
Temporal Variation of Plankton Community in Typical Lake in Middle Reaches of Yangtze River: Structure, Environmental Response and Interactions

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
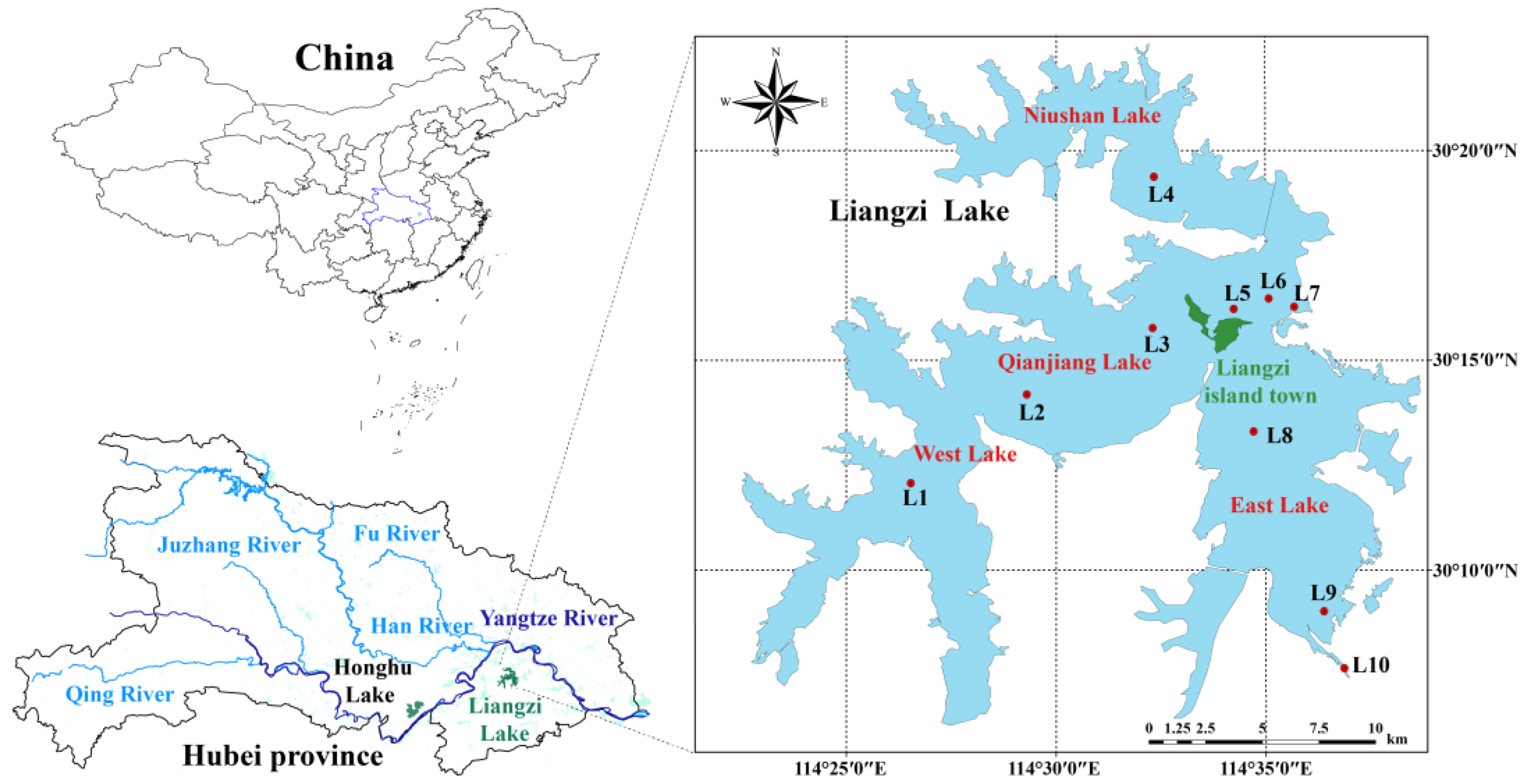
2.1. Sampling and Laboratory Analysis
2.2. DNA Extraction, and Sequencing Analysis
2.3. Calculation Methods
2.4. Statistical Analysis
3. Results
3.1. Environmental Characteristics
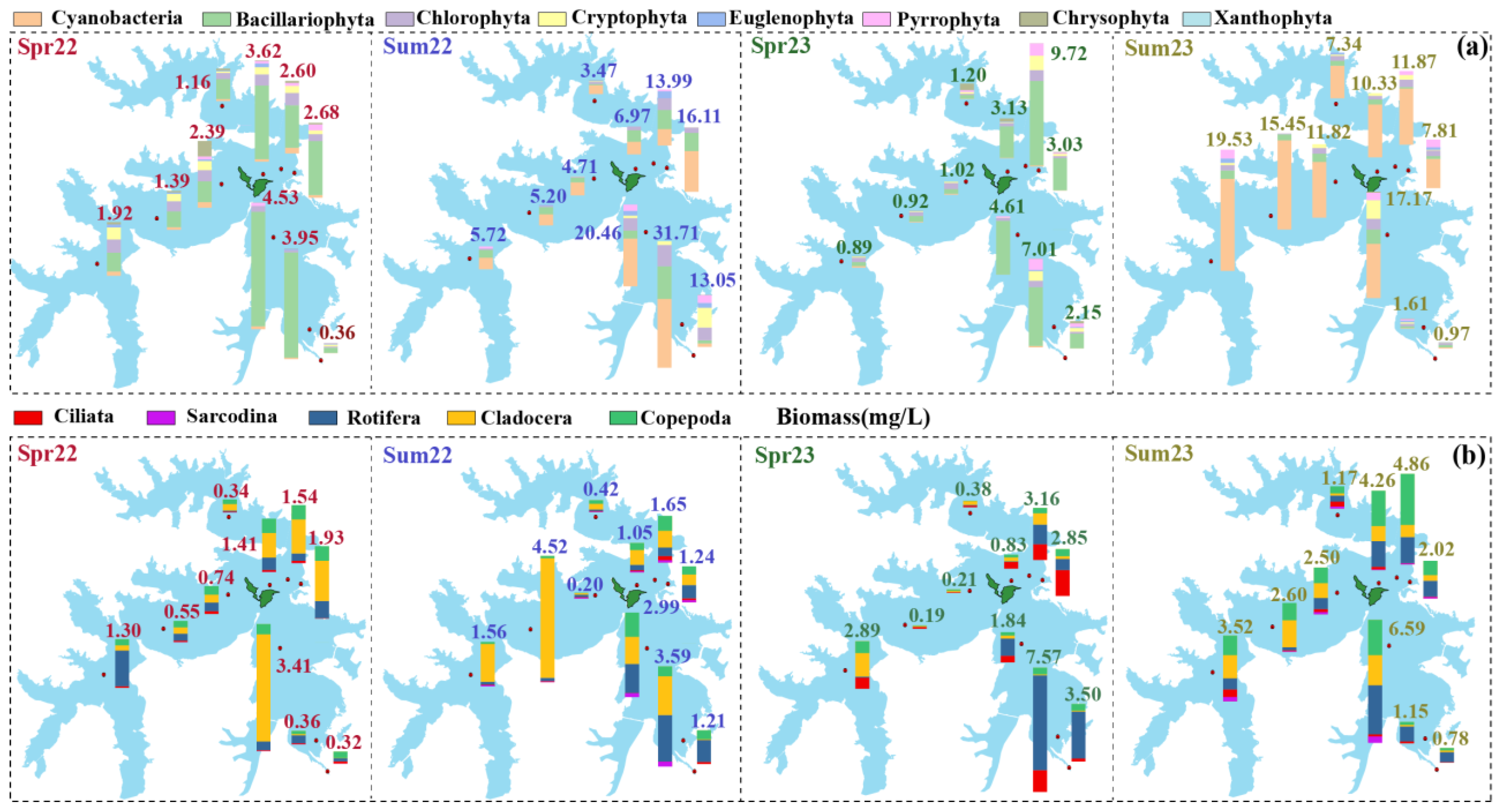
3.2. Biomass of Planktonic Communities
3.3. Abundance of Planktonic Communities
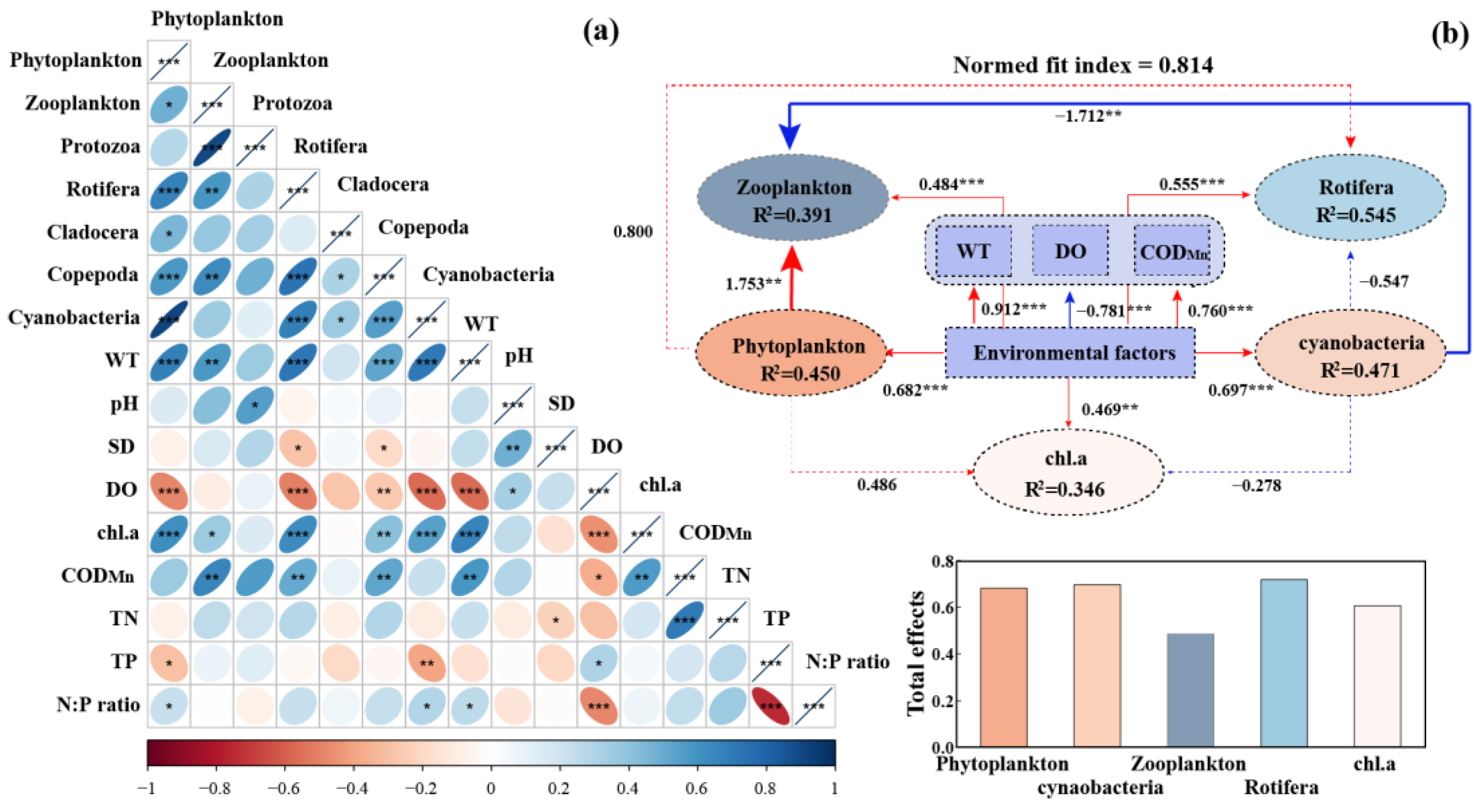
3.4. Environmental Drivers of Planktonic Communities
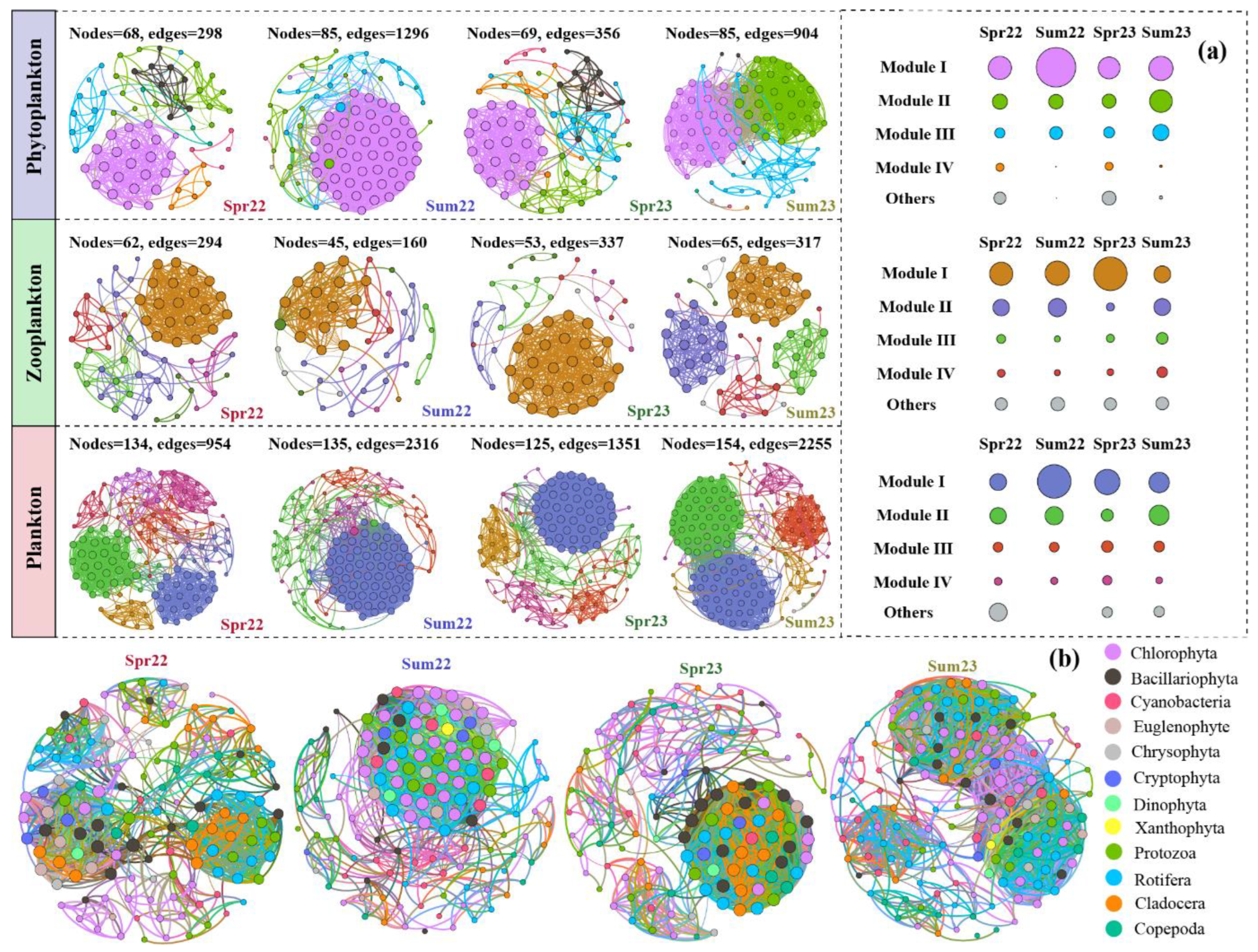
3.5. Planktonic Co-Existence Patterns
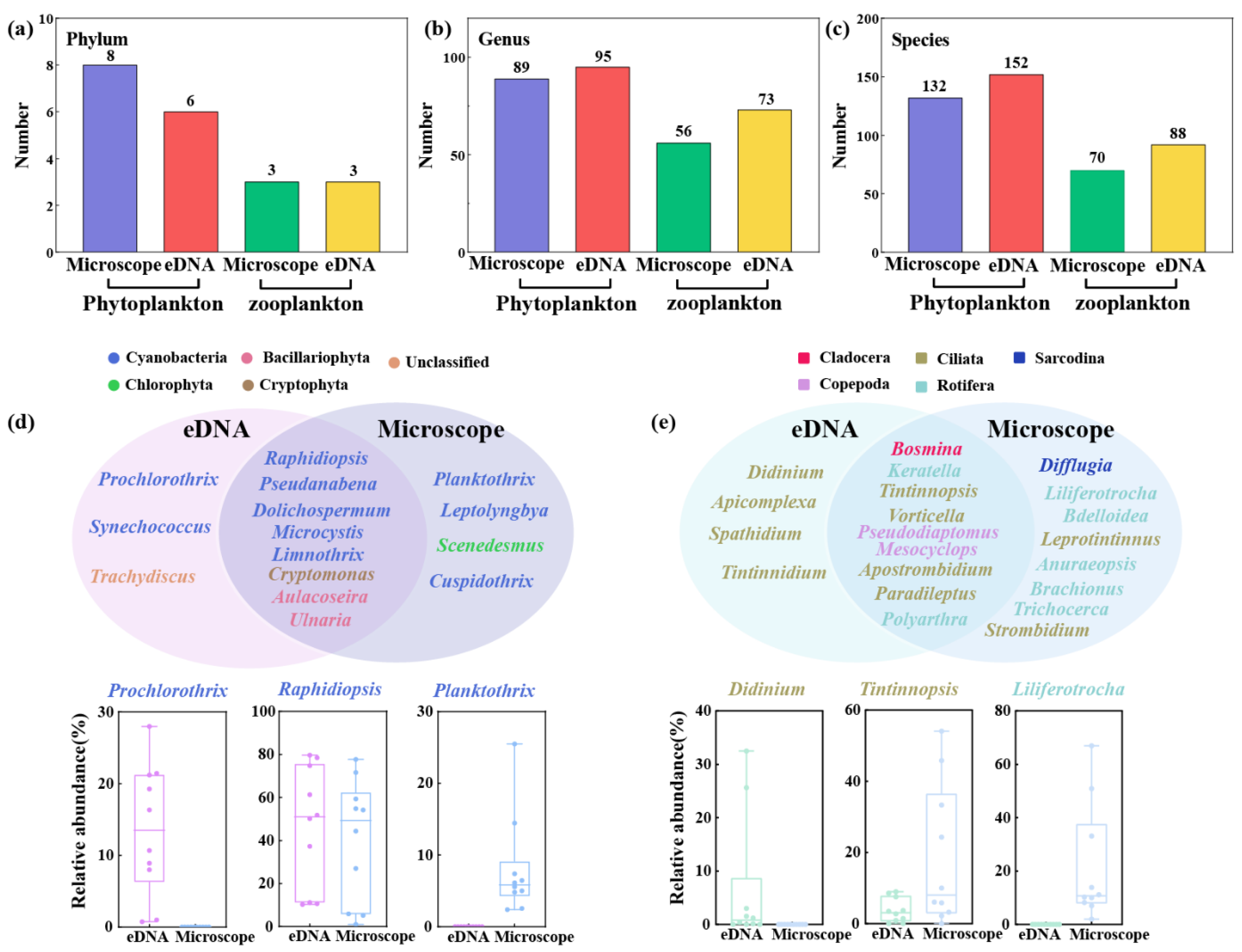
3.6. Comparison of eDNA Metabarcoding and Microscopical Analysis
4. Discussion
4.1. Temporal Dynamics and Environmental Response of Planktonic Communities
4.2. Interactions of Planktonic Communities
4.3. Difference Between Morphological Analysis and eDNA Metabarcoding
