

Short- and Mid-Term Spatiotemporal Diversity Patterns of Post-Fire Insect-Pollinated Plant Communities in the Mediterranean



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Sites and Field Sampling
2.3. Beta Diversity Patterns and Statistical Analysis
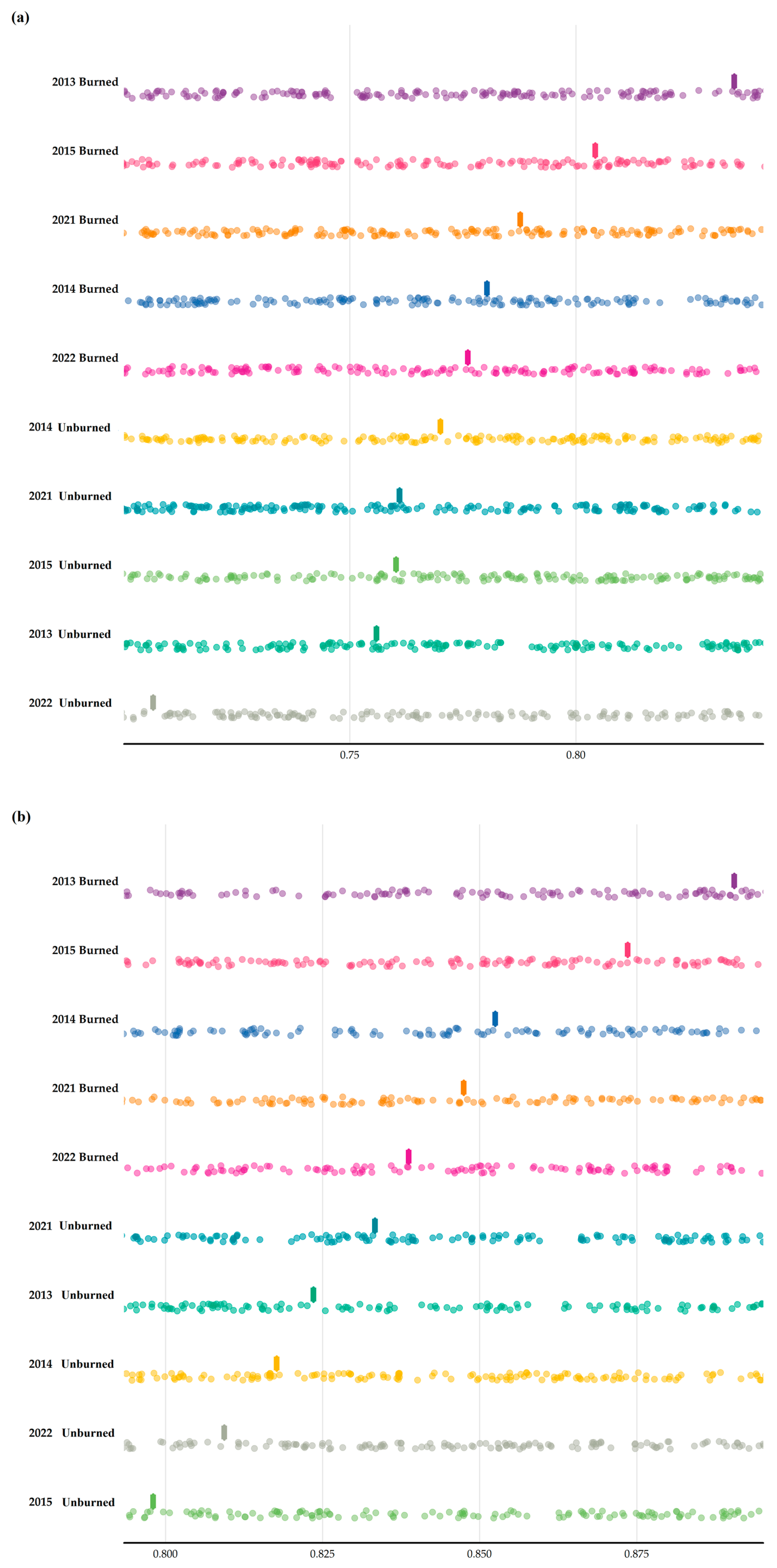
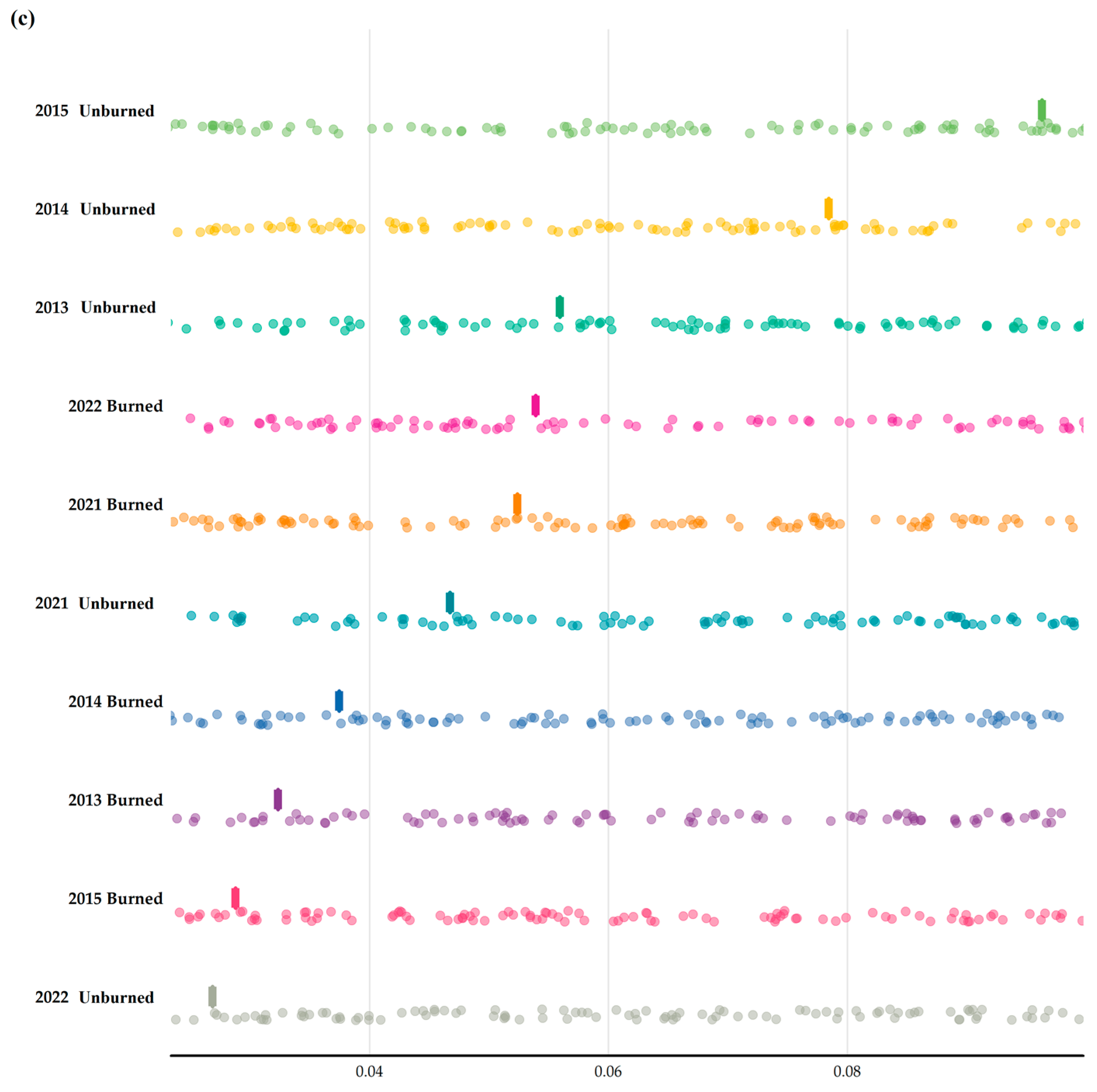
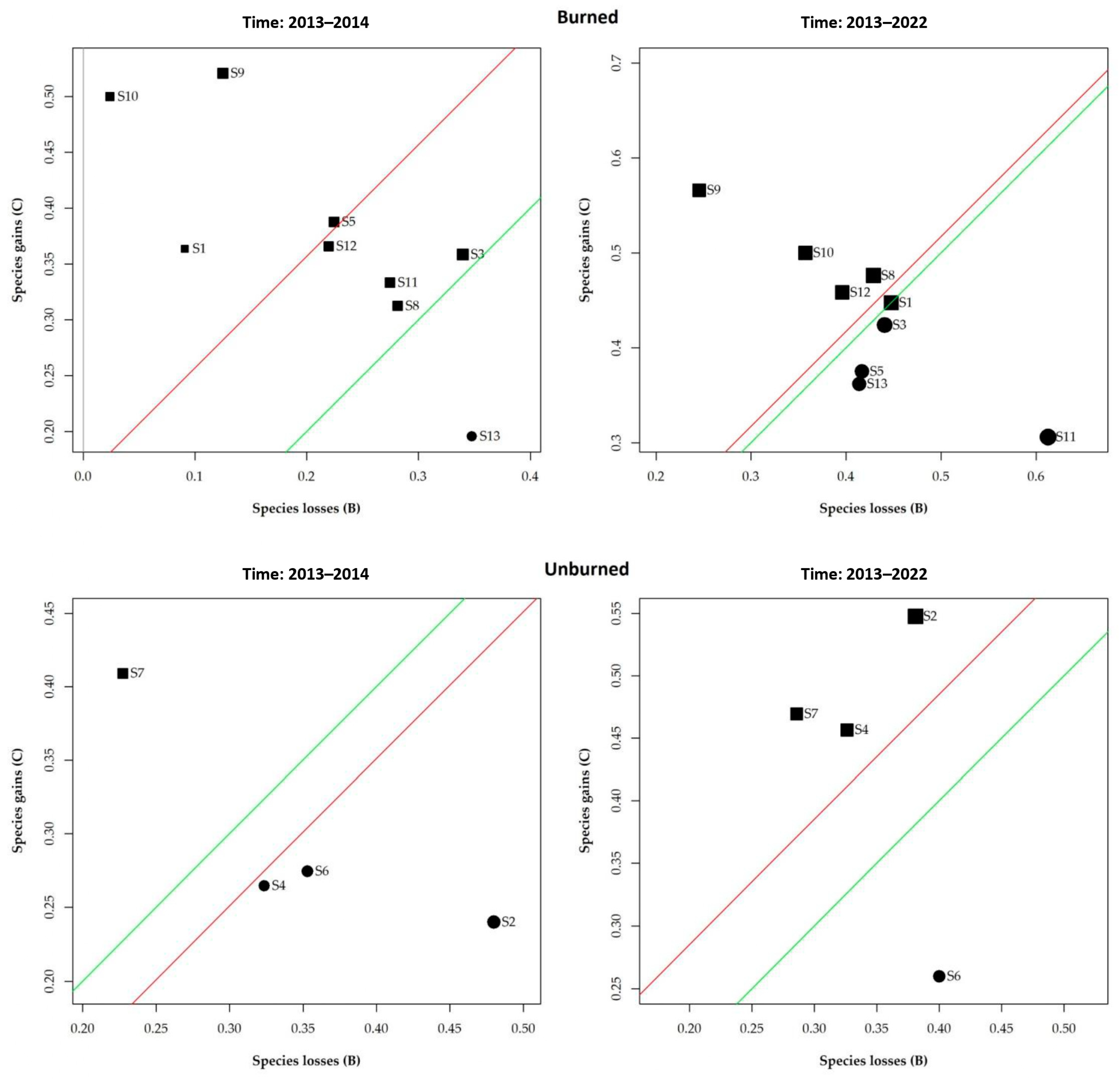
3. Results
3.1. Floristic Composition
3.2. Multiple Site Beta Diversity
3.3. Temporal Beta Diversity Index
4. Discussion
4.1. Beta Diversity Patterns
4.2. Temporal Beta Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the earth system. Science 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Keeley, J.E.; Bond, W.J.; Bradstock, R.A.; Pausas, J.G.; Rundel, P.W. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management; Cambridge University Press: New York, NY, USA, 2012. [Google Scholar]
- Chuvieco, E.; Giglio, L.; Justice, C. Global characterization of fire activity: Toward defining fire regimes from Earth observation data. Glob. Chang. Biol. 2008, 14, 1488–1502. [Google Scholar] [CrossRef]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Blondel, J.; Aronson, J.; Bodiou, J.-Y.; Boeuf, G. The Mediterranean Region: Biological Diversity in Space and Time, 2nd ed.; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Pausas, J.G.; Keeley, J.E. A burning story: The role of fire in the history of life. BioScience 2009, 59, 593–601. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Pyke, G.H. Fire-Stimulated Flowering: A Review and Look to the Future. Crit. Rev. Plant Sci. 2017, 36, 179–189. [Google Scholar] [CrossRef]
- Ne’eman, G.; Lev-Yadun, S.; Arianoutsou, M. Fire-Related Traits in Mediterranean Basin Plants. Isr. J. Ecol. Evol. 2012, 58, 177–194. [Google Scholar] [CrossRef]
