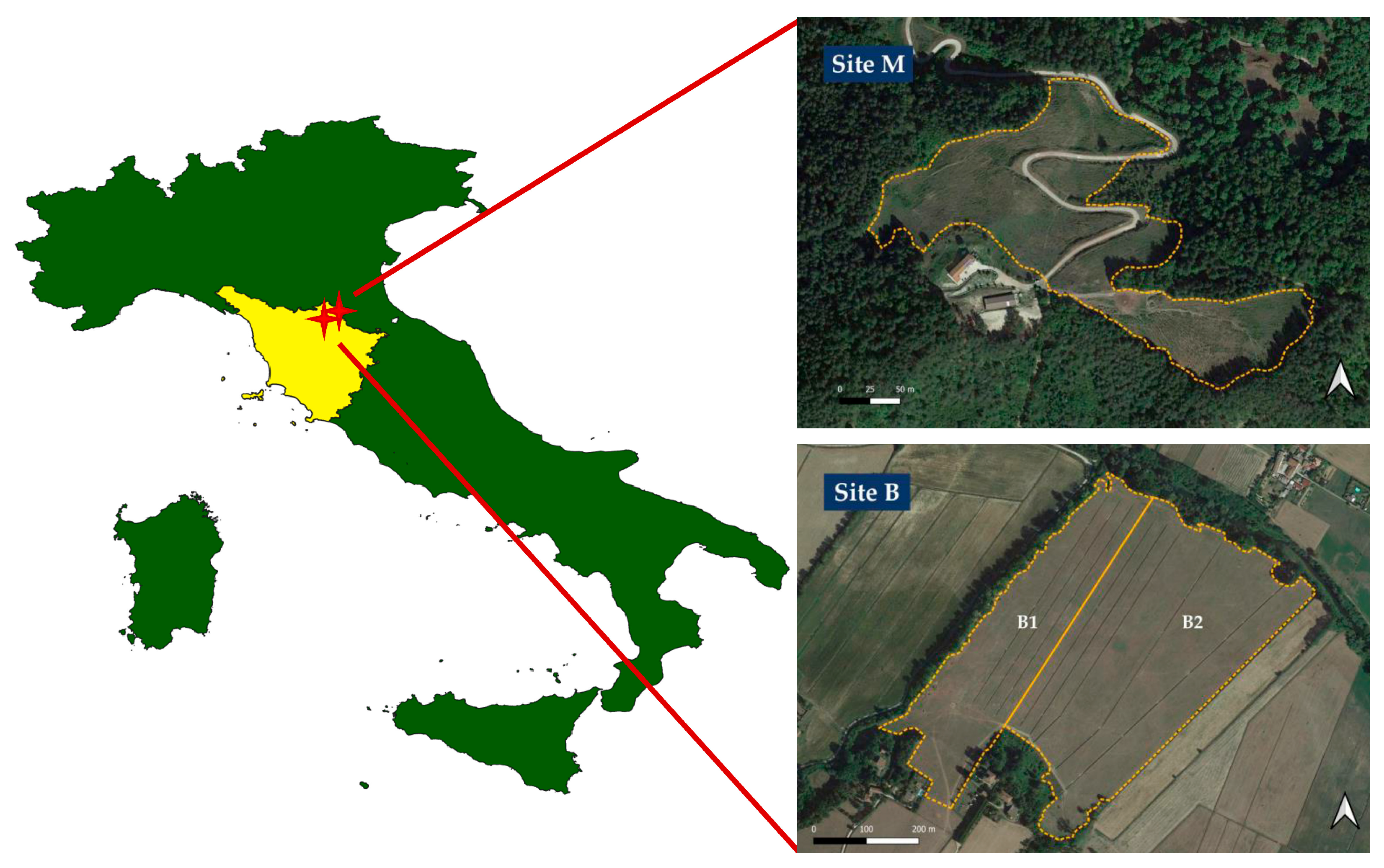{kind=link}
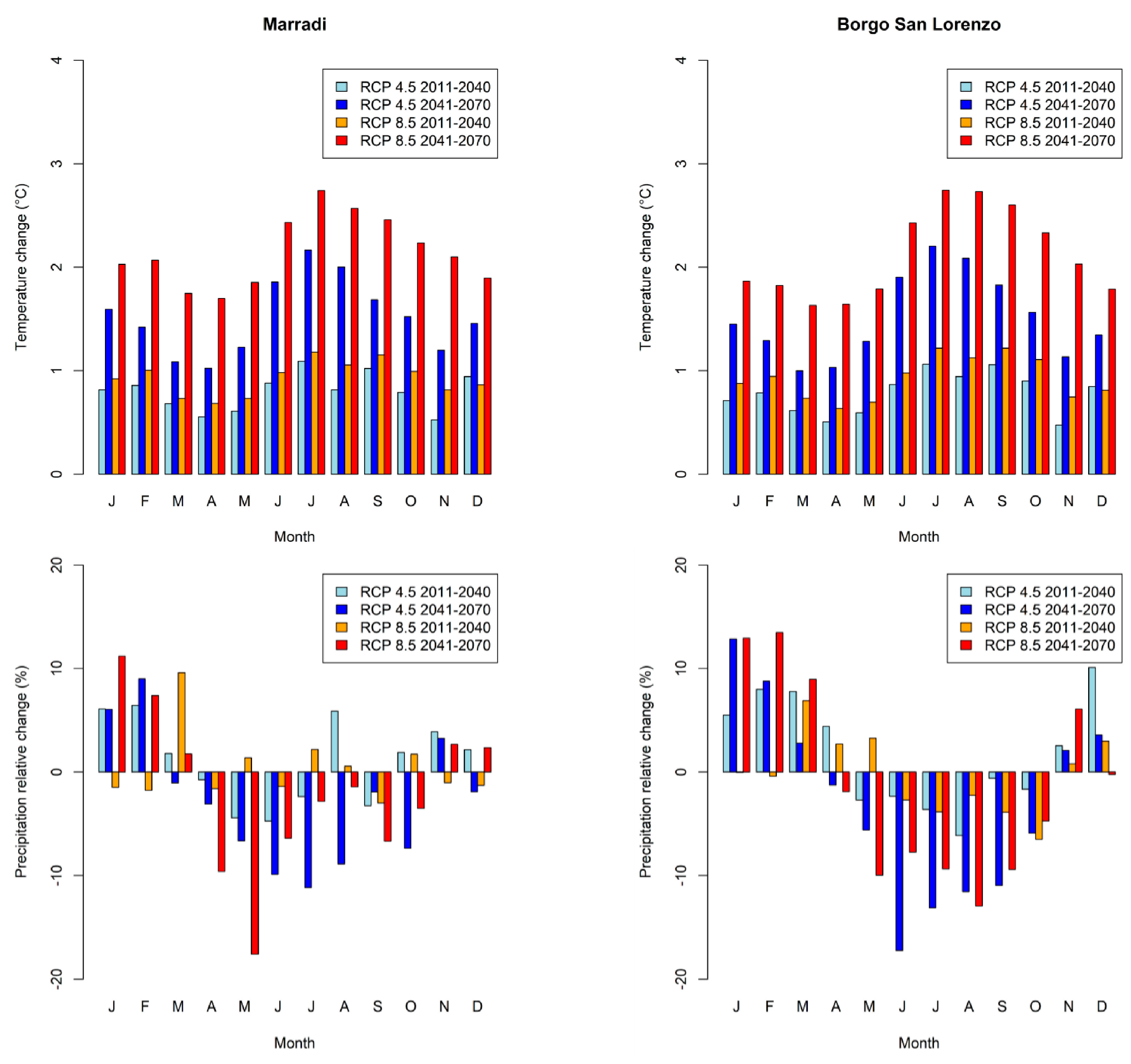{kind=link}
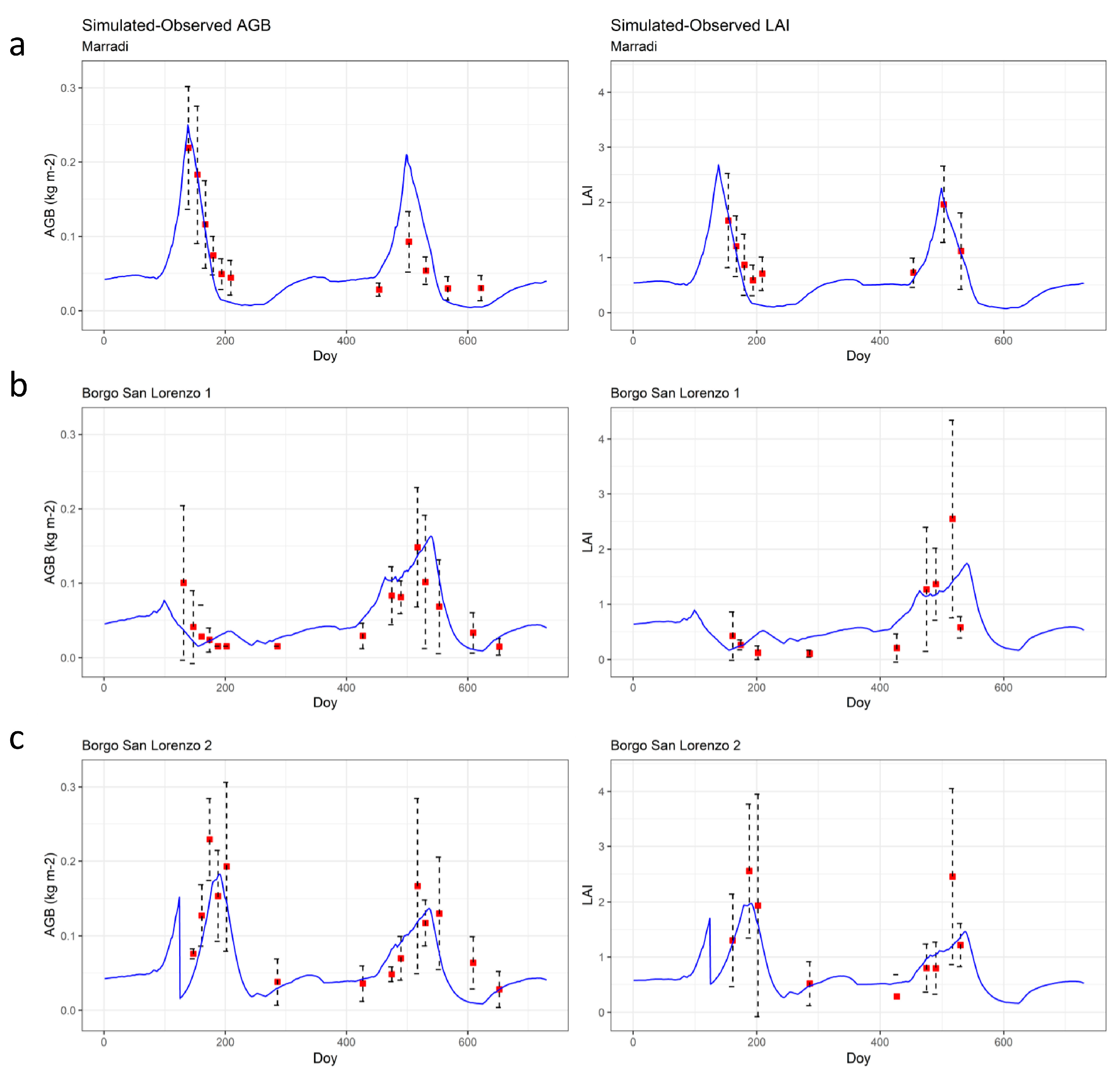{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Output | Acronym | Unit | Description |
|---|---|---|---|---|
| Date | Growing season start | GSs | day of year (doy) | Day after seven consecutive days with a mean air temperature ≥8 °C from 1 January onwards [67] |
| Growing season end | GSe | Day after seven consecutive days with a mean air temperature <8 °C from 1 July onwards [67] | ||
| Biomass peak date | BPd | Day of the year with the highest value of aboveground biomass | ||
| Count | Growing season length | GS | days | Number of days between the GSs and GSe |
| Amount | Biomass peak | BP | kg DM m−2 | Aboveground biomass value at the peak date |
| Aboveground biomass | AGB | kg DM m−2 | Aboveground biomass values | |
| Net ecosystem exchange | NEE | kg C m−2 yr−1 | C-N fluxes (annual balance) (These include emissions from ecosystem respiration, RECO = plant + soil + animal respiration, as well as estimates of the plant production of organic compounds from atmospheric CO2 (GPP: gross primary production) and other system variables: NEE = RECO - GPP, enteric emissions of CH4 from grazing animals and N2O emissions from the N cycle) | |
| Methane | CH4 | kg C m−2 yr−1 | ||
| Nitrous oxide | N2O | kg N m−2 yr−1 | ||
| Soil water content | SWC | m3 m−3 | Annual mean of daily soil water content values (0.35-m topsoil). In Supplementary Materials. |
3. Results
3.1. Climate Analysis
3.2. Suite 1 of Simulations: Evaluation of the Model against Observed Data
3.3. Suites 2, 3, and 4 of Simulations: Impacts of Future Scenarios, Sensitivity to Weather Inputs, and Adaptation Strategies
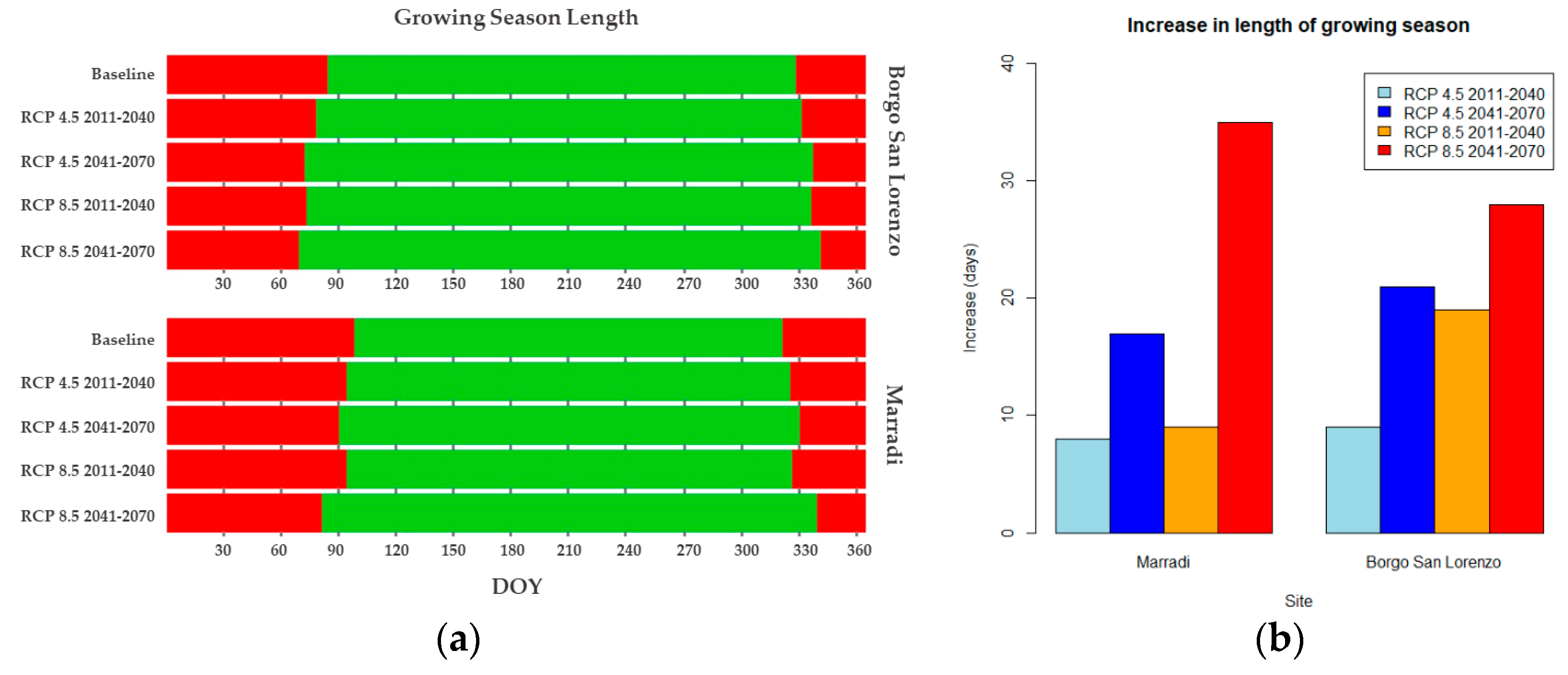
3.4. Growing Season
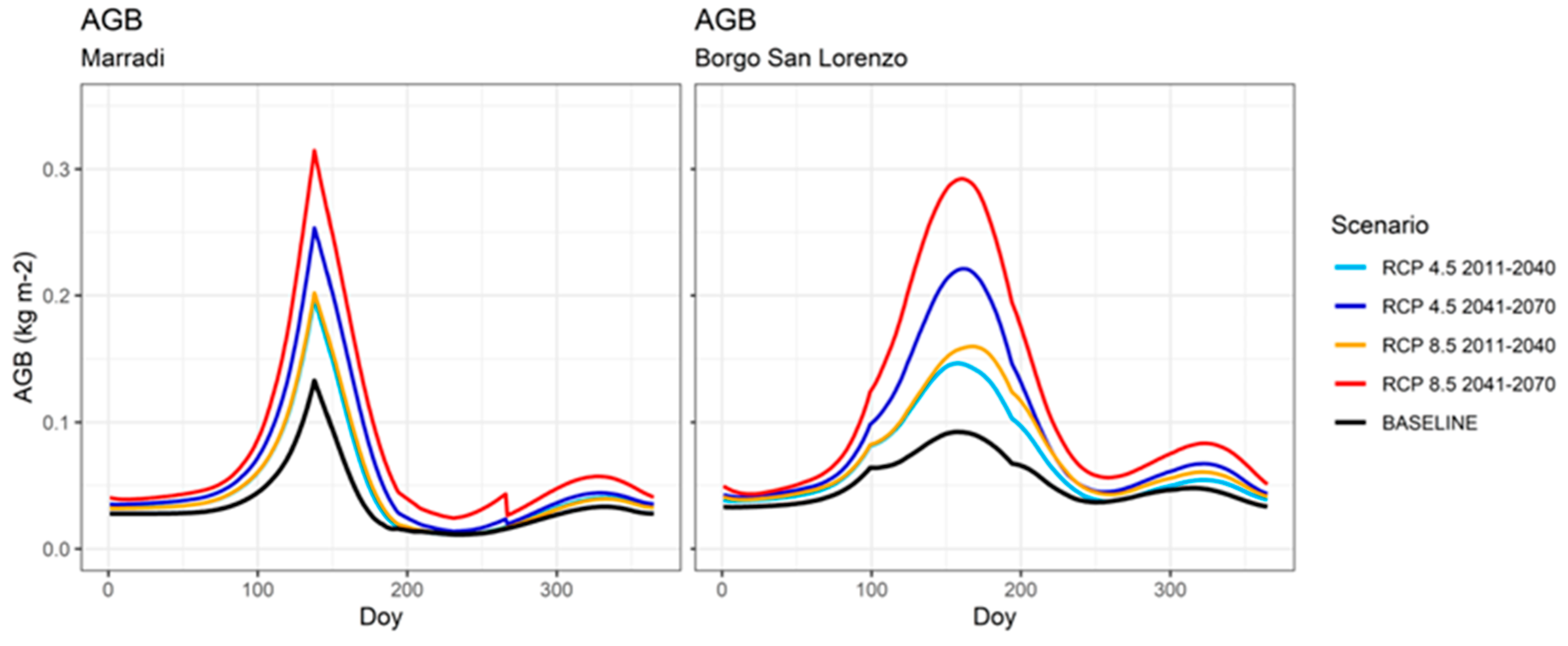
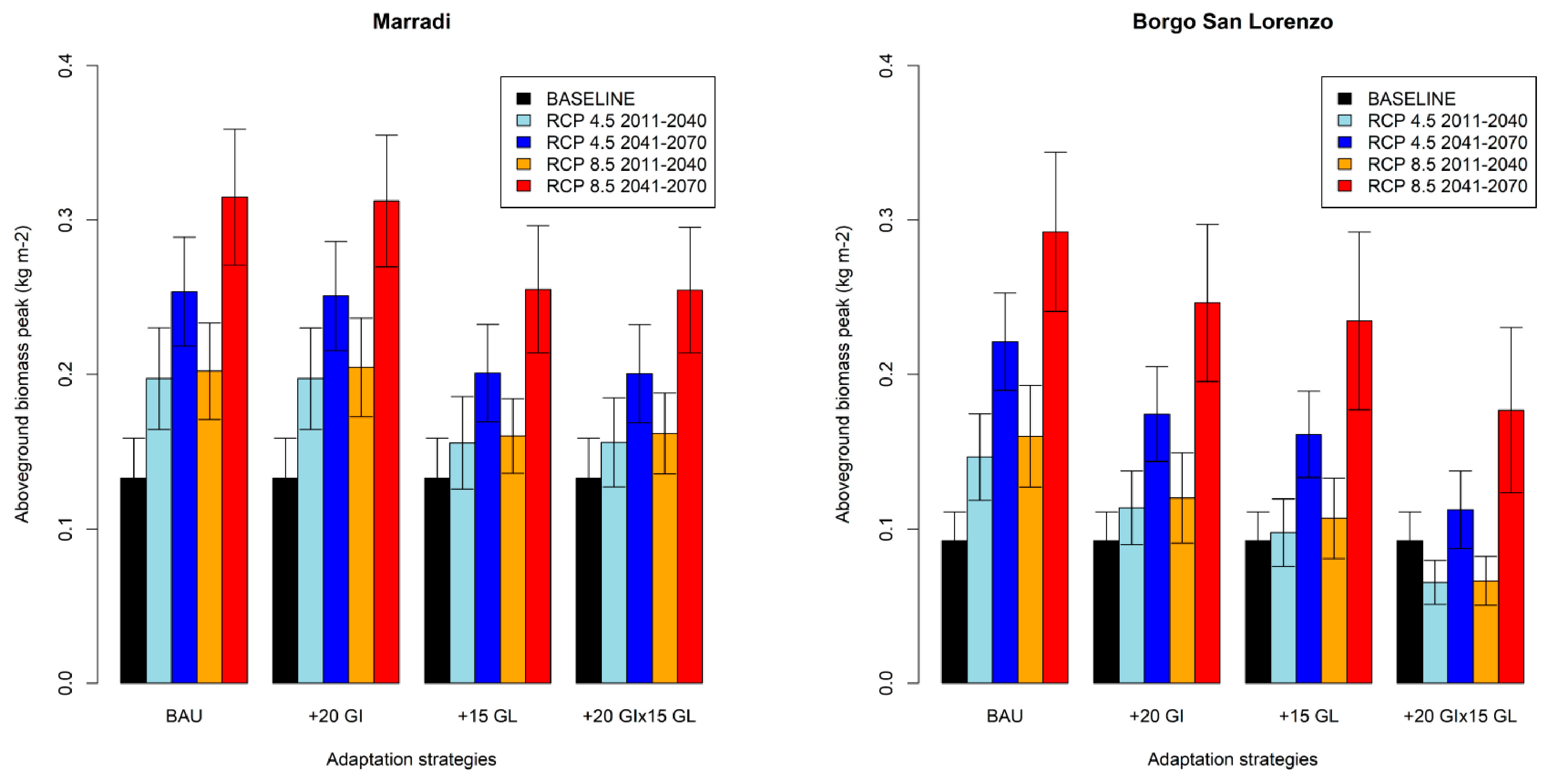
3.5. Aboveground Biomass
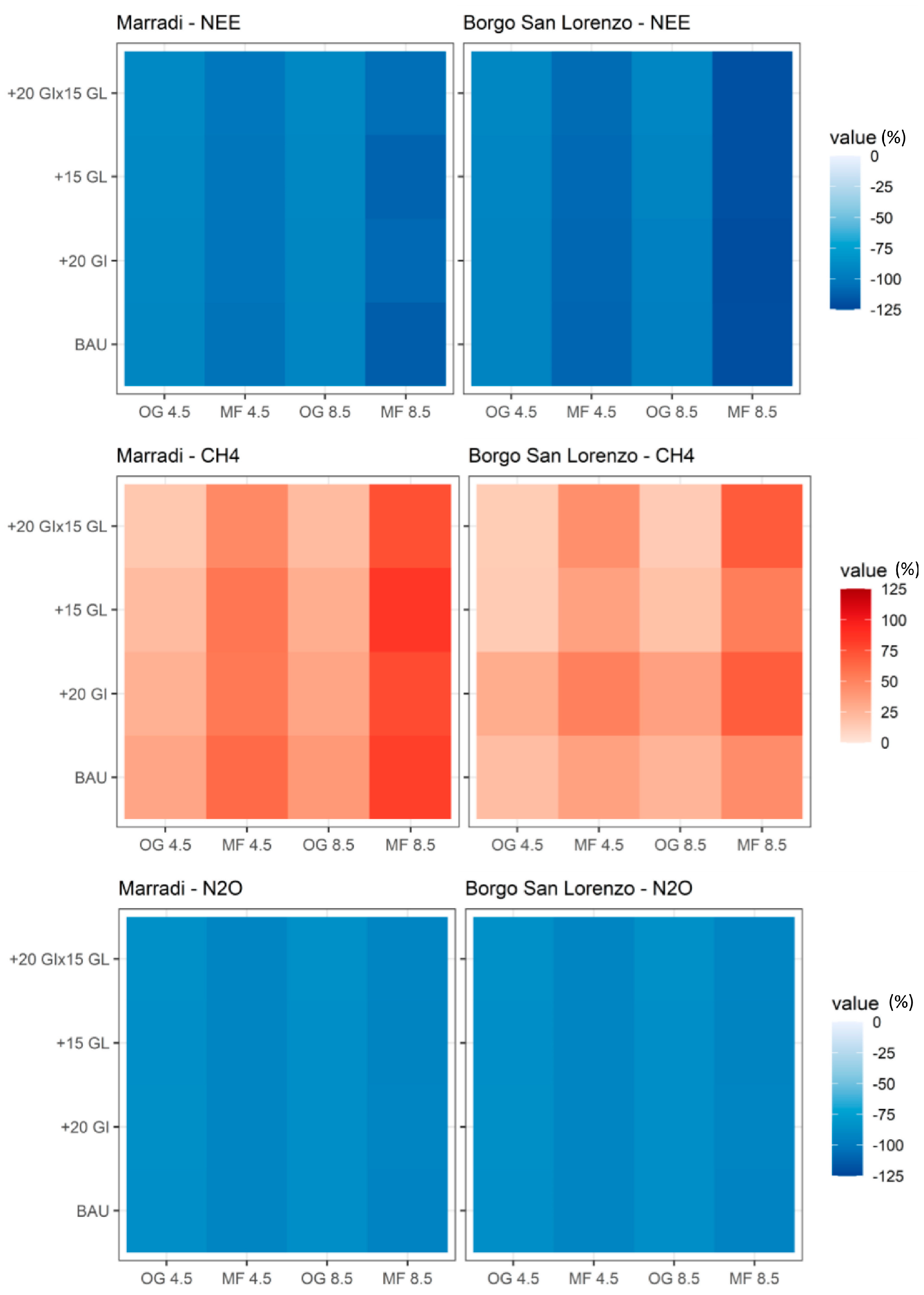
3.6. Carbon–nitrogen Fluxes
4. Discussion
4.1. Model Parameterisation
4.2. Uncertainties in Climate Change Impacts and Adaptation Strategies
4.3. Consequences for Grassland Sustainability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dillon, P. The Evolution of Grassland in the European Union in Terms of Utilisation, Productivity, Food Security and the Importance of Adoption of Technical Innovations in Increasing Sustainability of Pasture-Based Ruminant Production Systems. Grassl. Sci. Eur. 2018, 23, 3–15. [Google Scholar]
- Mara, F.P.O. The Role of Grasslands in Food Security and Climate Change. Ann. Bot. 2012, 210, 1263–1270. [Google Scholar] [CrossRef]
- Barbour, R.; Young, R.H.; Wilkinson, J.M. Production of Meat and Milk from Grass in the United Kingdom. Agronomy 2022, 12, 914. [Google Scholar] [CrossRef]
- ISTAT. Available online: Http://Dati.Istat.It/Index.Aspx?Queryid=33704 (accessed on 5 December 2022).
- Burrascano, S.; Caccianiga, M.; Gigante, D. Dry Grasslands Habitat Types in Italy. 2010. Bulletin of European Dry Grassland Group. Available online: https://air.unimi.it/handle/2434/155321 (accessed on 20 January 2023).
- Dibari, C.; Pulina, A.; Argenti, G.; Aglietti, C.; Bindi, M.; Moriondo, M.; Mula, L.; Pasqui, M.; Seddaiu, G.; Roggero, P.P. Climate Change Impacts on the Alpine, Continental and Mediterranean Grassland Systems of Italy: A Review. Ital. J. Agron. 2021, 16, 1843. [Google Scholar] [CrossRef]
- Cavallero, A.; Aceto, P.; Gorlier, A.; Lombardi, G.; Lonati, M.; Martinasso, B.; Tagliatori, C. I Tipi Pastorali Delle Alpi Piemontesi; Alberto Perdisa Editore: Bologna, Italy, 2007. (In Italian) [Google Scholar]
- Argenti, G.; Bottai, L.; Chiesi, M.; Maselli, F.; Staglianò, N.; Targetti, S. Analisi e Valutazione Di Pascoli Montani Attraverso l’integrazione Di Dati Multispettrali e Ausiliari. Riv. Ital. Telerilevamento 2011, 43, 45–57. (In Italian) [Google Scholar]
