
The Benefits of Combining Global and Local Data—A Showcase for Valuation and Mapping of Mangrove Climate Regulation and Food Provisioning Services within a Protected Area in Pará, North Brazil


 ,
,
Abstract
1. Introduction
- How can globally mapped data on regulating ES be refined by taking into account local empirical data and recent modelling approaches?
- How can the value of provisioning services be estimated and mapped combining globally available databases and local empirical data?
2. Materials and Methods
2.1. Case Study Area RESEXM Caeté-Taperaçu
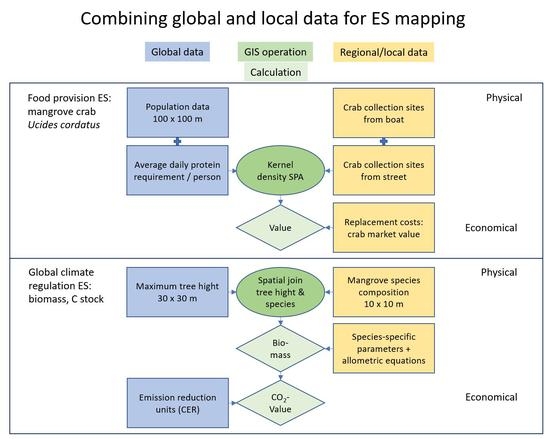
2.2. Valuation Techniques and GIS-Based Mapping
2.2.1. General Overview
- PV (i,N) = Sum of present values with discount rate i for N years
- CFt = Cash flow in year t in US Dollar
- N = Total number of years (10)
- i = 10-year average discount rate (monetary policy interest) (10%: [38])
- t = Number of the year starting in 2019 until 2029 (2020 = year 1 and so on)
2.2.2. Valuation and Mapping of the Food Provisioning ES
- PV = Present value for each grid cell
- Dis_fact = Distribution factor to spatially distribute the whole number of crabs, i.e., subsistence crab demand of the four village communities, to each grid cell of the mangroves within the RESEXM area [28]
- Gc = Grid cell number of the population raster data in the service benefitting area [36]
- Sub_crab = Proportion of population within a population grid cell relying on crabs as a protein source (40%: [6])
- Person = Number of persons per grid cell 2019 in the population raster data [36]
- Prot_requ = Daily human protein requirement in grams (40.23 g: [40])
- Days = Days per year (365)
- i = 10-year average discount rate (monetary policy interest rate) (10%: [38])
- t = Time of the cash flow in years starting in 2019 until 2029 (2020 = year 1 and so on)
2.2.3. Valuation and Mapping of “Carbon Stock for Global Climate Regulation ES”
- AGBSumX = Stand-specific total aboveground biomass in Mg for R. mangle dominated stand (AGBSumRz), A. germinans dominated stand (AGBSumAv) and L. racemosa dominated stand (AGBSumLa) for each grid cell
