
A Review of Eco-Corona Formation on Micro/Nanoplastics and Its Effects on Stability, Bioavailability, and Toxicity

Highlights
- Eco-corona formation significantly regulated the hetero-aggregation dynamics and colloidal stability of M/NPs.
- The eco-corona modulated the bioaccumulation and bioavailability of M/NPs across various aquatic organisms.
- The eco-corona reshaped the ecotoxicity of M/NPs, specifically influencing growth inhibition, oxidative stress, and genotoxicity.
- Humic acid corona played a pivotal role in alleviating the toxicity of M/NPs.
Abstract
1. Introduction
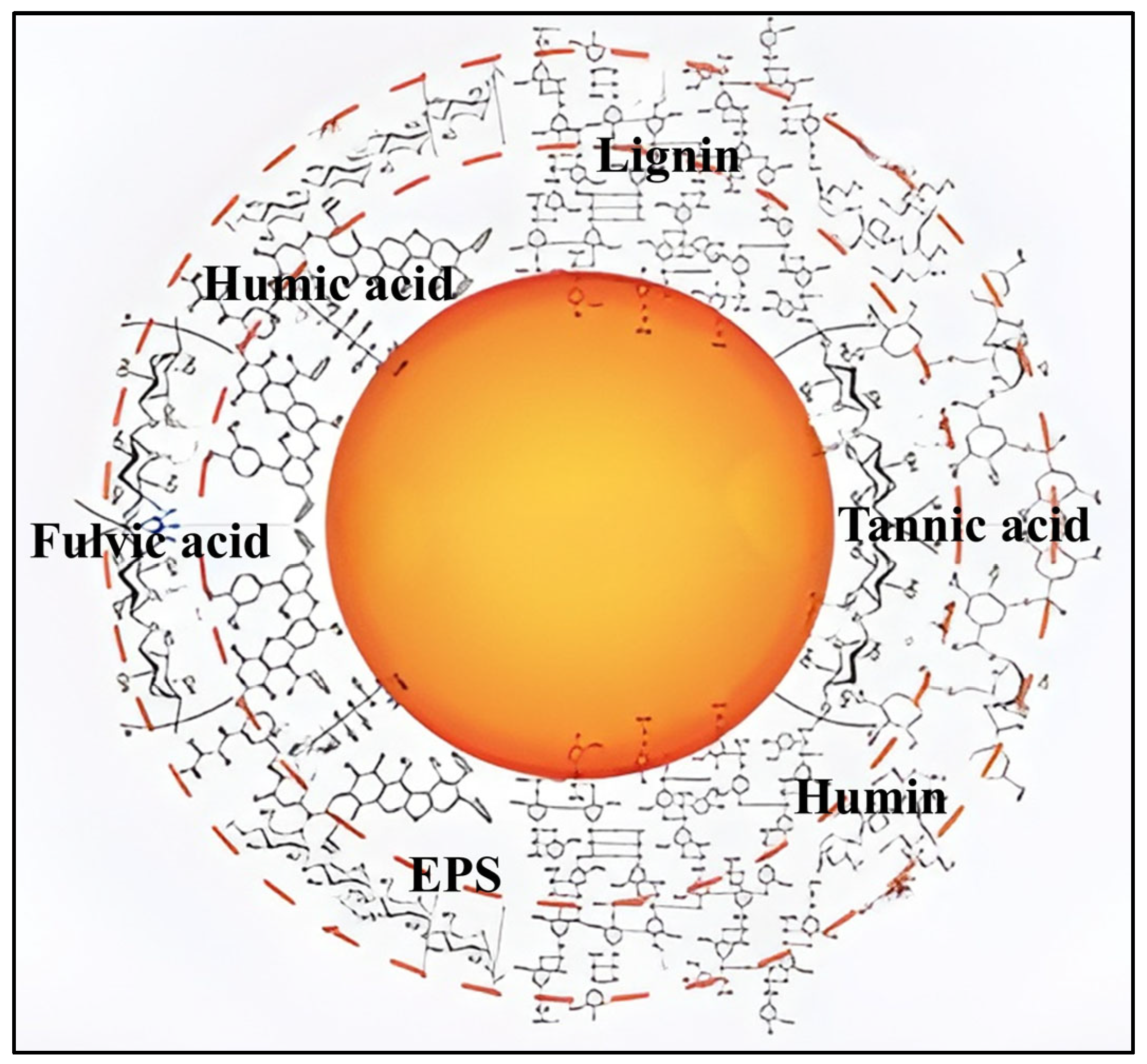
2. Physicochemical Properties of Typical Natural Organic Macromolecules
2.1. Natural Organic Matter
2.2. Extracellular Polymeric Substances
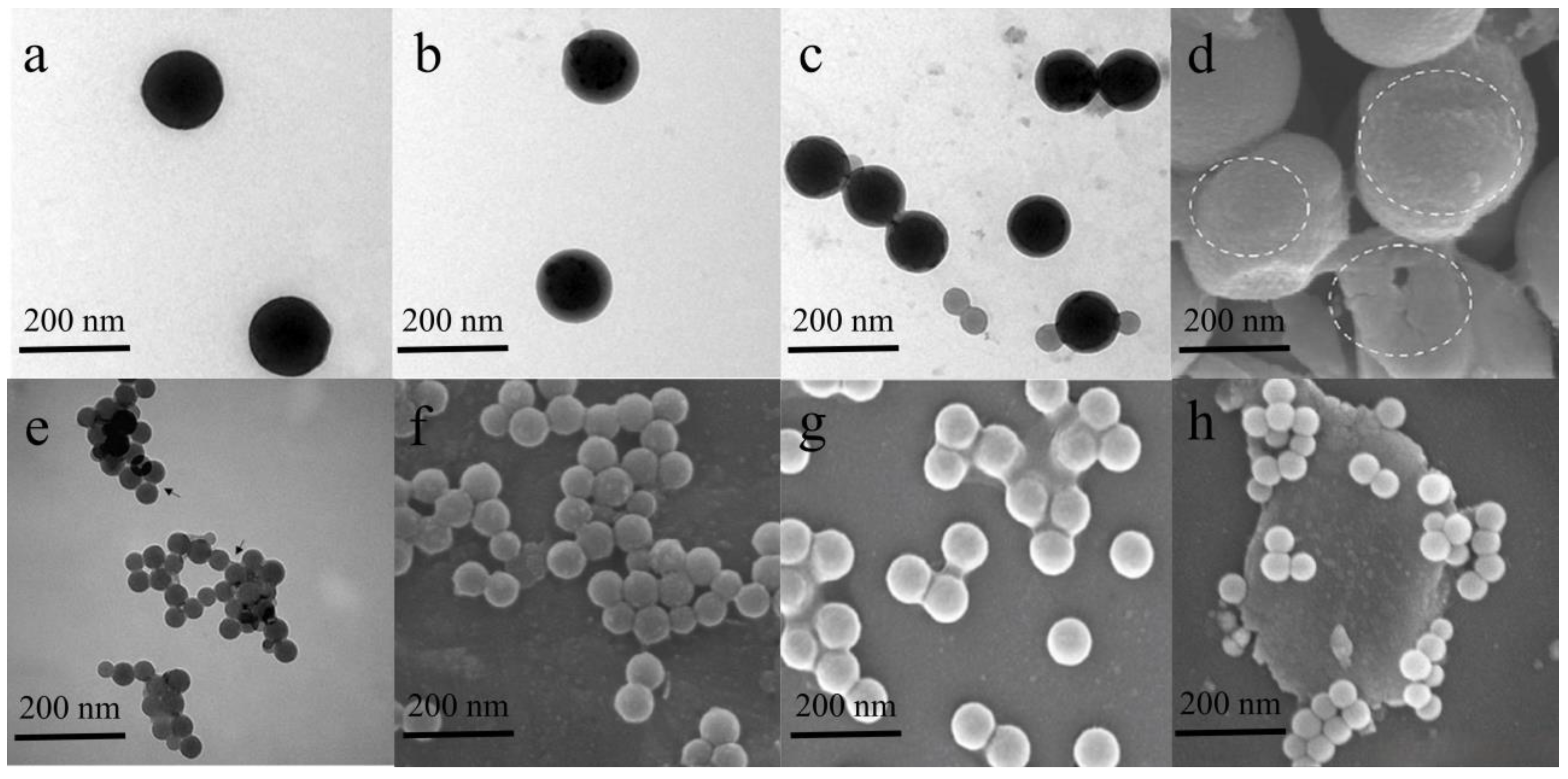
3. Formation Mechanism of the Eco-Corona
3.1. Electrostatic and Hydrophobic Interactions
3.2. Hydrogen Bonding and van der Waals Forces
4. Effects of Eco-Corona Formation on the Hetero-Aggregation Stability of M/NPs
4.1. Applications and Limitations of DLVO and XDLVO Theories
4.2. Regulatory Mechanisms of Eco-Corona on Hetero-Aggregation of M/NPs
5. Effects of Eco-Corona on the Bioavailability of M/NPs
5.1. Bioaccumulation
5.1.1. Algae
5.1.2. Invertebrates
5.1.3. Fish
5.2. Biological Effects
5.2.1. Growth Inhibition and Oxidative Stress
5.2.2. Photoaging M/NPs
5.3. Molecular Mechanism
6. Perspectives
- (1)
- The properties of the environmental medium significantly determine the aggregation state, stability, and bioavailability of contaminants in aquatic environments. For instance, higher ionic strength could reduce the electrostatic repulsion of TiO2 NPs and increase particle aggregation [162]. Higher temperatures not only facilitate the formation of EPS corona on Ag NPs surfaces but also significantly enhance the structural stability of the EPS-Ag NPs composite system [163]. So far, only the effects of salinity, pH, and different electrolytes on the formation of eco-corona on the surface of M/NPs have been studied, while the role of other influencing factors (such as temperature, conductivity, ion valence, etc.) deserves further investigation.
