1. Introduction
In many areas of the world, fire is an important ecological factor that influences the structure and functions of ecosystems by selecting organisms, affecting ecological biodiversity, nutrient cycles, and the biological, chemical, and physical properties of soils [
1,
2,
3,
4]. Mediterranean-climate areas are among the most fire-prone regions in the world, and for millions of years, wildfires have represented a periodic disturbance factor that affected plant species’ traits, compositions, and structures, as well as the dynamics of natural vegetation and, consequently, shaped landscapes [
1,
2,
5]. Among others, it is worth considering that in the Mediterranean Basin, landscapes also derive from a very long history of interactions between man and environmental conditions [
6]. Periodical fires, deforestation, and grazing were largely adopted in less favored areas as strategies to maximize the production of goods and ecosystem services, generating typical agro-sylvo-pastoral landscapes currently recognized for their historical and natural values [
6,
7,
8]. Indeed, many habitats of community importance of the Habitat Directive [
9] depend on the traditional agro-sylvo-pastoral activities of Mediterranean countries [
7,
8,
10,
11,
12,
13,
14].
The effects of fire on ecosystems and landscapes are mainly related to the sudden loss of biomass and habitats, the heat released, and the changes in physicochemical and biological soil properties [
1,
3,
15,
16]. Depending on the severity, size, and frequency of fires; the type of vegetation; and other site-specific factors, the consequences can be twofold. Sporadic and low-severity fires can contribute to renewing vegetation and promoting structural complexity and biodiversity of ecosystems, but too frequent, large, and severe fires can cause long-term consequences such as shifts in ecological succession, permanent changes in plant community composition and diversity, ecosystem services loss, changes in soil properties, soil erosion, and runoff [
1,
3,
17,
18]. Moreover, frequent and large wildfires can, directly and indirectly, threaten human lives, compromise economic activities and natural resources and contribute to carbon emissions into the atmosphere [
17,
19,
20].
It is well known that in fire-prone regions such as the Mediterranean Basin, plant species are adapted (more resistant or resilient), and the natural vegetation is more resilient to fire [
2]. Throughout a wildfire, the individuals of many Mediterranean plant species are adapted to sacrifice themselves or their above-ground portion to guarantee the persistence of plant life. Post-fire vegetation recovery mainly relies on resprouting of new shoots from survived belowground parts of plants or germination of heat tolerant seeds [
21,
22,
23]. For example, in many species, such as those belonging to the
Fabaceae and
Cistaceae families, seed germination is even increased by exposure to heat, smoke, or ash generated by fire [
23,
24]. The cork oak (
Quercus suber L.) has developed a thick cork bark that forms a fire-resistant cuirass, making it the only European tree capable of epicormic resprouting after high-intensity fires [
25,
26]. As a result of these strategies, generally, the post-fire vegetation recovery in Mediterranean environments follows an auto-succession model, which takes a relatively short time if compared with other ecoregions, for example only a few years for shrublands [
1,
2]. Nevertheless, particularly in forest ecosystems, an increasing frequency and severity of fires can induce transitions to other stable non-forest communities [
1,
27].
In the Mediterranean Basin, an increasing frequency and severity of wildfire events have been observed in the last few years [
1,
28] and 98% of them are human-induced [
2]. This trend is mainly linked to the intensification of droughts and heat waves, and to the progressive land abandonment, which contributes to flammable phytomass accumulation and continuity at the landscape scale [
1]. Under the climate change scenario, the frequency and severity of wildfires are expected to increase especially in the Mediterranean Basin, recognized as being among the most affected by intensification in droughts and heat waves in the future [
19,
22,
29]. This evidence raises concern about the possible negative effects on post-fire vegetation recovery dynamics, biodiversity, traditional landscapes, ecosystem functions, and services, and, therefore, on human well-being [
2,
21].
Consequently, much research has been focused on the effects of fire on vegetation and soil, e.g., [
3,
30,
31,
32,
33]; post-fire vegetation dynamics, e.g., [
1,
2,
34]; the effects of fire on biodiversity, e.g., [
35,
36,
37]; landscape recovery and fire-risk mitigation, e.g., [
38,
39,
40]; methods based on remote sensing to quantify burned areas, severity levels, vegetation recovery rates and effects of fires on the carbon cycle, e.g., [
19,
41,
42,
43,
44]. The aim of all these studies can be summarized as the need to comprehend the complex roles and effects of wildfires on ecological systems, the repercussion on human life, and to prevent or mitigate the negative consequences, particularly from the perspective of climate change.
As shown by the ever-growing literature, remote sensing has become an important tool for the analysis of burned areas. The multispectral images captured by the sensors aboard the satellites, such as those of the European Union’s Earth Observation Programme (Copernicus), offer the opportunity to easily estimate the areas affected by wildfires, the impact on vegetation, and the post-fire vegetation dynamics. With these purposes, various spectral indices have been conceived to compare pre- and post-fire spectral reflectance, or to measure changes over time. Among the spectral indices, those based on short-wave infrared (SWIR) and near-infrared (NIR) wavelengths (e.g., the differenced normalized burn ratio (dNBR)) are recognized as the most reliable for quantifying burned areas, estimating burn severity, and detecting post-fire vegetation recovery [
19,
20,
21]. Although spectral indices provide information on spectral vegetation changes and contribute to assessing the post-fire vegetation recovery rates, they do not provide information on vegetation structural changes [
1]. To assess the effective impact of fire on vegetation and soil, and to better understand the trends of vegetation dynamics in terms of structure and species composition, remote sensing spectral analyses may be combined with field surveys [
45,
46].