- He, T.; Lamont, B.B.; Pausas, J.G. Fire as a key driver of Earth’s biodiversity. Biol. Rev. 2019, 94, 1983–2010. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Kavgacı, A.; Čarni, A.; Başaran, S.; Başaran, M.A.; Košir, P.; Marinšek, A.; Šilc, U. Long-term post-fire succession of Pinus brutia forest in the east Mediterranean. Int. J. Wildland Fire 2010, 19, 599–605. [Google Scholar] [CrossRef]
- Christopoulou, A.; Fyllas, N.M.; Andriopoulos, P.; Koutsias, N.; Dimitrakopoulos, P.G.; Arianoutsou, M. Post-fire regeneration patterns of Pinus nigra in a recently burned area in Mount Taygetos, Southern Greece: The role of unburned forest patches. For. Ecol. Manag. 2014, 327, 148–156. [Google Scholar] [CrossRef]
- Christopoulou, A.; Kazanis, D.; Fyllas, N.; Arianoutsou, M. Post-fire recovery of Abies cephalonica forest communities: The case of Mt Parnitha National Park, Attica, Greece. iForest 2018, 11, 757–764. [Google Scholar] [CrossRef]
- Pausas, J.G.; Llovet, J.; Rodrigo, A.; Vallejo, R. Are wildfires a disaster in the Mediterranean basin? A review. Int. J. Wildland Fire 2008, 17, 713–723. [Google Scholar] [CrossRef]
- Arianoutsou-Faraggitaki, M. Post-fire successional recovery of a phryganic (East Mediterranean) ecosystem. Acta Oecologica 1984, 5, 387–394. [Google Scholar]
- Kazanis, D.; Arianoutsou, M. Long-term post-fire vegetation dynamics in Pinus halepensis forests of Central Greece: A functional group approach. Plant Ecol. 2004, 171, 101–121. [Google Scholar] [CrossRef]
- Hanes, T.L. Succession after Fire in the Chaparral of Southern California. Ecol. Monogr. 1971, 41, 27–52. [Google Scholar] [CrossRef]