- Orlandi, S.; Probo, M.; Sitzia, T.; Trentanovi, G.; Garbarino, M.; Lombardi, G.; Lonati, M. Environmental and Land Use Determinants of Grassland Patch Diversity in the Western and Eastern Alps under Agro-Pastoral Abandonment. Biodivers. Conserv. 2016, 25, 275–293. [Google Scholar] [CrossRef]
- Targetti, S.; Messeri, A.; Staglianò, N.; Argenti, G. Leaf Functional Traits for the Assessment of Succession Following Management in Semi-Natural Grasslands: A Case Study in the North Apennines, Italy. Appl. Veg. Sci. 2013, 16, 325–332. [Google Scholar] [CrossRef]
- Metzger, M.J.; Bunce, R.G.H.; Jongman, R.H.G.; Mücher, C.A.; Watkins, J.W. A Climatic Stratification of the Environment of Europe. Glob. Ecol. Biogeogr. 2005, 14, 549–563. [Google Scholar] [CrossRef]
- Bengtsson, J.; Bullock, J.M.; Egoh, B.; Everson, C.; Everson, T.; O’Connor, T.; O’Farrell, P.J.; Smith, H.G.; Lindborg, R. Grasslands—More Important for Ecosystem Services than You Might Think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- Hao, R.; Yu, D.; Liu, Y.; Liu, Y.; Qiao, J.; Wang, X.; Du, J. Impacts of Changes in Climate and Landscape Pattern on Ecosystem Services. Sci. Total Environ. 2017, 579, 718–728. [Google Scholar] [CrossRef]
- Ponzetta, M.P.; Cervasio, F.; Crocetti, C.; Messeri, A.; Argenti, G. Habitat Improvements with Wildlife Purposes in a Grazed Area on the Apennine Mountains. Ital. J. Agron. 2010, 5, 233–238. [Google Scholar] [CrossRef]
- Tamburini, G.; Aguilera, G.; Öckinger, E. Grasslands Enhance Ecosystem Service Multifunctionality above and Below-Ground in Agricultural Landscapes. J. Appl. Ecol. 2022, 3061–3071. [Google Scholar] [CrossRef]
- Wepking, C.; Mackin, H.C.; Raff, Z.; Shrestha, D.; Orfanou, A.; Booth, E.G.; Kucharik, C.J.; Gratton, C.; Jackson, R.D. Perennial Grassland Agriculture Restores Critical Ecosystem Functions in the U.S. Upper Midwest. Front. Sustain. Food Syst. 2022, 6, 1010280. [Google Scholar] [CrossRef]
- Sándor, R.; Ehrhardt, F.; Brilli, L.; Carozzi, M.; Recous, S.; Smith, P.; Snow, V.; Soussana, J.F.; Dorich, C.D.; Fuchs, K.; et al. The Use of Biogeochemical Models to Evaluate Mitigation of Greenhouse Gas Emissions from Managed Grasslands. Sci. Total Environ. 2018, 642, 292–306. [Google Scholar] [CrossRef]
- Smith, P.; Martino, D.; Cai, Z.; Gwary, D.; Janzen, H.; Kumar, P.; McCarl, B.; Ogle, S.; O’Mara, F.; Rice, C.; et al. Greenhouse Gas Mitigation in Agriculture. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 789–813. [Google Scholar] [CrossRef]
- Oates, L.G.; Jackson, R.D. Livestock Management Strategy Affects Net Ecosystem Carbon Balance of Subhumid Pasture. Rangel. Ecol. Manag. 2014, 67, 19–29. [Google Scholar] [CrossRef]
- Franzluebbers, A.J. Cattle Grazing Effects on the Environment: Greenhouse Gas Emissions and Carbon Footprint; Elsevier Inc.: Cambridge, MA, USA, 2020; ISBN 9780128144749. [Google Scholar]
- Guillaume, T.; Makowski, D.; Libohova, Z.; Elfouki, S.; Fontana, M.; Leifeld, J.; Bragazza, L.; Sinaj, S. Geoderma Carbon Storage in Agricultural Topsoils and Subsoils Is Promoted by Including Temporary Grasslands into the Crop Rotation. Geoderma 2022, 422, 115937. [Google Scholar] [CrossRef]
- Soussana, J.F.; Tallec, T.; Blanfort, V. Mitigating the Greenhouse Gas Balance of Ruminant Production Systems through Carbon Sequestration in Grasslands. Anim. Int. J. Anim. Biosci. 2010, 4, 334–350. [Google Scholar] [CrossRef]
- Dass, P.; Houlton, B.Z.; Wang, Y.; Warlind, D. Grasslands May Be More Reliable Carbon Sinks than Forests in California. Environ. Res. Lett. 2018, 13, 074027. [Google Scholar] [CrossRef]
- Steinfeld, H.; Wassenaar, T. The Role of Livestock Production in Carbon and Nitrogen Cycles. Annu. Rev. Environ. Resour. 2007, 32, 271–296. [Google Scholar] [CrossRef]
