1. Introduction
Invasive species are a threat to global biodiversity, as they can adversely affect ecosystems by displacing native and endemic species and altering ecosystem functions [
1,
2]. Preserving the integrity and biodiversity of ecosystems with high conservation values requires management actions to control the spread of invasive species [
3]. The costs of the economic loss due to invasive species and their control are very high on a global scale, with annual costs of an estimated USD 120 billion in the US [
4], USD 14.45 billion in China [
5], and up to USD 626 million over 34 years in Ecuador [
6]. Since funds for controlling these species are often scarce, it is important to allocate funds to priority areas, requiring accurate species distribution maps [
7]. Such maps can help establish baseline ecosystem conditions [
8], localize and target early infestations [
9], model invasion patterns, or monitor management outcomes [
10], all of which are essential to protect ecosystems threatened by invasive species [
2].
Remote sensing technologies offer an important resource for mapping vegetation [
11]. Multispectral satellite images, i.e., images composed of multiple spectral bands containing the amount of radiation of a range of wavelengths, have widely been used to characterize vegetation according to its spectral properties [
12,
13,
14]. Satellite sensors have different spatial resolutions, which are the ground representation of one pixel in the image. They range from low resolution, where a single pixel encompasses several trees or plants, to high resolution, where a tree or plant is represented by one or a few pixels [
15]. Lower resolutions are often used to map vegetation communities or ecosystems, whereas higher resolutions are commonly used to map single species [
16,
17]. While higher resolution imagery is costlier to acquire [
17] and has a lower temporal frequency and geographic coverage in tropical areas [
16], it has the advantage of greater detail that enhances the recognition of features. What allows the recognition of a plant species, apart from its spectral characteristics, is a combination of several features visible from above, for example, whether the plant species forms a tree crown, large monospecific stands, small patches mixed with other species, or how dense its foliage is. In this study, we refer to these characteristics as the “growth form” of the species.
