
Remote-Sensed Tree Crown Diameter as a Predictor of Stem Diameter and Above-Ground Biomass in Betula pendula Roth and Populus tremuloides Michx. × Populus tremula L. Plantations

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
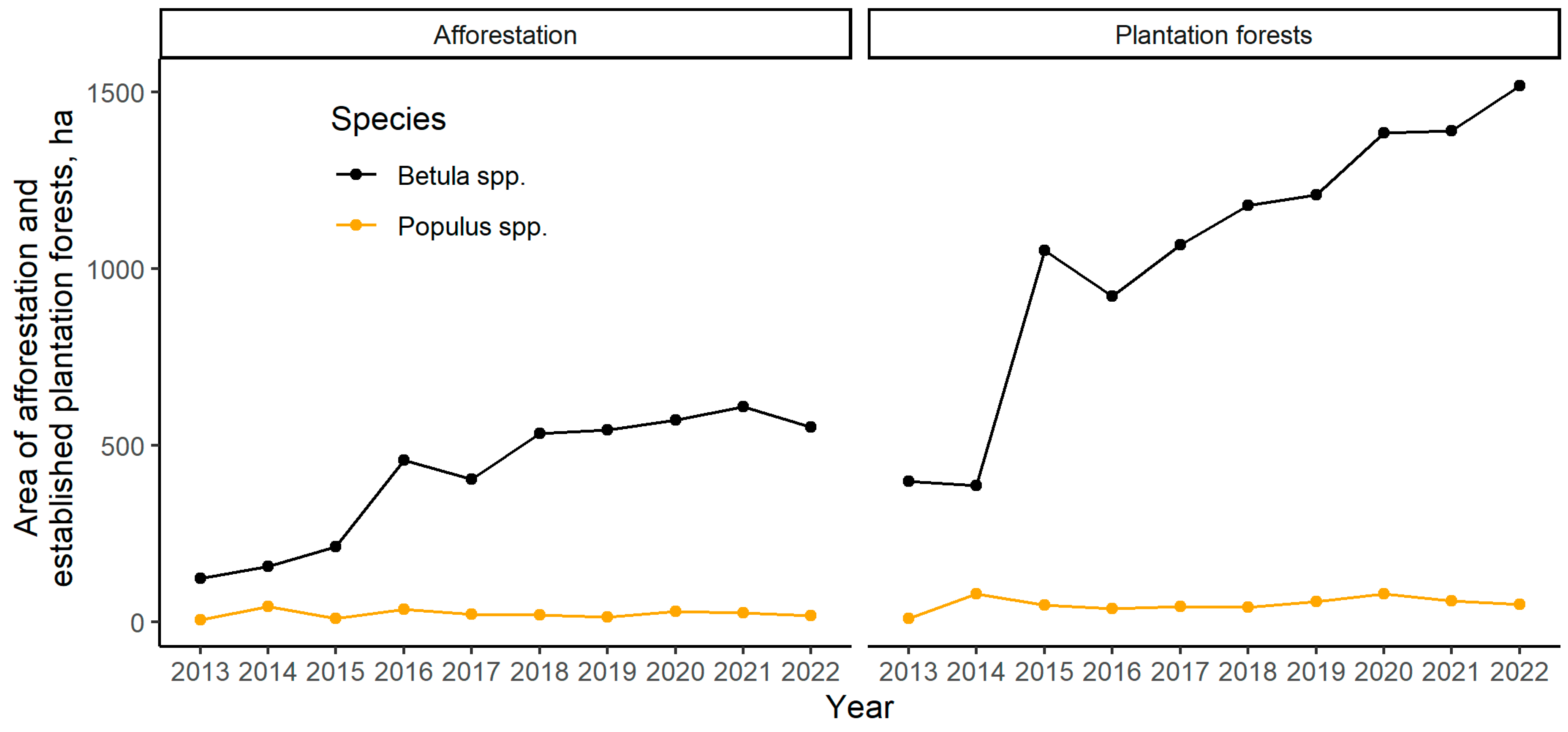
:1. Introduction
2. Materials and Methods
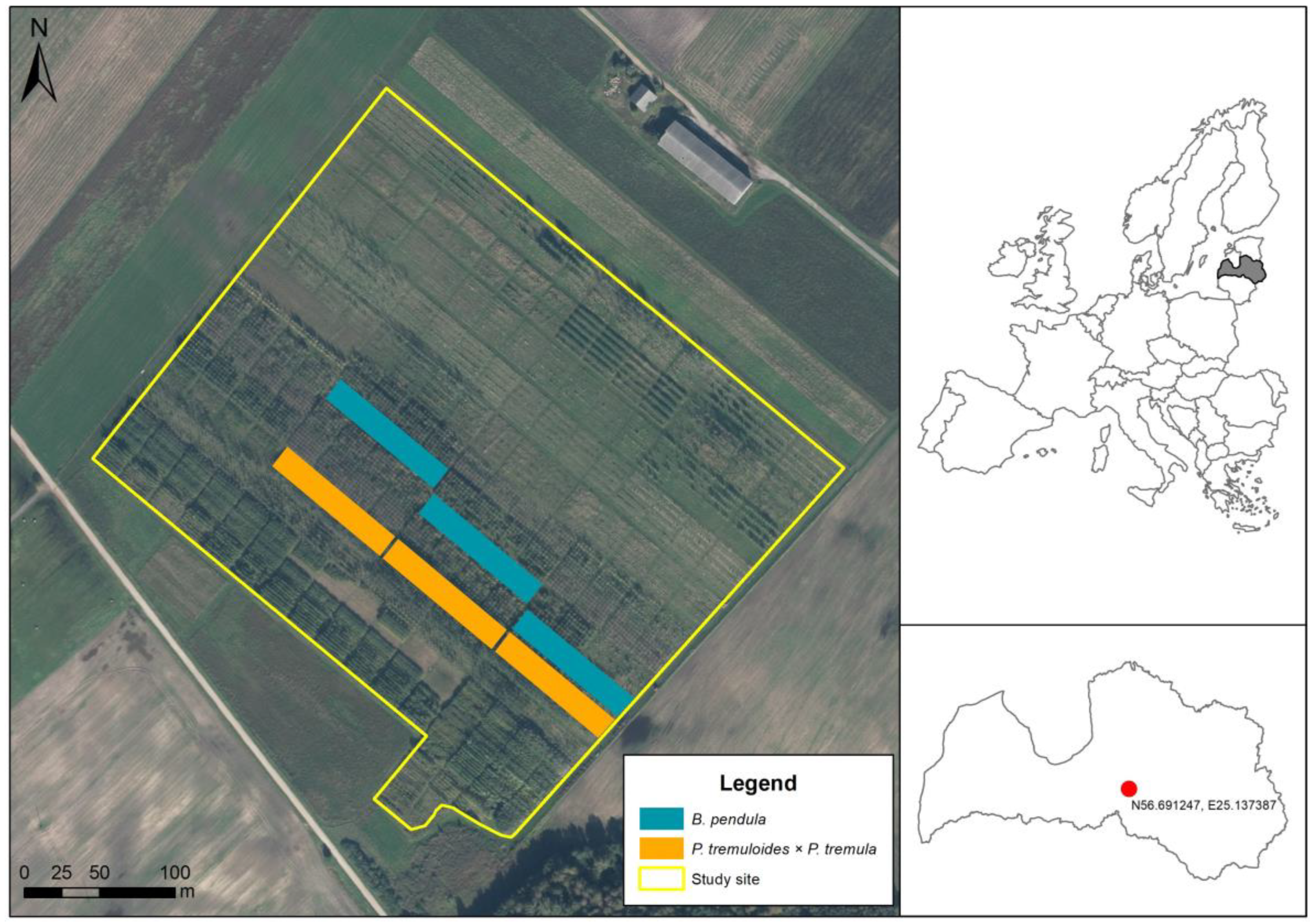
2.1. Study Site
2.2. Estimation of Tree Crown Diameter
2.3. Measurements and Calculations of Additional Variables
2.4. Statistical Analysis
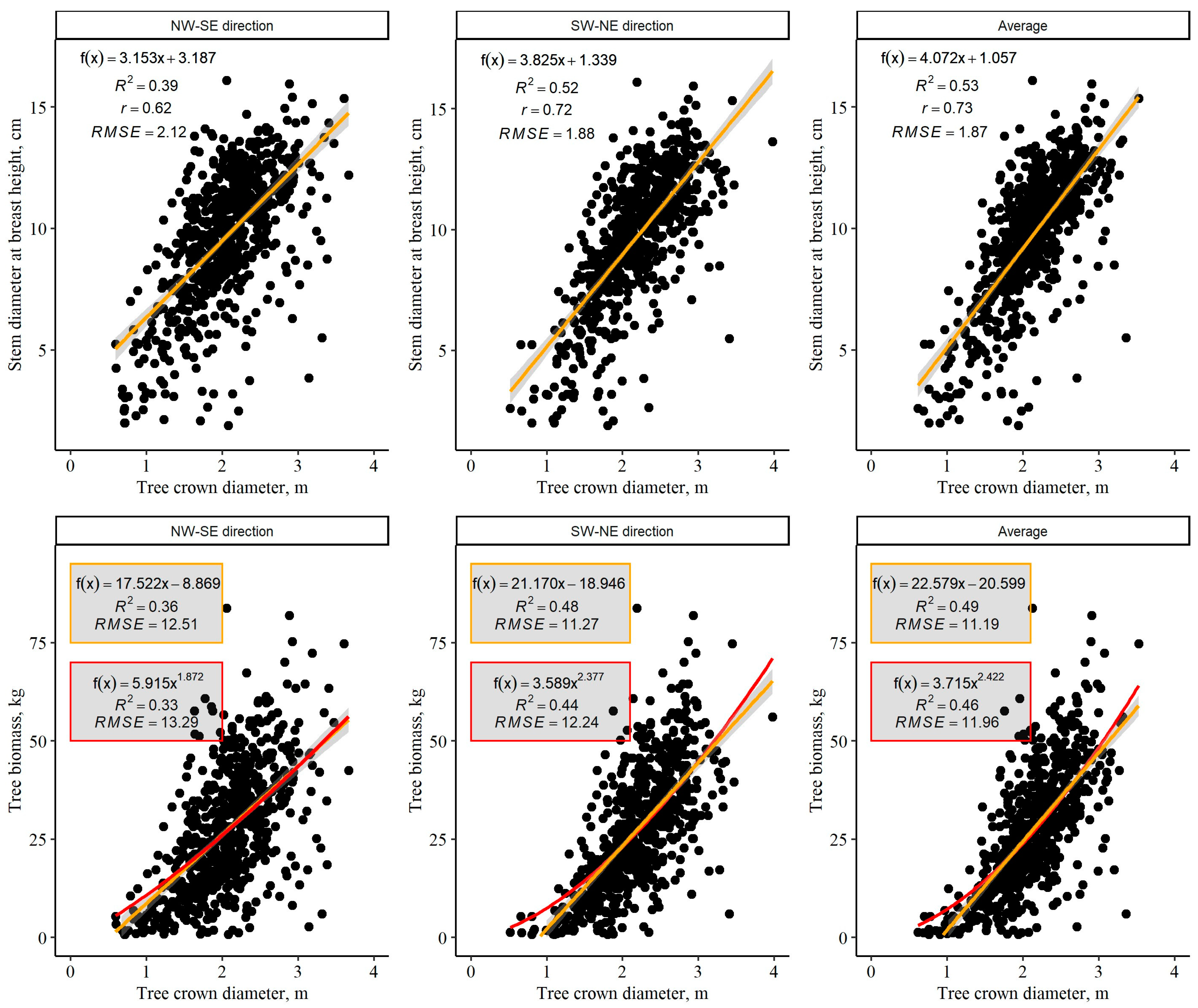
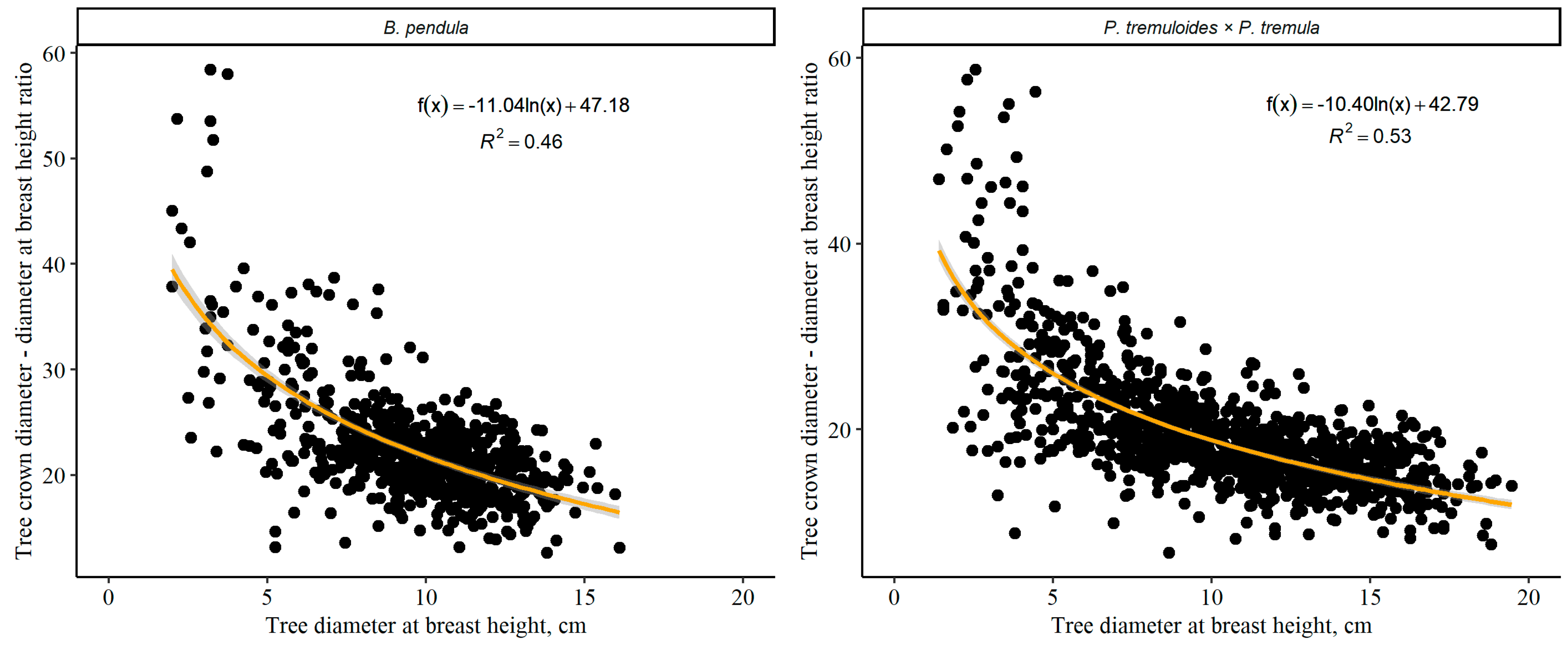