- Hortnagl, L.; Barthel, M.; Buchmann, N.; Eugster, W.; Butterbach-bahl, K.; Eugenio, D.; Zeeman, M.; Lu, H.; Kiese, R.; Bahn, M.; et al. Greenhouse Gas Fluxes over Managed Grasslands in Central Europe S. Glob. Chang. Biol. 2018, 24, 1843–1872. [Google Scholar] [CrossRef] [PubMed]
- Soussana, J.F.; Allard, V.; Pilegaard, K.; Ambus, P.; Amman, C.; Campbell, C.; Raschi, A.; Baronti, S.; Rees, R.M.; Skiba, U.; et al. Full Accounting of the Greenhouse Gas (CO2, N2O, CH4) Budget of Nine European Grassland Sites. Agric. Ecosyst. Environ. 2007, 121, 121–134. [Google Scholar] [CrossRef]
- Toreti, A.; Desiato, F. Temperature Trend over Italy from 1961 to 2004. Theor. Appl. Climatol. 2008, 91, 51–58. [Google Scholar] [CrossRef]
- Tomozeiu, R.; Pasqui, M.; Quaresima, S. Future Changes of Air Temperature over Italian Agricultural Areas: A Statistical Downscaling Technique Applied to 2021–2050 and 2071–2100 Periods. Meteorol. Atmos. Phys. 2018, 130, 543–563. [Google Scholar] [CrossRef]
- Scocco, P.; Piermarteri, K.; Malfatti, A.; Tardella, F.M.; Catorci, A. Increase of Drought Stress Negatively Affects the Sustainability of Extensive Sheep Farming in Sub-Mediterranean Climate. J. Arid Environ. 2016, 128, 50–58. [Google Scholar] [CrossRef]
- Chelli, S.; Wellstein, C.; Campetella, G.; Canullo, R.; Tonin, R.; Zerbe, S.; Gerdol, R. Climate Change Response of Vegetation across Climatic Zones in Italy. Clim. Res. 2017, 71, 249–262. [Google Scholar] [CrossRef]
- Petriccione, B.; Bricca, A. Thirty Years of Ecological Research at the Gran Sasso d’Italia LTER Site: Climate Change in Action. Nat. Conserv. 2019, 34, 9–39. [Google Scholar] [CrossRef]
- Dibari, C.; Costafreda-Aumedes, S.; Argenti, G.; Bindi, M.; Carotenuto, F.; Moriondo, M.; Padovan, G.; Pardini, A.; Staglianò, N.; Vagnoli, C.; et al. Expected Changes to Alpine Pastures in Extent and Composition under Future Climate Conditions. Agronomy 2020, 10, 926. [Google Scholar] [CrossRef]
- Stanisci, A.; Frate, L.; Morra Di Cella, U.; Pelino, G.; Petey, M.; Siniscalco, C.; Carranza, M.L. Short-Term Signals of Climate Change in Italian Summit Vegetation: Observations at Two GLORIA Sites. Plant Biosyst. 2016, 150, 227–235. [Google Scholar] [CrossRef]
- Evangelista, A.; Frate, L.; Carranza, M.L.; Attorre, F.; Pelino, G.; Stanisci, A. Changes in Composition, Ecology and Structure of High-Mountain Vegetation: A Re-Visitation Study over 42 Years. AoB Plants 2016, 8, 1–11. [Google Scholar] [CrossRef]
- Bebeley, J.F.; Kamara, A.Y.; Jibrin, J.M.; Akinseye, F.M.; Tofa, A.I.; Adam, A.M. Evaluation and Application of the CROPGRO-Soybean Model for Determining Optimum Sowing Windows of Soybean in the Nigeria Savannas. Sci. Rep. 2022, 12, 6747. [Google Scholar] [CrossRef] [PubMed]
- Kamilaris, C.; Dewhurst, R.J.; Sykes, A.J.; Alexander, P. Modelling Alternative Management Scenarios of Economic and Environmental Sustainability of Beef Fi Nishing Systems. J. Clean. Prod. 2020, 253, 119888. [Google Scholar] [CrossRef]
- Fullman, T.J.; Bunting, E.L.; Kiker, G.A.; Southworth, J. Predicting Shifts in Large Herbivore Distributions under Climate Change and Management Using a Spatially-Explicit Ecosystem Model. Ecol. Modell. 2017, 352, 1–18. [Google Scholar] [CrossRef]
- Kalaugher, E.; Beukes, P.; Bornman, J.F.; Clark, A.; Campbell, D.I. Modelling Farm-Level Adaptation of Temperate, Pasture-Based Dairy Farms to Climate Change. Agric. Syst. 2017, 153, 53–68. [Google Scholar] [CrossRef]
- Moore, A.D.; Ghahramani, A. Climate Change and Broadacre Livestock Production across Southern Australia. 3. Adaptation Options via Livestock Genetic Improvement. Anim. Prod. Sci. 2014, 54, 111–124. [Google Scholar] [CrossRef]
- Snow, V.O.; Rotz, C.A.; Moore, A.D.; Martin-Clouaire, R.; Johnson, I.R.; Hutchings, N.J.; Eckard, R.J. The Challenges–and Some Solutions–to Process-Based Modelling of Grazed Agricultural Systems. Environ. Model. Softw. 2014, 62, 420–436. [Google Scholar] [CrossRef]
