
Soil Infiltration Characteristics and Driving Mechanisms of Three Typical Forest Types in Southern Subtropical China

Abstract
1. Introduction
2. Materials and Methods
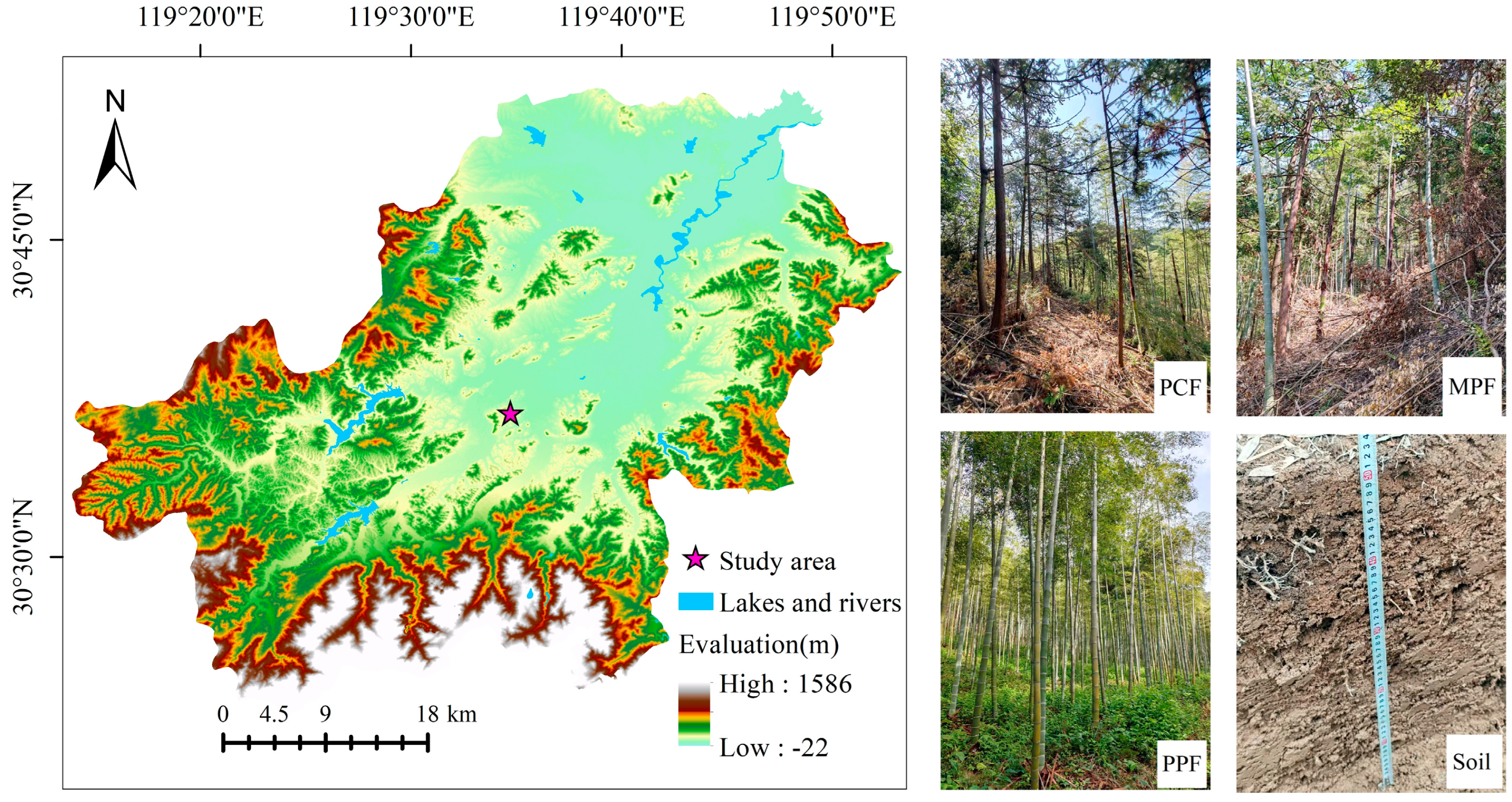
2.1. Study Area
2.2. Experimental Design
2.2.1. Plot Establishment
2.2.2. Soil Infiltration Measurements
2.2.3. Soil Sampling and Root Analysis
2.3. Calculation Methods
- (1)
- Root Morphological Parameters
- (2)
- Soil Infiltration Rate Calculation
2.4. Data Processing and Analysis
3. Results
3.1. Soil Infiltration Characteristics Across Three Stand Types
3.2. Soil Physicochemical Properties and Root Morphological Traits Across Stand Types
3.2.1. Soil Physicochemical Characteristics
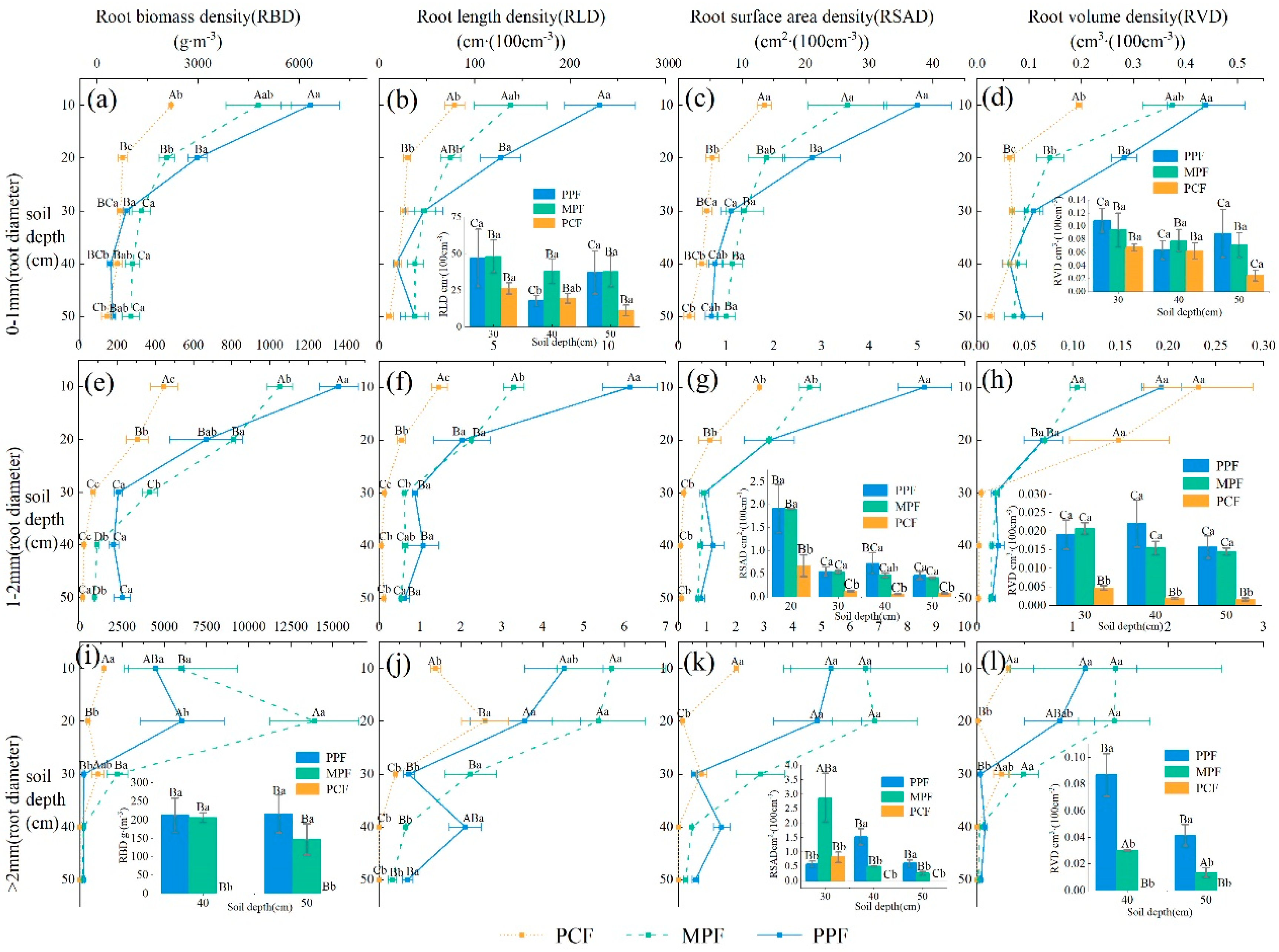
3.2.2. Root Traits Across Stand Types
3.2.3. Relationships Between Soil Properties and Root Traits
3.3. Analysis of Influencing Factors on Soil Infiltration Rates
3.3.1. Principal Component Analysis
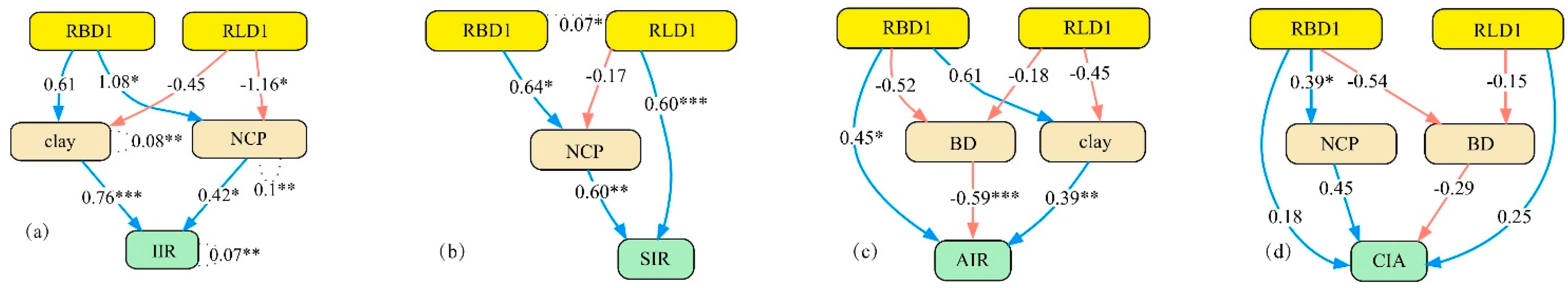
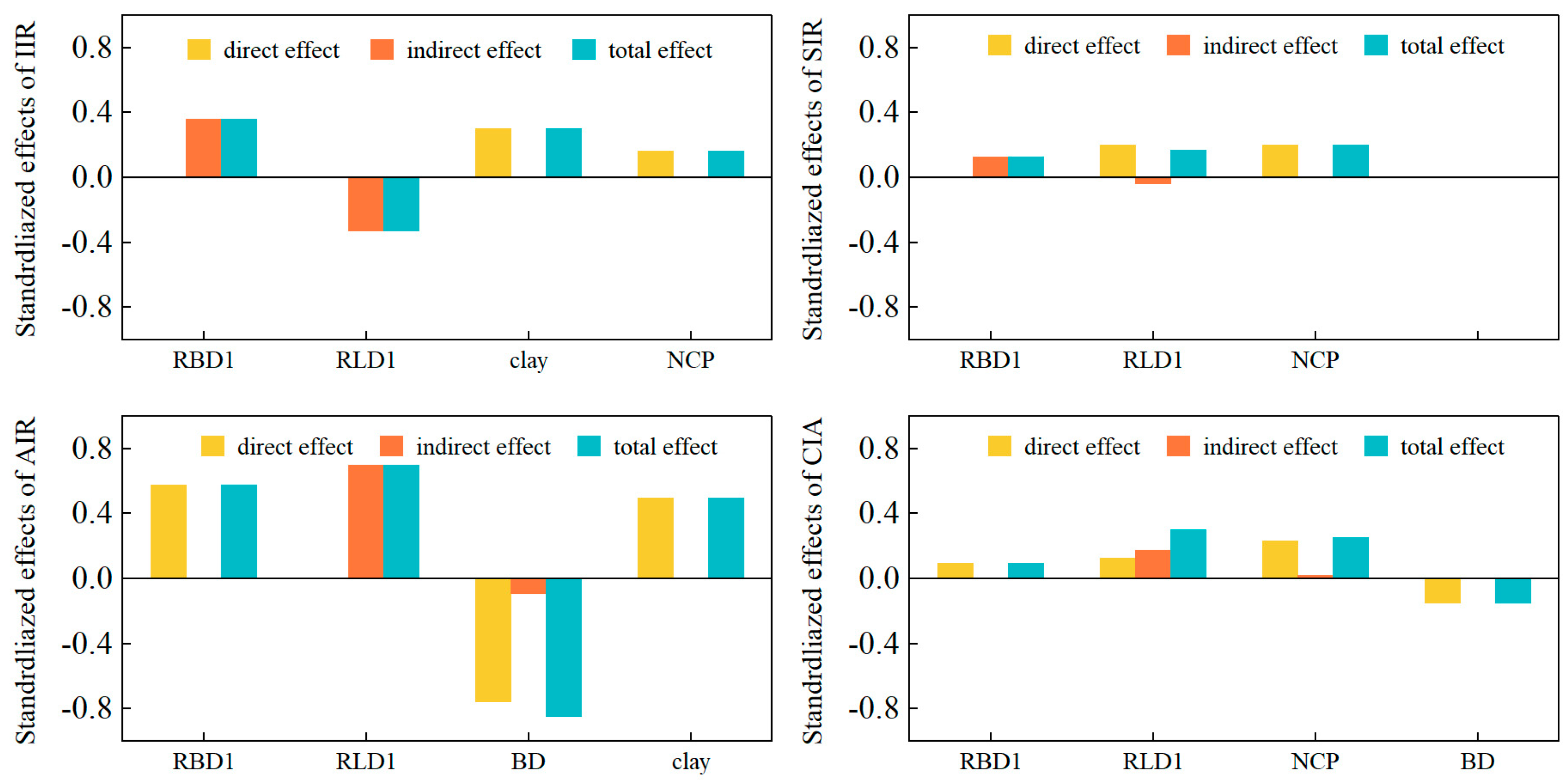
3.3.2. Structural Equation Model of the Infiltration Process
4. Discussion
4.1. Effects of Forest Types on Root Traits and Soil Properties
4.2. Mechanisms of Soil Infiltration Regulation by Root–Soil Interactions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ren, Z.P.; Zhu, L.J.; Wang, B.; Chen, S.D. Soil hydraulic conductivity as affected by vegetation restoration age on the Loess Plateau, China. J. Arid Land 2016, 8, 546–557. [Google Scholar] [CrossRef]
- Li, J.B.; Zang, F.C.; Duan, C.X. Study on soil infiltration characteristics and water and nitrogen migration under magnetization irrigation of nitrogen fertilizer solution. Trans. Chin. Soc. Agric. Mach. 2022, 53, 316–324. (In Chinese) [Google Scholar]