- (2)
- In aquatic environments, the interaction between M/NPs and co-pollutants could influence the uptake and accumulation of plastics or contaminants in aquatic organisms. Current studies have demonstrated that the formation of eco-corona altered the adsorption and desorption behavior of M/NPs toward heavy metal ions [164]. However, little is known about the effects of eco-corona on the interaction of M/NPs with other types of pollutants (e.g., metal nanoparticles, persistent organic pollutants, pharmaceutical pollutants), and further investigation of potential interfacial reactions is needed.
- (3)
- When M/NPs coated with eco-corona migrate from the external water environment into organisms or cells, the affinity of ecological macromolecules to the M/NPs may change with the changes due to the environment [165]. If biomolecules such as lipids, proteins, and nucleic acids within the organism exhibit a higher affinity for M/NPs, these biomolecules can be exchanged with the adsorbed components on the eco-corona [166]. However, it remains unclear which specific biomolecules would cover or replace the ecological macromolecules adsorbed on the surface of M/NPs, thereby forming a new eco-corona. The consequences of biomolecule replacement and its interaction process are also unclear. Therefore, further research is needed on the fate and transformation of the corona in cells or organisms, which is crucial for evaluating the biological effects of M/NPs.
- (4)
- Currently, knowledge of the biological effects and mechanisms of eco-corona is relatively limited. Studies on the biological effects of M/NPs by eco-corona have mainly centered on membrane adhesion, cellular uptake, growth inhibition, and oxidative stress. However, the impact of eco-corona on genotoxicity and reproductive toxicity has only recently started to be investigated. Therefore, beyond specific toxic endpoints such as DNA damage and apoptosis, the mechanisms of intracellular responses, including enzyme inactivation, cell differentiation, epigenetic modifications, and gene mutations, should be thoroughly explored.
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Plastics Europe. Plastics—The Fast Facts. 2023. Available online: https://plasticseurope.org/knowledge-hub/plastics-the-fast-facts-2023/ (accessed on 23 January 2025).
- Gaylarde, C.C.; Neto, J.A.B.; da Fonseca, E.M. Nanoplastics in aquatic systems-are they more hazardous than microplastics? Environ. Pollut. 2021, 272, 115950. [Google Scholar] [CrossRef]
- Singh, N.; Abdullah, M.M.; Ma, X.; Sharma, V.K. Microplastics and nanoplastics in the soil-plant nexus: Sources, uptake, and toxicity. Crit. Rev. Environ. Sci. Technol. 2023, 53, 1613–1642. [Google Scholar] [CrossRef]
- Gigault, J.; El Hadri, H.; Nguyen, B.; Grassl, B.; Rowenczyk, L.; Tufenkji, N.; Feng, S.; Wiesner, M. Nanoplastics are neither microplastics nor engineered nanoparticles. Nat. Nanotechnol. 2021, 16, 501–507. [Google Scholar] [CrossRef]
- Cai, Z.; Li, M.; Zhu, Z.; Wang, X.; Huang, Y.; Li, T.; Gong, H.; Yan, M. Biological degradation of plastics and microplastics: A recent perspective on associated mechanisms and influencing factors. Microorganisms 2023, 11, 1661. [Google Scholar] [CrossRef]
- Jaiswal, S.; Sharma, B.; Shukla, P. Integrated approaches in microbial degradation of plastics. Environ. Technol. Innov. 2020, 17, 100567. [Google Scholar] [CrossRef]
- Susanti, N.K.Y.; Mardiastuti, A.; Wardiatno, Y. Microplastics and the impact of plastic on wildlife: A literature review. IOP Conf. Ser. Earth Environ. Sci. 2020, 528, 12013. [Google Scholar] [CrossRef]
- Liu, Z.; Malinowski, C.R.; Sepúlveda, M.S. Emerging trends in nanoparticle toxicity and the significance of using Daphnia as a model organism. Chemosphere 2022, 291, 132941. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Y.; Pérez, E.; Jiang, Q.; Chen, Q.; Jiao, Y.; Huang, Y.; Yang, Y.; Zhao, Y. Polystyrene nanoplastic induces oxidative stress, immune defense, and glycometabolism change in Daphnia pulex: Application of transcriptome profiling in risk assessment of nanoplastics. J. Hazard. Mater. 2021, 402, 123778. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Chou, P.I.; Liu, J.; Zhu, Y.; Jun, Y.S. Oxidative roles of polystyrene-based nanoplastics in inducing manganese oxide formation under light illumination. ACS Nano 2022, 16, 20238–20250. [Google Scholar] [CrossRef]
- Rafa, N.; Ahmed, B.; Zohora, F.; Bakya, J.; Ahmed, S.; Ahmed, S.F.; Mofijur, M.; Chowdhury, A.; Almomani, F. Microplastics as carriers of toxic pollutants: Source, transport, and toxicological effects. Environ. Pollut. 2024, 343, 123190. [Google Scholar] [CrossRef]
- Li, X.; Luo, J.; Zeng, H.; Yang, X.; Hou, X.; Lu, X. Preferential adsorption of medium molecular weight proteins in extracellular polymeric substance alleviates toxicity of small-sized microplastics to Skeletonema costatum. J. Hazard. Mater. 2024, 476, 135034. [Google Scholar] [CrossRef]
- Bradford, S.A.; Sasidharan, S.; Kim, H.; Gomez-Flores, A.; Li, T.; Shen, C. Colloid interaction energies for surfaces with steric effects and incompressible and/or compressible roughness. Langmuir 2023, 37, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Flores, A.; Bradford, S.A.; Hong, G.; Kim, H. Statistical analysis, machine learning modeling, and text analytics of aggregation attachment efficiency: Mono and binary particle systems. J Hazard Mater. 2023, 454, 131482. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Flores, A.; Bradford, S.A.; Cai, L.; Urík, M.; Kim, H. Prediction of attachment efficiency using machine learning on a comprehensive database and its validation. Water Res. 2023, 229, 119429. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Cai, W.; Xia, J.; Sheng, L.; Wang, W.; Liu, H. Aggregation kinetics of fragmental PET nanoplastics in aqueous environment: Complex roles of electrolytes, pH and humic acid. Environ. Pollut. 2021, 268, 115828. [Google Scholar] [CrossRef]
- Liu, S.B.; Zhang, X.R.; Zeng, K.; He, C.T.; Huang, Y.C.; Xin, G.R.; Huang, X.C. Insights into eco-corona formation and its role in the biological effects of nanomaterials from a molecular mechanisms perspective. Sci. Total Environ. 2023, 858, 159867. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, M.; Zhao, J.; Cao, Z.; Zou, W.; Zhou, Q. Photoaging enhanced the adverse effects of polyamide microplastics on the growth, intestinal health, and lipid absorption in developing zebrafish. Environ. Int. 2022, 158, 106922. [Google Scholar] [CrossRef]
- Liu, G.; Jiang, R.; You, J.; Muir, D.C.G.; Zeng, E.Y. Microplastic impacts on microalgae growth: Effects of size and humic acid. Environ. Sci. Technol. 2020, 54, 1782–1789. [Google Scholar] [CrossRef]
- Ramsperger, A.F.R.M.; Narayana, V.K.B.; Gross, W.; Mohanraj, J.; Thelakkat, M.; Greiner, A.; Schmalz, H.; Kress, H.; Laforsch, C. Environmental exposure enhances the internalization of microplastic particles into cells. Sci. Adv. 2020, 6, eabd1211. [Google Scholar] [CrossRef]