- DBHX = Species- and stand-specific diameter at breast height in cm for R. mangle (DBHRz), A. germinans (DBHAv) and L. racemosa (DBHLa) [34]
- Rel_stock_CO2 = Releasable amount, total aboveground CO2 in Mg CO2 per grid cell
- AGBSum = Sum of aboveground biomass in Mg per grid cell (obtained from Equations (3) to (5)
- AGBSumtoC = Coefficient to convert from aboveground biomass to aboveground carbon weight (coefficient obtained via email from authors, same parameters used in [18] to value total carbon stocks)
- CtoCO2 = Coefficient to convert carbon weight to CO2 weight [45]
- PV = Present value for each grid cell
- Net_prim_prod_CO2 = Average aboveground net primary productivity of the mangrove ecosystem in Mg CO2 per year and grid cell (2.94 Mg CO2/year and grid cell: [47])
- Rel_stock_CO2 = Releasable amount, total aboveground CO2 in Mg CO2 per grid cell (Assumption: linear decay within 5 years, for year 6 to 10 the whole biomass is decayed and not part of the equation anymore)
- US $/Mg CO2 = Assumed price of one certificate of the certified emission reduction market (CER) in US Dollar, equivalent to one Mg CO2 (USD 5/Mg CO2: [46])
- i = 10-year average discount rate (monetary policy interest) (10%: [38])
- t = Time of the cash flow in years starting in 2019 until 2029 (2020 = year 1 and so on)
3. Results
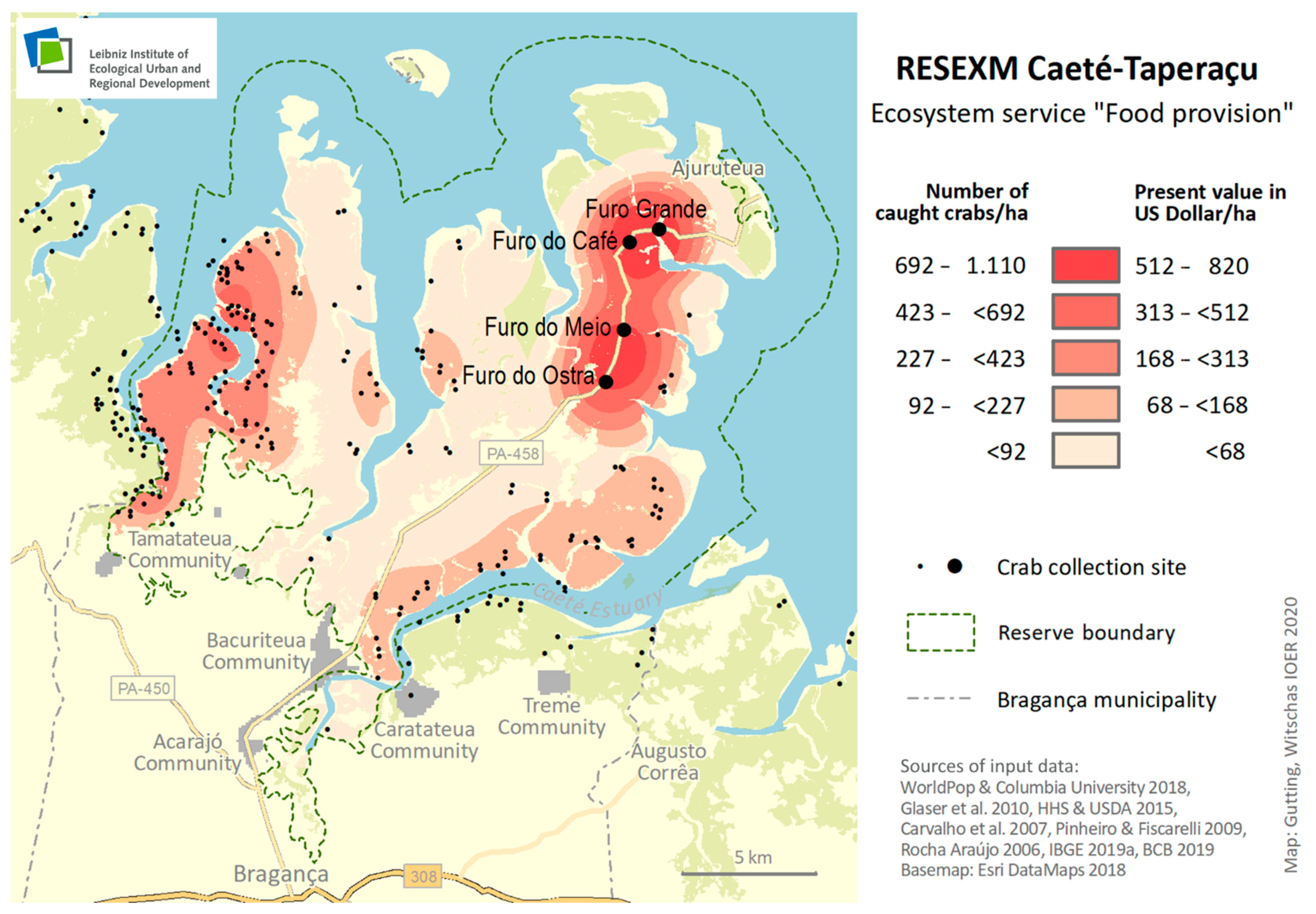
3.1. Food Provisioning ES
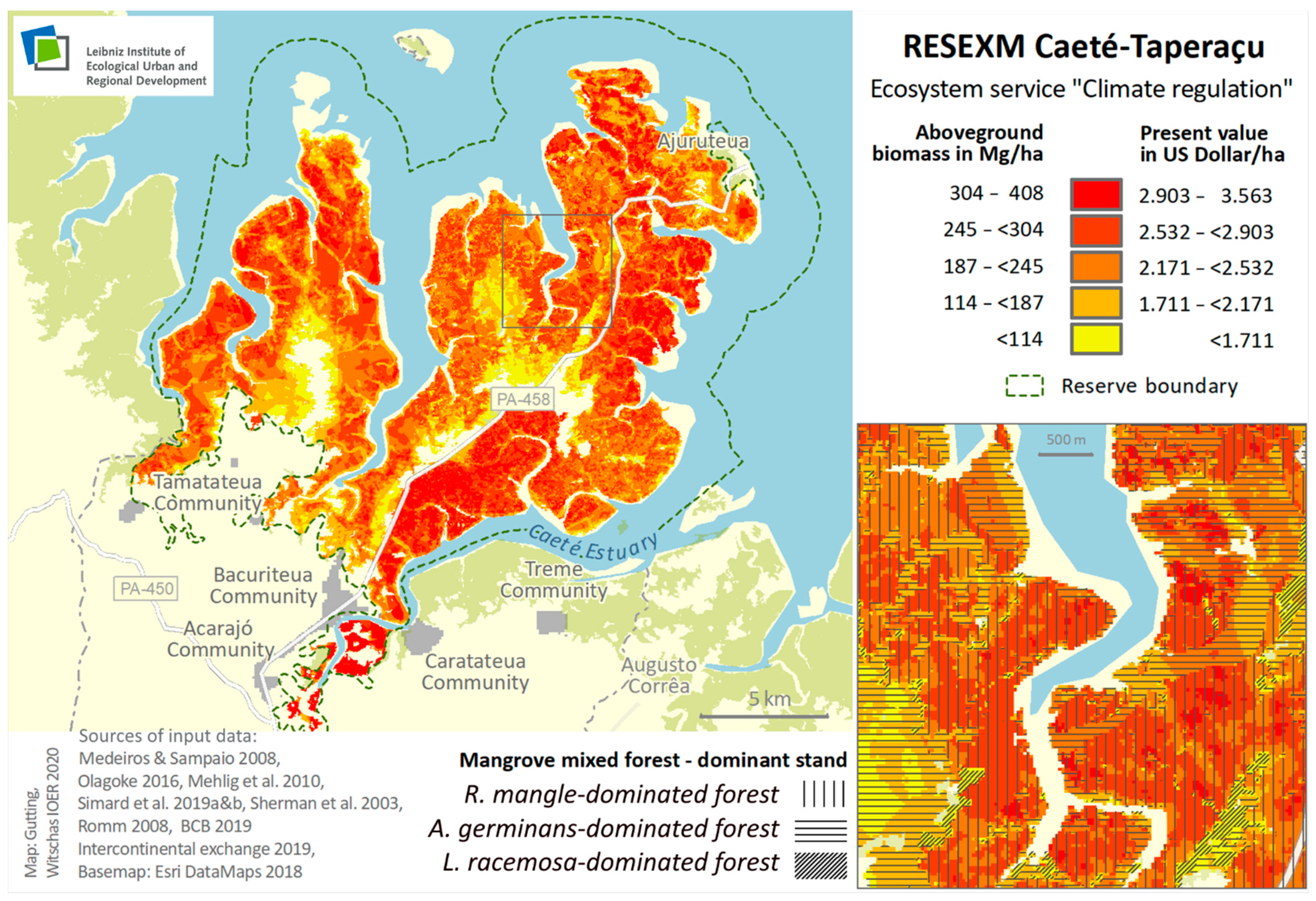
3.2. Carbon Stock for Global Climate Regulation ES
4. Discussion
4.1. General Issue: ES Mapping of Remote Areas Using Local and Worldwide Available Data
4.2. Food Provisioning ES
4.3. Carbon Stock for Global Climate Regulation ES
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Syrbe, R.U.; Grunewald, K. Ecosystem service supply and demand—the challenge to balance spatial mismatches. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2017, 13, 148–161. [Google Scholar] [CrossRef]
- Syrbe, R.U.; Walz, U. Spatial indicators for the assessment of ecosystem services: Providing, benefiting and connecting areas and landscape metrics. Ecol. Indic. 2012, 21, 80–88. [Google Scholar] [CrossRef]