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Choo, S.; Kwak, M.T.; Cho, Y.K.; Yoon, Y.H.; Soh, H.Y. Effects of water masses on the zooplankton community structure in the northern East China Sea during the East Asian Summer Monsoon in 2020. Ecol. Indic. 2023, 154, 110847. [Google Scholar] [CrossRef]
- Seymour, J.R.; Amin, S.A.; Raina, J.B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton-bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef]
- Zeng, L.; Wen, J.; Huang, B.; Yang, Y.; Huang, Z.; Zeng, F.; Fang, H.; Du, H. Environmental DNA metabarcoding reveals the effect of environmental selection on phytoplankton community structure along a subtropical river. Environ. Res. 2024, 243, 117708. [Google Scholar] [CrossRef] [PubMed]
- Park, T.; Lee, G.; Shin, J.; Park, J.-H.; Choi, B.; Kim, D.-K.; Cha, Y. Changes in zooplankton community in response to a shift from lentic to lotic conditions in a regulated river. Ecol. Inform. 2023, 77, 102236. [Google Scholar] [CrossRef]
- Ding, X.; Liu, J.; Liu, W.; Dai, S.; Ke, Z.; Guo, J.; Lai, Y.; Tan, Y. Phytoplankton Communities Miniaturization Driven by Extreme Weather in Subtropical Estuary under Climate Changes. Water Res. 2023, 245, 120588. [Google Scholar] [CrossRef]