Although several studies have investigated the post-fire vegetation recovery in different terrestrial ecosystems, e.g., [
42,
47,
48,
49,
50,
51,
52,
53,
54], there have not been many studies that specifically explored the response of natural vegetation in a relatively short span of time after a fire in the context of Mediterranean landscapes. Particularly, in the first years after a fire, vegetation is heavily reduced or simplified in its structure and diversity [
2,
51], landscapes are affected in their ecological complexity [
55] and identity value [
56], and soil runoff and erosion increase by several times compared to natural conditions [
18]. Therefore, the first years after a fire are the most critical and deserve more attention to promptly understand the short-term effects of fire, analyze the strength of the first stages of vegetation recovery, and identify the most vulnerable areas where to concentrate possible restoration plans. This is especially important in terms of adaptation in the context of climate change.
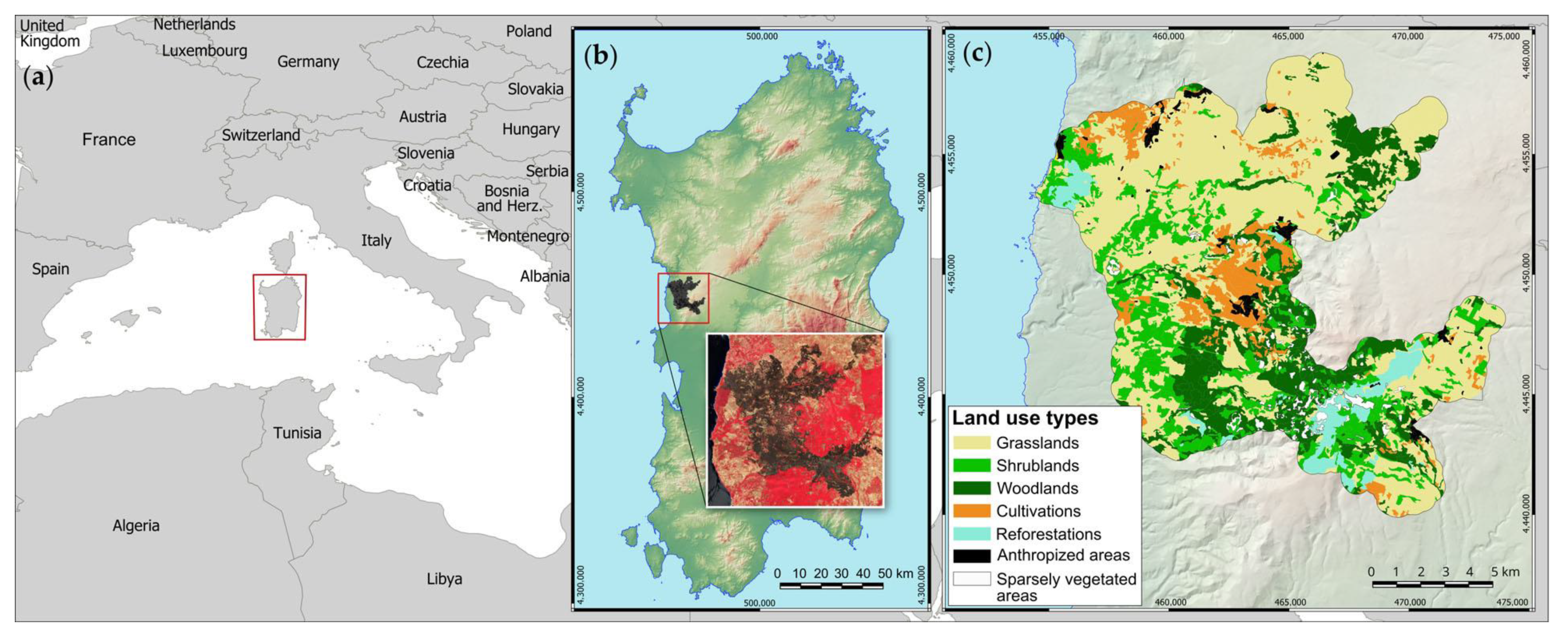
In this framework, the aim of this study was to investigate the short-term vegetation response in the context of an agro-sylvo-pastoral Mediterranean landscape struck by a “megafire”, i.e., a wildfire that exceeds 10,000 ha in area [
57]. More precisely, it was the largest wildfire in Italy in 2021 [
58]. Vegetation recovery one year after the fire has been studied in three main types of vegetation that represent different stages of the vegetation series: semi-natural grasslands, shrublands, and woodlands. Multispectral data from the Copernicus Sentinel-2 mission were used to estimate burn severity levels after the fire extinction and vegetation regeneration rate one year later. In addition, one year after the fire, field surveys were carried out to estimate total vegetation cover and height, plant species presence, and abundance in burned and unburned areas.