- Liu, Y.; Guo, L.; Huang, Z.; Wu, G.L. Root morphological characteristics and soil water infiltration capacity in semi-arid artificial grassland soils. Agric. Water Manag. 2022, 235, 106153. [Google Scholar] [CrossRef]
- Qiu, D.X.; Xu, R.R.; Gao, P.; Mu, X.M. Effect of vegetation restoration type and topography on soil water storage and infiltration capacity in the Loess Plateau, China. CATENA 2024, 241, 108079. [Google Scholar] [CrossRef]
- Zhu, P.Z.; Zhang, G.H.; Wang, C.S.; Wang, C.S.; Chen, S.Q.; Wan, Y.Q. Variation in soil infiltration properties under different land use/cover in the black soil region of Northeast China. Int. Soil Water Conserv. Res. 2024, 12, 379–387. [Google Scholar] [CrossRef]
- Neris, C.; Jimenez, J.; Fuentes, J. Vegetation and land-use effects on soil properties and water infiltration of Andisols in Tenerife (Canary Islands, Spain). CATENA 2012, 98, 55–62. [Google Scholar] [CrossRef]
- Qiu, D.X.; Xu, R.R.; Wu, C.X.; Mu, X.M.; Zhao, G.J.; Gao, P. Effects of vegetation restoration on soil infiltrability and preferential flow in hilly gully areas of the Loess Plateau, China. CATENA 2023, 221, 106770. [Google Scholar] [CrossRef]
- Robinson, D.A.; Nemes, A.; Reinsch, S.; Radbourne, A.; Bentley, L.; Keith, A.M. Global meta-analysis of soil hydraulic properties on the same soils with differing land use. Sci. Total Environ. 2022, 852, 158506. [Google Scholar] [CrossRef]
- Basset, C.; Abou, N.M.; Ghezzehei, T.; Hao, X.X.; Daccache, A. How does soil structure affect water infiltration? A meta-data systematic review. Soil Tillage Res. 2023, 226, 105577. [Google Scholar] [CrossRef]
- Zhang, W.J.; Zhu, X.A.; Xiong, X.; Wu, T.; Zhou, S.D.; Lie, Z.Y.; Jiang, X.J.; Liu, J.X. Changes in soil infiltration and water flow paths: Insights from subtropical forest succession sequence. CATENA 2023, 221, 106748. [Google Scholar] [CrossRef]