- Fu, X.; Shi, W.; Liu, Z.; Wang, X.; Zhang, G.; Sun, J. Impact of environmental variables on the distribution of phytoplankton communities in the Southern Yellow Sea. Environ. Res. 2024, 243, 117862. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yang, A.; Liu, J.; Xing, C.; Huang, S.; Huo, Y.; Yang, Z.; Huang, J.; Liu, W. Turnover of phytoplankton and zooplankton communities driven by human-induced disturbances and climate changes in a small urban coastal wetland. Ecol. Indic. 2023, 157, 111271. [Google Scholar] [CrossRef]
- Song, J.; Liang, D. Community structure of zooplankton and its response to aquatic environmental changes based on eDNA metabarcoding. J. Hydrol. 2023, 622, 129692. [Google Scholar] [CrossRef]
- Liao, N.; Chen, Z.; Zhang, L.; Chen, M.; Zhang, Y.; Li, J.; Wang, H. Study on the spatiotemporal distribution of algal blooms and its influencing factors in young reservoirs based on remote sensing interpretation. J. Environ. Manag. 2024, 354, 120333. [Google Scholar] [CrossRef]
- Hu, Q.; Chen, Y.; Xia, R.; Liu, X.; Jia, R.; Zhang, K.; Li, X.; Yan, C.; Wang, Y.; Yin, Y.; et al. Weakened hydrological oscillation period increased the frequency of river algal blooms. Water Res. 2024, 255, 121496. [Google Scholar] [CrossRef]