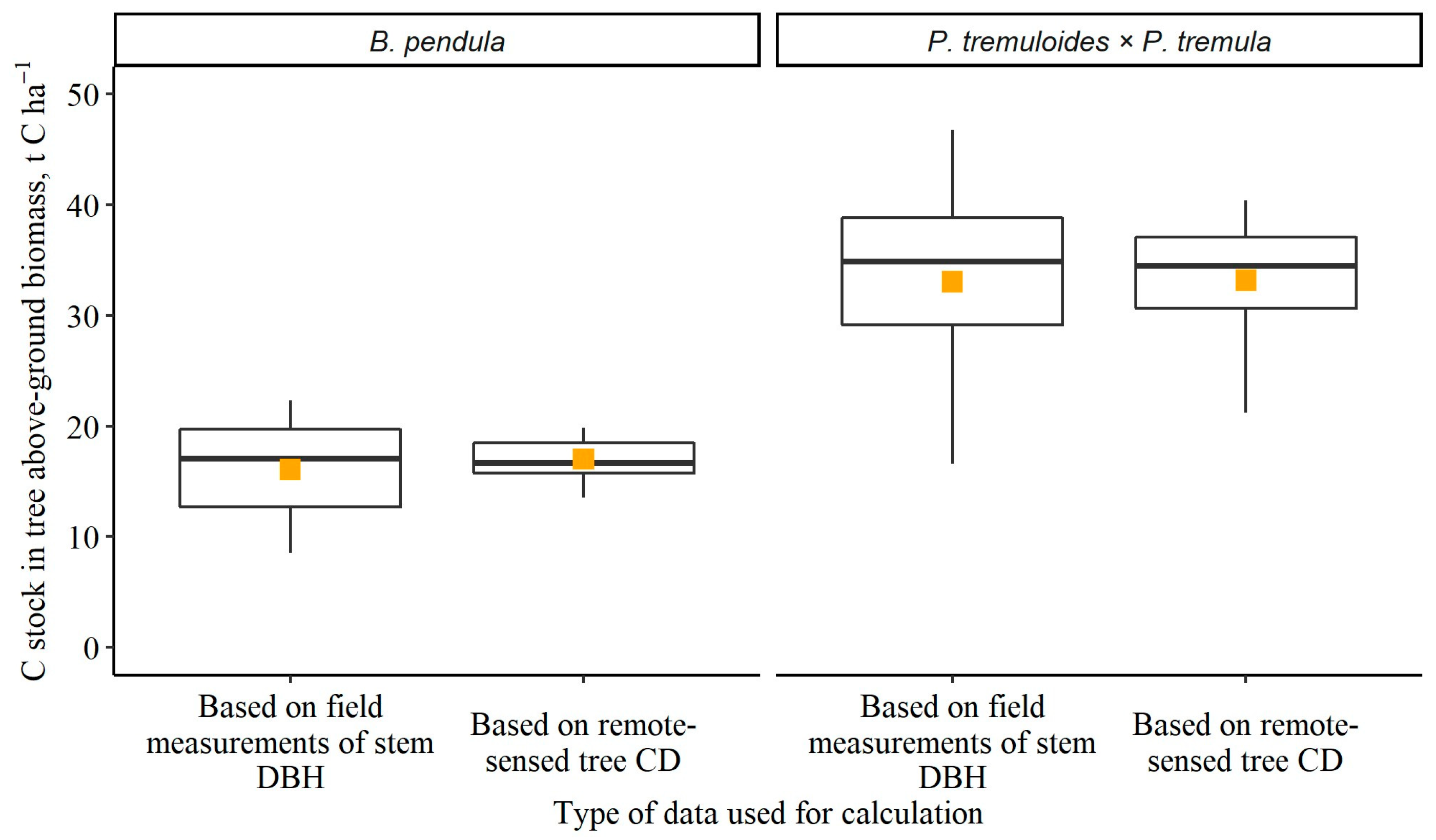
3. Results
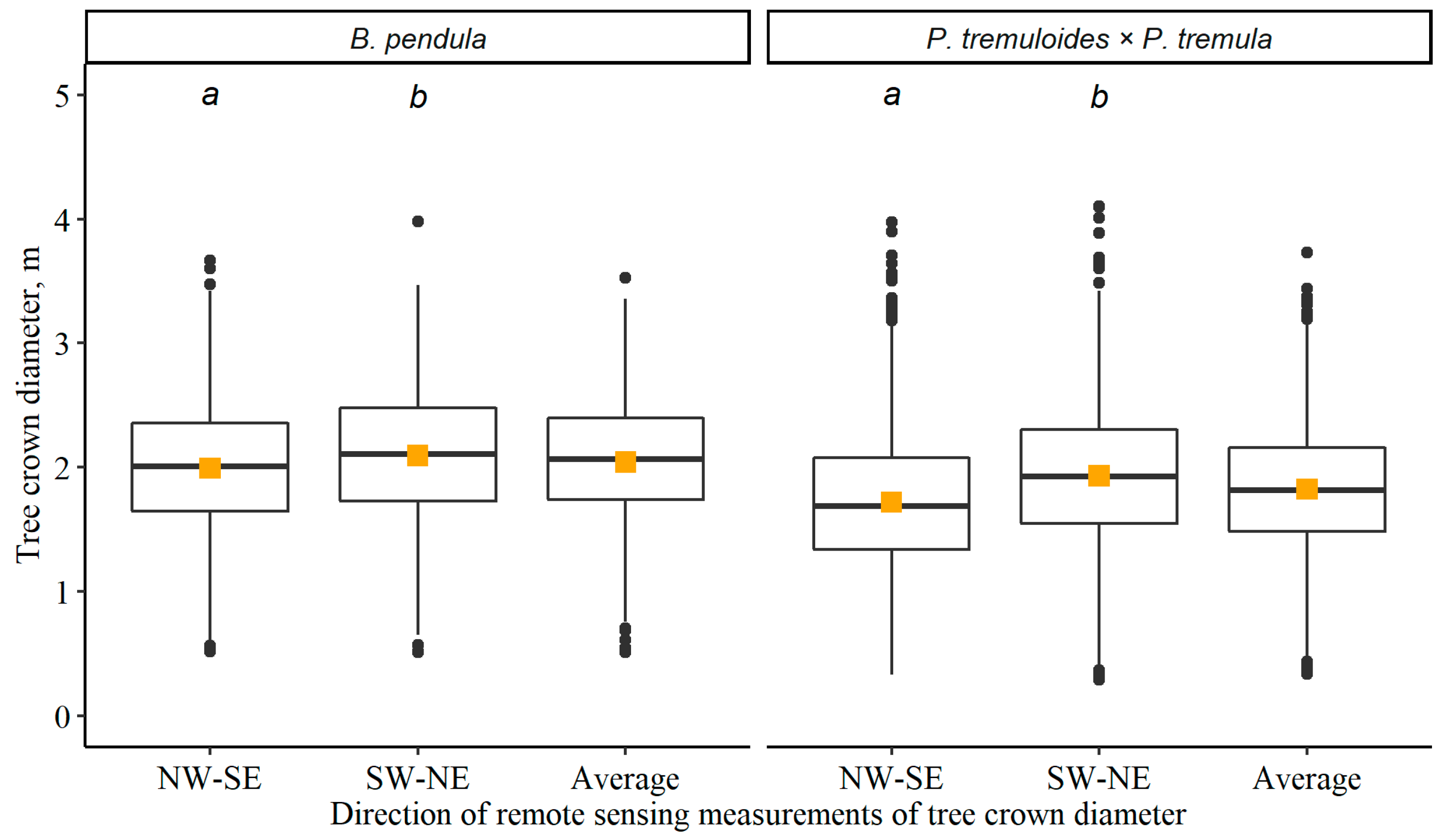
3.1. Tree Crown Diameter and Impact of Survival of the Adjacent Trees
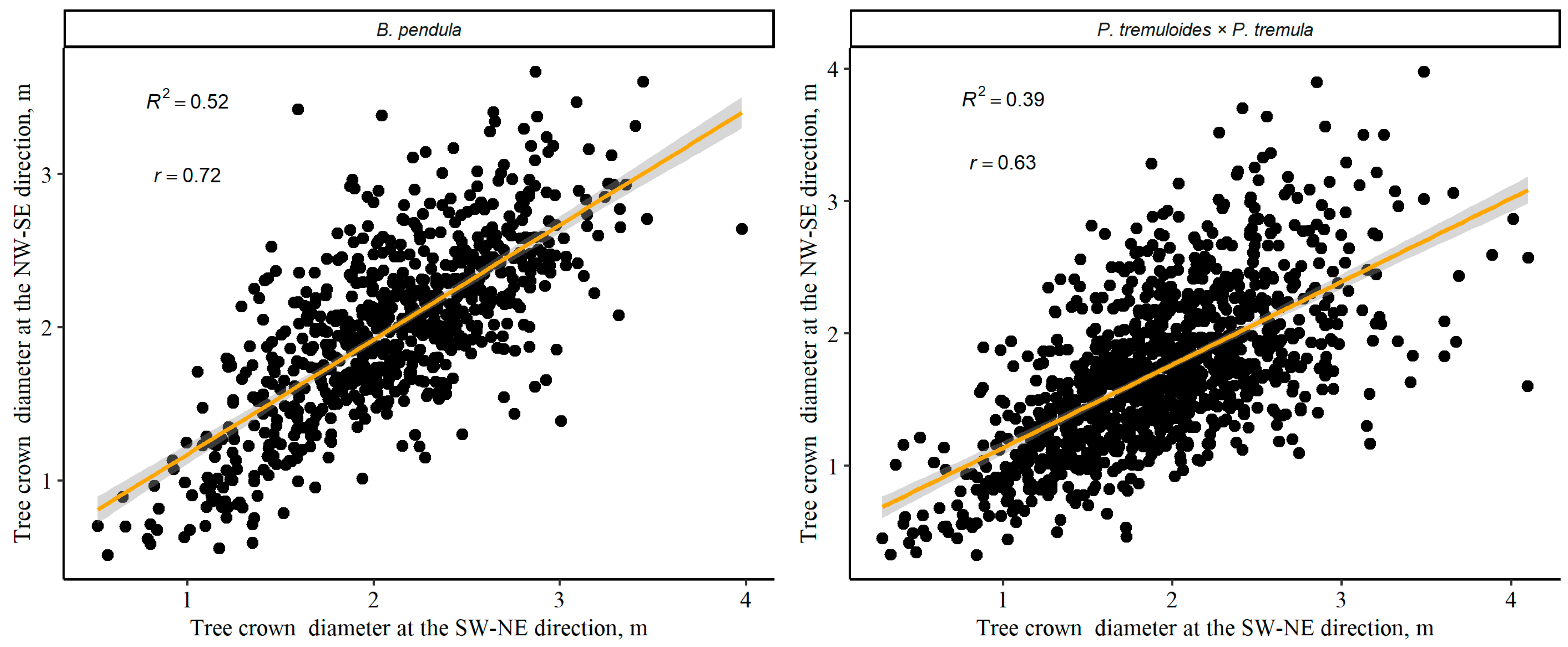
3.2. Relationship between Individual Tree Crown Diameter and Other Tree Variables
3.3. Estimation of Approximate C Stock in Tree Above-Ground Biomass
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Popescu, S.C.; Wynne, R.H.; Nelson, R.F. Measuring individual tree crown diameter with lidar and assessing its influence on estimating forest volume and biomass. Can. J. Remote Sens. 2003, 29, 564–577. [Google Scholar] [CrossRef]