- Peng, J.; Wu, Y.; Geng, T. Soil pore dynamics and infiltration characteristics as affected by cultivation duration for Mollisol in northeast China. Geoderma 2024, 449, 117021. [Google Scholar] [CrossRef]
- Horn, R. The Effect of Aggregation of Soils on Water, Gas, and Heat Transport. In Flux Control in Biological Systems; Academic Press: Cambridge, MA, USA, 1994; pp. 335–361. [Google Scholar]
- Huang, Z.; Tian, F.P.; Wu, G.L.; Liu, Y.; Dang, Z.Q. Legume Grasslands Promote Precipitation Infiltration better than Gramineous Grasslands in arid Regions. Land Degrad. Dev. 2017, 28, 309–316. [Google Scholar] [CrossRef]
- Wang, P.P.; Su, X.M.; Zhou, Z.C.; Wang, N.; Liu, J.; Zhu, B.B. Differential effects of soil texture and root traits on the spatial variability of soil infiltrability under natural revegetation in the Loess Plateau of China. CATENA 2023, 220, 106693. [Google Scholar] [CrossRef]
- Cui, Z.; Huang, Z.; Liu, Y.; López, V.M.; Wu, G.L. Natural compensation mechanism of soil water infiltration through decayed roots in semi-arid vegetation species. Sci. Total Environ. 2021, 819, 151985. [Google Scholar] [CrossRef]
- Shi, X.Q.; Qin, T.L.; Yan, D.H.; Tian, F.Q.; Wang, H. A meta-analysis on effects of root development on soil hydraulic properties. Geoderma 2021, 403, 115363. [Google Scholar] [CrossRef]
- Bodner, G.; Leitner, D.; Kaul, H.P. Coarse and fine root plants affect pore size distributions differently. Plant Soil 2014, 380, 133–151. [Google Scholar] [CrossRef]
- Gerke, H.H. Preferential flow descriptions for structured soils. Z. Pflanzenernähr. Bodenk 2006, 169, 382–400. [Google Scholar] [CrossRef]
- Jiang, X.J.; Liu, W.; Chen, C.; Liu, J.; Yuan, Z.Q.; Jin, B.; Yu, X. Effects of three morphometric features of roots on soil water flow behavior in three sites in China. Geoderma 2018, 320, 161–171. [Google Scholar] [CrossRef]