- Wang, N.; Mark, N.; Launer, N.; Hirtler, A.; Weston, C.; Cleckner, L.; Faehndrich, C.; LaGorga, L.; Xia, L.; Pyrek, D.; et al. Harmful algal blooms in Cayuga lake, NY: From microbiome analysis to eDNA monitoring. J. Environ. Manag. 2024, 354, 120128. [Google Scholar] [CrossRef]
- Meng, J.; Xu, F.; Yang, H.; Li, X.; Zhao, P. Exploring microbiome and plankton responses and interactions in the mangrove ecosystem through eDNA and network analysis. Sci. Total Environ. 2024, 930, 172581. [Google Scholar] [CrossRef]
- Djurhuus, A.; Pitz, K.; Sawaya, N.A.; Rojas-Márquez, J.; Michaud, B.; Montes, E.; Muller-Karger, F.; Breitbart, M. Evaluation of marine zooplankton community structure through environmental DNA metabarcoding. Limnol. Oceanogr. 2018, 16, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.W.; Oh, H.J.; Chang, K.H.; Park, Y.S.; Kwak, I.S. A Comparative Analyzing of Zooplankton Community Diversity in Surface Layer Water of Reservoir Via eDNA Metabarcoding and Microscopy. Diversity 2022, 14, 797. [Google Scholar] [CrossRef]