- Panagiotidis, D.; Abdollahnejad, A.; Surový, P.; Chiteculo, V. Determining tree height and crown diameter from high-resolution UAV imagery. Int. J. Remote Sens. 2017, 38, 2392–2410. [Google Scholar] [CrossRef]
- Jucker, T.; Caspersen, J.; Chave, J.; Antin, C.; Barbier, N.; Bongers, F.; Dalponte, M.; van Ewijk, K.Y.; Forrester, D.I.; Haeni, M.; et al. Allometric equations for integrating remote sensing imagery into forest monitoring programmes. Glob. Chang. Biol. 2017, 23, 177–190. [Google Scholar] [CrossRef]
- Dalponte, M.; Frizzera, L.; Ørka, H.O.; Gobakken, T.; Næsset, E.; Gianelle, D. Predicting stem diameters and aboveground biomass of individual trees using remote sensing data. Ecol. Indic. 2018, 85, 367–376. [Google Scholar] [CrossRef]
- Liu, Z.; Long, J.; Lin, H.; Du, K.; Xu, X.; Liu, H.; Yang, P.; Zhang, T.; Ye, Z. Interpretation and mapping tree crown diameter using spatial heterogeneity in relation to the radiative transfer model extracted from GF-2 images in planted boreal forest ecosystems. Remote Sens. 2023, 15, 1806. [Google Scholar] [CrossRef]
- Guo, Q.; Su, Y.; Hu, T. Applications of LiDAR in biodiversity conservation, ecohydrology, and ecological process modeling of forest ecosystems. In LiDAR Principles, Processing and Applications in Forest Ecology; Guo, Q., Su, Y., Hu, T., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 407–442. [Google Scholar]
- Hu, L.; Xu, X.; Wang, J.; Xu, H. Individual tree crown width detection from unmanned aerial vehicle images using a revised local transect method. Ecol. Inform. 2023, 75, 102086. [Google Scholar] [CrossRef]
- Ivanovs, J.; Lazdiņš, A.; Lang, M. The influence of forest tree species composition on the forest height predicted from airborne laser scanning data—A case study in Latvia. Balt. For. 2023, 29, 663. [Google Scholar] [CrossRef]
- Wynne, R.H. Forest mensuration with remote sensing: A retrospective and a vision for the future. In Forest Mensuration with Remote Sensing; Gen. Tech. Rep. SRS 75; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2004; pp. 109–116. [Google Scholar]
- AWF-Wiki. Crown Attributes. Available online: http://wiki.awf.forst.uni-goettingen.de/wiki/index.php/Crown_attributes (accessed on 14 September 2023).