- Feng, H.; Liang, Y.N.; Hu, X. Natural organic matter (NOM), an underexplored resource for environmental conservation and remediation. Mater. Today Sustain. 2022, 19, 100159. [Google Scholar] [CrossRef]
- Junaid, M.; Wang, J. Interaction of micro(nano)plastics with extracellular and intracellular biomolecules in the freshwater environment. Crit. Rev. Environ. Sci. Technol. 2021, 52, 4241–4265. [Google Scholar] [CrossRef]
- Sharma, V.K.; Filip, J.; Zboril, R.; Varma, R.S. Natural inorganic nanoparticles–formation, fate, and toxicity in the environment. Chem. Soc. Rev. 2015, 44, 8410–8423. [Google Scholar] [CrossRef] [PubMed]
- Hashino, M.; Katagiri, T.; Kubota, N.; Ohmukai, Y.; Maruyama, T.; Matsuyama, H. Effect of membrane surface morphology on membrane fouling with sodium alginate. J. Membr. Sci. 2011, 366, 258–265. [Google Scholar] [CrossRef]
- Sharma, A.; Anthal, R. Humic substances in aquatic ecosystems: A review. Int. J. Innov. Res. Sci. Eng. Technol. 2016, 5, 18462–18470. [Google Scholar]
- Glover, C.N.; Pane, E.F.; Wood, C.M. Humic substances influence sodium metabolism in the freshwater crustacean Daphnia magna. Physiol. Biochem. Zool. 2015, 78, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Ampong, K.; Thilakaranthna, M.S.; Gorim, L.Y. Understanding the role of humic acids on crop performance and soil health. Front Agronom. 2022, 4, 848621. [Google Scholar] [CrossRef]
- Zhao, T.; Fang, M.; Tang, Z.; Zhao, X.; Xie, F.; Wu, F.; Giesy, J.P. Effects of fulvic acid on aggregation, sedimentation, and adsorption of Fe3O4 magnetic nanoparticles. Environ. Sci. Pollut. Res. 2019, 26, 21463–21474. [Google Scholar] [CrossRef]
- Siddiqui, V.U.; Ilyas, R.A.; Sapuan, S.M.; Hamid, N.H.A.; Khoo, P.S.; Chowdhury, A.; Atikah, M.S.N.; Rani, M.S.A.; Asyraf, M.R.M. Alginate-based materials as adsorbent for sustainable water treatment. Int. J. Biol. Macromol. 2025, 298, 139946. [Google Scholar] [CrossRef]
- Siddharth, T.; Sridhar, P.; Vinila, V.; Tyagi, R.D. Environmental applications of microbial extracellular polymeric substance (EPS): A review. J. Environ. Manag. 2021, 287, 112307. [Google Scholar] [CrossRef]
- Nasser, F.; Lynch, I. Secreted protein eco-corona mediates uptake and impacts of polystyrene nanoparticles on Daphnia magna. J. Proteome. 2016, 137, 45–51. [Google Scholar] [CrossRef]
- Ye, T.; Yang, A.; Wang, Y.; Song, N.; Wang, P.; Xu, H. Changes of the physicochemical properties of extracellular polymeric substances (EPS) from Microcystis aeruginosa in response to microplastics. Environ. Pollut. 2022, 315, 120354. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshanee, M.; Das, S. Bacterial extracellular polymeric substances: Biosynthesis and interaction with environmental pollutants. Chemosphere 2023, 332, 138876. [Google Scholar]
- Tang, W.; Wu, M.; Lou, W.; Yang, C. Role of extracellular polymeric substances and enhanced performance for biological removal of carbonaceous organic matters and ammonia from wastewater with high salinity and low nutrient concentrations. Bioresour. Technol. 2021, 326, 124764. [Google Scholar] [CrossRef]
- Wang, S.; Huang, X.; Liu, L.; Shen, Y.; Yan, P.; Chen, Y.; Guo, J.; Fang, F. Understanding the mechanism in aggregation ability between aerobic and anammox granular sludge from the perspective of exopolysaccharides. J. Water Process Eng. 2020, 38, 101629. [Google Scholar] [CrossRef]
- Hong, P.N.; Taing, C.; Phan, P.T.; Honda, R. Polarity-molecular weight profile of extracellular polymeric substances in a membrane bioreactor: Comparison between bulk sludge and cake layers. J. Water Environ. Technol. 2018, 16, 40–53. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, Z.; Zhou, J.; Tang, J.; Yang, C.; Chen, C.; Huang, W.; Dang, Z. Influence of environmental and biological macromolecules on aggregation kinetics of nanoplastics in aquatic systems. Water Res. 2020, 186, 116316. [Google Scholar] [CrossRef] [PubMed]
- Messina, C.M.; Faggio, C.; Laudicella, V.A.; Sanfilippo, M.; Trischitta, F.; Santulli, A. Effect of sodium dodecyl sulfate (SDS) on stress response in the Mediterranean mussel (Mytilus galloprovincialis): Regulatory volume decrease (Rvd) and modulation of biochemical markers related to oxidative stress. Aquat. Toxicol. 2014, 157, 94–100. [Google Scholar] [CrossRef]
- Liu, L.; Ma, Y.; Xu, Y.; Liu, B.; Wang, C.; Feng, J.; Li, M.; Yin, H.; Sun, L.; Li, P.; et al. Mechanisms of eco-corona effects on micro(nano)plastics in marine medaka: Insights into translocation, immunity, and energy metabolism. J. Hazard. Mater. 2024, 480, 136236. [Google Scholar] [CrossRef]
- Schmidt, J.R.; Wilhelm, S.W.; Boyer, G.L. The Fate of Microcystins in the Environment and challenges for monitoring. Toxins. 2014, 6, 3354. [Google Scholar] [CrossRef] [PubMed]
- Chetwynd, A.J.; Zhang, W.; Thorn, J.A.; Lynch, I.; Ramautar, R. The nanomaterial metabolite corona determined using a quantitative metabolomics approach: A pilot study. Small 2020, 16, 2000295. [Google Scholar] [CrossRef]
- Schefer, R.B.; Armanious, A.; Mitrano, D.M. Eco-corona formation on plastics: Adsorption of dissolved organic matter to pristine and photochemically weathered polymer surfaces. Environ. Sci. Technol. 2023, 57, 14707–14716. [Google Scholar] [CrossRef] [PubMed]
- Khoshnamvand, M.; You, D.; Xie, Y.; Feng, Y.; Sultan, M.; Wei, X.; Li, J.; Fu, A.; Pei, D.S. Presence of humic acid in the environment holds promise as a potential mitigating factor for the joint toxicity of polystyrene nanoplastics and herbicide atrazine to Chlorella vulgaris: 96-H acute toxicity. Chemosphere 2024, 357, 142061. [Google Scholar] [CrossRef]
- Zhang, J.; Zhan, S.; Zhong, L.B.; Wang, X.; Qiu, Z.; Zheng, Y.M. Adsorption of typical natural organic matter on microplastics in aqueous solution: Kinetics, isotherm, influence factors and mechanism. J. Hazard Mater. 2023, 443, 130130. [Google Scholar] [CrossRef]
- Pedroza, R.H.P.; David, C.; Lodeiro, P.; Rey-Castro, C. Interactions of humic acid with pristine poly (lactic acid) microplastics in aqueous solution. Sci. Total Environ. 2024, 908, 168366. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Baimanov, D.; Yuan, H.; Xie, H.; Yu, S.; Zhang, Z.; Yang, J.; Zhao, F.; You, Y.; Guan, Y.; et al. Protein corona-directed cellular recognition and uptake of polyethylene nanoplastics by macrophages. Environ. Sci. Technol. 2024, 58, 14158–14168. [Google Scholar] [CrossRef]
- Giri, S.; Mukherjee, A. Ageing with algal EPS reduces the toxic effects of polystyrene nanoplastics in freshwater microalgae Scenedesmus obliquus. J. Environ. Chem. Eng. 2021, 9, 105978. [Google Scholar] [CrossRef]
- Li, X.; Ji, S.; He, E.; Peijnenburg, W.J.; Cao, X.; Zhao, L.; Xu, X.; Zhang, P.; Qiu, H. UV/ozone induced physicochemical transformations of polystyrene nanoparticles and their aggregation tendency and kinetics with natural organic matter in aqueous systems. J. Hazard Mater. 2022, 433, 128790. [Google Scholar] [CrossRef]
- Yu, S.; Li, Q.; Shan, W.; Hao, Z.; Li, P.; Liu, J. Heteroaggregation of different surface-modified polystyrene nanoparticles with model natural colloids. Sci. Total Environ. 2021, 784, 147190. [Google Scholar] [CrossRef]