- Ma, L.; Derner, J.D.; Harmel, R.D.; Tatarko, J.; Moore, A.D.; Rotz, C.A.; Augustine, D.J.; Boone, R.B.; Coughenour, M.B.; Beukes, P.C.; et al. Application of Grazing Land Models in Ecosystem Management: Current Status and next Frontiers. Adv. Agron. 2019, 158, 173–215. [Google Scholar] [CrossRef]
- Ferrarini, A.; Alatalo, J.M.; Gervasoni, D.; Foggi, B. Exploring the Compass of Potential Changes Induced by Climate Warming in Plant Communities. Ecol. Complex. 2017, 29, 1–9. [Google Scholar] [CrossRef]
- Frate, L.; Carranza, M.L.; Evangelista, A.; Stinca, A.; Schaminée, J.H.J.; Stanisci, A. Climate and Land Use Change Impacts on Mediterranean High-Mountain Vegetation in the Apennines since the 1950s. Plant Ecol. Divers. 2018, 11, 85–96. [Google Scholar] [CrossRef]
- Dibari, C.; Argenti, G.; Catolfi, F.; Moriondo, M.; Staglianò, N.; Bindi, M. Pastoral Suitability Driven by Future Climate Change along the Apennines. Ital. J. Agron. 2015, 10, 109–116. [Google Scholar] [CrossRef]
- Vigan, A.; Lasseur, J.; Benoit, M.; Mouillot, F.; Eugène, M.; Mansard, L.; Vigne, M.; Lecomte, P.; Dutilly, C. Evaluating Livestock Mobility as a Strategy for Climate Change Mitigation: Combining Models to Address the Specificities of Pastoral Systems. Agric. Ecosyst. Environ. 2017, 242, 89–101. [Google Scholar] [CrossRef]
- Insua, J.R.; Utsumi, S.A.; Basso, B. Estimation of Spatial and Temporal Variability of Pasture Growth and Digestibility in Grazing Rotations Coupling Unmanned Aerial Vehicle (UAV) with Crop Simulation Models. PLoS ONE 2019, 14, e0212773. [Google Scholar] [CrossRef]
- Gao, Q.Z.; Li, Y.; Xu, H.M.; Wan, Y.F.; Jiangcun, W. zha Adaptation Strategies of Climate Variability Impacts on Alpine Grassland Ecosystems in Tibetan Plateau. Mitig. Adapt. Strateg. Glob. Chang. 2014, 19, 199–209. [Google Scholar] [CrossRef]
- Riedo, M.; Grub, A.; Rosset, M. A Pasture Simulation Model for Dry Matter Production, and Fluxes of Carbon, Nitrogen, Water and Energy. Ecol. Model. 1998, 105, 141–183. [Google Scholar] [CrossRef]
- Wilcke, R.A.I.; Bärring, L. Selecting Regional Climate Scenarios for Impact Modelling Studies. Environ. Model. Softw. 2016, 78, 191–201. [Google Scholar] [CrossRef]
- Pierce, D.W.; Barnett, T.P.; Santer, B.D.; Gleckler, P.J. Selecting Global Climate Models for Regional Climate Change Studies. Proc. Natl. Acad. Sci. USA 2009, 106. [Google Scholar] [CrossRef]
- Ruti, P.M.; Somot, S.; Giorgi, F.; Dubois, C.; Flaounas, E.; Obermann, A.; Dell’Aquila, A.; Pisacane, G.; Harzallah, A.; Lombardi, E.; et al. Med-CORDEX Initiative for Mediterranean Climate Studies. Bull. Am. Meteorol. Soc. 2016, 97, 1187–1208. [Google Scholar] [CrossRef]
- Cornes, R.C.; van der Schrier, G.; van den Besselaar, E.J.M.; Jones, P.D. An Ensemble Version of the E-OBS Temperature and Precipitation Data Sets. J. Geophys. Res. Atmos. 2018, 123, 9391–9409. [Google Scholar] [CrossRef]
- Lange, S. Trend-Preserving Bias Adjustment and Statistical Downscaling with ISIMIP3BASD (v1.0). Geosci. Model Dev. 2019, 12, 3055–3070. [Google Scholar] [CrossRef]
- Bristow, K.L.; Campbell, G.S. On the Relationship between Incoming Solar Radiation and Daily Maximum and Minimum Temperature. Agric. For. Meteorol. 1984, 31, 159–166. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. FAO Irrigation and Drainage Paper No. 56 - Crop Evapotranspiration. Rome Food Agric. Organ. United Nations 1998, 56, e156. [Google Scholar]
- IPCC Annex III: Tables of Historical and Projected Well-Mixed Greenhouse Gas Mixing Ratios and Effective Radiative Forcing of All Climate Forcers. In Climate Change 2021: The Physical Science Basis. Contribution of Wor; Dentener, F.J., Hall, B., Smith, C., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 2139–2152. [Google Scholar]
- Graux, A.I.; Bellocchi, G.; Lardy, R.; Soussana, J.F. Ensemble Modelling of Climate Change Risks and Opportunities for Managed Grasslands in France. Agric. For. Meteorol. 2013, 170, 114–131. [Google Scholar] [CrossRef]