- Hemery, G.E.; Savill, P.S.; Pryor, S.N. Applications of the crown diameter–stem diameter relationship for different species of broadleaved trees. For. Ecol. Manag. 2005, 215, 285–294. [Google Scholar] [CrossRef]
- Freudenberg, M.; Magdon, P.; Nölke, N. Individual tree crown delineation in high-resolution remote sensing images based on U-Net. Neural. Comput. Appl. 2022, 34, 22197–22207. [Google Scholar] [CrossRef]
- Bonnor, G.M. Stem diameter estimates from crown width and tree height. Commonw. For. Rev. 1968, 47, 8–13. [Google Scholar]
- Lockhart, B.R.; Weih, R.C., Jr.; Smith, K.M. Crown radius and diameter at breast height relationships for six bottomland hardwood species. J. Ark. Acad. Sci. 2005, 59, 110–150. [Google Scholar]
- Kalliovirta, J.; Tokola, T. Functions for Estimating Stem Diameter and Tree Age Using Tree Height, Crown Width and Existing Stand Database Information. Silva Fenn. 2005, 39, 227–248. [Google Scholar] [CrossRef]
- The Paris Agreement. Available online: https://unfccc.int/process-and-meetings/the-paris-agreement (accessed on 11 October 2023).
- European Commission. The European Green Deal. Available online: https://commission.europa.eu/strategy-and-policy/priorities-2019-2024/european-green-deal_en (accessed on 11 October 2023).
- European Commission. Carbon farming. Available online: https://climate.ec.europa.eu/eu-action/sustainable-carbon-cycles/carbon-farming_en (accessed on 14 September 2023).
- Zomer, R.J.; Bossio, D.A.; Trabucco, A.; Noordwijk, M.; Xu, J. Global carbon sequestration potential of agroforestry and increased tree cover on agricultural land. Circ. Agric. Syst. 2022, 2, 1–10. [Google Scholar] [CrossRef]
- Oficiālās Statistikas Portals. Afforestation (ha) 2013–2022. Available online: https://data.stat.gov.lv/pxweb/lv/OSP_PUB/START__NOZ__ME__MEA/MEP042 (accessed on 14 September 2023).
- Lutter, R.; Stål, G.; Arnesson Ceder, L.; Lim, H.; Padari, A.; Tullus, H.; Nordin, A.; Lundmark, T. Climate benefit of different tree species on former agricultural land in northern Europe. Forests 2021, 12, 1810. [Google Scholar] [CrossRef]
- Ministru Kabineta Noteikumi Nr. 198 “Tiešo Maksājumu Piešķiršanas Kārtība Lauksaimniekiem”. Available online: https://likumi.lv/ta/id/341260-tieso-maksajumu-pieskirsanas-kartiba-lauksaimniekiem#piel1&pd=1 (accessed on 14 September 2023).
- Lauksaimniecības un Lauku Attīstības Likums. Available online: https://likumi.lv/ta/id/87480-lauksaimniecibas-un-lauku-attistibas-likums (accessed on 14 September 2023).
- Lauku Atbalsta Dienests. Platību Maksājumu Statistika. Available online: https://www.lad.gov.lv/lv/platibu-maksajumu-statistika (accessed on 14 September 2023).
- White, J.C.; Coops, N.C.; Wulder, M.A.; Vastaranta, M.; Hilker, T.P.; Tompalski, P. Remote sensing technologies for enhancing forest inventories: A review. Can. J. Remote Sens. 2016, 42, 619–641. [Google Scholar]
- Kārkliņš, A.; Rancāne, S. Augsnes Apraksts, Reģ. Nr. AI0103; LLU Augsnes un Augu Zinātņu Institūts: Skrīveri, Latvija, 2012; pp. 1–2. [Google Scholar]
- Kārkliņš, A.; Rancāne, S. Augsnes Apraksts, Reģ. Nr. AI0104; LLU Augsnes un Augu Zinātņu Institūts: Skrīveri, Latvija, 2012; pp. 1–2. [Google Scholar]
- Latvian Environment, Geology and Meteorology Centre. Meteorological Network. Available online: https://videscentrs.lvgmc.lv/ (accessed on 14 September 2023).
- Ministru Kabineta Noteikumi Nr. 108 “Meža Ieaudzēšanas un Plantāciju Mežu Noteikumi”. Available online: https://likumi.lv/ta/id/4754-meza-ieaudzesanas-un-plantaciju-mezu-noteikumi (accessed on 21 September 2023).