- Xu, Y.; Ou, Q.; He, Q.; Wu, Z.; Ma, J.; Huangfu, X. Influence of dissolved black carbon on the aggregation and deposition of polystyrene nanoplastics: Comparison with dissolved humic acid. Water Res. 2021, 196, 117054. [Google Scholar] [CrossRef]
- Zhang, Y.-N.; Cheng, F.; Zhang, T.; Li, C.; Qu, J.; Chen, J.; Peijnenburg, W.J.G.M. Dissolved organic matter enhanced the aggregation and oxidation of nanoplastics under simulated sunlight irradiation in water. Environ. Sci. Technol. 2022, 56, 3085–3095. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, Z.; Wang, S.; Fang, H.; Wang, D. Aquatic behavior and toxicity of polystyrene nanoplastic particles with different functional groups: Complex roles of pH, dissolved organic carbon and divalent cations. Chemosphere 2019, 228, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.; Ortiz, D.; Nieto-Sandoval, J.; de Pedro, Z.M.; Casas, J.A. Adsorption of micropollutants onto realistic microplastics: Role of microplastic nature, size, age, and NOM fouling. Chemosphere 2021, 283, 131085. [Google Scholar] [CrossRef]
- Ding, L.; Luo, Y.; Yu, X.; Ouyang, Z.; Liu, P.; Guo, X. Insight into interactions of polystyrene microplastics with different types and compositions of dissolved organic matter. Sci. Total Environ. 2022, 824, 153883. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Junaid, M.; Wang, C.; Wang, J. Eco-corona enhanced the interactive effects of nanoplastics and 6:2 chlorinated polyfluorinated ether sulfonate in zebrafish embryos. Sci. Total Environ. 2024, 953, 176223. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhao, J.; Li, L.; Wang, Y.; Dai, X.; Yu, F.; Ma, J. Interfacial interaction between micro/nanoplastics and typical PPCPs and nanoplastics removal via electrosorption from an aqueous solution. Water Res. 2020, 184, 116100. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, Z.; Wei, W.; Chen, J.; Ni, B.J. Toxicity of micro/nanoplastics in the environment: Roles of plastisphere and eco-corona. Soil Environ. Health 2023, 1, 100002. [Google Scholar] [CrossRef]
- Song, Y.; Zhao, J.; Zheng, L.; Zhu, W.; Xue, X.; Yu, Y.; Deng, Y.; Wang, H. Adsorption behaviors and mechanisms of humic acid on virgin and aging microplastics. J. Mol. Liq. 2022, 363, 119819. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, Y.; Liu, G.; He, G.; Liu, W. Adsorption mechanism of cadmium on microplastics and their desorption behavior in sediment and gut environments: The roles of water pH, lead ions, natural organic matter and phenanthrene. Water Res. 2020, 184, 116209. [Google Scholar] [CrossRef]
- Wu, X.; Liu, P.; Gong, Z.; Wang, H.; Huang, H.; Shi, Y.; Zhao, X.; Gao, S. Humic acid and fulvic acid hinder long-term weathering of microplastics in lake water. Environ. Sci. Technol. 2021, 55, 15810–15820. [Google Scholar] [CrossRef]
- Fadare, O.O.; Wan, B.; Guo, L.H.; Xin, Y.; Qin, W.; Yang, Y. Humic acid alleviates the toxicity of polystyrene nanoplastic particles to Daphnia magna. Environ. Sci. J. Integr. Environ. Res. Nano 2019, 6, 1466–1477. [Google Scholar] [CrossRef]
- Zhang, M.; Hou, J.; Xia, J.; Wu, J.; Zeng, Y.; Miao, L.; Lv, B. Transport of polystyrene nanoplastics in porous media: Combined effects of two co-existing substances. Sci. Total Environ. 2023, 897, 165275. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yue, T.; Liu, L.; Zhang, B.; Feng, H.; Li, S.; Liu, X.; Dai, Y.; Zhao, J. Molecular assembly of extracellular polymeric substances regulating aggregation of differently charged nanoplastics and subsequent interactions with bacterial membrane. J. Hazard. Mater. 2023, 457, 131825. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Ma, X.; Guo, B.; Zhang, K. Environmental factors-mediated behavior of microplastics and nanoplastics in water: A review. Chemosphere 2021, 271, 129597. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Liu, Y.; Xu, Y.; Li, S.; Liu, X.; Dai, Y.; Zhao, J.; Yue, T. Benzo[a]pyrene and heavy metal ion adsorption on nanoplastics regulated by humic acid: Cooperation/competition mechanisms revealed by molecular dynamics simulations. J. Hazard Mater. 2022, 424, 127431. [Google Scholar] [CrossRef]
- Shiu, R.F.; Vazquez, C.I.; Chiang, C.Y.; Chiu, M.H.; Chen, C.S.; Ni, C.W.; Gong, G.C.; Quigg, A.; Santschi, P.H.; Chin, W.C. Nano- and microplastics trigger secretion of protein-rich extracellular polymeric substances from phytoplankton. Sci. Total Environ. 2020, 748, 141469. [Google Scholar] [CrossRef]
- Zafar, R.; Arshad, Z.; Choi, N.E.; Li, X.; Hur, J. Unravelling the complex adsorption behavior of extracellular polymeric substances onto pristine and UV-aged microplastics using two-dimensional correlation spectroscopy. Chem. Eng. J. 2023, 470, 144031. [Google Scholar] [CrossRef]
- Xu, S.Y.; Wang, C.Q.; Zhu, P.F.; Zhang, D.Y.; Pan, X.L. Temporospatial nano-heterogeneity of self-assembly of extracellular polymeric substances on microplastics and water environmental implications. J. Hazard. Mater. 2022, 440, 129773. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, X.; Xia, S.; Zhao, J. Cu(II) adsorption on Poly(Lactic Acid) microplastics: Significance of microbial colonization and degradation. Chem. Eng. J. 2022, 429, 132306. [Google Scholar] [CrossRef]
- Yang, C.W.; Yuan, L.; Zhou, H.Z.; Zhang, X.; Sheng, G.P. Coating ligand-mediated dynamic formation of natural organic matter (NOM) corona on engineered nanoparticles in natural environments. Environ. Sci. Nano 2021, 8, 1029–1041. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Q.; Yang, Y.; Hou, L.; Zheng, W.; Wu, Z.; Wang, Z. Revealing the contradiction between DLVO/XDLVO theory and membrane fouling propensity for oil-in-water emulsion separation. J Hazard Mater. 2024, 466, 133594. [Google Scholar] [CrossRef]
- Giri, S.; Mukherjee, A. Eco-corona reduces the phytotoxic effects of polystyrene nanoplastics in Allium cepa: Emphasizing the role of ROS. Environ. Exp. Bot. 2022, 198, 104850. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Z.; Bai, H.; Xue, M.; Tang, J. Photochemically induced aging of polystyrene nanoplastics and its impact on norfloxacin adsorption behavior. Sci. Total Environ. 2024, 930, 172511. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Gabellieri, E.; Cioni, P.; Paccagnini, E.; Faleri, C.; Lupetti, P.; Corsi, I.; Morelli, E. Interplay between extracellular polymeric substances (EPS) from a marine diatom and model nanoplastic through eco-corona formation. Sci. Total Environ. 2020, 725, 138457. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jiang, R.; Liu, Q.; Ouyang, G. Impact of different modes of adsorption of natural organic matter on the environmental fate of nanoplastics. Chemosphere 2021, 263, 127967. [Google Scholar] [CrossRef]
- Zhang, M.; Hou, J.; Xia, J.; Wu, J.; You, G.; Miao, L. The long-term release and particle fracture behaviors of nanoplastics retained in porous media: Effects of surfactants, natural organic matters, antibiotics, and bacteria. Sci. Total Environ. 2024, 925, 171563. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Q.; Maa, J.P.Y.; Shen, X.; Liang, J.; Yu, L.; Ge, L.; Wang, G. Effects of organic matter on interaction forces between polystyrene microplastics: An experimental study. Sci. Total Environ. 2022, 844, 157186. [Google Scholar] [CrossRef]