- Trabaud, L.; Lepart, J. Diversity and stability in garrigue ecosystems after fire. Vegetatio 1980, 43, 49–57. [Google Scholar] [CrossRef]
- Strand, E.K.; Satterberg, K.L.; Hudak, A.T.; Byrne, J.; Khalyani, A.H.; Smith, A.M.S. Does burn severity affect plant community diversity and composition in mixed conifer forests of the United States Intermountain West one decade post fire? Fire Ecol. 2019, 15, 25. [Google Scholar] [CrossRef]
- Moradizadeh, H.; Heydari, M.; Omidipour, R.; Mezbani, A.; Prévosto, B. Ecological effects of fire severity and time since fire on the diversity partitioning, composition and niche apportionment models of post-fire understory vegetation in semi-arid oak forests of Western Iran. Ecol. Eng. 2020, 143, 105694. [Google Scholar] [CrossRef]
- Tsafrir, A.; Osem, Y.; Shemesh, H.; Carmel, Y.; Soref, C.; Ovadia, O. Fire season modifies the perennial plant community composition through a differential effect on obligate seeders in eastern Mediterranean woodlands. Appl. Veg. Sci. 2019, 22, 115–126. [Google Scholar] [CrossRef]
- Fernández-García, V.; Fulé, P.Z.; Marcos, E.; Calvo, L. The role of fire frequency and severity on the regeneration of Mediterranean serotinous pines under different environmental conditions. For. Ecol. Manag. 2019, 444, 59–68. [Google Scholar] [CrossRef]
- Tessler, N.; Sapir, Y.; Wittenberg, L.; Greenbaum, N. Recovery of Mediterranean Vegetation after Recurrent Forest Fires: Insight from the 2010 Forest Fire on Mount Carmel, Israel. Land Degrad. Dev. 2016, 27, 1424–1431. [Google Scholar] [CrossRef]
- Arnan, X.; Rodrigo, A.; Retana, J. Post-fire regeneration of Mediterranean plant communities at a regional scale is dependent on vegetation type and dryness. J. Veg. Sci. 2007, 18, 111–122. [Google Scholar] [CrossRef]