4. Discussion
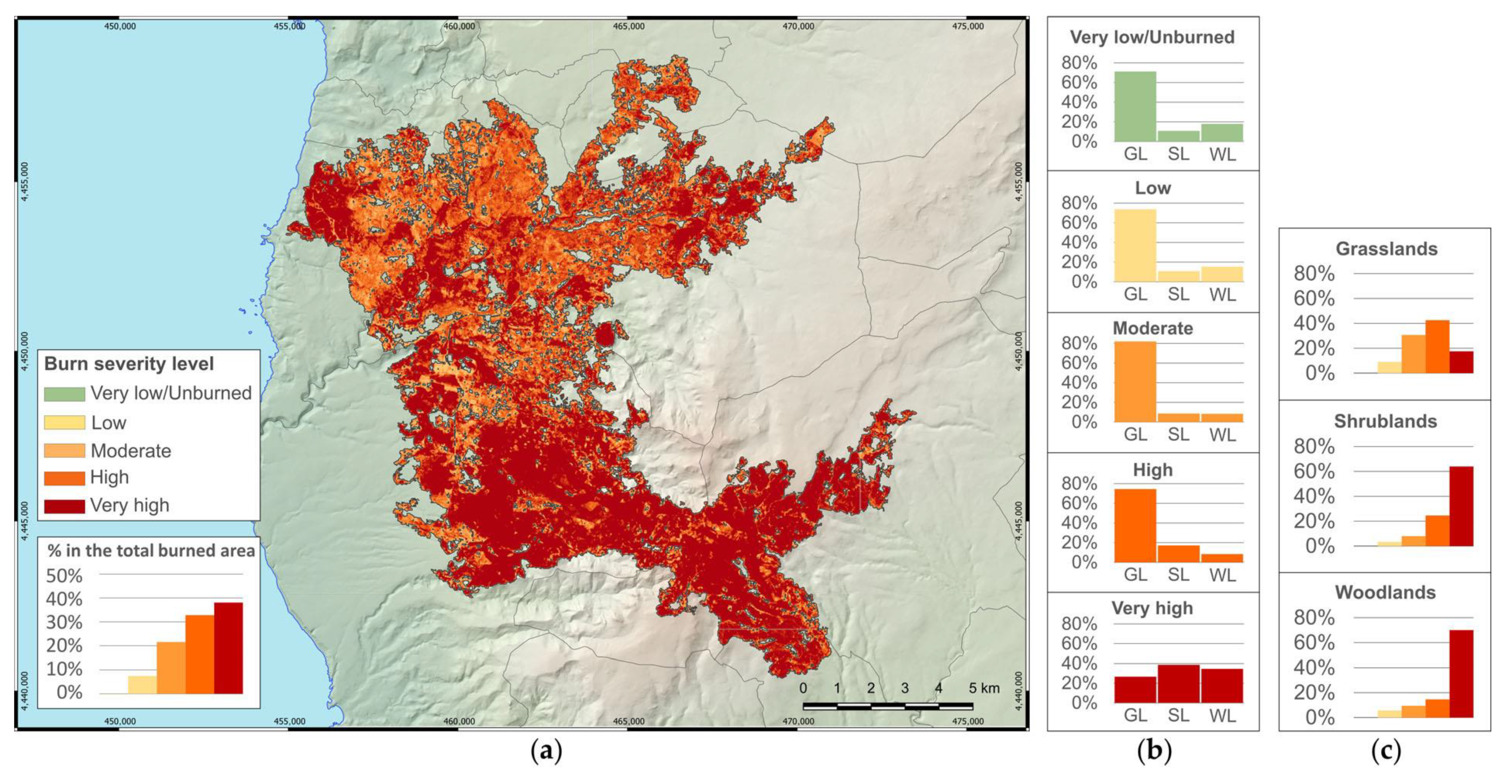
Regarding the effects of the megafire, the total burned area calculated through the dNBR index is 8.5% smaller than that estimated by EFFIS (13,278 ha), which uses MODIS images. This result confirms what has been observed by Llorens et al. 2021 [
19] about the better accuracy provided by the high spatial resolution images of Sentinel-2 (resolution 10 m) with respect to MODIS (resolution 250 m) when mapping burned areas. The burned area reveals the magnitude of the megafire (0.5% of the Sardinian territory) which represents 63% of the total burned area in the region in 2021 [
58].
Burn severity levels, estimated through the dNBR index, showed that the largest part of the area was affected by the highest severity levels and the very high severity level was the most represented. The relevance of the highest severity levels in the study area was probably related to the widespread presence of semi-natural vegetation at the landscape scale and, consequently, to the large availability of flammable phytomass [
5,
67] and the high flammability of Mediterranean plant species [
68,
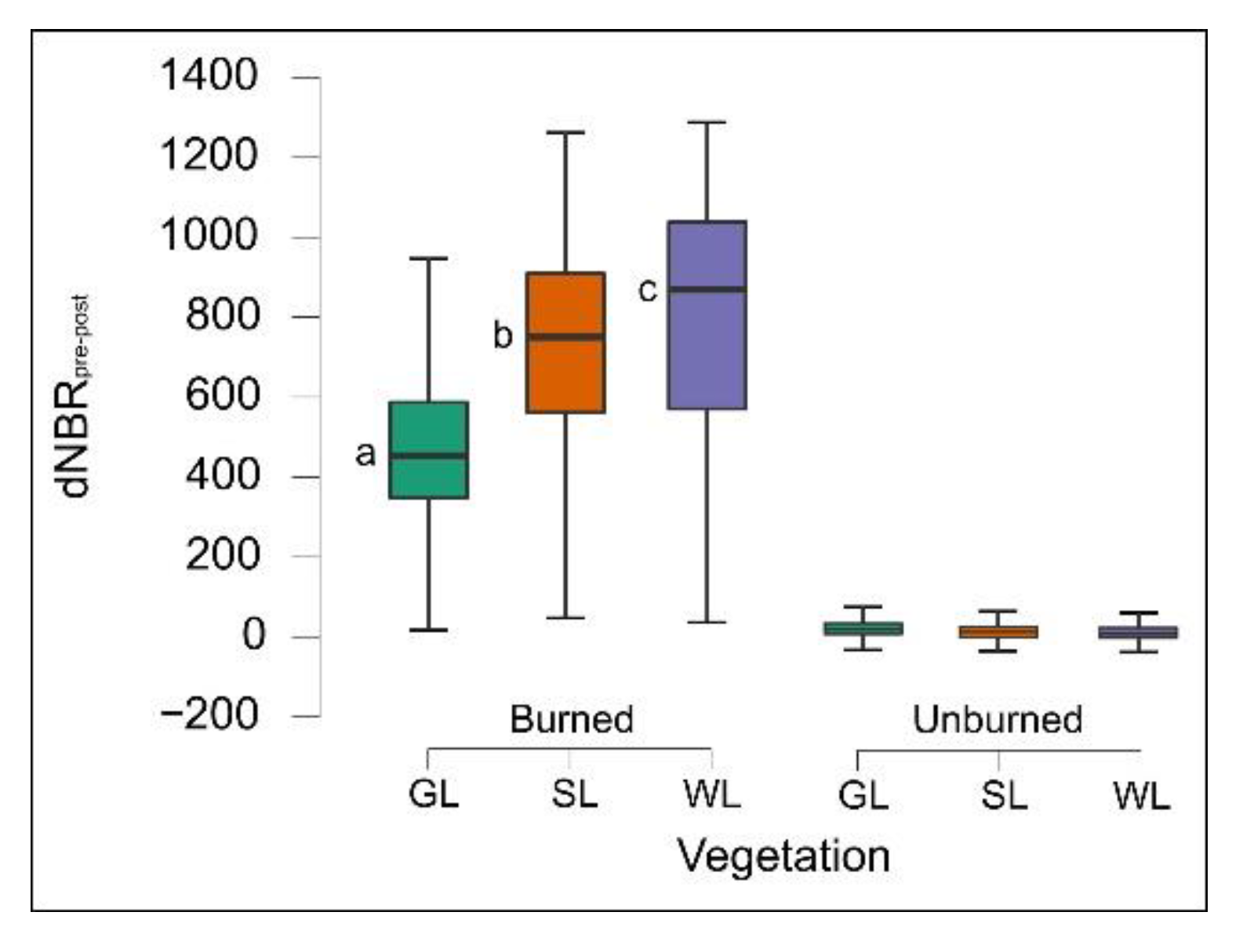
69]. In the study area, patches of woodlands and shrublands are surrounded by a landscape matrix of grasslands mainly consisting of semi-natural plant communities that often include scattered woody species. In fact, the areas classified as very high severity were mostly shrublands and woodlands due to the high presence of flammable phytomass, but an important part was grasslands as well. On the other hand, as expected, when considering the three types of vegetation separately, woodlands and shrublands were mostly affected by very high severity, while grasslands were mainly affected by high and moderate severity. The nonparametric analyses confirmed the significantly different partitioning of dNBR
