
Relationships Between Land Use and Stream Macroinvertebrate Biotic Integrity in Central Ohio, USA


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Spatial Analysis
2.3. Field Analysis
2.4. Statistical Analysis
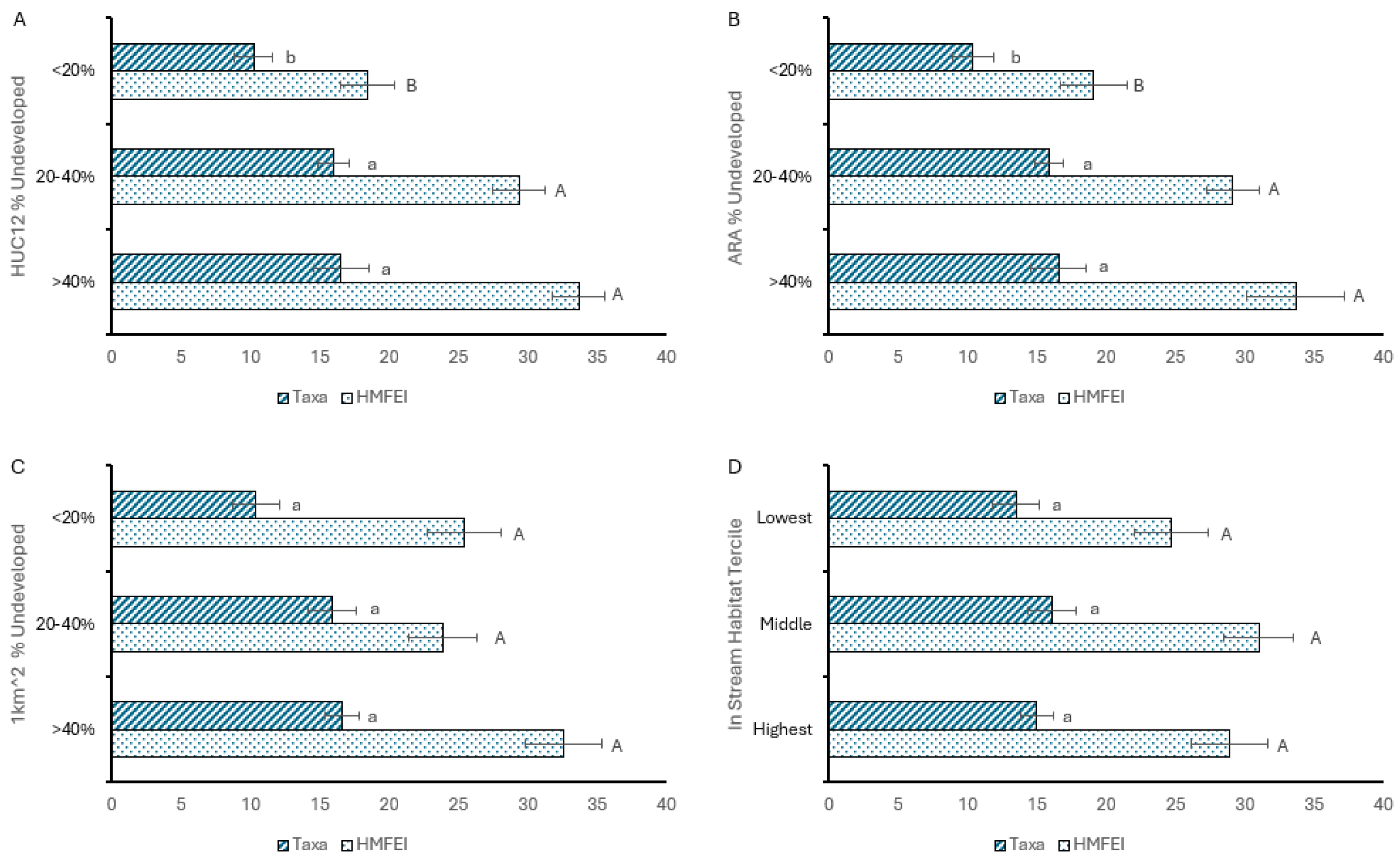
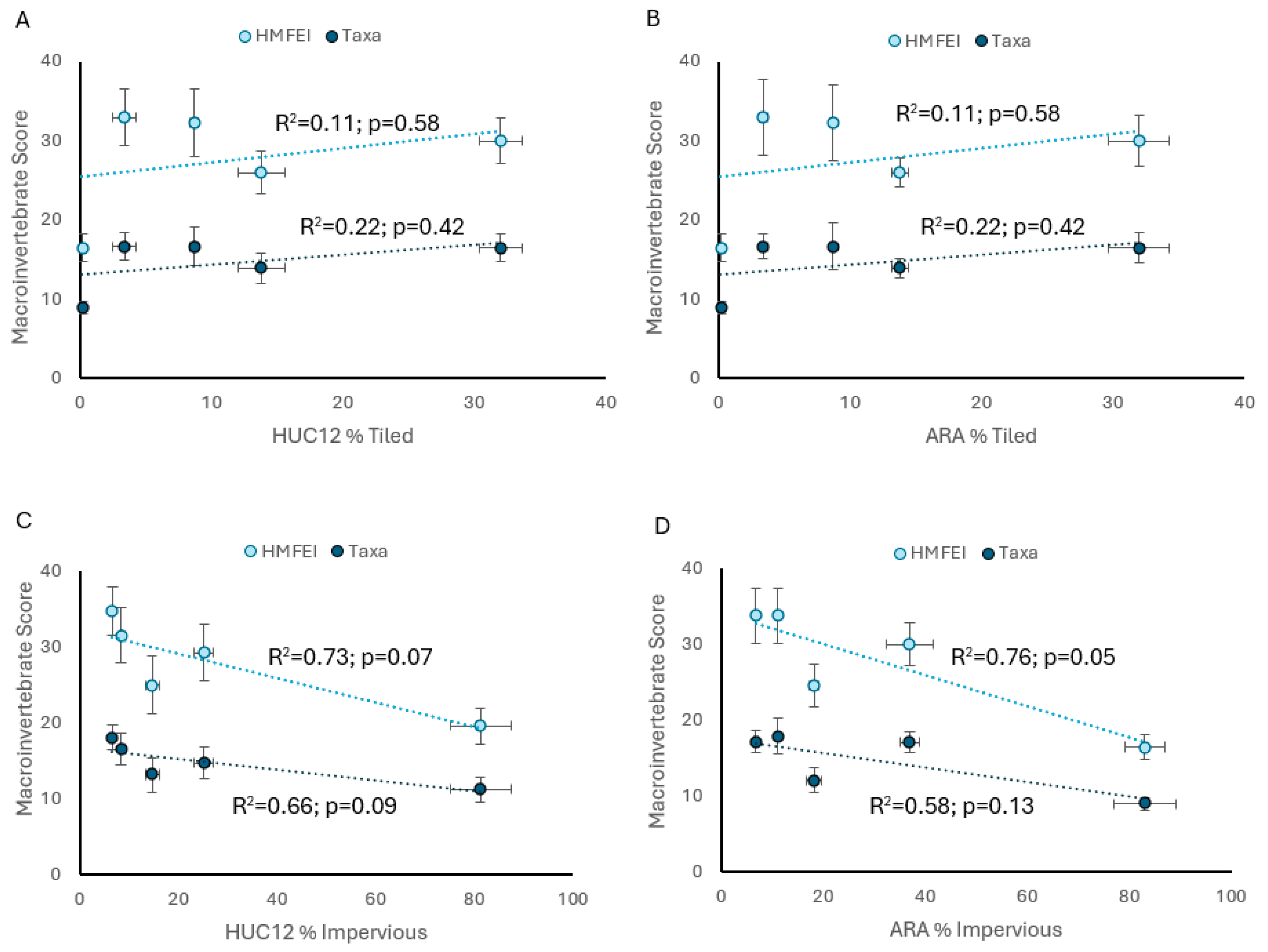
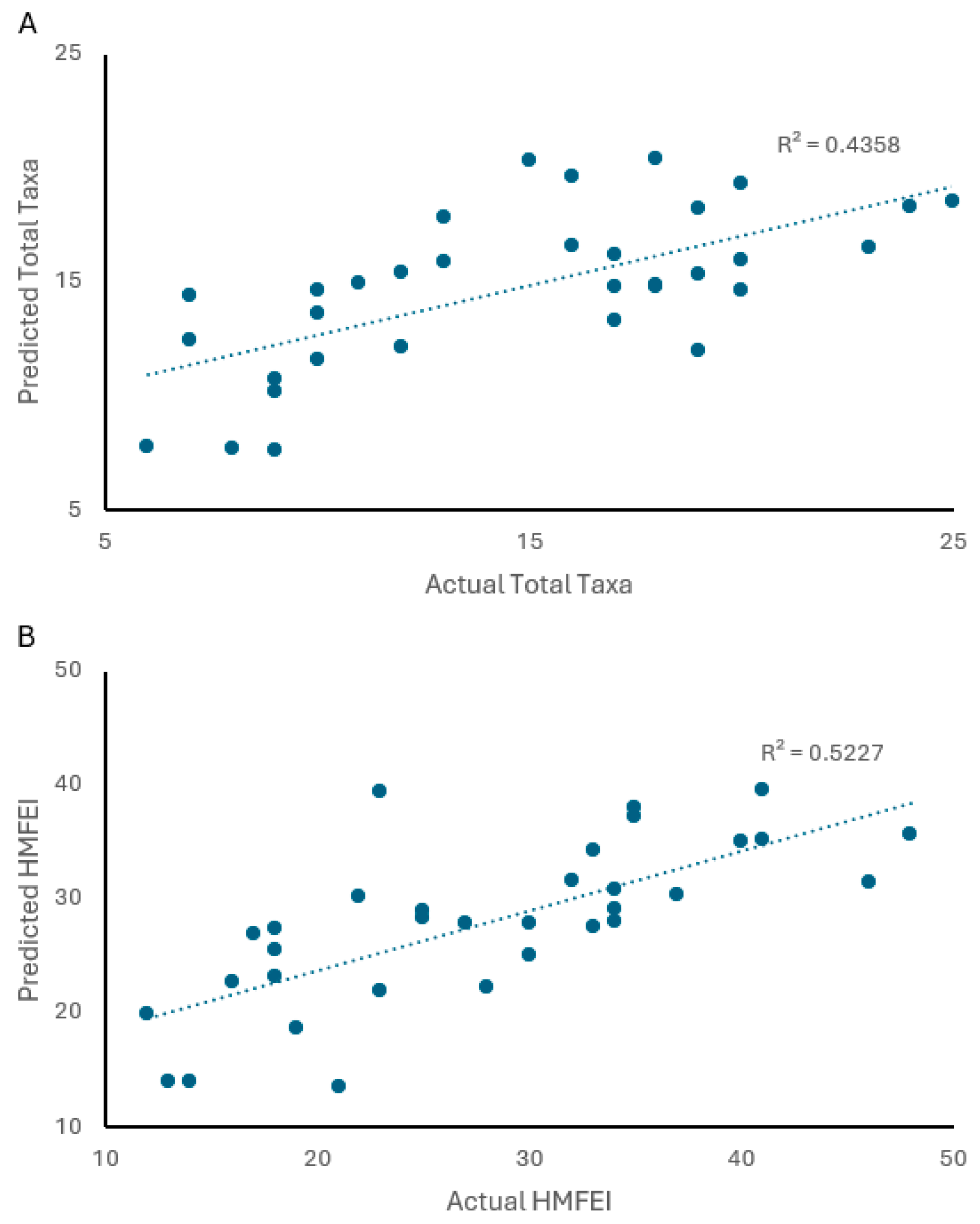
3. Results
3.1. Descriptive Statistics
3.2. Analysis of Hypotheses
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, C.; Zhang, F.; Shi, J.; Kung, H.T. What is the relationship between land use and surface water quality? A review and prospects from remote sensing perspective. Environ. Sci. Pollut. Res. 2022, 29, 56887–56907. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.J.; DeFries, R. Ecological mechanisms linking protected areas to surrounding lands. Ecol. Appl. 2007, 17, 974–988. [Google Scholar] [CrossRef]
- Lin, J.; Huang, J.; Prell, C.; Bryan, B.A. Changes in supply and demand mediate the effects of land-use change on freshwater ecosystem services flows. Sci. Total Environ. 2021, 763, 143012. [Google Scholar] [CrossRef]
- Fierro, P.; Bertrán, C.; Tapia, J.; Hauenstein, E.; Peña-Cortés, F.; Vergara, C.; Cerna, C.; Vargas-Chacoff, L. Effects of local land-use on riparian vegetation, water quality, and the functional organization of macroinvertebrate assemblages. Sci. Total Environ. 2017, 609, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.R.; Stanley, E.H.; Vander Zanden, M.J. State of the world’s freshwater ecosystems: Physical, chemical, and biological changes. Ann. Rev. Environ. Res. 2011, 36, 75–99. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Deng, X.; Du, J. Land Quality: Environmental and Human Health Effects. In Encyclopedia of Environmental Health; Nriagu, J.O., Ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 362–365. [Google Scholar] [CrossRef]