- Ge, Y.; Zhang, Q.; Dong, X.; Yang, X. Revealing anthropogenic effects on lakes and wetlands: Pollen-based environmental changes of Liangzi Lake, China over the last 150 years. Catena 2021, 207, 105605. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, W.; Shan, B. Distribution of nitrogen and phosphorus and estimation of nutrient fluxes in the water and sediments of Liangzi Lake, China. Environ. Sci. Pollut. Res. Int. 2020, 27, 7096–7104. [Google Scholar] [CrossRef]
- Chen, N.; Wang, S.; Zhang, X.; Yang, S. A risk assessment method for remote sensing of cyanobacterial blooms in inland waters. Sci. Total. Environ. 2020, 740, 140012. [Google Scholar] [CrossRef]
- Wang, C.; Zhu, Y.; Wang, L.; Wu, W. Assessment on Ecosystem Health in the Liangzi Lake. Environ. Sci. Technol. 2015, 38, 398–404. (In Chinese) [Google Scholar]
- Yi, M.; Li, L.; Li, H.; Liu, C.; Deng, Y.; Wu, Z.; Li, Y. Spatiotemporal variations of plankton communities in different water bodies of the Yellow River: Structural characteristics, biogeographic patterns, environmental responses, and community assembly. J. Hydrol. 2024, 640, 131702. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, University of Galway. Available online: https://www.algaebase.org (accessed on 5 December 2024).