pre-post values among vegetation types, with higher values in woodlands, lower values in shrublands, and the lowest in grasslands, following the different amounts of live above-ground biomass and dead organic matter stocks among the vegetation types [
70].
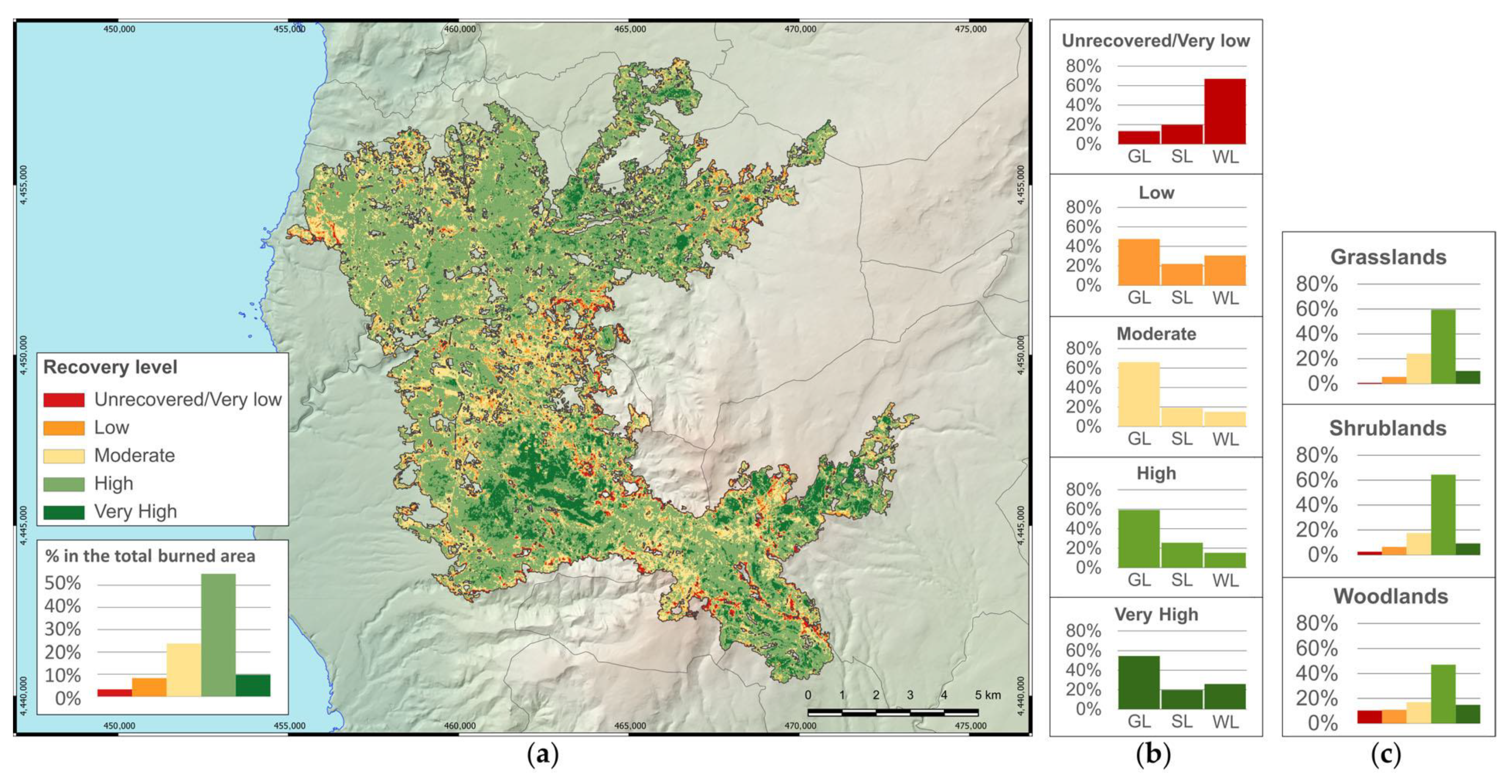
One year after the fire, a telling change in severity levels was observed. The unburned/very low severity level, which was 0.1% of the burned area after fire extinction, covered more than half of the study area one year later. Differences among vegetation types became more explicit, with the highest dNBR
pre-1yr values in woodlands, intermediate values in shrublands, and the lowest in grasslands. This is also displayed in a quite clear way by the nonparametric analyses, showing the different resilience of the three vegetation types. It confirmed the high resilience of grasslands with respect to the other types of vegetation, which need more time to regain their original condition [
46,
71]. In fact, most of the areas classified as unburned/very low severity fell in grasslands and more than four-fifths of grasslands were classified as unburned/very low severity level. The resilience of grasslands has to be considered as a positive aspect from a landscape scale point of view, for different reasons: in the study area, semi-natural grasslands represent the main landscape matrix; therefore, they contribute to a faster recovery of landscape patterns and connectivity [
72], and also of cultural ecosystem services such as aesthetic and identity values [
56,
72]. Herbaceous vegetation of grasslands can quickly protect soil from erosion risk [
72] and grasslands are related to extensive grazing activities, which are among the main sources of income in the area. Additionally, some habitats of community interest and a relevant part of plant diversity are linked to semi-natural grasslands, e.g., [
7].