- Atkinson, S.C.; Jupiter, S.D.; Adams, V.M.; Ingram, J.C.; Narayan, S.; Klein, C.J.; Possingham, H.P. Prioritising Mangrove Ecosystem Services Results in Spatially Variable Management Priorities. PLoS ONE 2016, 11, 1–21. [Google Scholar] [CrossRef]
- Cannicci, S.; Burrows, D.; Fratini, S.; Smith, T.J.; Offenberg, J.; Dahdouh-Guebas, F. Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review. Aquat. Bot. 2008, 89, 186–200. [Google Scholar] [CrossRef]
- Fernandes, M.E.B.; Oliveira, F.P.; Eyzaguirre, I.A.L. Mangroves on the Brazilian Amazon Coast: Uses and Rehabilitation. In Threats to Mangrove Forests; Coastal Research Library; Makowski, C., Finkl, C.W., Eds.; Springer: Cham, Switzerland, 2018; Volume 25, pp. 621–635. [Google Scholar] [CrossRef]
- Glaser, M.; Krause, G.; Oliveira, R.S. Fontalvo-Herazo, M. Mangroves and People: A Social-Ecological System. In Mangrove Dynamics and Management in North Brazil; Ecological Studies; Saint-Paul, U., Schneider, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 211, pp. 307–351. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Blaber, S.J.M.; Bouillon, S.; Green, P.; Haywood, M.; Kirton, L.G.; Meynecke, J.O.; Pawlik, J.; Penrose, H.M.; Sasekumar, A.; et al. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat. Bot. 2008, 89, 155–185. [Google Scholar] [CrossRef]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C. The Loss of Species: Mangrove Extinction Risk and Geographic Areas of Global Concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef]
- Worthington, T.A.; Andradi-Brown, D.A.; Bhargava, R.; Buelow, C.; Bunting, P.; Duncan, C.; Fatoyinbo, L.; Friess, D.A.; Goldberg, L.; Hilarides, L.; et al. Harnessing Big Data to Support the Conservation and Rehabilitation of Mangrove Forests Globally. One Earth 2020, 2, 429–443. [Google Scholar] [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob. Chang. Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.E.; Casey, D. Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Sandilyan, S.; Kathiresan, K. Mangrove conservation: A global perspective. Biodivers. Conserv. 2012, 21, 3523–3542. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Himes-Cornell, A.; Pendleton, L.; Atiyah, P. Valuing ecosystem services from blue forests: A systematic review of the valuation of salt marshes, sea grass beds and mangrove forests. Ecosyst. Serv. 2018, 30, 36–48. [Google Scholar] [CrossRef]
- Mehvar, S.; Filatova, T.; Dastgheib, A.; De Ruyter van Steveninck, E.; Ranasinghe, R. Quantifying economic value of coastal ecosystem services: A review. J. Mar. Sci. Eng. 2018, 6, 5. [Google Scholar] [CrossRef]
- Vo, Q.T.; Kuenzer, C.; Vo, Q.M.; Moder, F.; Oppelt, N. Review of valuation methods for mangrove ecosystem services. Ecol. Indic. 2012, 23, 431–446. [Google Scholar] [CrossRef]
- Sanderman, J. Global mangrove soil carbon: Dataset and spatial maps. Harv. Dataverse 2017, 4. [Google Scholar] [CrossRef]
- Simard, M.; Fatoyinbo, L.; Smetanka, C.; Rivera-monroy, V.H.; Castaneda-Moya, E.; Thomas, N.; Van der Stocken, T. Mangrove canopy height globally related to precipitation, temperature and cyclone frequency. Nat. Geosci. 2019, 12, 40–45. [Google Scholar] [CrossRef]
- Simard, M.; Fatoyinbo, L.; Smetanka, C.; Rivera-monroy, V.H.; Castaneda-Moya, E.; Thomas, N.; Van der Stocken, T. Global Mangrove Distribution, Aboveground Biomass, and Canopy Height; ORNL DAAC: Oak Ridge, TN, USA, 2019. [CrossRef]
- Alongi, D.M. Present State and Future of the World’s Mangrove Forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Syrbe, R.U.; Schröder, M.; Grunewald, K.; Walz, U.; Burkhard, B. What to map? In Mapping Ecosystem Services; Burkhard, B., Maes, J., Eds.; Pensoft Publishers: Sofia, Bulgaria, 2017; pp. 151–158. [Google Scholar] [CrossRef]
- ICMBio (Instituto Chico Mendes de Conservação da Biodiversidade). Plano de manejo da reserve extrativista marinha de Caeté-Taperaçu. In Portuguese, English Title: Caete-Taperacu Marine Reserve Management Plan PA; 2012; p. 339. Available online: https://repositorio.unb.br/handle/10482/14358 (accessed on 1 January 2021).