- Burkle, L.A.; Myers, J.A.; Belote, R.T. Wildfire disturbance and productivity as drivers of plant species diversity across spatial scales. Ecosphere 2015, 6, art202. [Google Scholar] [CrossRef]
- Weeks, J.; Miller, J.E.D.; Steel, Z.L.; Batzer, E.E.; Safford, H.D. High-severity fire drives persistent floristic homogenization in human-altered forests. Ecosphere 2023, 14, e4409. [Google Scholar] [CrossRef]
- Ponisio, L.C.; Wilkin, K.; M’Gonigle, L.K.; Kulhanek, K.; Cook, L.; Thorp, R.; Griswold, T.; Kremen, C. Pyrodiversity begets plant–pollinator community diversity. Glob. Chang. Biol. 2016, 22, 1794–1808. [Google Scholar] [CrossRef]
- Lazarina, M.; Devalez, J.; Neokosmidis, L.; Sgardelis, S.P.; Kallimanis, A.S.; Tscheulin, T.; Tsalkatis, P.; Kourtidou, M.; Mizerakis, V.; Nakas, G.; et al. Moderate fire severity is best for the diversity of most of the pollinator guilds in Mediterranean pine forests. Ecology 2019, 100, e02615. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Vulliamy, B.; Dafni, A.; Ne’eman, G.; O’Toole, C.; Roberts, S.; Willmer, P. Response of plant-pollinator communities to fire: Changes in diversity, abundance and floral reward structure. Oikos 2003, 101, 103–112. [Google Scholar] [CrossRef]
- Potts, S.G.; Vulliamy, B.; Dafni, A.; Ne’eman, G.; Willmer, P. Linking bees and flowers: How do floral communities structure pollinator communities? Ecology 2003, 84, 2628–2642. [Google Scholar] [CrossRef]