- Hao, H.X.; Wei, Y.J.; Cao, D.N.; Guo, Z.L.; Shi, Z.H. Vegetation restoration and fine roots promote soil infiltrability in heavy-textured soils. Soil Tillage Res. 2020, 198, 104542. [Google Scholar] [CrossRef]
- Lu, C.; Wang, F.; Lin, Y.Y.; Long, Q.; Zhao, Y.L.; Han, J.Q.; Ge, W.Y.; Chen, H. Changes in preferential flow caused by root effects in black locust plantations of different stand ages in the semi-arid region of the Loess Plateau. J. Hydrol. 2024, 634, 131086. [Google Scholar]
- Li, Y.C.; Liang, X.; Tang, C.X. Moso bamboo invasion into broadleaf forests is associated with greater abundance and activity of soil autotrophic bacteria. Plant Soil 2018, 428, 163–177. [Google Scholar] [CrossRef]
- Liu, L.; Gudmundsson, L.; Hauser, M.; Qin, D.; Seneviratne, S.I. Soil moisture dominates dryness stress on ecosystem production globally. Nat. Commun. 2020, 11, 4892. [Google Scholar] [CrossRef]
- Bouwer, H. Intake Rate: Cylinder Infiltrometer. In Methods of Soil Analysis; Klute, A., Ed.; Wiley: Hoboken, NJ, USA, 1986. [Google Scholar]
- Zhang, Y.; Niu, J.; Zhang, M.; Xiao, Z.X.; Zhu, W.L. Interaction Between Plant Roots and Soil Water Flow in Response to Preferential Flow Paths in Northern China. Land Degrad. Dev. 2017, 28, 648–663. [Google Scholar] [CrossRef]
- Xu, Q.F.; Liang, C.F.; Chen, J.H.; Li, Y.C.; Qin, H.Q. Rapid bamboo invasion (expansion) and its effects on biodiversity and soil processes. Glob. Ecol. Conserv. 2020, 21, e00787. [Google Scholar] [CrossRef]
- Liu, J.; Yang, Q.P.; Song, Q.N.; Yu, D.K. Strategy of fine root expansion of Phyllostachys pubescens population into evergreen broad- leaved forest. Chin. J. Plant Ecol. 2013, 37, 230–238. (In Chinese) [Google Scholar] [CrossRef]