- Ruiz-Pérez, M.; Almeida, M.; Dewi, S.; Costa, E.M.; Pantoja, M.C.; Puntodewo, A.; de Postigo, A.A.; de Andrade, A.G. Conservation and development in Amazonian extractive reserves: The case of Alto Juruá. Ambio 2005, 34, 218–223. [Google Scholar] [CrossRef]
- Glaser, M.; Krause, G. Integrated coastal management in federal Brazil: Institutional sectoral and legal structures and the limits of participative planning. (In German, title: Integriertes Küstenmanagement im föderalen Brasilien: Institutionelle, sektorale und legale Strukturen und die Grenzen der partizipativen Planung). In Kueste, Oekologie und Mensch—Integriertes Kuestenmanagement als Instrument nachhaltiger Entwicklung; Glaeser, B., Ed.; Humanoekologie: Oekom, Munchen, 2005; Volume 2, pp. 37–54. [Google Scholar]
- Saint-Paul, U.; Schneider, H. (Eds.) Mangrove Dynamics and Management in North Brazil, Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2010; Volume 211. [Google Scholar] [CrossRef]
- Bunting, P.; Rosenqvist, A.; Lucas, R.; Rebelo, L.-M.; Hilarides, L.; Thomas, N.; Hardy, A.; Itoh, T.; Shimada, M.; Finlayson, C.M. The Global Mangrove Watch—A New 2010 Global Baseline of Mangrove Extent. Remote Sens. 2018, 10, 1669. [Google Scholar] [CrossRef]
- UNEP-WCMC; IUCN; Protected Planet. The World Database on Protected Areas (WDPA); UNEP-WCMC and IUCN: Cambridge, UK, 2019. [Google Scholar]
- Rocha Araújo, A.R. Fishery Statistics and Commercialization of the mangrove crab, Ucides cordatus (L.) in Bragança, Pará, Brazil. Dissertation/Ph.D. Thesis, Universityof Bremen, Bremen, Germany, 2006. [Google Scholar]
- European Commission. Mapping and Assessment of Ecosystems and their Services—An Analytical Framework for Mapping and Assessment of Ecosystem Condition in EU. Discussion Paper. 2018, p. 60. Available online: https://ec.europa.eu/environment/nature/knowledge/ecosystem_assessment/pdf/2ndMAESWorkingPaper.pdf (accessed on 1 January 2021).
- European Environment Agency CICES—Towards a Common Classification of Ecosystem Services. 2020. Available online: https://cices.eu/ (accessed on 17 March 2021).