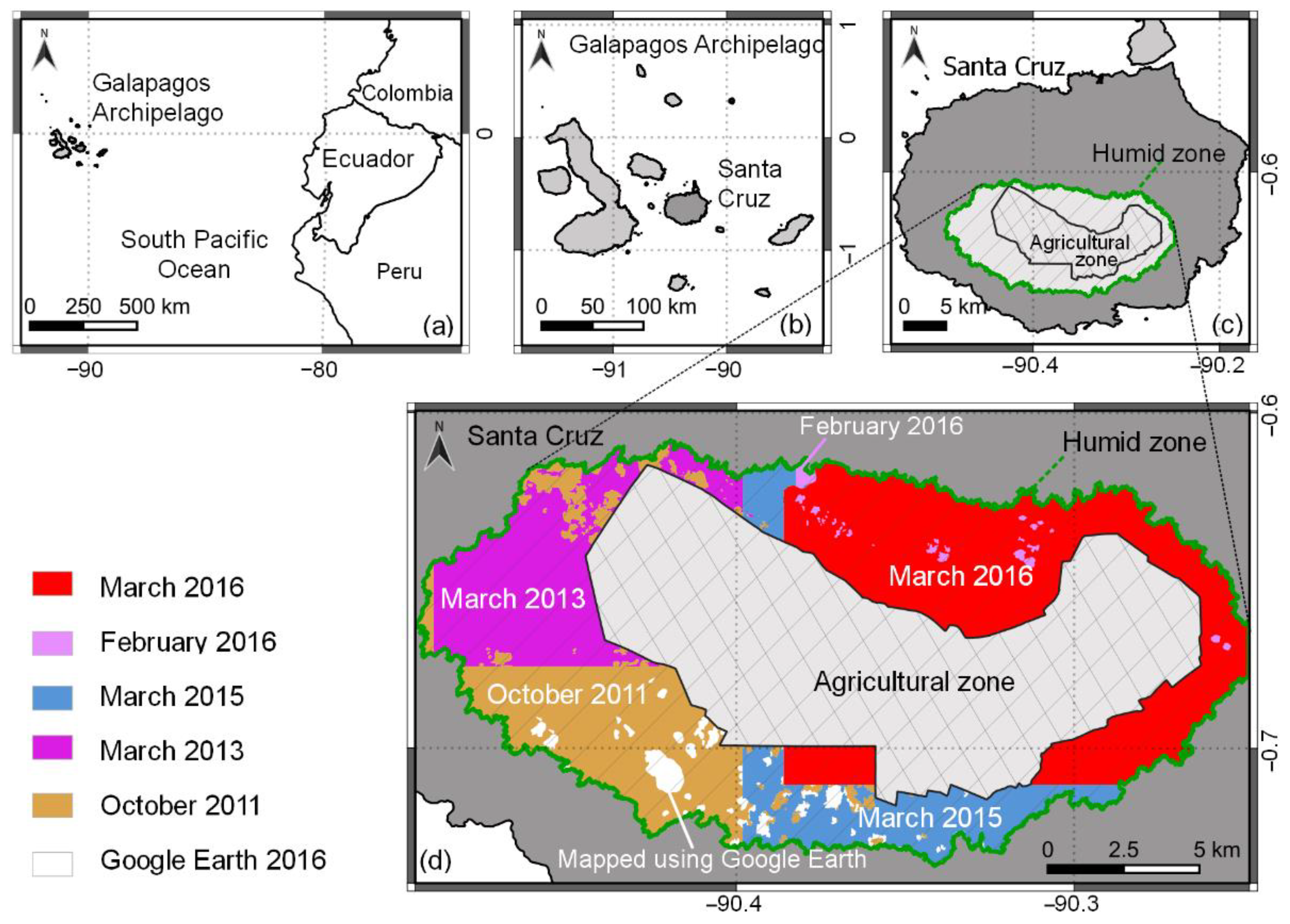
Our research was conducted in the Galapagos archipelago, a UNESCO World Heritage Site, which is known for its high endemism of species [
18]. Sadly, the unique biodiversity of terrestrial ecosystems is currently under threat from invasive plant species [
8,
19]. To be able to prioritize management actions for these invasive species, accurate distribution maps of individual species are required. Both very-high-resolution (VHR) and medium-resolution (MR) satellite imagery have been used to map the extension of invasive and other plant species in the Galapagos Islands in the past. For example, Trueman et al. [
20] used VHR imagery from the WorldView-2 imaging satellite to produce a spatial database of canopy plant densities in the protected areas of the highlands of Santa Cruz Island. The database consisted of manually delineated polygons, which included a density measure for several vegetation cover classes. Rivas-Torres et al. [
21] used MR from the Landsat 8 imaging satellite to develop an object-based methodology for mapping native and invasive vegetation cover for the protected area on all islands in the archipelago, resulting in a spatial database of vegetation units at an “ecosystem scale” that included units dominated by invasive species. Laso et al. [
22] mapped the agricultural zone, and a buffer surrounding this area, on the inhabited islands, using a combination of high resolution (HR) PlanetScope and MR Sentinel-2 satellite imagery.
The objective of our study was to advance this previous work by incorporating the detailed information of VHR imagery to model the distribution and abundance of dominant invasive plant species. Our goal was to show the effects that the spatial resolution of satellite imagery has on the outcomes of vegetation mapping in the highlands of Santa Cruz in the Galapagos National Park (GNP) area by comparing our maps produced with VHR imagery with maps produced with MR imagery by Rivas-Torres et al. [
21]. We hypothesized that VHR imagery, apart from delivering more accurate results, was crucial for the mapping of plant species with less distinctive growth forms. We argue that whereas lower resolution could be suitable for the mapping of plant species with more distinctive growth forms, higher-resolution imagery may be indispensable for the mapping of species with less distinctive growth forms.