- Lin, X.W.; Zhang, X.G.; Tan, L.; Tian, Z.; Li, J.T.; Luo, Q.Y. Temporal ecological processes have different seasonal influences on multiple dimensions of riverine insect diversity in China. Insect Conserv. Div. 2024, 18, 246–259. [Google Scholar] [CrossRef]
- Clarke, A.; Mac Nally, R.; Bond, N.; Lake, P.S. Macroinvertebrate diversity in headwater streams: A review. Freshw. Biol. 2008, 53, 1707–1721. [Google Scholar] [CrossRef]
- Atkinson, C.L.; Capps, K.A.; Rugenski, A.T.; Vanni, M.J. Consumer-driven nutrient dynamics in freshwater ecosystems: From individuals to ecosystems. Biol. Rev. 2017, 92, 2003–2023. [Google Scholar] [CrossRef]
- Graça, M.A. The role of invertebrates on leaf litter decomposition in streams–a review. Int. Rev. Hydrobiol. 2001, 86, 383–393. [Google Scholar] [CrossRef]
- Chakraborty, A.; Saha, G.K.; Aditya, G. Macroinvertebrates as engineers for bioturbation in freshwater ecosystem. Environ. Sci. Pollut. Res. 2022, 29, 64447–64468. [Google Scholar] [CrossRef] [PubMed]
- Macadam, C.R.; Stockan, J.A. More than just fish food: Ecosystem services provided by freshwater insects. Ecol. Entomol. 2015, 40, 113–123. [Google Scholar] [CrossRef]
- Carter, J.L.; Resh, V.H.; Hannaford, M.J. Macroinvertebrates as biotic indicators of environmental quality. In Methods in Stream Ecology, 3rd ed.; Lamberti, G.A., Hauer, F.R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 293–318. [Google Scholar] [CrossRef]