- Jia, J.; Xi, X.; Li, X.; Hu, H.; Chen, K.; Wu, C. Characteristics of microbial communities and antibiotic resistance genes in typical rivers of the western Qinghai Lake basin. Water Bio. Sec. 2024, 3, 100249. [Google Scholar] [CrossRef]
- Jia, J.; Guan, Y.; Cheng, M.; Chen, H.; He, J.; Wang, S.; Wang, Z. Occurrence and distribution of antibiotics and antibiotic resistance genes in Ba River, China. Sci. Total Environ. 2018, 642, 1136–1144. [Google Scholar] [CrossRef]
- Wu, S.; Dong, Y.; Stoeck, T.; Wang, S.; Fan, H.; Wang, Y.; Zhuang, X. Geographic characteristics and environmental variables determine the diversities and assembly of the algal communities in interconnected river-lake system. Water Res. 2023, 233, 119792. [Google Scholar] [CrossRef]
- Jia, J.; Cheng, M.; Xue, X.; Guan, Y.; Wang, Z. Characterization of tetracycline effects on microbial community, antibiotic resistance genes and antibiotic resistance of Aeromonas spp. in gut of goldfish Carassius auratus Linnaeus. Ecotoxicol. Environ. Saf. 2020, 191, 110182. [Google Scholar] [CrossRef]
- Ministry of Ecology and Environment of the People’s Republic of China (2011, March). Methods for the Assessment of Surface Water Environmental Quality of China. Available online: https://www.mee.gov.cn/gkml/hbb/bgt/201104/W020110401583735386081.pdf (accessed on 20 March 2025).
- Ren, Y.; Yu, G.; Shi, C.; Liu, L.; Guo, Q.; Han, C.; Zhang, D.; Zhang, L.; Liu, B.; Gao, H.; et al. Majorbio Cloud: A one-stop, comprehensive bioinformatic platform for multiomics analyses. iMeta 2022, 1, e12. [Google Scholar] [CrossRef]
- Chen, Y.; Su, J.Q.; Zhang, J.; Li, P.; Chen, H.; Zhang, B.; Gin, K.Y.; He, Y. High-throughput profiling of antibiotic resistance gene dynamic in a drinking water river-reservoir system. Water Res. 2019, 149, 179–189. [Google Scholar] [CrossRef]
- Li, B.; Yang, Y.; Ma, L.; Ju, F.; Guo, F.; Tiedje, J.M.; Zhang, T. Metagenomic and network analysis reveal wide distribution and co-occurrence of environmental antibiotic resistance genes. ISME J. 2015, 9, 2490–2502. [Google Scholar] [CrossRef]
- Sommer, U.; Adrian, R.; De Senerpont Domis, L.; Elser, J.J.; Gaedke, U.; Ibelings, B.; Jeppesen, E.; Lürling, M.; Molinero, J.C.; Mooij, W.M.; et al. Beyond the Plankton Ecology Group (PEG) Model: Mechanisms Driving Plankton Succession. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- dos Santos Severiano, J.; dos Santos Almeida-Melo, V.L.; Bittencourt-Oliveira, M.d.C.; Chia, M.A.; do Nascimento Moura, A. Effects of increased zooplankton biomass on phytoplankton and cyanotoxins: A tropical mesocosm study. Harmful Algae 2018, 71, 10–18. [Google Scholar] [CrossRef]
- Shi, J.; Wang, D.; Wang, J.; Sheng, J.; Peng, K.; Hu, B.; Zeng, L.; Xiao, M.; Hong, Y. Comparative analysis of the complete mitochondrial genomes of three geographical topmouth culter (Culter alburnus) groups and implications for their phylogenetics. Biosci. Biotechnol. Biochem. 2017, 81, 482–490. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, W.; Xiong, P.; Li, H.; Liu, Z.; Ai, J.; Yuan, D.; Wan, F.; Wan, Y.; Zou, H.; et al. Temporal variation of plankton and zoobenthos communities in a freshwater reservoir: Structure feature, construction mechanism, associated symbiosis and environmental response. Ecol. Indic. 2023, 154, 110774. [Google Scholar] [CrossRef]
- Zhou, S.Y.; Lie, Z.; Liu, X.; Zhu, Y.G.; Peñuelas, J.; Neilson, R.; Su, X.; Liu, Z.; Chu, G.; Meng, Z.; et al. Distinct patterns of soil bacterial and fungal community assemblages in subtropical forest ecosystems under warming. Glob. Chang. Biol. 2023, 29, 1501–1513. [Google Scholar] [CrossRef]
- Xu, S.; Xiao, Y.; Xu, Y.; Su, L.; Cai, Y.; Qi, Z.; Liu, Y.; Chen, Z.; Lakshmikandan, M. Effects of seasonal variations and environmental factors on phytoplankton community structure and abundance in Beibu Gulf, China. Ocean Coast Manag. 2024, 248, 106982. [Google Scholar] [CrossRef]