- Touhami, H.B.; Lardy, R.; Barra, V.; Bellocchi, G. Screening Parameters in the Pasture Simulation Model Using the Morris Method. Ecol. Modell. 2013, 266, 42–57. [Google Scholar] [CrossRef]
- Ma, S.; Lardy, R.; Graux, A.I.; Ben Touhami, H.; Klumpp, K.; Martin, R.; Bellocchi, G. Regional-Scale Analysis of Carbon and Water Cycles on Managed Grassland Systems. Environ. Model. Softw. 2015, 72, 356–371. [Google Scholar] [CrossRef]
- Pulina, A.; Lai, R.; Salis, L.; Seddaiu, G.; Roggero, P.P.; Bellocchi, G. Modelling Pasture Production and Soil Temperature, Water and Carbon Fluxes in Mediterranean Grassland Systems with the Pasture Simulation Model. Grass Forage Sci. 2018, 73, 272–283. [Google Scholar] [CrossRef]
- Sándor, R.; Picon-Cochard, C.; Martin, R.; Louault, F.; Klumpp, K.; Borras, D.; Bellocchi, G. Plant Acclimation to Temperature: Developments in the Pasture Simulation Model. Field Crop. Res. 2018, 222, 238–255. [Google Scholar] [CrossRef]
- Ehrhardt, F.; Soussana, J.F.; Bellocchi, G.; Grace, P.; McAuliffe, R.; Recous, S.; Sándor, R.; Smith, P.; Snow, V.; de Antoni Migliorati, M.; et al. Assessing Uncertainties in Crop and Pasture Ensemble Model Simulations of Productivity and N2O Emissions. Glob. Chang. Biol. 2018, 24, e603–e616. [Google Scholar] [CrossRef]
- Bojanowski, J.S.; Donatelli, M.; Skidmore, A.K.; Vrieling, A. An Auto-Calibration Procedure for Empirical Solar Radiation Models. Environ. Model. Softw. 2013, 49, 118–128. [Google Scholar] [CrossRef]
- Poggio, L.; De Sousa, L.M.; Batjes, N.H.; Heuvelink, G.B.M.; Kempen, B.; Ribeiro, E.; Rossiter, D. SoilGrids 2.0: Producing Soil Information for the Globe with Quantified Spatial Uncertainty. Soil 2021, 7, 217–240. [Google Scholar] [CrossRef]
- Richter, K.; Atzberger, C.; Hank, T.B.; Mauser, W. Derivation of Biophysical Variables from Earth Observation Data: Validation and Statistical Measures. J. Appl. Remote Sens. 2012, 6, 063557-1. [Google Scholar] [CrossRef]
- Meinshausen, M.; Smith, S.J.; Calvin, K.; Daniel, J.S.; Kainuma, M.L.T.; Lamarque, J.; Matsumoto, K.; Montzka, S.A.; Raper, S.C.B.; Riahi, K.; et al. The RCP Greenhouse Gas Concentrations and Their Extensions from 1765 to 2300. Clim. Chang. 2011, 109, 213–241. [Google Scholar] [CrossRef]
- Movedi, E.; Bellocchi, G.; Argenti, G.; Paleari, L.; Vesely, F.; Staglianò, N.; Dibari, C.; Confalonieri, R. Development of Generic Crop Models for Simulation of Multi-Species Plant Communities in Mown Grasslands. Ecol. Modell. 2019, 401, 111–128. [Google Scholar] [CrossRef]
- Sándor, R.; Barcza, Z.; Acutis, M.; Doro, L.; Hidy, D.; Köchy, M.; Minet, J.; Lellei-Kovács, E.; Ma, S.; Perego, A.; et al. Multi-Model Simulation of Soil Temperature, Soil Water Content and Biomass in Euro-Mediterranean Grasslands: Uncertainties and Ensemble Performance. Eur. J. Agron. 2017, 88, 22–40. [Google Scholar] [CrossRef]
- Schucknecht, A.; Seo, B.; Krämer, A.; Asam, S.; Atzberger, C.; Kiese, R. Estimating Dry Biomass and Plant Nitrogen Concentration in Pre-Alpine Grasslands with Low-Cost UAS-Borne Multispectral Data-a Comparison of Sensors, Algorithms, and Predictor Sets. Biogeosciences 2022, 19, 2699–2727. [Google Scholar] [CrossRef]
- Kollas, C.; Kersebaum, K.C.; Nendel, C.; Manevski, K.; Müller, C.; Palosuo, T.; Armas-Herrera, C.M.; Beaudoin, N.; Bindi, M.; Charfeddine, M.; et al. Crop Rotation Modelling-A European Model Intercomparison. Eur. J. Agron. 2015, 70, 98–111. [Google Scholar] [CrossRef]
- Vuichard, N.; Soussana, J.F.; Ciais, P.; Viovy, N.; Ammann, C.; Calanca, P.; Clifton-Brown, J.; Fuhrer, J.; Jones, M.; Martin, C. Estimating the Greenhouse Gas Fluxes of European Grasslands with a Process-Based Model: 1. Model Evaluation from in Situ Measurements. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Schippers, P.; Van Groenendael, J.M.; Vleeshouwers, L.M.; Hunt, R. Herbaceous Plant Strategies in Disturbed Habitats. Oikos 2001, 95, 198–210. [Google Scholar] [CrossRef]