4. Discussion
Our study compared the outcome of mapping performed with very-high-resolution (VHR) and medium-resolution (MR) satellite imagery. We mapped three invasive plant species with different growth forms on Santa Cruz Island, Galapagos: tall tree, medium tree, and shrub. As expected, we found that for each growth form, VHR imagery produced models with significantly higher overall accuracy (OA) and Kappa estimates than those resulting from the models produced with MR imagery. Several studies have compared the effects of spatial resolution on the mapping of land cover classes or individual species. Müllernová et al. [
46] demonstrated that higher-resolution imagery detected
Heracleum mantegazzianum (giant hogweed) plants more reliably. Another study from the Interior Atlantic Forest in Paraguay found that satellite imagery of higher resolution delineated land cover classes better and identified smaller patches with greater accuracy [
17]. A study from south-eastern Brazil showed that higher spatial resolution was better at detecting land cover classes and estimating the total load of suspended solids in water bodies [
47]. What is novel about our study is that we carried out a comparison of the mapping results using VHR and MR imagery across an array of different growth forms. We showed that the map resulting from MR imagery of the tall tree growth form (Cuban cedar) was moderately similar to the one resulting from VHR imagery. However, we also found that for the medium tree (guava) and shrub (blackberry) growth forms, the maps based on MR imagery widely diverged from those produced with VHR imagery.
A model performance comparison for each growth form and resolution revealed the best OA results for the tall tree growth form (Cuban cedar) (92% VHR, 81% MR), followed by the shrub (blackberry) (82% VHR, 63% MR) and the medium tree (guava) (76% VHR, 47% MR) growth forms. The weighted average of class sensitivities produced with VHR imagery was also highest for the tall tree growth form (Cuban cedar) (90%), followed by the shrub (blackberry) (86%) and the medium tree (guava) (79%) growth forms. Kappa estimates ≥ 0.5 were obtained for all VHR models and for the MR model for the tall tree growth form (Cuban cedar) only. The Kappa estimates showed that these models had a likelihood of accuracy ≥ 50% better than expected by chance alone [