- LaManna, J.A.; Burkle, L.A.; Belote, R.T.; Myers, J.A. Biotic and abiotic drivers of plant–pollinator community assembly across wildfire gradients. J. Ecol. 2021, 109, 1000–1013. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Aizen, M.A.; Gemmill-Herren, B.; LeBuhn, G.; Minckley, R.; Packer, L.; Potts, S.G.; Roulston, T.A.; Steffan-Dewenter, I.; et al. Pollination and other ecosystem services produced by mobile organisms: A conceptual framework for the effects of land-use change. Ecol. Lett. 2007, 10, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Powney, G.D.; Carvell, C.; Edwards, M.; Morris, R.K.A.; Roy, H.E.; Woodcock, B.A.; Isaac, N.J.B. Widespread losses of pollinating insects in Britain. Nat. Commun. 2019, 10, 1018. [Google Scholar] [CrossRef]
- Han, J.; Shen, Z.; Li, Y.; Luo, C.; Xu, Q.; Yang, K.; Zhang, Z. Beta Diversity Patterns of Post-fire Forests in Central Yunnan Plateau, Southwest China: Disturbances Intensify the Priority Effect in the Community Assembly. Front. Plant Sci. 2018, 9, 1000. [Google Scholar] [CrossRef]
- Lazarina, M.; Sgardelis, S.P.; Tscheulin, T.; Devalez, J.; Mizerakis, V.; Kallimanis, A.S.; Papakonstantinou, S.; Kyriazis, T.; Petanidou, T. The effect of fire history in shaping diversity patterns of flower-visiting insects in post-fire Mediterranean pine forests. Biodivers. Conserv. 2017, 26, 115–131. [Google Scholar] [CrossRef]
- Meikle, R.D. A Survey of the Flora of Chios. Kew Bull. 1954, 9, 85–199. [Google Scholar] [CrossRef]
- Snogerup, S.; Snogerup, B.; Phitos, D.; Kamari, G. The flora of Chios island (Greece). Bot. Chron. 2001, 14, 5–197. [Google Scholar]
- Stefanaki, A.; Kokkini, S. Phytogeographical affinities at the crossroads of two continents: Distribution patterns of Lamiaceae in Chios Island (East Aegean Islands, Greece) and Çeşme–Karaburun Peninsula (West Anatolia, Turkey). Syst. Biodivers. 2015, 13, 307–325. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Valli, A.T.; Georgopoulou, E.; Simaiakis, S.M.; Triantis, K.A.; Trigas, P. Network biogeography of a complex island system: The Aegean Archipelago revisited. J. Biogeogr. 2017, 44, 651–660. [Google Scholar] [CrossRef]