- Wang, X.; Bolan, N.; Tsang, D.C.W.; Sarkar, B.; Bradney, L.; Li, Y. A review of microplastics aggregation in aquatic environment: Influence factors, analytical methods, and environmental implications. J. Hazard. Mater. 2021, 402, 123496. [Google Scholar] [CrossRef]
- Pradel, A.; Ferreres, S.; Veclin, C.; El Hadri, H.; Gautier, M.; Grassl, B.; Gigault, J. Stabilization of fragmental polystyrene nanoplastic by natural organic matter: Insight into mechanisms. ACS EST Water 2021, 1, 1198–1208. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chen, K.F.; Lin, K.Y.A.; Chen, J.K.; Jiang, X.Y.; Lin, C.H. The nephrotoxic potential of polystyrene microplastics at realistic environmental concentrations. J. Hazard. Mater. 2022, 427, 127871. [Google Scholar] [CrossRef]
- Ruan, J.; Yang, J.; Wang, X.; Liang, C.; Li, L.; Zeng, Y.; Wang, J.; Li, Y.; Huang, W.; Chen, C. Heteroaggregation kinetics of oppositely charged nanoplastics in aquatic environments: Effects of particle ratio, solution chemistry, and interaction sequence. J Hazard Mater. 2024, 475, 134857. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, X.; van der Hoek, J.P.; Liu, G.; Lompe, K.M. Natural organic matter stabilizes pristine nanoplastics but destabilizes photochemical weathered nanoplastics in monovalent electrolyte solutions. Environ. Sci. Technol. 2025, 59, 1822–1834. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Shen, M.; Li, S.; Fu, Y.; Zhang, D.; Liu, H.; Liu, J. Aggregation kinetics of different surface-modified polystyrene nanoparticles in monovalent and divalent electrolytes. Environ. Pollut. 2019, 255, 113302. [Google Scholar] [CrossRef]
- Huang, S.H.; Zhang, B.; Cui, F.Y.; He, Y.K.; Shi, J.Y.; Yang, X.Y.; Lens, P.N.L.; Shi, W.X. Mechanisms underlying the detrimental impact of micro(nano)plastics on the stability of aerobic granular sludge: Interactions between micro(nano)plastics and extracellular polymeric substances. J. Hazard. Mater. 2024, 478, 135512. [Google Scholar] [CrossRef]
- Huang, R.; Han, Z.; Ma, C.; Liu, H.; Huangfu, X. Stability and mobility of zinc oxide nanoparticles in aquatic environment: Influence of extracellular polymeric substances from cyanobacteria and microalgae. J. Environ. Chem. Eng. 2023, 11, 109069. [Google Scholar] [CrossRef]
- Li, L.; Luo, D.; Luo, S.; Yue, J.; Li, X.; Chen, L.; Chen, X.; Wen, B.; Luo, X.; Li, Y.; et al. Heteroaggregation, disaggregation, and migration of nanoplastics with nanosized activated carbon in aquatic environments: Effects of particle property, water chemistry, and hydrodynamic condition. Water Res. 2024, 266, 122399. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Jiao, R.; Yu, J.; Wang, D. Combined effects of particle size and humic acid corona on the aggregation kinetics of nanoplastics in aquatic environments. Sci. Total. Environ. 2023, 901, 165987. [Google Scholar] [CrossRef]
- Zhang, M.; Hou, J.; Xia, J.; Zeng, Y.; Miao, L. Influence of natural organic matters on fate of polystyrene nanoplastics in porous media. Sci. Total Environ. 2023, 893, 64504. [Google Scholar] [CrossRef]
- Kong, Y.; Li, X.; Tao, M.; Cao, X.; Wang, Z.; Xing, B. Cation-π mechanism promotes the adsorption of humic acid on polystyrene nanoplastics to differently affect their aggregation: Evidence from experimental characterization and DFT calculation. J. Hazard. Mater. 2023, 459, 132071. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, H.; Li, H.; Yin, Y.; Song, W.; Guo, H.; Huang, T.; Xing, B. Molecular-level insight into the behavior of metal cations and organic matter during the aggregation of polystyrene nanoplastics. J Hazard Mater. 2024, 473, 134665. [Google Scholar] [CrossRef]
- Mao, Y.; Li, H.; Huangfu, X.; Liu, Y.; He, Q. Nanoplastics display strong stability in aqueous environments: Insights from aggregation behaviour and theoretical calculations. Environ. Pollut. 2020, 258, 113760. [Google Scholar] [CrossRef]
- Guckeisen, T.; Orghici, R.; Rathgeber, S. Correlative effects on nanoplastic aggregation in model extracellular biofilm substances investigated with fluorescence correlation spectroscopy. Polymers 2024, 16, 2170. [Google Scholar] [CrossRef]
- Xu, Y.; Ou, Q.; Li, X.; Wang, X.; van der Hoek, J.P.; Liu, G. Combined effects of photoaging and natural organic matter on the colloidal stability of nanoplastics in aquatic environments. Water Res. 2022, 226, 119313. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jiang, R.; Lin, W.; Ouyang, G. Effect of salinity and humic acid on the aggregation and toxicity of polystyrene nanoplastics with different functional groups and charges. Environ. Pollut. 2019, 245, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Lian, F.; Han, Y.; Zhang, Y.; Li, J.; Sun, B.; Geng, Z.; Wang, Z.; Xing, B. Exposure order to photoaging and humic acids significantly modifies the aggregation and transformation of nanoplastics in aqueous solutions. Environ. Sci. Technol. 2023, 57, 6520–6529. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Chen, C.; Huang, Z.; Gu, J.; Zhang, Z.; Cai, T.; Peng, J.; Huang, W.; Dang, Z.; Yang, C. Influence of macromolecules and electrolytes on heteroaggregation kinetics of polystyrene nanoplastics and goethite nanoparticles in aquatic environments. J. Hazard. Mater. 2024, 477, 135257. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Du, J.; Pu, L.; Chen, S. Effects of chlorella extracellular polymeric substances on the aggregation and stability of TiO2 nanoparticles as electrolytes. Desalination Water Treat. 2021, 209, 334–341. [Google Scholar] [CrossRef]
- Xiong, S.; Cao, X.; Eggleston, I.; Chi, Y.; Li, A.; Liu, X.; Zhao, J.; Xing, B. Role of extracellular polymeric substances in the aggregation and biological response of micro(nano)plastics with different functional groups and sizes. J. Hazard. Mater. 2023, 446, 130713. [Google Scholar] [CrossRef]
- Cai, L.; Hu, L.; Shi, H.; Ye, J.; Zhang, Y.; Kim, H. Effects of inorganic ions and natural organic matter on the aggregation of nanoplastics. Chemosphere 2018, 197, 142–151. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, H.; Zhao, J.; Luo, X.; Wang, Z.; Xing, B. Photodegradation elevated the toxicity of polystyrene microplastics to grouper (Epinephelus moara) through disrupting hepatic lipid homeostasis. Environ. Sci. Technol. 2020, 54, 6202–6212. [Google Scholar] [CrossRef]
- Sun, H.; Jiao, R.; Gao, Y.; Wang, D. Insight into the aggregation kinetics of nanoplastics in aquatic environments: Combined effects of protein corona and particle size. Sep. Purif. Technol. 2025, 362, 131899. [Google Scholar] [CrossRef]
- Beddoes, C.M.; Case, C.P.; Briscoe, W.H. Understanding nanoparticle cellular entry: A physicochemical perspective. Adv. Colloid Interface Sci. 2015, 218, 48–68. [Google Scholar] [CrossRef]