45], thus indicating the models’ suitability. The highest Kappa estimate was obtained for the VHR maps of the tall tree growth form (Cuban cedar) (0.84), followed by the VHR maps of the shrub growth form (blackberry) (0.64), the MR maps of the tall tree growth form (Cuban cedar) (0.62), and the VHR maps of the medium tree growth form (guava) (0.52). While the MR model for the tall tree growth form (Cuban cedar) did not perform as well as the VHR model, its Kappa estimate showed that it still performed ≥ 50% better than by chance alone. Moreover, the OA for this MR model was only 11% lower than that for the VHR model (
Table 3). In contrast, the OA for the shrub (blackberry) model based on MR imagery was 19% lower than the one based on VHR imagery (
Table 5). The OA for the medium tree (guava) model based on MR imagery was as much as 29% lower than the one based on VHR imagery (
Table 4). Furthermore, the Kappa estimates revealed the unsuitability of MR imagery for mapping the medium tree (guava) and the shrub (blackberry) growth forms. The VHR and MR maps of these last two growth forms occupied different spatial distributions, with an overlap of as little as 2.1% of the total area mapped with VHR and/or MR.
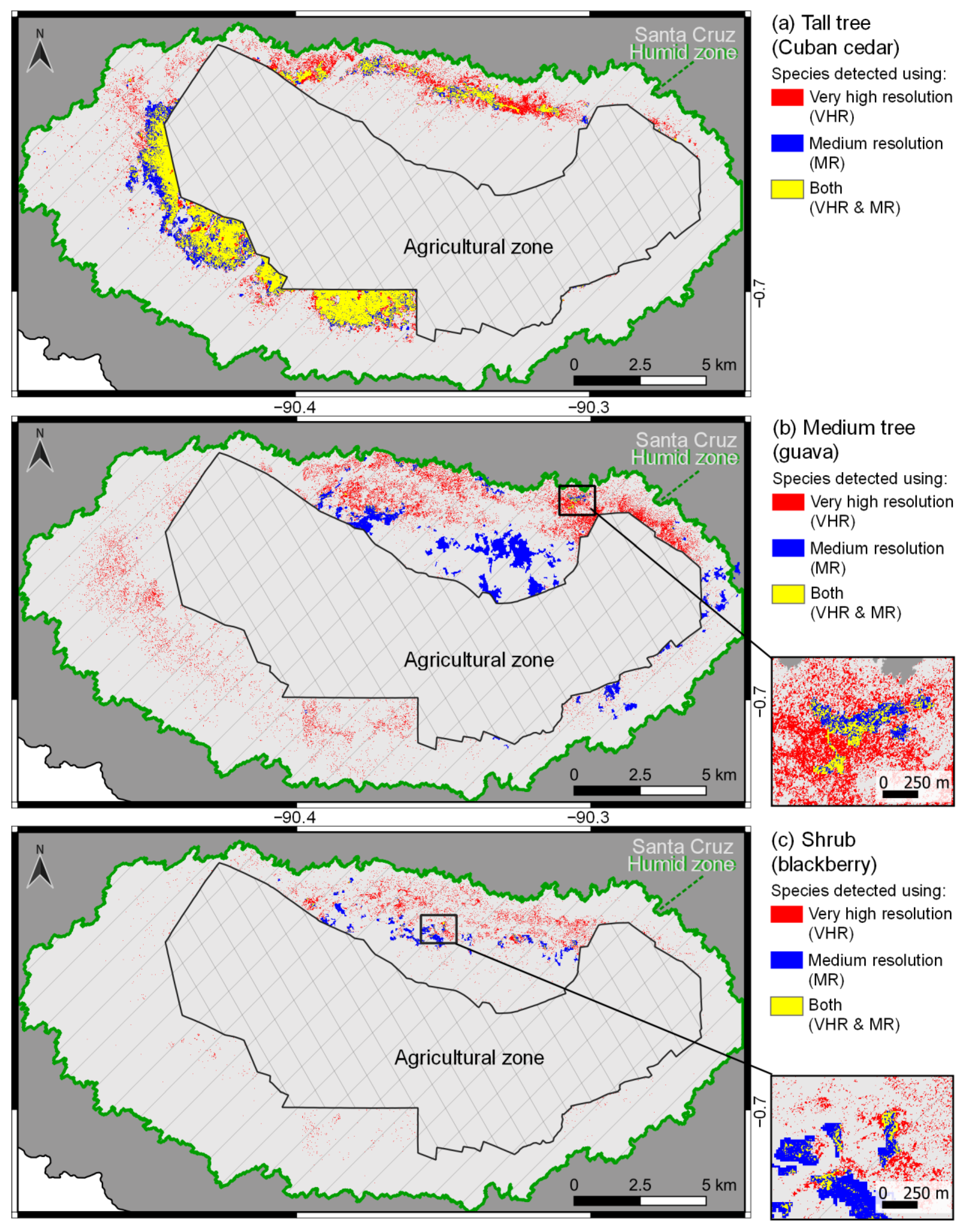
The diverging spatial distributions for the medium tree (guava) and shrub (blackberry) growth forms in maps produced with VHR and MR imagery (
Figure 2b,c), and the low Kappa estimates (<0.5) for these MR models (
Table 4 and
Table 5), showed that the MR area estimates were unreliable. Reliable estimates of the area covered by invasive species are crucial to help assess the extent of an invasion and to plan the resources needed to control these species [
48,
49]. However, while higher resolutions usually provide the most accurate estimates, there are cases where lower resolutions can deliver comparable results. For example, when comparing forest cover in eastern Paraguay, only marginal differences were encountered between the MR and VHR total area estimates [
17]. We found a similar result when mapping the tall tree growth form (Cuban cedar) produced by MR, which underestimated the area by only 6.9% (
Table 6). The comparability of MR and VHR maps for the tall tree growth form is further evidenced by the similar spatial distribution of both maps and the ≥0.5 Kappa estimates in the models produced with both resolutions (
Table 3).