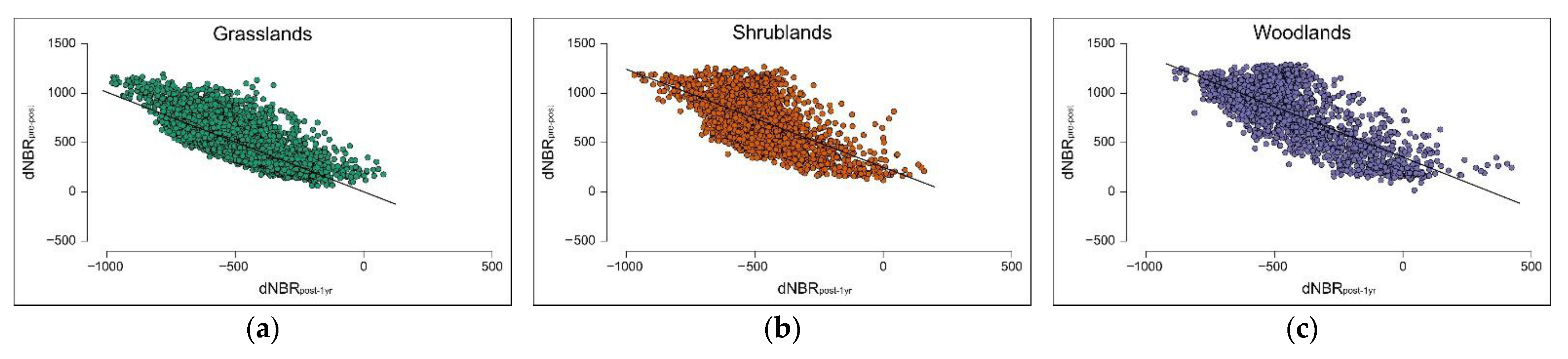
In terms of spectral recovery, one year after the fire, more than half of the burned area showed high and very high recovery levels. Moderate recovery was important as well, covering almost a quarter of the total burned area. These findings confirm that all the vegetation types substantially started to recover, but most of the areas with the highest and moderate recovery levels were included in grasslands. In contrast to dNBRpre-1yr, dNBRpost-1yr values did not differ between shrublands and woodlands. This finding suggests that above-ground phytomass regrowth in shrublands and woodlands, at least during the first year after the fire, proceeds at a similar pace. Areas classified as unrecovered (3.1% of the burned area) mostly fell in woodlands and appear to be located close to the boundaries of the burned area, where low and unburned/very low post-fire severity levels were detected. This observation becomes more evident when correlating the index of spectral recovery (dNBRpost-1yr) with the index of burn severity (dNBRpre-post): all vegetation types showed a highly significant inverse relationship between the indices, suggesting that higher burn severity levels were linked to higher vegetation recovery levels.
One year after the fire, grasslands had totally recovered. In woodlands, the lowest values of burn severity were partially related to positive dNBR
post-1yr values, which indicate phytomass loss. This finding needs further investigations to be clarified, but it can be explained with delayed fire mortality, i.e., tree mortality started with fire, but occurred at a later time, mainly affecting older trees and fire-resistant species such as
Quercus spp. [
73]. Another explanation can be related to improper attribution of severity levels in some areas, due to commission errors in the computation of dNBR
pre-post values. This can occur, for example, in the case of a sub-canopy burn, i.e., when a fire burns only phytomass underneath a dense tree canopy cover. In this case, the satellite sensor cannot detect spectral changes under the tree canopy. Sub-canopy burns are common in low-severity fires; therefore, they can occur near the fire perimeters [
74]. Furthermore, post-fire phytomass loss can be attributable to forest fire cleanup operations after fire extinction, such as the removal of hazardous trees or flammable vegetation near the boundaries of the burned area to avoid reignition.
Areas with the highest burn severity and the lowest recovery levels one year after the fire deserve more attention in terms of vegetation dynamics and factors that hamper or slow down it. If any plan for vegetation restoration should be realized, it should be focused on these areas [
18,
75]. In particular, in burned woodlands low recovery rates were observed, suggesting that in some areas phytomass loss took place during the first year after the fire. Soil erosion risk can be higher if these areas fall in steep slopes [
75,
76], therefore these should be the areas where to concentrate monitoring activities and, if necessary, plans for vegetation restoration.