- T-Krasznai, E.; Török, P.; Borics, G.; Lukács, Á.; Kókai, Z.; Lerf, V.; Görgényi, J.; B-Béres, V. Functional dynamics of phytoplankton assemblages in hypertrophic lakes: Functional- and species diversity is highly resistant to cyanobacterial blooms. Ecol. Indic. 2022, 145, 109583. [Google Scholar] [CrossRef]
- Karpowicz, M.; Kornijów, R.; Ejsmont-Karabin, J.; Nawrocka, L.; Kobos, J.; Jakubowska, A.; Grzonkowski, K. Seasonal dynamics of phytoplankton and zooplankton communities in the estuarine Elblag Bay (Vistula Lagoon, southern Baltic) dominated by floating-leaved plants. Ecohydrol. Hydrobiol. 2024, 25, 215–224. [Google Scholar] [CrossRef]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef]
- Wu, Y.; Peng, C.; Li, G.; He, F.; Huang, L.; Sun, X.; Wu, S. Integrated evaluation of the impact of water diversion on water quality index and phytoplankton assemblages of eutrophic lake: A case study of Yilong Lake. J. Environ. Manag. 2024, 357, 120707. [Google Scholar] [CrossRef]
- Cao, J.; Hou, Z.Y.; Li, Z.K.; Zheng, B.H.; Chu, Z.S. Spatiotemporal dynamics of phytoplankton biomass and community succession for driving factors in a meso-eutrophic lake. J. Environ. Manag. 2023, 345, 118693. [Google Scholar] [CrossRef]
- Shi, J.; Ni, L.; Liu, J.; Xu, C.; Zhang, J.; Chen, X. Spatiotemporal distribution of phytoplankton community structure and its relationship with environmental factors in Hongze Lake, China. Urban Clim. 2023, 52, 101746. [Google Scholar] [CrossRef]
- Jabir, T.; Vipindas, P.V.; Jesmi, Y.; Valliyodan, S.; Parambath, P.M.; Singh, A.; Abdulla, M.H. Nutrient stoichiometry (N:P) controls nitrogen fixation and distribution of diazotrophs in a tropical eutrophic estuary. Mar. Pollut. Bull. 2020, 151, 110799. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Xu, H.; Kang, L.; Zhao, X. Spatial and seasonal change in algal community structure and its interaction with nutrient dynamics in a gravel-bed urban river. J. Hazard. Mater. 2022, 425, 127775. [Google Scholar] [CrossRef] [PubMed]
- Bustos-Díaz, E.D.; Barona-Gómez, F.; Cibrián-Jaramillo, A. Cyanobacteria in Nitrogen-Fixing Symbioses. In Cyanobacteria, 2nd ed.; Mishra, A.K., Tiwari, D.N., Rai, A.N., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 29–42. [Google Scholar]
- Carneiro, F.M.; Santos, A.M.C.; Medina, N.G.; De Marco Júnior, P.; Hortal, J. Drivers of phytoplankton diversity in tropical artificial ponds. Perspect. Ecol. Conser. 2024, 22, 167–176. [Google Scholar] [CrossRef]
- Tao, T.; Wang, H.; Na, X.; Liu, Y.; Zhang, N.; Lu, X.; Fan, Y. Temperate urban wetland plankton community stability driven by environmental variables, biodiversity, and resource use efficiency: A case of Hulanhe Wetland. Front. Ecol. Evol. 2023, 11, 1148580. [Google Scholar] [CrossRef]
- Wang, L.; Chen, J.; Su, H.J.; Ma, X.F.; Wu, Z.X.; Shen, H.; Yu, J.; Liu, J.R.; Wu, Y.; Ding, G.Y.; et al. Is Zooplankton Body Size an Indicator of Water Quality in (Sub)tropical Reservoirs in China? Ecosystems 2022, 25, 308–319. [Google Scholar] [CrossRef]
- Shen, J.; Qin, G.; Yu, R.; Zhao, Y.; Yang, J.; An, S.; Liu, R.; Leng, X.; Wan, Y. Urbanization has changed the distribution pattern of zooplankton species diversity and the structure of functional groups. Ecol. Indic. 2021, 120, 106944. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.; Gong, Y.; Zhang, P.; Zhang, H.; Wang, T.; Xie, J.; Xu, J.; Wang, H.; Kong, X. Benefits of combined environmental DNA and microscopy for diversity monitoring in rotifer community: A mesocosm experiment. Ecol. Indic. 2023, 155, 110930. [Google Scholar] [CrossRef]
- Marshall, C.C.; Watkins, J.M.; Connolly, J.K.; Boynton, P.V.; Schaefer, S.L.; Currie, W.J.S.; Rudstam, L.G. Patterns and drivers of seasonal succession and vertical distribution in the rotifer community of Lake Ontario in 2018. J. Great Lakes Res. 2024, 50, 102283. [Google Scholar] [CrossRef]