- Diele, K.; Koch, V.; Saint-Paul, U. Population structure, catch composition and CPUE of the artisanally harvested mangrove crab Ucides cordatus (Ocypodidae) in the Caeté estuary, North Brazil: Indications for overfishing? Aquat. Living Resour. 2005, 18, 169–178. [Google Scholar] [CrossRef]
- Sullivan, M.J.P.; Talbot, J.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Begne, S.K.; Chave, J.; Cuni-Sanchez, A.; Hubau, W.; Lopez-Gonzalez, G.; et al. Diversity and carbon storage across the tropical forest biome. Sci. Rep. 2017, 7, 39102. [Google Scholar] [CrossRef]
- Medeiros, T.C.C.; Sampaio, E.V.S.B. Allometry of aboveground biomasses in mangrove species in Itamaracá, Pernambuco, Brazil. Wetl. Ecol. Manag. 2008, 16, 323–330. [Google Scholar] [CrossRef]
- Mehlig, U.; Menezes, M.P.M.; Reise, A.; Schories, D.; Medina, E. Mangrove Vegetation of the Caeté Estuary. In Mangrove Dynamics and Management in North Brazil; Saint-Paul, U., Schneider, H., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2010; Volume 211, pp. 71–107. [Google Scholar] [CrossRef]
- Olagoke, A. Towards a Better Characterization of Morphological Plasticity and Biomass Partitioning of Trees in Structural Dynamics of Mangrove Forests. Dissertation/Ph.D. Thesis, Technical University of Dresden, Dresden, Germany, 2016. [Google Scholar]
- WorldPop (School of Geography and Environmental Science, University of Southampton; Department of Geography and Geosciences, University of Louisville; Departement de Geographie, Universite de Namur) and Center for International Earth Science Information Network (CIESIN), Columbia University 2018, Global High Resolution Population Denominators Project—Funded by The Bill and Melinda Gates Foundation (OPP1134076), Internet Database. Available online: https://www.worldpop.org/geodata/summary?id=6414 (accessed on 17 April 2021).
- Kruschwitz, L.; Loeffler, A. Discounted Cash Flow: A Theory of the Valuation of Firms; Wiley: Hoboken, NJ, USA, 2006. [Google Scholar]
- BCB (Banco Central do Brasil) Interest rates—Actual SELIC rate % p.y. 2019. Available online: https://www.bcb.gov.br/en/legacy?url=https:%2F%2Fwww.bcb.gov.br%2FPec%2FCopom%2FIngl%2FtaxaSelic-i.asp#notas (accessed on 17 March 2020).
- IBGE (Instituto Brasileiro de Geografia e Estatistica) Continuous National Household Sample Survey—Continuous PNAD. 2019. Available online: https://www.ibge.gov.br/en/statistics/social/housing/18083-annual-dissemination-pnadc3.html?edicao=24560&t=resultados (accessed on 17 March 2020).
- HHS (U.S. Department of Health and Human Services); USDA (U.S. Department of Agriculture). 2015–2020 Dietary Guidelines for Americans, 8th ed.; U.S. Department of Health and Human Services: Rockville, MD, USA, 2015; 125p.
- Carvalho, A.F.U.; Farias, D.F.; Barroso, C.X.; Sombra, C.M.L.; Silvino, A.S.; Menezes, M.O.T.; Soares, M.O.; Fernandes, D.A.O.; Gouveia, S.T. Nutritive value of three organisms from mangrove ecosystem: Ucides cordatus (Linnaeus, 1763), Mytella sp. (Soot-Ryen, 1955) and Crassostrea rhizophorae (Guilding, 1828). Braz. J. Biol. 2007, 67, 787–788. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pinheiro, M.A.A.; Fiscarelli, A.G. Length-weight relationship and condition factor of the mangrove crab Ucides cordatus (Linnaeus, 1763) (Crustacea, Brachyura, Ucididae). Braz. Arch. Biol. Technol. 2009, 52, 397–406. [Google Scholar] [CrossRef]
- Pascal, N.; Bulu, M. Economic Valuation of Mangrove Ecosystem Services in Vanuatu: Case Study of Crab Bay (Malekula is.) and Eratap (Efate is.); Technical Report for Project Mescal: Suva, Fiji, 2013. [Google Scholar]
- IBGE (Instituto Brasileiro de Geografia e Estatistica) Índice Nacional de Preços ao Consumidor Amplo—IPCA. 2019. Available online: https://www.ibge.gov.br/estatisticas/economicas/precos-e-custos/9256-indice-nacional-de-precos-ao-consumidor-amplo.html?=&t=o-que-e (accessed on 17 March 2020).
- Romm, J. The biggest source of mistake: C vs. CO2. 2008. Available online: https://archive.thinkprogress.org/the-biggest-source-of-mistakes-c-vs-CO2-c0b077313b/ (accessed on 17 March 2020).