- Liu, J.; Yang, Q.P.; Yu, D.K.; Yang, G.Y.; Qi, H.Y.; Shi, J.M. Contribution of fine root to soil nutrient heterogeneity at two sides of the bamboo and broad-leaved forest interface: Contribution of fine root to soil nutrient heterogeneity at two sides of the bamboo and broad-leaved forest interface. Chin. J. Plant Ecol. 2014, 37, 739–749. [Google Scholar] [CrossRef]
- Ni, H.J.; Su, W.H. Spatial distribution of fine root traits in relation to soil properties and aggregate stability of intensively managed Moso bamboo (Phyllostachys edulis) plantations in subtropical China. Plant Soil 2024, 498, 487–503. [Google Scholar] [CrossRef]
- Cai, C.J.; Fan, S.H.; Liu, X.Z. Fine root adaptation strategies during the expansion of moso bamboo to Chinese fir forest. Chin. J. Ecol. 2019, 38, 967–972. (In Chinese) [Google Scholar]
- Chen, J.Y.; Zhu, Z.Y.; Hu, H.B.; Luo, Z.X.; Yang, X.Y.; Jia, X.C.; Fang, P.; Mansour, L. Soil quality assessment of oak forests invaded by moso bamboo (Phyllostachys edulis) in the northern subtropics. Glob. Ecol. Conserv. 2024, 51, e02945. [Google Scholar] [CrossRef]
- Song, Q.N.; Yang, Q.P.; Liu, J. Effects of bamboo expansion on soil nitrogen mineralization and availability in evergreen broad-leaved forest. Chin. J. Appl. Ecol. 2013, 24, 338–344. (In Chinese) [Google Scholar]
- Bai, S.; Conant, R.T.; Zhou, G.; Wang, Y.; Wang, N.; Li, Y.; Zhang, K. Effects of Moso bamboo encroachment into native, broad-leaved forests on soil carbon and nitrogen pools. Sci. Rep. 2016, 6, 31480. [Google Scholar] [CrossRef]
- Xie, Y.Y.; Guo, Z.W.; Lin, S.Y.; Zuo, K.Y.; Yang, L.T.; Xu, S.; Gu, S.; Chen, S.L. Soil particle composition and water infiltration characteristics during the succession of understory vegetation in Moso bamboo forest. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2024, 48, 108–116. (In Chinese) [Google Scholar]