- Strid, A. Phytogeographia Aegaea and the flora Hellenica database. Ann. Naturhistorischen Mus. Wien 1996, 98, 279–289. [Google Scholar]
- Hammoud, C.; Kougioumoutzis, K.; Rijsdijk, K.F.; Simaiakis, S.M.; Norder, S.J.; Foufopoulos, J.; Georgopoulou, E.; Van Loon, E.E. Past connections with the mainland structure patterns of insular species richness in a continental-shelf archipelago (Aegean Sea, Greece). Ecol. Evol. 2021, 11, 5441–5458. [Google Scholar] [CrossRef] [PubMed]
- Panitsa, M.; Kagiampaki, A.; Kougioumoutzis, K. Plant diversity and biogeography of the Aegean Archipelago: A New Synthesis. In Biogeography and Biodiversity of the Aegean. In Honour of Prof. Moysis Mylonas; Sfenthourakis, S., Pafilis, P., Parmakelis, A., Poulakakis, N., Triantis, K., Eds.; Broken Hill Publishers Ltd.: Nicosia, Cyprus, 2018; pp. 223–244. [Google Scholar]
- Nakas, G.; Kantsa, A.; Vujić, A.; Mescher, M.C.; De Moraes, C.Μ.; Petanidou, T. Recent fire in a Mediterranean ecosystem strengthens hoverfly populations and their interaction networks with plants. Ecol. Evol. 2023, 13, e9803. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A.; Bonthoux, S.; Balent, G. Temporal Beta Diversity of Bird Assemblages in Agricultural Landscapes: Land Cover Change vs. Stochastic Processes. PLoS ONE 2015, 10, e0127913. [Google Scholar] [CrossRef]
- Aizaki, H. Mded: Measuring the Difference Between Two Empirical Distributions. R Package Version 0.1-2; 2015. Available online: https://CRAN.R-project.org/package=mded (accessed on 24 November 2023).