- Fuchs, K.; Merbold, L.; Buchmann, N.; Bellocchi, G.; Bindi, M.; Brilli, L.; Conant, R.T.; Dorich, C.D.; Ehrhardt, F.; Fitton, N.; et al. Evaluating the Potential of Legumes to Mitigate N2O Emissions From Permanent Grassland Using Process-Based Models. Glob. Biogeochem. Cycles 2020, 34, e2020GB006561. [Google Scholar] [CrossRef]
- Vital, J.A.; Gaurut, M.; Lardy, R.; Viovy, N.; Soussana, J.F.; Bellocchi, G.; Martin, R. High-Performance Computing for Climate Change Impact Studies with the Pasture Simulation Model. Comput. Electron. Agric. 2013, 98, 131–135. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, Y.; Zhang, Y.; Shang, J. Review of Remote Sensing Applications in Grassland Monitoring. Remote Sens. 2022, 14, 2903. [Google Scholar] [CrossRef]
- Bellini, E.; Moriondo, M.; Dibari, C.; Leolini, L.; Staglianò, N.; Stendardi, L.; Filippa, G.; Galvagno, M.; Argenti, G. Impacts of Climate Change on European Grassland Phenology: A 20-Year Analysis of MODIS Satellite Data. Remote Sens. 2023, 15, 218. [Google Scholar] [CrossRef]
- Ren, S.; Vitasse, Y.; Chen, X.; Peichl, M.; An, S. Assessing the Relative Importance of Sunshine, Temperature, Precipitation, and Spring Phenology in Regulating Leaf Senescence Timing of Herbaceous Species in China. Agric. For. Meteorol. 2022, 313, 108770. [Google Scholar] [CrossRef]
- Ellsworth, D.S.; Reich, P.B.; Naumburg, E.S.; Koch, G.W.; Kubiske, M.E.; Smith, S.D. Photosynthesis, Carboxylation and Leaf Nitrogen Responses of 16 Species to Elevated PCO2 across Four Free-Air CO2 Enrichment Experiments in Forest, Grassland and Desert. Glob. Chang. Biol. 2004, 10, 2121–2138. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What Have We Learned from 15 Years of Free-Air CO2 Enrichment (FACE)? A Meta-Analytic Review of the Responses of Photosynthesis, Canopy Properties and Plant Production to Rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Rogers, A. The Response of Photosynthesis and Stomatal Conductance to Rising [CO2]: Mechanisms and Environmental Interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Shrestha, S.; Abdalla, M.; Hennessy, T.; Forristal, D.; Jones, M.B. Irish Farms under Climate Change - Is There a Regional Variation on Farm Responses? J. Agric. Sci. 2015, 153, 385–398. [Google Scholar] [CrossRef]
- Zarrineh, N.; Abbaspour, K.C.; Holzkämper, A. Integrated Assessment of Climate Change Impacts on Multiple Ecosystem Services in Western Switzerland. Sci. Total Environ. 2020, 708, 135212. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Ciais, P.; Gasser, T.; Smith, P.; Herrero, M.; Havlík, P.; Obersteiner, M.; Guenet, B.; Goll, D.S.; Li, W.; et al. Climate Warming from Managed Grasslands Cancels the Cooling Effect of Carbon Sinks in Sparsely Grazed and Natural Grasslands. Nat. Commun. 2021, 12, 118. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Purdy, K.J.; Eyice, Ö.; Shen, L.; Harpenslager, S.F.; Yvon-Durocher, G.; Dumbrell, A.J.; Trimmer, M. Disproportionate Increase in Freshwater Methane Emissions Induced by Experimental Warming. Nat. Clim. Chang. 2020, 10, 685–690. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous Oxide Emissions from Soils: How Well Do We Understand the Processes and Their Controls? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Cullen, B.R.; Johnson, I.R.; Eckard, R.J.; Lodge, G.M.; Walker, R.J.; Rawnsley, R.P.; McCaskillF, M.R. Climate Change Effects on Pasture Systems in South-Eastern Australia. Crop Pasture Sci. 2009, 60, 933–942. [Google Scholar] [CrossRef]
- Parton, W.J.; Ojima, D.S.; Cole, C.V.; Schimel, D.S. A General Model for Soil Organic Matter Dynamics: Sensitivity to Litter Chemistry, Texture and Management. Quant. Model. Soil Form. Process. Proc. Symp. Minneapolis 1994, 39, 147–167. [Google Scholar] [CrossRef]