- Petsch, D.K.; Blowes, S.A.; Melo, A.S.; Chase, J.M. A synthesis of land use impacts on stream biodiversity across metrics and scales. Ecology 2021, 102, e03498. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.M.; Herlihy, A.T.; Comeleo, R.; Peck, D.; Mitchell, R.M.; Paulsen, S.G. Patterns in and predictors of stream and river macroinvertebrate genera and fish species richness across the conterminous USA. Knowl. Manag. Aquat. Ecosyst. 2023, 424, 19. [Google Scholar] [CrossRef]
- Rumschlag, S.L.; Mahon, M.B.; Jones, D.K.; Battaglin, W.; Behrens, J.; Bernhardt, E.S.; Bradley, P.; Brown, E.; de Laender, F.; Hill, R.; et al. Density declines, richness increases, and composition shifts in stream macroinvertebrates. Sci. Adv. 2023, 9, 18. [Google Scholar] [CrossRef]
- Vinson, M.R.; Hawkins, C.P. Broad-scale geographical patterns in local stream insect genera richness. Ecography 2003, 26, 751–767. [Google Scholar] [CrossRef]
- Sandefur, H.; Cummings, E.; Johnston, R.; Matlock, M.; Gustafson, D. Analyzing correlations between stream benthic macroinvertebrate community structure, water quality, and watershed land use in the continental United States. Biol. Eng. Trans. 2014, 7, 169–182. [Google Scholar] [CrossRef]
- Camana, M.; Ortega, J.C.; Brejão, G.L.; Melo, A.S.; Dias, M.S.; Becker, F.G. A global meta-analysis of the effects of land use on the diversity of stream fish and macroinvertebrates. Aqua. Sci. 2024, 86, 86. [Google Scholar] [CrossRef]
- Karaouzas, I.; Gritzalis, K.C.; Skoulikidis, N. Land use effects on macroinvertebrate assemblages and stream quality along an agricultural river basin. Fres. Environ. Bull. 2007, 16, 645–653. [Google Scholar]
- Robinson, C.T.; Schuwirth, N.; Baumgartner, S.; Stamm, C. Spatial relationships between land-use, habitat, water quality and lotic macroinvertebrates in two Swiss catchments. Aqua. Sci. 2014, 76, 375–392. [Google Scholar] [CrossRef]