- Kihara, S.; Ashenden, A.; Kaur, M.; Glasson, J.; Ghosh, S.; van der Heijden, N.; Brooks, A.E.S.; Mata, J.P.; Holt, S.; Domigan, L.J.; et al. Cellular interactions with polystyrene nanoplastics-The role of particle size and protein corona. Biointerphases 2021, 16, 041001. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Nakamura, H.; Ohsaki, S.; Watano, S. Direct translocation of a negatively charged nanoparticle across a negatively charged model cell membrane. Phys. Chem. Chem. Phys. 2021, 23, 10591–10599. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, G.; Wei, W. Simulation of nanoparticles interacting with a cell membrane: Probing the structural basis and potential biomedical application. NPG Asia Mater. 2021, 13, 52. [Google Scholar] [CrossRef]
- Li, N.; Wang, P.; Wang, S.; Wang, C.; Zhou, H.; Kapur, S.; Zhang, J.; Song, Y. Electrostatic charges on microalgae surface: Mechanism and applications. J. Environ. Chem. Eng. 2022, 10, 1–12. [Google Scholar] [CrossRef]
- Hanachi, P.; Khoshnamvand, M.; Walker, T.R.; Hamidian, A.H. Nano-sized polystyrene plastics toxicity to microalgae Chlorella vulgaris: Toxicity mitigation using humic acid. Aquat. Toxicol. 2022, 245, 106123. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.; Christudoss, A.C.; Chandrasekaran, N.; Peijnenburg, W.J.G.M.; Mukherjee, A. The role of algal EPS in reducing the combined toxicity of BPA and polystyrene nanoparticles to the freshwater algae Scenedesmus obliquus. Plant Physiol. Biochem. 2023, 197, 107664. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, W.; Meng, L.; Zeb, A.; Mo, F.; Wang, J.; Shi, R. The interfacial interaction between Dechlorane plus (DP) and polystyrene nanoplastics (PSNPs): An overlooked influence factor for the algal toxicity of PSNPs. Sci. Total Environ. 2023, 905, 167129. [Google Scholar] [CrossRef]
- Debroy, A.; Saravanan, J.S.; Joyce Nirmala, M.; Pulimi, M.; Mukherjee, D.A. Algal EPS modifies the toxicity potential of the mixture of polystyrene nanoplastics (PSNPs) and flame retardant, triphenyl phosphate in freshwater microalgae Chlorella sp. Chemosphere 2024, 366, 143471. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Gao, R.Y.; Wang, Z.J.; Shao, Q.Q.; Hu, Y.W.; Jia, H.B.; Liu, X.J.; Dong, F.Q.; Fu, L.M.; Zhang, J.P. Daphnia magna uptake and excretion of luminescence-labelled polystyrene nanoparticle as visualized by high sensitivity real-time optical imaging. Chemosphere 2023, 326, 138341. [Google Scholar] [CrossRef]
- Guo, Y.; Tang, N.; Lu, L.; Li, N.; Hu, T.; Guo, J.; Zhang, J.; Zeng, Z.; Liang, J. Aggregation behavior of polystyrene nanoplastics: Role of surface functional groups and protein and electrolyte variation. Chemosphere. 2024, 350, 140998. [Google Scholar] [CrossRef]
- Canesi, L.; Balbi, T.; Fabbri, R.; Salis, A.; Damonte, G.; Volland, M.; Blasco, J. Biomolecular coronas in invertebrate species: Implications in the environmental impact of nanoparticles. NanoImpact 2017, 8, 89–98. [Google Scholar] [CrossRef]
- Ding, J.; Huang, Y.; Liu, S.; Zhang, S.; Zou, H.; Wang, Z.; Geng, J. Toxicological effects of nano-and micro-polystyrene plastics on red tilapia: Are larger plastic particles more harmless? J. Hazard. Mater. 2020, 396, 122693. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Ding, J.; Xu, H.; Tian, W.; Xu, J.; Zou, H.; Zhu, W. Influences of molecular weight fractionated humic acids on polyamide 66 microplastic stability and toxicity in red tilapia (Oreochromis niloticus). Front. Mar. Sci. 2022, 9, 1060582. [Google Scholar] [CrossRef]
- Lei, C.; Sun, Y.; Tsang, D.C.W.; Lin, D. Environmental transformations and ecological effects of iron-based nanoparticles. Environ. Pollut. 2018, 232, 10–30. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, L.; Zhao, J.; Xing, B. Environmental processes and toxicity of metallic nanoparticles in aquatic systems as affected by natural organic matter. Environ. Sci. Nano 2016, 3, 240–255. [Google Scholar] [CrossRef]
- Luo, H.; Du, Q.; Zhong, Z.; Xu, Y.; Peng, J. Protein-coated microplastics corona complex: An underestimated risk of microplastics. Sci. Total Environ. 2022, 851, 157948. [Google Scholar] [CrossRef]
- Li, S.; Wang, S.; Yan, B.; Yue, T. Surface properties of nanoparticles dictate their toxicity by regulating adsorption of humic acid molecules. ACS Sustain. Chem. Eng. 2021, 9, 13705–13716. [Google Scholar] [CrossRef]
- Zou, W.; Xia, M.L.; Jiang, K.; Cao, Z.; Zhang, X.; Hu, X. Photo-oxidative degradation mitigated the developmental toxicity of polyamide microplastics to zebrafish larvae by modulating macrophage-triggered proinflammatory responses and apoptosis. Environ. Sci. Technol. 2020, 54, 13888–13898. [Google Scholar] [CrossRef]
- Xu, Y.; Ou, Q.; van der Hoek, J.P.; Liu, G.; Lompe, K.M. Photo-oxidation of micro- and nanoplastics: Physical, chemical, and biological effects in environments. Environ. Sci. Technol. 2024, 58, 991–1009. [Google Scholar] [CrossRef]
- Qiao, R.; Deng, Y.; Zhang, S.; Wolosker, M.B.; Zhu, Q.; Ren, H.; Zhang, Y. Accumulation of different shapes of microplastics initiates intestinal injury and gut microbiota dysbiosis in the gut of zebrafish. Chemosphere 2019, 236, 124334. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, X.; Wu, A.; Tang, Z.; Niu, L.; Wu, F.; Wang, F.; Zhao, T.; Fu, Z. Aggregation and stability of sulfate-modified polystyrene nanoplastics in synthetic and natural waters. Environ. Pollut. 2020, 268, 114240. [Google Scholar] [CrossRef]
- Triebskorn, R.; Braunbeck, T.; Grummt, T.; Hanslik, L.; Huppertsberg, S.; Jekel, M.; Knepper, T.P.; Krais, S.; Müller, Y.K.; Pittroff, M.; et al. Relevance of nano- and microplastics for freshwater ecosystems: A critical review. Trends Analyt. Chem. 2019, 110, 375–392. [Google Scholar] [CrossRef]
- Merrill, G.B.; Hermabessiere, L.; Rochman, C.M.; Nowacek, D.P. Microplastics in marine mammal blubber, melon, & other tissues: Evidence of translocation. Environ. Pollut. 2023, 335, 122252. [Google Scholar] [PubMed]
- Zhang, Y.; Goss, G.G. The “Trojan Horse” effect of nanoplastics: Potentiation of polycyclic aromatic hydrocarbon uptake in rainbow trout and the mitigating effects of natural organic matter. Environ. Sci. Nano 2021, 8, 3685–3698. [Google Scholar] [CrossRef]
- Monikh, F.A.; Vijver, M.G.; Guo, Z.; Zhang, P.; Darbha, G.K.; Peijnenburg, W.J. Metal sorption onto nanoscale plastic debris and trojan horse effects in Daphnia magna: Role of dissolved organic matter. Water Res. 2020, 186, 116410. [Google Scholar] [CrossRef]
- Qiao, R.; Lu, K.; Deng, Y.; Ren, H.; Zhang, Y. Combined effects of polystyrene microplastics and natural organic matter on the accumulation and toxicity of copper in zebrafish. Sci. Total Environ. 2019, 682, 128–137. [Google Scholar] [CrossRef]
- Chen, W.; Sandoval, H.; Kubiak, J.Z.; Li, X.C.; Ghobrial, R.M.; Kloc, M. The phenotype of peritoneal mouse macrophages depends on the mitochondria and ATP/ADP homeostasis. Cell Immunol. 2018, 324, 1–7. [Google Scholar] [CrossRef]
- Mao, R.; Lang, M.; Yu, X.; Wu, R.; Yang, X.; Guo, X. Aging mechanism of microplastics with UV irradiation and its effects on the adsorption of heavy metals. J. Hazard. Mater. 2020, 393, 122515. [Google Scholar] [CrossRef]