- Intercontinental exchange CER Futures. 2019. Available online: https://www.theice.com/products/814666/CER-Futures/data?marketId=5236635&span=2 (accessed on 17 March 2020).
- Sherman, R.; Fahey, T.; Martinez, P. Spatial Patterns of Biomass and Aboveground Net Primary Productivity in a Mangrove Ecosystem in the Dominican Republic. Ecosystems 2003, 6, 384–398. [Google Scholar] [CrossRef]
- Quadros, A.F.; Zimmer, M. Dataset of “true mangroves” plant species traits. Biodivers. Data J. 2017, 5, e22089. [Google Scholar] [CrossRef]
- Gardunho, D.C.L. Estimativa de biomassa e carbono acima do solo das especies arbóreas dominantes nas florestas de mangue da peninsula de Ajuruteua, nordeste do Pará, Costa Amazônica Brasileira 2018. Ph.D. Thesis, University of Bragança, Bragança, Brazil. Unpublished Dissertation.
- Herault, B.; Beauchene, J.; Muller, F.; Wagner, F.; Baraloto, C.; Blanc, L.; Martin, J.M. Modeling decay rates of dead wood in a neotropical forest. Oecologia 2010, 164, 243–251. [Google Scholar] [CrossRef]
- Pingoud, K.; Skog, K.E.; Martino, D.L.; Tonosaki, M.; Xiaoquan, Z. Chapter 12—Harvested wood products. In IPCC—National Guidelines for Greenhouse Gas Inventories; Eggleston, S., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Eds.; The Institute for Global Environmental Strategies (IGES) for the IPCC 2006: Hayama, Japan, 2006; Volume 4, Available online: https://www.ipcc-nggip.iges.or.jp/public/2006gl/pdf/4_Volume4/V4_12_Ch12_HWP.pdf (accessed on 1 January 2021).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecosystem Service Category | Ecosystem Service |
|---|---|
| Provisioning | Food from wild animals (e.g., crab, fish, honey) (Highest values 9 × 5) |
| Timber and fibers from trees or other plants (Highest values 6 × 5) | |
| Water for general use (Highest values 6 × 5) | |
| Regulating | Nursery populations and habitats (Highest values 9 × 5) |
| Flood protection/Storm protection and air purification/ CO2-storage and global climate regulation (Highest values 6 × 5) | |
| Coastal stabilization and control of erosion rates (Highest values 6 × 5) | |
| Cultural | Aesthetic interactions (Highest values 7 × 5) |
| Spiritual/existence (Highest values 4 × 5) | |
| Educational interactions (Highest values 7 × 5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutting, R.; Syrbe, R.-U.; Grunewald, K.; Mehlig, U.; Helfer, V.; Zimmer, M. The Benefits of Combining Global and Local Data—A Showcase for Valuation and Mapping of Mangrove Climate Regulation and Food Provisioning Services within a Protected Area in Pará, North Brazil. Land 2021, 10, 432. https://doi.org/10.3390/land10040432
Gutting R, Syrbe R-U, Grunewald K, Mehlig U, Helfer V, Zimmer M. The Benefits of Combining Global and Local Data—A Showcase for Valuation and Mapping of Mangrove Climate Regulation and Food Provisioning Services within a Protected Area in Pará, North Brazil. Land. 2021; 10(4):432. https://doi.org/10.3390/land10040432
Chicago/Turabian StyleGutting, Robin, Ralf-Uwe Syrbe, Karsten Grunewald, Ulf Mehlig, Véronique Helfer, and Martin Zimmer. 2021. "The Benefits of Combining Global and Local Data—A Showcase for Valuation and Mapping of Mangrove Climate Regulation and Food Provisioning Services within a Protected Area in Pará, North Brazil" Land 10, no. 4: 432. https://doi.org/10.3390/land10040432
APA StyleGutting, R., Syrbe, R.-U., Grunewald, K., Mehlig, U., Helfer, V., & Zimmer, M. (2021). The Benefits of Combining Global and Local Data—A Showcase for Valuation and Mapping of Mangrove Climate Regulation and Food Provisioning Services within a Protected Area in Pará, North Brazil. Land, 10(4), 432. https://doi.org/10.3390/land10040432