- Palt, M.; Le Gall, M.; Piffady, J.; Hering, D.; Kail, J. A metric-based analysis on the effects of riparian and catchment land use on macroinvertebrates. Sci. Total Environ. 2022, 816, 151590. [Google Scholar] [CrossRef]
- Sulikowski, T. An Examination of the Effects of Land Use and land Cover on Macroinvertebrate Communities at Two Landscape Scales. Master’s Thesis, The Faculty of Montclair State University, Montclair, NJ, USA, August 2013. [Google Scholar]
- Hoover, G.W.; Wood, J. Analysis of localized water chemistry and land use drivers of macroinvertebrate community structure in the Upper Ohio Watershed using multivariate modelling and ordination techniques. Proc. West Va. Acad. Sci. 2021, 93, 1. [Google Scholar] [CrossRef]
- Hrodey, P.J.; Sutton, T.M.; Frimpong, E.A.; Simon, T.P. Land-use impacts on watershed health and integrity in Indiana warmwater streams. Am. Mid. Nat. 2009, 161, 76–95. [Google Scholar] [CrossRef]
- Henderson, N.D.; Christian, A.D. Freshwater invertebrate assemblage composition and water quality assessment of an urban coastal watershed in the context of land-use land-cover and reach-scale physical habitat. Ecologies 2022, 3, 376–394. [Google Scholar] [CrossRef]
- Stepenuck, K.F.; Crunkilton, R.L.; Wang, L. Impacts of urban landuse on macroinvertebrate communities in southeastern Wisconsin streams. J. Am. Water Res. Assoc. 2007, 38, 1041–1051. [Google Scholar] [CrossRef]
- Wang, B.; Liu, D.; Liu, S.; Zhang, Y.; Lu, D.; Wang, L. Impacts of urbanization on stream habitats and macroinvertebrate communities in the tributaries of Qiangtang River, China. Hydrobiologia 2012, 680, 39–51. [Google Scholar] [CrossRef]