- Murphy, K.L.; Burke, I.C.; Vinton, M.A.; Lauenroth, W.K.; Aguiar, M.R.; Wedin, D.A.; Virginia, R.A.; Lowe, P.N. Regional Analysis of Litter Quality in the Central Grassland Region of North America. J. Veg. Sci. 2002, 13, 395–402. [Google Scholar] [CrossRef]
- Ouled Belgacem, A.; Louhaichi, M. The Vulnerability of Native Rangeland Plant Species to Global Climate Change in the West Asia and North African Regions. Clim. Chang. 2013, 119, 451–463. [Google Scholar] [CrossRef]
- He, P.; Ma, X.; Sun, Z.; Han, Z.; Ma, S.; Xiaoyu, M. Compound Drought Constrains Gross Primary Productivity in Chinese Grasslands. Environ. Res. Lett. 2022, 17, 104054. [Google Scholar] [CrossRef]
- Rounsevell, M.D.A.; Brignall, A.P.; Siddons, P.A. Potential Climate Change Effects on the Distribution of Agricultural Grassland in England and Wales. Soil Use Manag. 1996, 12, 44–51. [Google Scholar] [CrossRef]
- Edmonds, J.A.; Rosenberg, N.J. Climate Change Impacts for the Conterminous USA: An Integrated Assessment Summary. Clim. Chang. Impacts Conterminous USA Integr. Assess. 2005, 151–162. [Google Scholar] [CrossRef]
- Morales, P.; Hickler, T.; Rowell, D.P.; Smith, B.; Sykes, M.T. Changes in European Ecosystem Productivity and Carbon Balance Driven by Regional Climate Model Output. Glob. Chang. Biol. 2007, 13, 108–122. [Google Scholar] [CrossRef]
- Harrison, M.T.; Cullen, B.R.; Armstrong, D. Management Options for Dairy Farms under Climate Change: Effects of Intensification, Adaptation and Simplification on Pastures, Milk Production and Profitability. Agric. Syst 2017, 155, 19–32. [Google Scholar] [CrossRef]
- Joyce, L.A.; Briske, D.D.; Brown, J.R.; Polley, H.W.; McCarl, B.A.; Bailey, D.W. Climate Change and North American Rangelands: Assessment of Mitigation and Adaptation Strategies. Rangel. Ecol. Manag. 2013, 66, 512–528. [Google Scholar] [CrossRef]
- Cheng, M.; Mccarl, B.; Fei, C. Climate Change and Livestock Production: A Literature Review. Atmosphere 2022, 13, 140. [Google Scholar] [CrossRef]
- Briske, D.D.; Joyce, L.A.; Polley, H.W.; Brown, J.R.; Wolter, K.; Morgan, J.A.; McCarl, B.A.; Bailey, D.W. Climate-Change Adaptation on Rangelands: Linking Regional Exposure with Diverse Adaptive Capacity. Front. Ecol. Environ. 2015, 13, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Degaetano, A.T.; Rotz, C.A.; Hoberg, E.; Skinner, R.H.; Felix, T.; Li, H.; Patterson, P.H.; Roth, G.; Hall, M.; et al. Climate Change Effects on Livestock in the Northeast US and Strategies for Adaptation. Clim. Chang. 2018, 146, 33–45. [Google Scholar] [CrossRef]
- Xu, C.; Liu, H.; Williams, A.P.; Yin, Y.; Wu, X. Trends toward an Earlier Peak of the Growing Season in Northern Hemisphere Mid-Latitudes. Glob. Chang. Biol. 2016, 22, 2852–2860. [Google Scholar] [CrossRef] [PubMed]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; McIvor, J.; Milne, J.; Morris, C.; Peeters, A.; et al. An International Terminology for Grazing Lands and Grazing Animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Li, W.; Cao, W.; Wang, J.; Li, X.; Xu, C.; Shi, S. Effects of Grazing Regime on Vegetation Structure, Productivity, Soil Quality, Carbon and Nitrogen Storage of Alpine Meadow on the Qinghai-Tibetan Plateau. Ecol. Eng. 2017, 98, 123–133. [Google Scholar] [CrossRef]
- Liu, J.; Isbell, F.; Ma, Q.; Chen, Y.; Xing, F.; Sun, W.; Wang, L.; Li, J.; Wang, Y.; Hou, F.; et al. Overgrazing, Not Haying, Decreases Grassland Topsoil Organic Carbon by Decreasing Plant Species Richness along an Aridity Gradient in Northern China. Agric. Ecosyst. Environ. 2022, 332, 107935. [Google Scholar] [CrossRef]
- Porro, F.; Tomaselli, M.; Abeli, T.; Gandini, M.; Gualmini, M.; Orsenigo, S.; Petraglia, A.; Rossi, G.; Carbognani, M. Could Plant Diversity Metrics Explain Climate-Driven Vegetation Changes on Mountain Summits of the GLORIA Network? Biodivers. Conserv. 2019, 28, 3575–3596. [Google Scholar] [CrossRef]