- Nwinyimagu, A.J.; Eyo, J.E.; Okogwu, O.I. Seasonal variation in abundance and diversity of zooplankton in Asu River, Ebonyi state, Nigeria. Acta Ecol. Sin. 2021, 41, 591–596. [Google Scholar] [CrossRef]
- Shao, Z.; Xie, P.; Zhuge, Y. Long-term changes of planktonic rotifers in a subtropical Chinese lake dominated by filter-feeding fishes. Freshw. Biol. 2001, 46, 973–986. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, H.; Shen, W.; Du, X.; Li, S.; Wei, Z.; Zhang, Z.; Feng, K.; Deng, Y. The large-scale spatial patterns of ecological networks between phytoplankton and zooplankton in coastal marine ecosystems. Sci. Total. Environ. 2022, 827, 154285. [Google Scholar] [CrossRef]
- Shuwang, X.; Zhang, G.; Li, D.; Wen, Y.; Zhang, G.; Sun, J. Spatial and temporal changes in the assembly mechanism and co-occurrence network of the chromophytic phytoplankton communities in coastal ecosystems under anthropogenic influences. Sci. Total Environ. 2023, 877, 162831. [Google Scholar] [CrossRef]
- Bockwoldt, K.A.; Nodine, E.R.; Mihuc, T.B.; Shambaugh, A.D.; Stockwell, J.D. Reduced phytoplankton and zooplankton diversity associated with increased cyanobacteria in Lake Champlain, USA. J. Contemp. Water Res. Educ. 2017, 160, 100–118. [Google Scholar] [CrossRef]
- Marrec, P.; McNair, H.; Franzè, G.; Morison, F.; Strock, J.P.; Menden-Deuer, S. Seasonal variability in planktonic food web structure and function of the Northeast U.S. Shelf. Limnol. Oceanogr. 2021, 66, 1440–1458. [Google Scholar] [CrossRef]
- Bohmann, K.; Evans, A.; Gilbert, M.T.P.; Carvalho, G.R.; Creer, S.; Knapp, M.; Yu, D.W.; de Bruyn, M. Environmental DNA for wildlife biology and biodiversity monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Deiner, K.; Fronhofer, E.A.; Mächler, E.; Walser, J.C.; Altermatt, F. Environmental DNA reveals that rivers are conveyer belts of biodiversity information. Nat. Commun. 2016, 7, 12544. [Google Scholar] [CrossRef]
- Harvey, J.B.J.; Johnson, S.B.; Fisher, J.L.; Peterson, W.T.; Vrijenhoek, R.C. Comparison of morphological and next generation DNA sequencing methods for assessing zooplankton assemblages. J. Exp. Mar. Biol. Ecol. 2017, 487, 113–126. [Google Scholar] [CrossRef]
- Liu, S.; Cui, Z.; Zhao, Y.; Chen, N. Composition and spatial-temporal dynamics of phytoplankton community shaped by environmental selection and interactions in the Jiaozhou Bay. Water Res. 2022, 218, 118488. [Google Scholar] [CrossRef]
- Cowart, D.A.; Murphy, K.R.; Cheng, C.H.C. Metagenomic sequencing of environmental DNA reveals marine faunal assemblages from the West Antarctic Peninsula. Mar. Genom. 2018, 37, 148–160. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, B.; Hu, H.; Jia, J.; Wu, W.; Li, X.; Chen, X.; Zeng, H.; Wang, Z.; Wu, C. Temporal Variation of Plankton Community in Typical Lake in Middle Reaches of Yangtze River: Structure, Environmental Response and Interactions. Water 2025, 17, 1021. https://doi.org/10.3390/w17071021
Zou B, Hu H, Jia J, Wu W, Li X, Chen X, Zeng H, Wang Z, Wu C. Temporal Variation of Plankton Community in Typical Lake in Middle Reaches of Yangtze River: Structure, Environmental Response and Interactions. Water. 2025; 17(7):1021. https://doi.org/10.3390/w17071021
Chicago/Turabian StyleZou, Borui, Hongjuan Hu, Jia Jia, Weiju Wu, Xin Li, Xiaofei Chen, Honghui Zeng, Zhi Wang, and Chenxi Wu. 2025. "Temporal Variation of Plankton Community in Typical Lake in Middle Reaches of Yangtze River: Structure, Environmental Response and Interactions" Water 17, no. 7: 1021. https://doi.org/10.3390/w17071021
APA StyleZou, B., Hu, H., Jia, J., Wu, W., Li, X., Chen, X., Zeng, H., Wang, Z., & Wu, C. (2025). Temporal Variation of Plankton Community in Typical Lake in Middle Reaches of Yangtze River: Structure, Environmental Response and Interactions. Water, 17(7), 1021. https://doi.org/10.3390/w17071021