- Huang, J.; Gergel, S.E. Effectiveness of landscape indicators for explaining the variability of benthic macro-invertebrates in urban streams. J. Environ. Manag. 2023, 342, 118130. [Google Scholar] [CrossRef]
- Blann, K.L.; Anderson, J.L.; Sands, G.R.; Vondracek, B. Effects of agricultural drainage on aquatic ecosystems: A review. Crit. Rev. Environ. Sci. Technol. 2009, 39, 909–1001. [Google Scholar] [CrossRef]
- Seaber, P.R.; Kapinos, F.P.; Knapp, G.L. Hydrologic Unit Maps; Water Supply Paper 2294; U.S. Department of the Interior, U.S. Geological Survey US Government Printing Office: Washington, DC, USA, 1987. [Google Scholar] [CrossRef]
- Dewitz, J. United States National Land Cover Database (NLCD) 2019 Products, v. 3.0.; U.S. Department of the Interior, U.S. Geological Survey: Washington, DC, USA, 2024. [Google Scholar] [CrossRef]
- Wickham, J.; Stehman, S.V.; Sorenson, D.G.; Gass, L.; Dewitz, J.A. Thematic accuracy assessment of the NLCD 2019 land cover for the conterminous United States. GISci. Remote Sens. 2023, 60, 2181143. [Google Scholar] [CrossRef]
- Valayamkunnath, P.; Barlage, M.; Chen, F.; Gochis, D.J.; Franz, K.J. Mapping of 30-meter resolution tile-drained croplands using a geospatial modeling approach. Sci. Data 2020, 7, 257. [Google Scholar] [CrossRef]
- Smith, M.P.; Schiff, R.; Olivero, A.; MacBroom, J. The Active River Area. In A Conservation Framework for Protecting Rivers and Streams; The Nature Conservancy: Boston, MA, USA, 2008; Eastern U.S. Freshwater Program 11 Avenue De Lafayette, 5th Floor; Available online: https://www.conservationgateway.org/Files/Pages/active-river-area-conserv.aspx (accessed on 1 June 2024).