- Löffler, J.; Pape, R. Thermal Niche Predictors of Alpine Plant Species. Ecology 2020, 101, e02891. [Google Scholar] [CrossRef]
- Piseddu, F.; Bellocchi, G.; Picon-Cochard, C. Mowing and Warming Effects on Grassland Species Richness and Harvested Biomass: Meta-Analyses. Agron. Sustain. Dev. 2021, 41, 74. [Google Scholar] [CrossRef]
- Lamprecht, A.; Semenchuk, P.R.; Steinbauer, K.; Winkler, M.; Pauli, H. Climate Change Leads to Accelerated Transformation of High-Elevation Vegetation in the Central Alps. New Phytol. 2018, 220, 447–459. [Google Scholar] [CrossRef] [PubMed]









| Description | Unit | Site M (Marradi) | Site B (Borgo San Lorenzo) |
|---|---|---|---|
| Location | |||
| Latitude (WGS84) | degree N | 44.08° | 43.95° |
| Longitude (WGS84) | degree E | 11.63° | 11.35° |
| Elevation | m a.s.l. | 600 | 200 |
| Climate | |||
| Mean annual temperature 1 | °C | 12.4 | 13.4 |
| Mean annual precipitation 2 | mm | 1330 | 990 |
| Soil 3 | |||
| Depth | m | 1 | 1 |
| Clay | % | 37 | 37 |
| Silt | % | 42 | 36 |
| Sand | % | 21 | 27 |
| Total organic carbon | g kg−1 | 33.6 | 23.5 |
| Total nitrogen | g kg−1 | 3.0 | 2.5 |
| Soil pH | - | 6.6 | 7.4 |
| Bulk density | g cm−3 | 1.29 | 1.44 |
| Saturated soil water content | m3 m−3 | 0.52 | 0.51 |
| Field capacity | m3 m−3 | 0.36 | 0.35 |
| Wilting point | m3 m−3 | 0.21 | 0.21 |
| Dominant vegetation | - | Dactylis glomerata, Lolium sp., Festuca arundinacea, Phleum pratense, and Onobrychis viciifolia, with other minor forbs and a large presence in some sectors of shrubs, such as Rubus ulmifolius. | Lolium sp., Dactylis glomerata, Trifolium pratense, Trifolium repens, Lotus corniculatus, and Festuca arundinacea, with other minor forbs. |
| Management | Unit | Site M (Marradi) | Site B (Borgo San Lorenzo) | ||||
|---|---|---|---|---|---|---|---|
| B1 | B2 | ||||||
| 2020 | 2021 | 2020 | 2021 | 2020 | 2021 | ||
| Surface | ha | 5.4 | 10 | 20 | 10 | ||
| Cut | day of year | - | - | - | 125 | - | |
| Grazing period | days of year (start, end) | 139–244 a; 244–267 b | 135–176 a; 176–276 b | 100–180 a; 186–300 b | 100–145 a; 145–306 b | 180–186 a; 186–300 b | 110–145 a; 145–306 b |
| Stocking rate | LSU ha−1 d−1 | 4.0 a; 3.4 b | 3.3 a; 2.0 b | 2.9 a; 1.0 b | 1.5 a; 1.0 b | 0.9; 1.2 b | |
| Output | Site M | Site B | ||||
|---|---|---|---|---|---|---|
| R2 | RRMSE | B1 | B2 | |||
| R2 | RRMSE | R2 | RRMSE | |||
| AGB | 0.76 | 14.9 | 0.66 | 13.5 | 0.68 | 10.0 |
| LAI | 0.96 | 9.6 | 0.47 | 24.5 | 0.71 | 12.6 |
| Site | NEE | CH4 | N2O |
|---|---|---|---|
| g C m−2 yr−1 | g N m−2 yr−1 | ||
| Site M | 381.3 ± 245.6 | 2.2 ± 0.3 | 4.6 ± 3.4 |
| Site B | 350.1 ± 236.1 | 1.8 ± 0.2 | 4.7 ± 3.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellini, E.; Martin, R.; Argenti, G.; Staglianò, N.; Costafreda-Aumedes, S.; Dibari, C.; Moriondo, M.; Bellocchi, G. Opportunities for Adaptation to Climate Change of Extensively Grazed Pastures in the Central Apennines (Italy). Land 2023, 12, 351. https://doi.org/10.3390/land12020351
Bellini E, Martin R, Argenti G, Staglianò N, Costafreda-Aumedes S, Dibari C, Moriondo M, Bellocchi G. Opportunities for Adaptation to Climate Change of Extensively Grazed Pastures in the Central Apennines (Italy). Land. 2023; 12(2):351. https://doi.org/10.3390/land12020351
Chicago/Turabian StyleBellini, Edoardo, Raphaël Martin, Giovanni Argenti, Nicolina Staglianò, Sergi Costafreda-Aumedes, Camilla Dibari, Marco Moriondo, and Gianni Bellocchi. 2023. "Opportunities for Adaptation to Climate Change of Extensively Grazed Pastures in the Central Apennines (Italy)" Land 12, no. 2: 351. https://doi.org/10.3390/land12020351
APA StyleBellini, E., Martin, R., Argenti, G., Staglianò, N., Costafreda-Aumedes, S., Dibari, C., Moriondo, M., & Bellocchi, G. (2023). Opportunities for Adaptation to Climate Change of Extensively Grazed Pastures in the Central Apennines (Italy). Land, 12(2), 351. https://doi.org/10.3390/land12020351