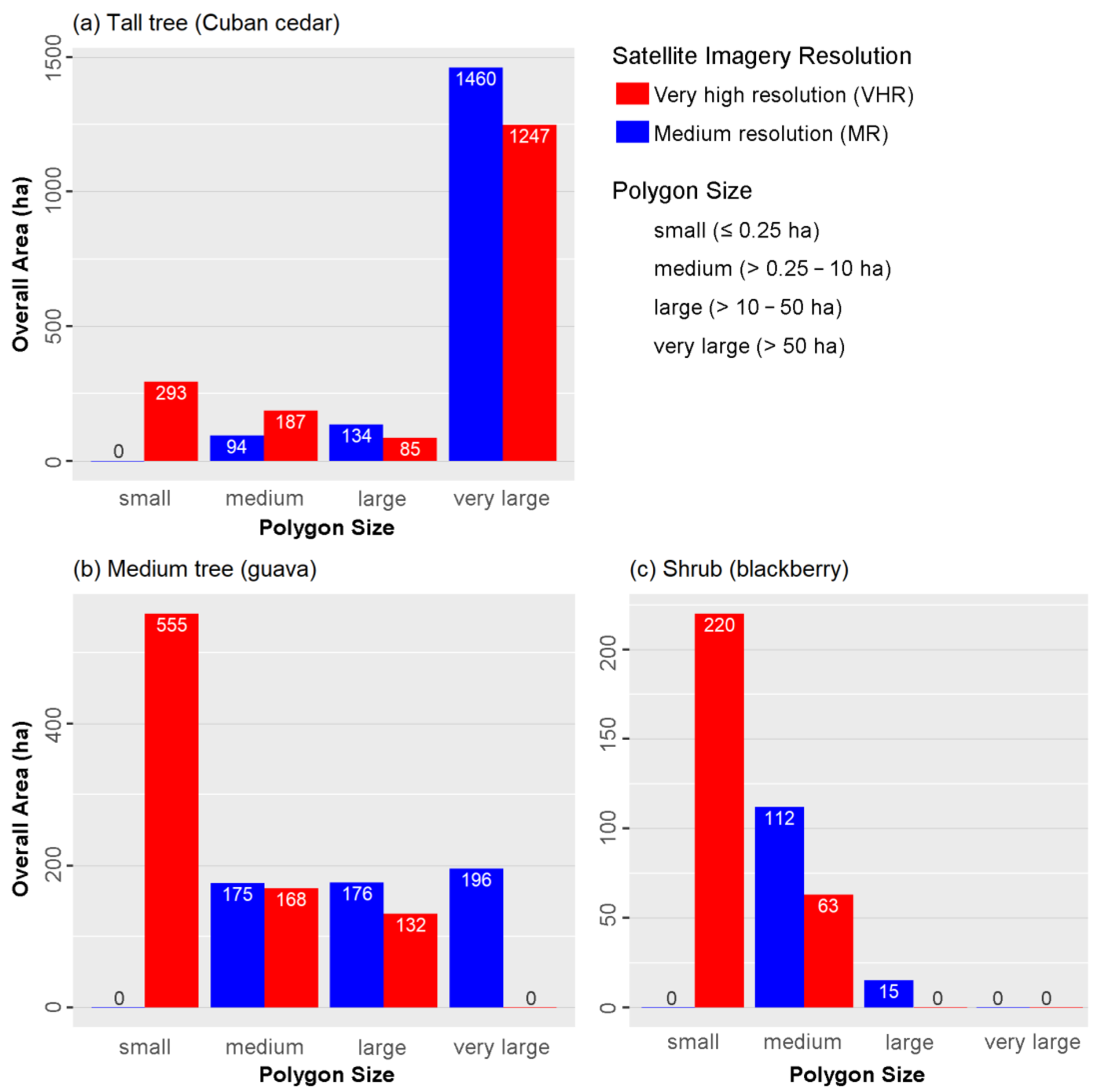
The similarity between the maps of the tall tree growth form (Cuban cedar) produced by VHR and MR (
Figure 2a) was probably due to the fact that most of the space occupied by Cuban cedar consisted of large monospecific stands, which were primarily detected with very large polygons. Only a minor portion was detected with the small polygons exclusively present in VHR maps (
Figure 3a). The high accuracy in detecting the tall tree growth form (Cuban cedar) with MR imagery can be explained by the distinctiveness of this plant species from others. Cuban cedar was the tallest tree in our study area, had the largest crown, and was hardly ever shaded by other vegetation. During the wet season, the crowns were lush green, resulting in a high photosynthetic activity and a mean NDVI of 0.85, the highest among the classes analyzed. The high JMD also evidenced the distinctiveness of Cuban cedar compared to all other classes.