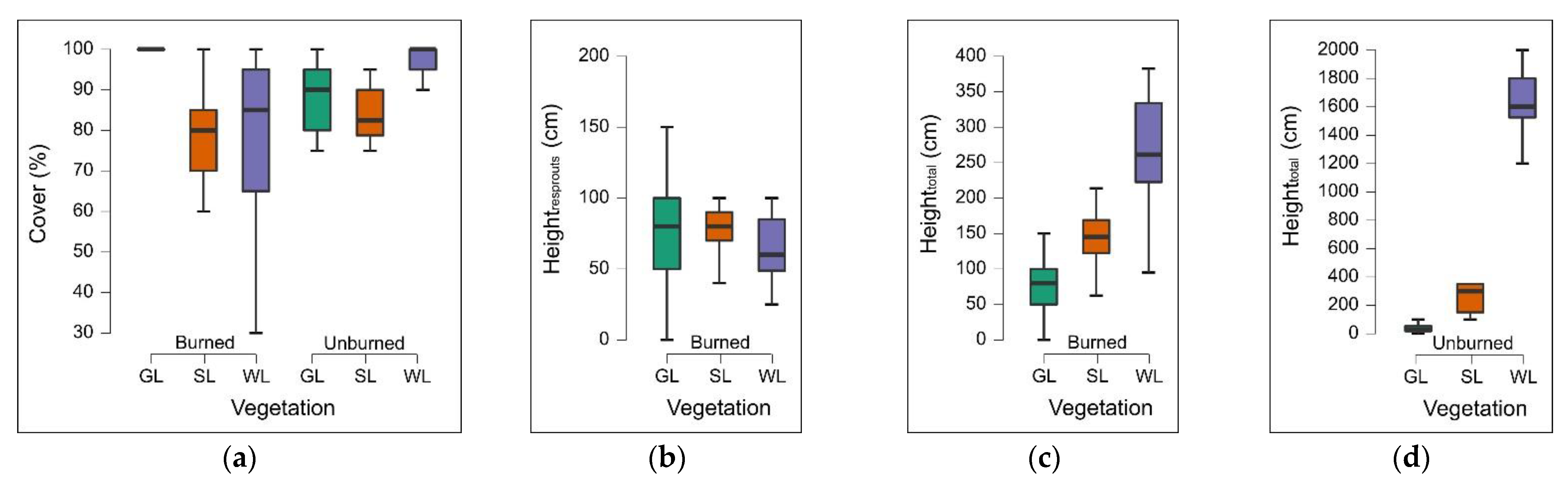
Field data confirmed the trend observed using spectral indices. Vegetation cover in burned grasslands was complete one year after the fire, slightly higher and less variable than in unburned grasslands. Even the median height in burned grasslands was higher than in unburned ones. These findings indicate that above-ground phytomass in burned grasslands is higher than in unburned ones, which could be counterintuitive, but it is possible to offer different explanations for this observation: (i) in the burned grasslands, an increasing presence of
Pteridium aquilinum (L.) Kuhn. was observed. This species became dominant in coverage and height in many of the sampled grasslands; (ii) fire increases the soil nutrients which can sustain plant growth and productivity in the short-term [
48,
77]; (iii) in burned grasslands, the phytomass may have had the possibility to grow more as an effect of the suspension or reduction of grazing activities and other forms of land management [
78,
79].
Values of dNBR
pre-1yr and dNBR
post-1yr showed to be mostly correlated with vegetation cover. The height of resprouts did not show any correlation with dNBR values. In fact, one year after the fire, the median height of new vegetation (Height
resprouts) was similar among vegetation types. It is probably necessary to wait at least the second year after the fire to see a significant correlation between the height of resprouts and dNBR values. Conversely, the total height of vegetation (Height
total) seems to be correlated with dNBR values in a reversed way with respect to vegetation cover. However, this result should be considered trivial because the total height of vegetation was the lowest in grasslands and the highest in shrublands and woodlands, while vegetation cover had the opposite trend. Snags of surviving trees and shrubs, which contributed to an increase in the total height of burned shrublands and woodlands, also did not contribute effectively to vegetation cover, as also observed by Bolton et al. (2017) [
80] and White et al. (2018) [
81] in boreal forests. Therefore, at least during the first year after the fire, in the study area, dNBR values prove to be more correlated with vegetation cover than with vegetation height, and it is due both to no differences in height of new regenerating vegetation and to the presence of residual structures in the canopy (snags of surviving trees and shrubs) that did not contribute to vegetation cover. It is reasonable to expect a clearer correlation between dNBR values and vegetation height with the progress of vegetation recovery.