- Stroud Water Research Center. Model My Watershed [Software], v.1.35.0. 2024. Available online: https://wikiwatershed.org/ (accessed on 1 June 2024).
- Pitt, R.; Voorhees, J. SLAMM, the source loading and management model. In Wet-Weather Flow in the Urban Watershed; CRC Press: Boca Raton, FL, USA, 2002; pp. 103–139. [Google Scholar] [CrossRef]
- United States Soil Conservation Service. Urban Hydrology for Small Watersheds; Tech. Rep. No. 55 (TR-55); United States Soil Conservation Service: Washington, DC, USA, 1986. Available online: https://www.nrc.gov/docs/ML1421/ML14219A437.pdf (accessed on 1 June 2024).
- United States Environmental Protection Agency. Spreadsheet Tool for the Estimation of Pollutant Load (STEPL); Tetra Tech, Inc.: Fairfax, VA, USA, 2001. Available online: https://19january2021snapshot.epa.gov/nps/spreadsheet-tool-estimating-pollutant-loads-stepl (accessed on 1 June 2024).
- Ohio Environmental Protection Agency. Methods for Assessing Habitat in Flowing Waters: Using the Qualitative Habitat Evaluation Index (QHEI); Division of Surface Water, Ecological Assessment Section: Columbus, OH, USA, 2006. Available online: https://dam.assets.ohio.gov/image/upload/epa.ohio.gov/Portals/35/documents/QHEIManualJune2006.pdf (accessed on 1 June 2024).
- Barbour, M.T. Rapid Bioassessment Protocols for Use in Wadable Streams and Rivers. In Periphyton, Benthic Macroinvertebrates and Fish; US Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999. Available online: https://www3.epa.gov/region1/npdes/merrimackstation/pdfs/ar/AR-1164.pdf (accessed on 1 June 2024).
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Ohio Environmental Protection Agency. Field Methods for Evaluating Primary Headwater Streams in Ohio, Version 4.1; Ohio EPA Division of Surface Water: Columbus, OH, USA, 2020. Available online: https://dam.assets.ohio.gov/image/upload/epa.ohio.gov/Portals/35/wqs/headwaters/PHWHManual_2020_Ver_4_1_May_2020_Final.pdf (accessed on 1 June 2024).
- Sterling, J.L.; Rosemond, A.D.; Wenger, S.J. Watershed urbanization affects macroinvertebrate community structure and reduces biomass through similar pathways in Piedmont streams, Georgia, USA. Freshw. Sci. 2016, 35, 676–688. [Google Scholar] [CrossRef]
- Weigel, B.M.; Lyons, J.; Paine, L.K.; Dodson, S.I.; Undersander, D.J. Using stream macroinvertebrates to compare riparian land use practices on cattle farms in southwestern Wisconsin. J. Freshw. Ecol. 2000, 15, 93–106. [Google Scholar] [CrossRef]
- Wang, L.; Kanehl, P. Influences of watershed urbanization and instream habitat on macroinvertebrates in cold water streams. J. Am. Water Res. Assoc. 2003, 39, 1181–1196. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HUC12 Watershed | Active River Area (ARA) | |||||