- Natarajan, L.; Omer, S.; Jetly, N.; Jenifer, M.A.; Chandrasekaran, N.; Suraishkumar, G.K.; Mukherjee, A. Eco-Corona formation lessens the toxic effects of polystyrene nanoplastics towards marine microalgae Chlorella sp. Environ. Res. 2020, 188, 109842. [Google Scholar] [CrossRef]
- Khoshnamvand, M.; You, D.; Xie, Y.; Feng, Y.; Sultan, M.; Pei, D.S.; Fu, A. Alleviating binary toxicity of polystyrene nanoplastics and atrazine to Chlorella vulgaris through humic acid interaction: Long-term toxicity using environmentally relevant concentrations. Chemosphere 2024, 358, 142111. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, K.; Kashian, D.R. Extracellular polymeric substances in green alga facilitate microplastic deposition. Chemosphere 2022, 286, 131814. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yuan, Y.; Li, Y.; Liu, X.; Wang, X.; Fan, Z. Polystyrene nanoplastics affect growth and microcystin production of Microcystis aeruginosa. Environ. Sci. Pollut. Res. 2021, 28, 13394–13403. [Google Scholar] [CrossRef]
- Abomohra, A.; Hanelt, D. Recent advances in micro-/nanoplastic (MNPs) removal by microalgae and possible integrated routes of energy recovery. Microorganisms 2022, 10, 2400. [Google Scholar] [CrossRef]
- Liu, T.; Hou, B.; Wang, Z.; Yang, Y. Polystyrene microplastics induce mitochondrial damage in mouse GC-2 cells. Ecotoxicol. Environ. Saf. 2022, 237, 113520. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cong, J.Y.; Wu, J.; Chen, Y.Z.; Fan, H.Y.; Wang, X.L.; Duan, Z.H.; Wang, L. Nanoplastic-protein corona interactions and their biological effects: A review of recent advances and trends. Trends Analyt. Chem. 2023, 166, 117206. [Google Scholar] [CrossRef]
- He, Y.; Peng, G.; Jiang, Y.; Zhao, M.; Wang, X.; Chen, M.; Lin, S. Environmental hazard potential of nano-photocatalysts determined by nano-bio interactions and exposure conditions. Small 2020, 16, 1907690. [Google Scholar] [CrossRef]
- Natarajan, L.; Annie Jenifer, M.; Peijnenburg, W.J.G.M.; Mukherjee, A. Algal extracellular polymeric substances (algal-EPS) for mitigating the combined toxic effects of polystyrene nanoplastics and nano-TiO2 in Chlorella sp. Nanotoxicology 2023, 17, 143–156. [Google Scholar] [CrossRef]
- Zhu, K.; Jia, H.; Sun, Y.; Dai, Y.; Zhang, C.; Guo, X.; Wang, T.; Zhu, L. Long-term phototransformation of microplastics under simulated sunlight irradiation in aquatic environments: Roles of reactive oxygen species. Water Res. 2020, 173, 115564. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, T.; Qu, G.; Jia, H.; Zhu, L. Probing the aging processes and mechanisms of microplastic under simulated multiple actions generated by discharge plasma. J. Hazard. Mater. 2020, 398, 122956. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, Y.; Zhang, L.; Xu, M.; Gao, L.; Zhao, B. The interaction of micro/nano plastics and the environment: Effects of ecological corona on the toxicity to aquatic organisms. Ecotoxicol. Environ. Saf. 2022, 243, 113997. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Wang, C.; Wang, Y.; Fu, L.; Man, M.; Chen, L. Realistic polyethylene terephthalate nanoplastics and the size-and surface coating-dependent toxicological impacts on zebrafish embryos. Environ. Sci. J. Integr. Environ. Res. Nano 2020, 7, 2313–2324. [Google Scholar] [CrossRef]
- Barabadi, H.; Najafi, M.; Samadian, H.; Azarnezhad, A.; Vahidi, H.; Mahjoub, M.A.; Koohiyan, M.; Ahmadi, A. A systematic review of the genotoxicity and antigenotoxicity of biologically synthesized metallic nanomaterials: Are green nanoparticles safe enough for clinical marketing? Medicina 2019, 55, 439. [Google Scholar] [CrossRef]
- Khan, A.R.; Ulhassan, Z.; Li, G.; Lou, J.; Iqbal, B.; Salam, A.; Azhar, W.; Batool, S.; Zhao, T.; Li, K.; et al. Micro/ nanoplastics: Critical review of their impacts on plants, interactions with other contaminants (antibiotics, heavy metals, and polycyclic aromatic hydrocarbons), and management strategies. Sci. Total Environ. 2024, 912, 169420. [Google Scholar] [CrossRef]
- Shukla, R.K.; Badiye, A.; Vajpayee, K.; Kapoor, N. Genotoxic potential of nanoparticles: Structural and functional modifications in DNA. Front. Genet. 2021, 12, 728250. [Google Scholar] [CrossRef]
- Gunderson, M.P.; Nguyen, B.T.; Reyes, J.C.C.; Holden, L.L.; French, J.M.; Smith, B.D.; Lineberger, C. Response of phase I and II detoxification enzymes, glutathione, metallothionein and acetylcholine esterase to mercury and dimethoate in signal crayfish (Pacifastacus leniusculus). Chemosphere 2018, 208, 749–756. [Google Scholar] [CrossRef]
- Flohé, L.; Toppo, S.; Orian, L. The glutathione peroxidase family: Discoveries and mechanism. Free Radic. Biol. Med. 2022, 187, 113–122. [Google Scholar] [CrossRef]
- Zámocký, M.; Koller, F. Understanding the structure and function of catalases: Clues from molecular evolution and in vitro mutagenesis. Prog. Biophys. Mol. Biol. 1999, 72, 19–66. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Staud, F.; Ceckova, M.; Micuda, S.; Pavek, P. Expression and function of pglycoprotein in normal tissues: Effect on pharmacokinetics. Methods Mol. Biol. 2010, 596, 199–222. [Google Scholar] [PubMed]
- Fadare, O.O.; Wan, B.; Liu, K.; Yang, Y.; Zhao, L.; Guo, L.H. Eco-corona vs protein corona: Effects of humic substances on corona formation and nanoplastic particle toxicity in Daphnia magna. Environ. Sci. Technol. 2020, 54, 8001–8009. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Ghosh, S. Therapeutic potential of fulvic acid in chronic inflammatory diseases and diabetes. J. Diabetes Res. 2018, 2018, 5391014. [Google Scholar] [CrossRef]
- Faught, E.; Schaaf, M.J.M. Molecular mechanisms of the stress-induced regulation of the inflammatory response in fish. Gen. Comp. Endocrinol. 2024, 345, 114387. [Google Scholar] [CrossRef]
- Khan, A.R.; Wallbom, N.; Sundh, H.; Sandblom, E.; Tort, L.; Jönsson, E. Sea water acclimation of rainbow trout (Oncorhynchus mykiss) modulates the mucosal transcript immune response induced by Vibrio anguillarum and Aeromonas salmonicida vaccine, and prevents further transcription of stress-immune genes in response to acutes. Fish Shellfish Immunol. 2024, 152, 109733. [Google Scholar]
- Brandts, I.; Balasch, J.C.; Gonçalves, A.P.; Martins, M.A.; Pereira, M.L.; Tvarijonaviciute, A.; Teles, M.; Oliveira, M. Immuno-modulatory effects of nanoplastics and humic acids in the European seabass (Dicentrarchus labrax). J. Hazard Mater. 2021, 414, 125562. [Google Scholar] [CrossRef]
- Fuse, Y.; Endo, Y.; Araoi, S.; Daitoku, H.; Suzuki, H.; Kato, M.; Kobayashi, M. The possible repositioning of an oral anti-arthritic drug, auranofin, for Nrf2-activating therapy: The demonstration of Nrf2-dependent anti-oxidative action using a zebrafish model. Free Radic. Biol. Med. 2018, 115, 405–411. [Google Scholar] [CrossRef]
- Raghunath, A.; Nagarajan, R.; Sundarraj, K.; Panneerselvam, L.; Perumal, E. Genomewide identification and analysis of Nrf2 binding sites—Antioxidant response elements in zebrafish. Toxicol. Appl. Pharmacol. 2018, 360, 236–248. [Google Scholar] [CrossRef]
- Zhou, J.; Miners, J.O. Enzyme kinetics of uridine diphosphate glucuronosyltransferases (UGTs). Methods Mol Biol. 2014, 1113, 203–228. [Google Scholar]