- Bārdulis, A.; Purviņa, D.; Makovskis, K.; Bārdule, A.; Lazdiņa, D. Soil-to-atmosphere GHG fluxes in hemiboreal deciduous tree and willow coppice based agroforestry systems with mineral soil. Land 2023, 12, 715. [Google Scholar] [CrossRef]
- Bardule, A.; Grinfelde, I.; Lazdina, D.; Bardulis, A.; Sarkanabols, T. Macronutrient leaching in a fertilized juvenile hybrid aspen (Populus tremula L. × P. tremuloides Michx.) plantation cultivated in an agroforestry system in Latvia. Hydrol. Res. 2018, 49, 407–420. [Google Scholar] [CrossRef]
- Naslund, M. Skogsförsöksanstaltens Gallringsförsök i Tallskog. [Forest Research Institute’s Thinning Experiments in Scots Pine Forests]; Meddelanden från Statens Skogsförsöksanstalt: Stockholm, Sweden, 1937; p. 249. [Google Scholar]
- Liepiņš, J.; Lazdiņš, A.; Liepiņš, K. Equations for estimating above- and belowground biomass of Norway spruce, Scots pine, birch spp. and European aspen in Latvia. Scand. J. For. Res. 2018, 33, 58–70. [Google Scholar] [CrossRef]
- Lamlom, S.H.; Savidge, R.A. A reassessment of carbon content in wood: Variation within and between 41 North American species. Biomass Bioenergy 2003, 25, 381–388. [Google Scholar] [CrossRef]
- IPCC. 2006 IPCC Guidelines for National Greenhouse Gas Inventories; Prepared by the National Greenhouse Gas Inventories Programme; Eggleston, H.S., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Eds.; IGES: Tokyo, Japan, 2006. Available online: https://www.ipcc-nggip.iges.or.jp/pub-lic/2006gl/index.html (accessed on 4 September 2023).
- R Core Team. The R Project for Statistical Computing. Available online: https://www.R-project.org (accessed on 14 September 2023).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 4 September 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. Available online: https://ggplot2.tidyverse.org (accessed on 4 September 2023).
- Niemistö, P. Influence of initial spacing and row-to-row distance on the growth and yield of silver birch (betula pendula). Scand. J. For. Res. 1995, 10, 245–255. [Google Scholar] [CrossRef]
- Broks, J. Meža Enciklopēdija, 1st ed.; Apgāds Zelta grauds: Rīga, Latvia, 2003; p. 368. [Google Scholar]
- Tuvikene, M. Growth and Management of Silver Birch and Hybrid Aspen in Southern Sweden. Master’s Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2021. [Google Scholar]
- Rouvinen, S.; Kuuluvainen, T. Structure and asymmetry of tree crowns in relation to local competition in a natural mature Scots pine forest. Can. J. For. Res. 1997, 27, 890–902. [Google Scholar]
- Aakala, T.; Shimatani, K.; Abe, T.; Kubota, Y.; Kuuluvainen, T. Crown asymmetry in high latitude forests: Disentangling the directional effects of tree competition and solar radiation. Oikos 2016, 125, 1035–1043. [Google Scholar]
- Dubois, H.; Claessens, H.; Ligot, G. Towards silviculture guidelines to produce large-sized Silver birch (Betula pendula Roth) logs in western Europe. Forests 2021, 12, 599. [Google Scholar]
- Malek, S.; Miglietta, F.; Gobakken, T.; Næsset, E.; Gianelle, D.; Dalponte, M. Prediction of stem diameter and biomass at individual tree crown level with advanced machine learning techniques. iForest 2019, 12, 323–329. [Google Scholar] [CrossRef]
- Nonini, L.; Fiala, M. Estimation of carbon storage of forest biomass for voluntary carbon markets: Preliminary results. J. For. Res. 2019, 32, 329–338. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štāls, T.A.; Bārdule, A.; Dūmiņš, K.; Makovskis, K.; Lazdiņa, D. Remote-Sensed Tree Crown Diameter as a Predictor of Stem Diameter and Above-Ground Biomass in Betula pendula Roth and Populus tremuloides Michx. × Populus tremula L. Plantations. Land 2023, 12, 2006. https://doi.org/10.3390/land12112006
Štāls TA, Bārdule A, Dūmiņš K, Makovskis K, Lazdiņa D. Remote-Sensed Tree Crown Diameter as a Predictor of Stem Diameter and Above-Ground Biomass in Betula pendula Roth and Populus tremuloides Michx. × Populus tremula L. Plantations. Land. 2023; 12(11):2006. https://doi.org/10.3390/land12112006
Chicago/Turabian StyleŠtāls, Toms Artūrs, Arta Bārdule, Kārlis Dūmiņš, Kristaps Makovskis, and Dagnija Lazdiņa. 2023. "Remote-Sensed Tree Crown Diameter as a Predictor of Stem Diameter and Above-Ground Biomass in Betula pendula Roth and Populus tremuloides Michx. × Populus tremula L. Plantations" Land 12, no. 11: 2006. https://doi.org/10.3390/land12112006
APA StyleŠtāls, T. A., Bārdule, A., Dūmiņš, K., Makovskis, K., & Lazdiņa, D. (2023). Remote-Sensed Tree Crown Diameter as a Predictor of Stem Diameter and Above-Ground Biomass in Betula pendula Roth and Populus tremuloides Michx. × Populus tremula L. Plantations. Land, 12(11), 2006. https://doi.org/10.3390/land12112006