|---|---|---|---|---|---|---|
| Area (km2) | Imperv% | Tiled% | Area (km2) | Imperv% | Tiled% | |
| Developed | ||||||
| Total (27) | 79 ± 5.7 | 61 ± 4.6 | 11 ± 2.5 | 21 ± 2.6 | 9.1 ± 1.0 | 1.8 ± 0.5 |
| Sampled (10) | 82 ± 9.1 | 64 ± 9.0 | 5.7 ± 3.0 | 37 ± 4.7 | 57 ± 9.1 | 3.2 ± 1.2 |
| Agriculture | ||||||
| Total (20) | 86 ± 8.4 | 8.6 ± 1.4 | 55 ± 1.5 | 23 ± 4.1 | 3.2 ± 1.2 | 12 ± 1.4 |
| Sampled (10) | 76 ± 9.6 | 15 ± 6.3 | 30 ± 1.8 | 37 ± 5.1 | 17 ± 5.7 | 18 ± 3.1 |
| Less Developed | ||||||
| Total (24) | 79 ± 5.2 | 8.3 ± 0.5 | 6.6 ± 1.0 | 19 ± 1.8 | 1.8 ± 0.2 | 1.4 ± 0.2 |
| Sampled (10) | 71 ± 6.9 | 12 ± 1.8 | 11 ± 2.6 | 26 ± 4.0 | 17 ± 3.5 | 7.1 ± 1.2 |
| Phylum | Class | Order | Family | Abundance | Locations |
|---|---|---|---|---|---|
| Annelida | Clitellata | Rhynchobdellida | Glossiphoniidae | 187 | 22 |
| Clitellata | Other | 4 | 3 | ||
| Arthropoda | Arachnida | Araneae | Pisauridae | 2 | 1 |
| Insecta | Coleoptera | Dryopidae | 10 | 6 | |
| Dytiscidae | 4 | 4 | |||
| Elmidae | 74 | 24 | |||
| Gyrinidae | 28 | 4 | |||
| Haliplidae | 4 | 3 | |||
| Hydrophilidae | 1 | 1 | |||
| Psephenidae | 241 | 17 | |||
| Ptilodactylidae | 1 | 1 | |||
| Diptera | Athericidae | 1 | 1 | ||
| Ceratopogonidae | 1 | 1 | |||
| Chironomidae | 59 | 20 | |||
| Ephydridae | 1 | 1 | |||
| Limoniidae | 2 | 2 | |||
| Simuliidae | 257 | 21 | |||
| Tipulidae | 24 | 11 | |||
| Ephemeroptera | Ameletidae | 11 | 3 | ||
| Baetidae | 213 | 26 | |||
| Caenidae | 1 | 1 | |||
| Ephemerellidae | 17 | 6 | |||
| Heptageniidae | 279 | 26 | |||
| Leptophlebiidae | 10 | 6 | |||
| Potamanthidae | 1 | 1 | |||
| Siphlonuridae | 3 | 3 | |||
| Hemiptera | Gerridae | 17 | 8 | ||
| Veliidae | 12 | 6 | |||
| Megaloptera | Corydalidae | 2 | 2 | ||
| Odonata | Aeshnidae | 6 | 6 | ||
| Calopterygidae | 3 | 2 | |||
| Coenagrionidae | 10 | 6 | |||
| Plecoptera | Capniidae | 1 | 1 | ||
| Chloroperlidae | 33 | 7 | |||
| Leuctridae | 3 | 1 | |||
| Nemouridae | 10 | 5 | |||
| Peltoperlidae | 1 | 1 | |||
| Perlidae | 27 | 7 | |||
| Perlodidae | 15 | 5 | |||
| Taeniopterygidae | 2 | 2 | |||
| Trichoptera | Apataniidae | 1 | 1 | ||
| Brachycentridae | 32 | 8 | |||
| Dipseudopsidae | 19 | 6 | |||
| Glossosomatidae | 27 | 2 | |||
| Helicopsychidae | 226 | 12 | |||
| Hydropsychidae | 814 | 32 | |||
| Hydroptilidae | 13 | 5 | |||
| Lepidostoma | 4 | 1 | |||
| Leptoceridae | 3 | 2 | |||
| Limnephilidae | 33 | 4 | |||
| Philopotamidae | 275 | 27 | |||
| Polycentropodidae | 18 | 4 | |||
| Psychomyiidae | 9 | 4 | |||
| Rhyacophilidae | 23 | 5 | |||
| Thremmatidae | 2 | 1 | |||
| other | 97 | 23 | |||
| Malacostraca | Amphipoda | Gammaridae | 7 | 5 | |