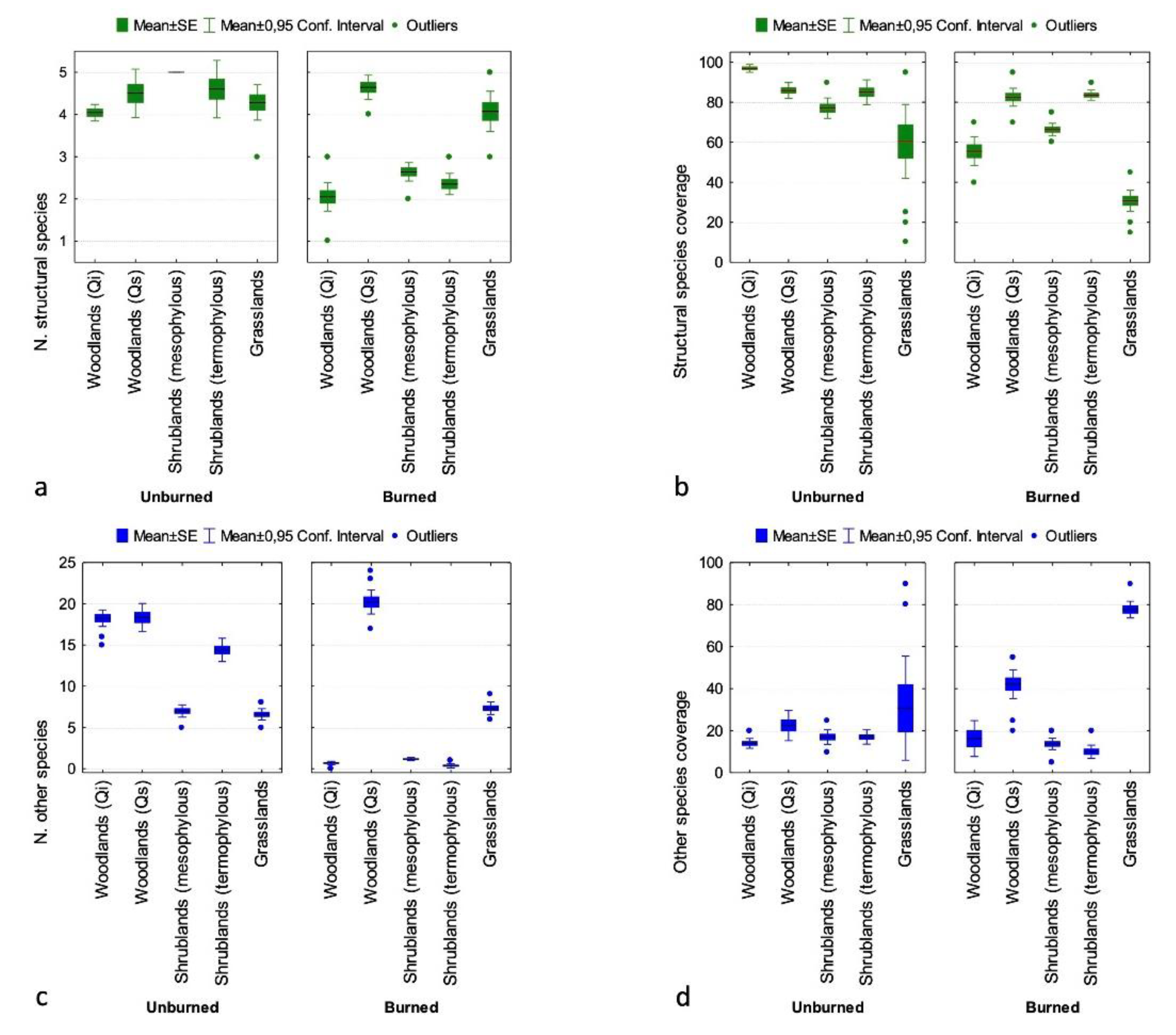
Detailed field vegetation surveys highlighted a general recovery of all vegetation types, although they followed different patterns related to the strategies adopted by plant species to respond to fire. In particular,
Q. suber-dominated communities showed a rapid recovery rate in mean height and coverage thanks to the epicormic resprouting capabilities of trees [
25,
26]. On the other side, a less considerable recovery has been observed in
Q. ilex woodlands, where the recovery was ensured only by basal resprouting. Therefore, in these formations, a significantly lower mean height was observed. A general reduction in mean height and coverage was always observed in both mesophilous and thermophilous shrublands, while grasslands completely recovered in terms of mean height and coverage. In shrublands, vegetation cover was similar between burned and unburned ones and significantly lower than in burned grasslands. Overall, in shrublands and woodlands, vegetation recovery through resprouts was observed, but vegetation recovery was also linked to plants that survived the fire. These surviving plants contribute to the total height of burned vegetation, but one year after the fire the median total height was still quite far from the unburned condition. The general absence of an herbaceous layer in burned shrublands and woodlands one year after the fire has to be pointed out. Hence, vegetation cover in burned shrublands and woodlands was mostly linked to resprouting of survived plants and the germination of shrub and tree seeds. The absence of a herbaceous layer can be explained by the absence of a soil seed bank of herbaceous species [
54]. It can be assumed that the absence of the herbaceous layer in burned shrublands and woodlands can expose these less resilient vegetation types to a higher soil erosion risk when the vegetation cover is still incomplete. Although it is not the main subject of this research, the effects of fire on soil properties deserve to be discussed. Indeed, fires not only affect flora, but also soil physical, chemical, and biological properties [
3]. Fires influence soil properties in complex ways, including changes in texture and aggregate stability [
82], density and porosity [
83], water content and repellency [
84], organic matter amount and quality [
20,
85], soil pH [
86], soil biota composition and activity [
87], nutrient recycling and availability [
82], and consequently, affecting the post-fire dynamics of vegetation [
3]. In the case study, the impacts of the megafire on soil properties were not addressed, but it is important to consider their possible effects on long-term vegetation dynamics. Moreover, the possible detrimental effects on soil properties and the lack of vegetation cover in the less resilient vegetation types could have an additive effect on soil erosion risk, which can be more serious, especially in steep slopes [
75,
76].
Field surveys showed a general loss of plant diversity in burned areas compared to unburned ones; this loss was probably related to the short time elapsed since the megafire and to the different capability and speed of response of each species. In fact, several plant species mainly linked to the
Q. ilex formations, such as
Taxus baccata L.,
Ilex aquifolium L., and
Crataegus monogyna Jacq., have not been found. This observation confirms what has been observed in previous studies about the gradual disappearance of
T. baccata as an effect of recurring fires [
88,
89]. Conversely, other species, such as
Cytisus villosus Pourr., reached remarkable coverage values in both
Q.ilex and
Q. suber woodlands, but also in shrublands, where
C. villosus, Teline monspessulana (L.) K.Koch.,
Genista desoleana Vals. subsp.
desoleana, and, rarely and limited to the lowest altitudes,
Myrtus communis L. reached high coverage values as well. The rapid response to fire of
C. villosus,
Teline monspessulana, and
G. desoleana subsp.
desoleana was expected since several studies demonstrated that
Fabaceae species show a positive germination response of seeds to heat shock, e.g., [
90,
91,
92], and some
Cytisus species also reveal high vegetative post-fire regeneration [
93]. Furthermore, the observed increase of
C. villosus coverage in the study area is in accordance with what has been observed by Xofis et al. (2021) [
2] in northeast Mediterranean ecosystems, where this opportunistic species can become dominant after a fire. In burned grasslands, instead, a pivotal role was played by
P. aquilinum, which becomes dominant in height and coverage. It is important to consider that
P. aquilinum is a toxic fern species that can be harmful to grazing animals, in particular bovines, and its toxins can probably pass into milk [
94,
95].