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guénard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. Adespatial: Multivariate Multiscale Spatial Analysis. R Package Version 0.3-23. 2023. Available online: https://CRAN.R-project.org/package=adespatial (accessed on 24 November 2023).
- Legendre, P. A temporal beta-diversity index to identify sites that have changed in exceptional ways in space–time surveys. Ecol. Evol. 2019, 9, 3500–3514. [Google Scholar] [CrossRef]
- Petanidou, T.; Ellis, W.N. Interdependence of native bee faunas and floras in changing Mediterranean communities. In The Conservation of Bees—Linnean Society Symposium Series No 18; Matheson, A., Buchmann, S.L., O’Toole, C., Westrich, P., Williams, I.H., Eds.; Academic Press: London, UK, 1996; Volume 18, pp. 201–226. [Google Scholar]
- Ulyshen, M.D.; Hiers, J.K.; Pokswinksi, S.M.; Fair, C. Pyrodiversity promotes pollinator diversity in a fire-adapted landscape. Front. Ecol. Environ. 2022, 20, 78–83. [Google Scholar] [CrossRef]
- Freeman, J.P.; Stohlgren, T.J.; Hunter, M.E.; Omi, P.N.; Martinson, E.J.; Chong, G.W.; Brown, C.S. Rapid assessment of postfire plant invasions in coniferous forests of the western United States. Ecol. Appl. 2007, 17, 1656–1665. [Google Scholar] [CrossRef] [PubMed]
- St. Clair, S.B.; O’Connor, R.; Gill, R.; McMillan, B. Biotic resistance and disturbance: Rodent consumers regulate post-fire plant invasions and increase plant community diversity. Ecology 2016, 97, 1700–1711. [Google Scholar] [CrossRef] [PubMed]
- Capitanio, R.; Carcaillet, C. Post-fire Mediterranean vegetation dynamics and diversity: A discussion of succession models. For. Ecol. Manag. 2008, 255, 431–439. [Google Scholar] [CrossRef]
- Kazanis, D.; Arianoutsou, M. Vegetation Composition in a Post-Fire Successional Gradient of Pinus halepensis Forests in Attica, Greece. Int. J. Wildland Fire 1996, 6, 83–91. [Google Scholar] [CrossRef]
- Fründ, J.; Linsenmair, K.E.; Blüthgen, N. Pollinator diversity and specialization in relation to flower diversity. Oikos 2010, 119, 1581–1590. [Google Scholar] [CrossRef]
- Paula, S.; Arianoutsou, M.; Kazanis, D.; Tavsanoglu, Ç.; Lloret, F.; Buhk, C.; Ojeda, F.; Luna, B.; Moreno, J.M.; Rodrigo, A.; et al. Fire-related traits for plant species of the Mediterranean Basin. Ecology 2009, 90, 1420. [Google Scholar] [CrossRef]
- Bond, W.J.; Keeley, J.E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef]
- Nakas, G.; Lazarina, M.; Chtenelli, T.-T.; Sgardelis, S.; Dimitrakopoulos, P.G.; Devalez, J.; Petanidou, T. The response to fire by two eusocial bee species. Ecol. Entomol. 2023, 1–13. [Google Scholar] [CrossRef]
- Arianoutsou, M.; Ne’eman, G. Post-fire regeneration of natural Pinus halepensis forests in the East Mediterranean Basin. In Ecology Biogeography and Management of Pinus halepensis and Pinus brutia Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000. [Google Scholar]