| Decapoda | Cambaridae | 93 | 43 | ||
| Mollusca | Bivalvia | Unionida | Unionidae | 43 | 13 |
| Gastropoda | Architaenioglossa | Viviparidae | 3 | 2 | |
| Basommatophora | Ancylidae | 16 | 4 | ||
| Lymnaeidae | 15 | 4 | |||
| Physidae | 179 | 25 | |||
| Planorbidae | 32 | 14 | |||
| Caenogastropoda | Pleuroceridae | 15 | 1 | ||
| Gyraulus | Planorbidae | 2 | 1 | ||
| Heterostropha | Valvatidae | 1 | 1 | ||
| Littorinimorpha | Amnicolidae | 2 | 1 | ||
| Hydrobiidae | 2 | 2 | |||
| Neotaenioglossa | Pleuroceridae | 334 | 15 | ||
| 334 | Pleurocera | 2 | 1 | ||
| Sorbeoconcha | Arionidae | 4 | 3 | ||
| Stylommatophora | 7 | 1 | |||
| Other | 26 | 4 |
| Macroinvertebrate Community Metrics | |||||
|---|---|---|---|---|---|
| Total Individuals | Total Taxa | Shannon Diversity | HMFEI Score | EPT Taxa | |
| HUC12 Level | |||||
| % Developed | −0.40 ** | −0.54 ** | −0.37 ** | −0.57 ** | −0.50 ** |
| % Agriculture | 0.56 ** | 0.41 ** | 0.20 | 0.38 | 0.25 |
| % Undeveloped | −0.06 | 0.45 ** | 0.42 ** | 0.57 ** | 0.63 ** |
| Hydrologic Load | −0.13 | −0.51 ** | −0.39 ** | −0.60 ** | −0.60 ** |
| Nitrogen Load | 0.25 | −0.28 | −0.26 | −0.40 ** | −0.47 ** |
| Phosphorus Load | 0.36 ** | −0.20 | −0.22 | −0.34 ** | −0.42 ** |
| Sediment Load | −0.20 | −0.29 * | −0.21 | −0.28 * | −0.26 |
| Reach Level | |||||
| QHEI Score | −0.05 | 0.15 | 0.35 ** | 0.23 | 0.22 |
| Habitat Score | 0.09 | 0.28 | 0.35 ** | 0.30 * | 0.28 |
| Sinuosity | 0.02 | 0.37 ** | 0.32 * | 0.42 ** | 0.43 ** |
| Canopy Cover | −0.01 | 0.19 | 0.23 | 0.23 | 0.26 |
| Substrate Score | −0.02 | −0.09 | −0.10 | −0.08 | −0.13 |
| % Riffle | −0.24 | −0.12 | −0.01 | −0.05 | 0.05 |
| Periphyton Cover | 0.43 ** | 0.15 | 0.08 | 0.15 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spieles, D.; Krashes, Z.; Nguyen, K.; Rodgers, S.; Ruiz, L.; Vigilante, M. Relationships Between Land Use and Stream Macroinvertebrate Biotic Integrity in Central Ohio, USA. Water 2025, 17, 895. https://doi.org/10.3390/w17060895
Spieles D, Krashes Z, Nguyen K, Rodgers S, Ruiz L, Vigilante M. Relationships Between Land Use and Stream Macroinvertebrate Biotic Integrity in Central Ohio, USA. Water. 2025; 17(6):895. https://doi.org/10.3390/w17060895
Chicago/Turabian StyleSpieles, Douglas, Zoe Krashes, Khiem Nguyen, Summer Rodgers, Lillian Ruiz, and Marco Vigilante. 2025. "Relationships Between Land Use and Stream Macroinvertebrate Biotic Integrity in Central Ohio, USA" Water 17, no. 6: 895. https://doi.org/10.3390/w17060895
APA StyleSpieles, D., Krashes, Z., Nguyen, K., Rodgers, S., Ruiz, L., & Vigilante, M. (2025). Relationships Between Land Use and Stream Macroinvertebrate Biotic Integrity in Central Ohio, USA. Water, 17(6), 895. https://doi.org/10.3390/w17060895