P. aquilinum is a cosmopolitan species that can become invasive in burned or deforested areas, thanks to fire-resistant spores and its long underground rhizomes that store carbohydrates (and are immune to fire) as well as a lot of dormant buds [
96]. The post-fire environment is, therefore, favorable for its survival and spread [
96,
97]. In addition, its high productivity and the emission of allelopathic components, which can hamper the establishment of other plant species, are well documented [
96]. For these reasons, the increasing presence of
P. aquilinum in burned grasslands is cause for concern since livestock activities based on wild grazing are the prevailing form of land use and one of the main sources of income in the study area.
An alarming observation of this study underlines the loss of several interesting plants from a conservation point of view; in fact, in all burned areas, no signal of recovery was observed for
T. baccata,
I. aquifolium, and
Laurus nobilis L., indicating that, although there was a general positive recovery of all vegetation types, several peculiar floristic elements could be definitively replaced by other species; this process could also imply a progressive loss of habitats, such as the “Arborescent matorral with
Laurus nobilis” [
98] or the “Mediterranean
Taxus baccata woods” [
99], which are habitats of European interest. To better investigate this critical aspect, further studies and repeated long-term monitoring activities should be planned in the Montiferru area.
5. Conclusions
This study is one of the first attempts to quantify the short-term response of natural vegetation after a megafire in a traditional agro-sylvo-pastoral Mediterranean landscape. It is mainly focused on the first stages of vegetation recovery in order to contribute to the assessment of natural vegetation resilience in the context of the Mediterranean Basin, which is among the most endangered by wildfire intensification and climate change. The study was carried out at two different spatial scales, but related to each other: the landscape scale, taking advantage of the viewpoint offered by satellites and the power of spectral indices; and the field scale, through floristic and vegetation surveys on the field, which are essential to better understand the true dynamics of plant life.
The results confirm that Mediterranean vegetation can considerably respond to fire in a short span of time. In the case study, post-fire vegetation response in terms of phytomass recovery is noticeable just after the first year, although high and very high post-fire burn severity levels were prevalent. This rapid response is mostly a result of the adaptations that Mediterranean plant species have evolved as post-fire regeneration strategies. Significant regrowth was dependent on basal resprouting strategy, but epicormic resprouting was also important in cork oak-dominated formations. We also confirmed the increased presence of plant taxa that mainly respond as seeders, probably because the seed germinability is favored by heat shock. Moreover, in terms of vegetation types, the study confirms the higher resilience of semi-natural grasslands, which can positively affect the recovery of landscape connectivity, patterns, biodiversity, and identity value. Even though shrublands and woodlands showed to be less resilient than grasslands, they showed a significant capability to respond to the damage caused by the fire and, against expectations, the regeneration rate seems to be proportional to burn severity.
All the findings of this study show the importance of temporal resolution when assessing the short-term effects of a wildfire. This is especially true for fire-prone areas such as the Mediterranean Basin, where the vegetation has developed specific adaptations to respond to fire. In such areas, the post-fire burn severity is not enough to correctly estimate the actual magnitude of the impact, because burn severity levels are not necessarily related to the apparent loss of above-ground phytomass. In fact, the burn severity concept includes short- and long-term impacts in the post-fire environment and it also considers vegetation response processes [
4,
45,
54]. In a Mediterranean context, the assessment of burn severity levels exclusively close to the fire extinction date can lead to misinterpretation of the damage the fire caused to ecosystems. Therefore, for example, some areas can be erroneously classified as having very high severity levels and, consequently, can draw more attention for possible recovery plans, while belowground phytomass is still alive and vegetation is ready to naturally recover. Conversely, at a glance, other areas could seem less damaged and not deserving of attention, but fire-resistant vegetation can be subjected to delayed mortality, plant species of conservation interest could disappear, or invasive species can be favored. Therefore, as also stated by Key 2006 [
45] and Key and Benson 2006 [
46], for the short-term assessment, it is important to monitor throughout the first year after a fire.
Moreover, the results obtained at the field scale show the relevance of a detailed analysis of flora and vegetation in addition to the remote sensing analysis. This is essential not only to assess the impact of fire on biodiversity, structure, and functions of ecosystems but also to promptly monitor the direction of the early stages of the vegetation dynamics. For example, as shown in the case study, plant communities of semi-natural grasslands used to feed livestock can be subjected to changes in species composition and richness, invasion of noxious species (e.g., P. aquilinum), and, consequently, loss of fodder value. Furthermore, species of conservation interest were not found again during the field surveys. Similarly, the relevance of field scale analyses can also be extended to the assessment of the effects of fire on soil properties, which can affect soil conservation and post-fire vegetation dynamics.
The findings of this study suggest further investigations and repeated monitoring activities in the Montiferru area to contribute to defining specific protocols to evaluate burn severity levels in Mediterranean environments, and to assess vegetation recovery over the medium and long term in order to better understand which factors can hamper or slow down the vegetation dynamics and, if necessary, identify those areas that really need restoration or conservation plans.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}