- Burkle, L.A.; Belote, R.T.; Myers, J.A. Wildfire severity alters drivers of interaction beta-diversity in plant–bee networks. Ecography 2022, 2022, e05986. [Google Scholar] [CrossRef]
- Flannigan, M.; Cantin, A.S.; de Groot, W.J.; Wotton, M.; Newbery, A.; Gowman, L.M. Global wildland fire season severity in the 21st century. For. Ecol. Manag. 2013, 294, 54–61. [Google Scholar] [CrossRef]
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J.S. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat. Commun. 2015, 6, 7537. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # of Plant Species | |||||||
|---|---|---|---|---|---|---|---|
| Site | Fire | Total | 2013 | 2014 | 2015 | 2021 | 2022 |
| S1 | B | 45 | 21 | 30 | 33 | 23 | 21 |
| S3 | B | 75 | 34 | 35 | 37 | 36 | 33 |
| S5 | B | 66 | 30 | 39 | 39 | 24 | 28 |
| S8 | B | 45 | 22 | 23 | 23 | 21 | 24 |
| S9 | B | 72 | 23 | 42 | 30 | 40 | 40 |
| S10 | B | 56 | 21 | 41 | 29 | 28 | 27 |
| S11 | B | 59 | 34 | 37 | 26 | 28 | 19 |
| S12 | B | 60 | 26 | 32 | 28 | 21 | 29 |
| S13 | B | 71 | 37 | 30 | 29 | 37 | 34 |
| S2 | U | 29 | 19 | 13 | 16 | 19 | 26 |
| S4 | U | 49 | 25 | 23 | 22 | 29 | 31 |
| S6 | U | 68 | 37 | 33 | 42 | 28 | 30 |
| S7 | U | 65 | 26 | 34 | 42 | 36 | 35 |
| Species Name | Family | Life Cycle | Chorology | # of Sites |
|---|---|---|---|---|
| Anagallis arvensis | Primulaceae | Annual | Cosmopolitan | 13 |
| Centaurium tenuiflorum | Gentianaceae | Annual | Mediterranean-European | 13 |
| Cistus creticus | Cistaceae | Perennial | Mediterranean | 12 |
| Crepis commutata | Asteraceae | Perennial | E-Mediterranean | 12 |
| Helichrysum stoechas | Asteraceae | Perennial | Mediterranean | 13 |
| Hypochaeris achyrophorus | Asteraceae | Annual | Mediterranean | 12 |
| Leontodon tuberosus | Asteraceae | Perennial | Mediterranean | 12 |
| Teucrium divaricatum | Lamiaceae | Perennial | E-Mediterranean | 12 |
| Thymbra capitata | Lamiaceae | Perennial | Mediterranean | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakas, G.; Kougioumoutzis, K.; Petanidou, T. Short- and Mid-Term Spatiotemporal Diversity Patterns of Post-Fire Insect-Pollinated Plant Communities in the Mediterranean. Land 2023, 12, 2109. https://doi.org/10.3390/land12122109
Nakas G, Kougioumoutzis K, Petanidou T. Short- and Mid-Term Spatiotemporal Diversity Patterns of Post-Fire Insect-Pollinated Plant Communities in the Mediterranean. Land. 2023; 12(12):2109. https://doi.org/10.3390/land12122109
Chicago/Turabian StyleNakas, Georgios, Konstantinos Kougioumoutzis, and Theodora Petanidou. 2023. "Short- and Mid-Term Spatiotemporal Diversity Patterns of Post-Fire Insect-Pollinated Plant Communities in the Mediterranean" Land 12, no. 12: 2109. https://doi.org/10.3390/land12122109
APA StyleNakas, G., Kougioumoutzis, K., & Petanidou, T. (2023). Short- and Mid-Term Spatiotemporal Diversity Patterns of Post-Fire Insect-Pollinated Plant Communities in the Mediterranean. Land, 12(12), 2109. https://doi.org/10.3390/land12122109