- Shrestha, S.; Wang, B.; Dutta, P. Nanoparticle processing: Understanding and controlling aggregation. Adv. Colloid. Interface Sci. 2020, 279, 102162. [Google Scholar] [CrossRef] [PubMed]
- Gasco, R.; Worms, I.A.M.; Kantarciyan, A.; Slaveykova, V.I. Diatom-derived extracellular polymeric substances form eco-corona and enhance stability of silver nanoparticles. Environ. Sci. Nano 2024, 11, 4138–4150. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yang, Q.; Jiang, J.; Dalu, T.; Kadushkin, A.; Singh, J.; Fakhrullin, R.; Wang, F.; Cai, X.; Li, R. Coronas of micro/nano plastics: A key determinant in their risk assessments. Part Fibre Toxicol. 2022, 19, 55. [Google Scholar] [CrossRef] [PubMed]
- Nasser, F.; Constantinou, J.; Lynch, I. Nanomaterials in the environment acquire an “eco-corona” impacting their toxicity to Daphnia magna—A call for updating toxicity testing policies. Proteomics 2020, 20, e1800412. [Google Scholar] [CrossRef]
- Soliman, M.G.; Martinez-Serra, A.; Antonello, G.; Dobricic, M.; Wilkins, T.; Serchi, T.; Fenoglio, I.; Monopoli, M.P. Understanding the role of biomolecular coronas in human exposure to nanomaterials. Environ. Sci. Nano 2024, 11, 4421–4448. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M/NPs | Original Surface State | Ecological Macromolecules | Formation Mechanisms | References |
|---|---|---|---|---|
| PS NPs | Pristine | EPS | Hydrogen bonding | [11] |
| PE, PP, PS, PET MPs | Pristine | Suwannee River FA, HA, NOM | Hydrophobic interactions | [42] |
| Polyvinyl chloride (PVC) MPs, PS MPs | Pristine | HA | Electrostatic interactions, hydrogen bonding, π–π and hydrophobic interactions | [44] |
| PLA MPs | Pristine | HA | Hydrophobic interactions, hydrogen bonding | [45] |
| PS NPs, PS NPs-COOH, PS NPs-NH2 | Pristine and functionalized | EPS | Electrostatic interactions, hydrophobic interactions, and van der Waals forces | [47] |
| PS NPs | Aged | HA | Electrostatic interactions and van der Waals forces | [48] |
| PS NPs, PS NPs-COOH, PS NPs-NH2 | Pristine and functionalized | Suwannee River NOM | Electrostatic interactions | [49] |
| PS NPs | Pristine and aged | HA | Electrostatic, hydrophobic interactions | [50] |
| PS NPs | Pristine | Suwannee River FA | Hydrophobic interactions | [51] |
| Polystyrene latex (PSL) NPs, PSL NPs-NH2, PSL NPs-COOH | Pristine and functionalized | FA | Electrostatic interactions | [52] |
| High-density polyethylene (HDPE), PP, PET, PS MPs | Pristine | NOM | Electrostatic and hydrophobic interactions | [53] |
| PS MPs | Pristine | FA, HA | Hydrophobic interactions | [54] |
| PS NPs | Pristine | HA | Electrostatic interactions | [55] |
| PS, PVC M/NPs | Pristine | HA | Hydrophobic interactions | [56] |
| PS NPs | Pristine | EPS | Electrostatic interactions and van der Waals forces | [57] |
| Polyamide (PA), PP MPs | Pristine | HA | Electrostatic, hydrophobic interactions | [58] |
| Acrylonitrile butadiene styrene, PA, PVC, PS, PET MPs | Pristine | HA | Electrostatic interactions | [59] |
| PP MPs | Pristine | Suwannee River HA, Pony Lake FA | Electrostatic, hydrophobic interactions | [60] |
| PS NPs-NH2, PS MPs-NH2 | Functionalized | HA | Electrostatic interactions | [61] |
| PS NPs | Pristine | HA | Electrostatic interactions | [62] |
| Theoretical Calculation | M/NPs | Ecological Macromolecules | Regulatory Mechanisms | References |
|---|---|---|---|---|
| DLVO | PS NPs | HA, SA | Electrostatic interactions, molecular bridging | [62] |
| PS NPs, PS NPs-NH2, PS NPs-COOH | HA | Electrostatic interactions, molecular bridging | [76] | |
| Aged PET NPs | HA | Electrostatic interactions | [77] | |
| PS NPs, PS NPs-NH2, PS NPs-COOH | NOM | Electrostatic interactions | [78] | |
| PS NPs, PS NPs-NH2, PS NPs-COOH | EPS | Electrostatic interactions, molecular bridging | [79] | |
| PS NPs | HA, FA | Electrostatic interactions | [80] | |
| PS NPs | HA, SA | Electrostatic interactions | [81] | |
| PS NPs | HA, SA | Electrostatic interactions | [82] | |
| XDLVO | PS NPs | HA, BSA | Electrostatic interactions, steric hindrance, molecular bridging | [37] |
| Aged PS NPs | HA, SA | Electrostatic interactions, steric hindrance, molecular bridging | [48] | |
| PS M/NPs | EPS | Electrostatic interactions, steric hindrance | [83] | |
| PS NPs | EPS | Electrostatic interactions, steric hindrance | [84] | |
| PS NPs | HA | Electrostatic interactions, steric hindrance | [85] | |
| PS NPs | NOM | Steric hindrance, molecular bridging | [86] | |
| PS NPs | BSA | Steric hindrance, molecular bridging | [87] | |
| PS NPs, PS NPs-NH2, PS NPs-COOH | BSA | Electrostatic interactions, steric hindrance | [88] |
| Biotrophic Level | Toxicity Enhancement/ Reduction Mechanisms | M/NPs | Ecological Macromolecules | Test Organisms | References |
|---|---|---|---|---|---|
| Algae | Alleviate oxidative stress | PS MPs | EPS | S. costatum | [11] |
| Prevent cell uptake, reduced ROS production | PS NPs | EPS | S. obliquus | [47] | |
| Alleviate oxidative stress by scavenging free radicals | PS NPs-COOH | EPS | P. tricornutum | [74] | |
| Reduce electrostatic attraction between NPs and algal cells | PS NPs | HA | C. vulgaris | [107] | |
| Alleviate oxidative stress | PS NPs-COOH, PS NPs-NH2 | EPS | S. obliquus | [108] | |
| Alleviate oxidative stress | PS NPs, PS NPs-COOH, PS NPs-NH2 | EPS | Chlorella sp. | [130] | |
| Reduce ROS production | PS NPs | EPS | Chlorella sp. | [131] | |
| Invertebrates | Damage feeding capacity | PS NPs-COOH, PS NPs-NH2 | Proteins | D. magna | [31] |
| Reduce the entanglement and body burden of NPs | PS NPs, PS NPs-COOH, PS NPs-NH2 | HA | D. magna | [112] | |
| Fish | Cause poor lipid adsorption and growth inhibition | PA MPs | HA, FA | D. rerio | [18] |
| Damage the immune system and induce immune suppression | PS M/NPs | EPS | O. melastigma | [39] | |
| Alleviate hepatic lipid peroxidation and protein damage | PA MPs | HA | O. niloticus | [115] | |
| Increase accumulation of MPs in the intestine, inhibit food intake, and promote ROS production | PS MPs | BSA | D. rerio | [118] | |
| Enhance oxidative stress | PS MPs | NOM | D. rerio | [126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Chen, Z.; Kong, L.; Xing, H.; Yang, Q.; Wu, J. A Review of Eco-Corona Formation on Micro/Nanoplastics and Its Effects on Stability, Bioavailability, and Toxicity. Water 2025, 17, 1124. https://doi.org/10.3390/w17081124
Yang H, Chen Z, Kong L, Xing H, Yang Q, Wu J. A Review of Eco-Corona Formation on Micro/Nanoplastics and Its Effects on Stability, Bioavailability, and Toxicity. Water. 2025; 17(8):1124. https://doi.org/10.3390/w17081124
Chicago/Turabian StyleYang, Haohan, Zhuoyu Chen, Linghui Kong, Hao Xing, Qihang Yang, and Jun Wu. 2025. "A Review of Eco-Corona Formation on Micro/Nanoplastics and Its Effects on Stability, Bioavailability, and Toxicity" Water 17, no. 8: 1124. https://doi.org/10.3390/w17081124
APA StyleYang, H., Chen, Z., Kong, L., Xing, H., Yang, Q., & Wu, J. (2025). A Review of Eco-Corona Formation on Micro/Nanoplastics and Its Effects on Stability, Bioavailability, and Toxicity. Water, 17(8), 1124. https://doi.org/10.3390/w17081124