The spatial distributions of the medium tree (guava) and the shrub (blackberry) growth forms were more scattered (
Figure 2b,c). Most of their overall area was detected by small polygons (≤0.25 ha) with VHR imagery, but not with MR imagery (
Figure 3b,c). Similar results were encountered for the mangrove mapping in Galapagos, where MR mapping was unable to detect 60% of the mangrove distribution because 85% of the mangrove patches were smaller than 0.5 ha [
44]. These results highlight the need to use VHR imagery to map small vegetation patches. This is particularly crucial at the early stages of plant invasions, when effective control or even eradication might still be possible [
50,
51].
The medium tree growth form (guava) was the most difficult to discern from other vegetation and its OA was the lowest of the growth forms for both resolutions (76% VHR, 47% MR). We believe that this was due to the difficulty in differentiating guava from other classes in the study area. Indices such as the Normalized Difference Vegetation Index (NDVI) have been used for coarse differentiation between native and non-native vegetation [
21,
52]. However, the mean NDVI obtained for guava in our study was relatively low and similar to other vegetation types, so a clear separation was impossible. Jeffries–Matusita distance (JMD) values for guava, mixed vegetation, Scalesia, guayabillo, and green moss were very low and confirmed the spectral similarity between these classes.
Modeling the distribution of the shrub growth form (blackberry) proved to be challenging because much of the blackberry was hidden in the understory and only visible at the top layer of vegetation as small patches. Additionally, blackberry has a spectral similarity to the native bracken (Pteridium arachnoideum), which makes separating both species difficult. Additional drone footage could be used to help distinguish the two classes and to further improve the model.
Other studies also show that VHR outperforms MR in land cover classification, achieving higher OA and detection of small units [
17,
44,
53]. Satellite imagery with lower spatial resolution is dominated by pixels that present a mixed signature average across multiple objects and, therefore, it is typically used to study ecological systems and map broad vegetation communities from regional to landscape scales [
15,
16]. At a higher spatial resolution, individual objects, such as tree crowns or small patches of plants, are recognizable [
15,
54] and, therefore, it is the preferred resolution for classifying individual species such as invasive species, or for biodiversity assessments in general [
9,
14,
15]. However, using MR to classify invasive species can be a cost-effective way to provide an overview of areas dominated by these species [
21].
Multispectral VHR imagery is not free to use and the costs can be unaffordable when management resources are scarce [
55,
56]. Multispectral MR imagery with global coverage is currently available at no cost, thanks to programs such as the joint NASA/USGS Landsat and the European Space Agency Sentinel. Therefore, MR imagery is an asset for low- to middle-income, but highly biodiverse countries such as Ecuador. Multispectral VHR imagery can also be prohibitive in terms of the increased computer power needed to analyze this type of imagery compared to MR, increasing the cost of the analysis [
46,
57]. Cost is an important factor when choosing the source resolution for mapping. Hopefully, with increasing technological advances, VHR programs will eventually provide free access to their imagery databases for academia and conservation.
One potential limitation of our study is that differences in the mapping results encountered could also have been caused by other factors besides spatial resolution, such as different acquisition dates of the imagery or the classification methods used. However, the acquisition dates for the MR and VHR imagery were very similar for most of the study area. The classification methods used with VHR imagery were similar to those used with MR imagery in that both used an object-based image analysis approach. However, they were different in the statistical classification method applied and in the segmentation methodology. For example, the segmentation methodology applied on VHR imagery produced segments that represented single trees, which would not be a discernable segment unit in MR. While we compared the results obtained with slightly different classification methods, we argue that classification methods are not always directly transferable across spatial resolutions. Therefore, we believe that a comparison of the results obtained with the methodology best suited for each resolution is still very relevant for understanding the strengths and limitations of different spatial resolutions in the mapping of different plant growth forms.
Future studies on the effects of using different spatial resolutions will improve our understanding of the outcomes of vegetation mapping, as will the exploration of different classification methods. Given the promising results obtained in this study, we recommend testing the ability of VHR imagery to detect other important plant species with similar growth forms in Galapagos and expanding the mapping to the rest of the islands in the archipelago, as well as to similar ecosystems around the world.






 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