- Xu, D.W.; Liu, J.F.; He, Z.S. Community species diversity characteristics of Phyllostachys heterocycla after expansion into Cunninghamia lanceolata forest. J. For. Environ. 2019, 39, 37–41. (In Chinese) [Google Scholar]
- Wu, J.S.; Jiang, P.K.; Wang, Z.L. Effects of bamboo expansion on soil fertility in Tianmu Mountain National Nature Reserve. Acta Agric. Univ. Jiangxiensis 2008, 689–692. (In Chinese) [Google Scholar]
- Alameda, D.; Villar, R. Linking root traits to plant physiology and growth in Fraxinus angustifolia Vahl. seedlings under soil compaction conditions. Environ. Exp. Bot. 2012, 79, 49–57. [Google Scholar] [CrossRef]
- Ali, H.E.; Reineking, B.; Münkemüller, T. Effects of plant functional traits on soil stability: Intraspecific variability matters. Plant Soil 2017, 411, 359–375. [Google Scholar] [CrossRef]
- Loades, K.W.; Bengough, A.G.; Bransby, M.F.; Hallett, P.D. Planting density influence on fibrous root reinforcement of soils. Ecol. Eng. 2010, 36, 276–284. [Google Scholar] [CrossRef]
- Gyssels, G.; Poesen, J.; Bochet, E.; Li, Y. Impact of plant roots on the resistance of soils to erosion by water: A review. Prog. Phys. Geogr. 2005, 29, 189–217. [Google Scholar] [CrossRef]
- Redelstein, R.; Dinter, T.; Hertel, D.; Leuschner, C. Effects of Inundation, Nutrient Availability and Plant Species Diversity on Fine Root Mass and Morphology Across a Saltmarsh Flooding Gradient. Front. Plant Sci. 2018, 9, 98. [Google Scholar] [CrossRef]
- Qu, Y.; Wu, Q.; Khan, F.U.; Wang, J.F.; Ren, X.Z.; Chai, X.H.; Xu, X.X.; Du, F. The impact of root systems on soil macropore abundance and soil infiltration capacity. Plant Soil 2025. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; Gerlinde, B.D.D.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef]
- Wu, G.L.; Liu, Y.; Yang, Z.; Cui, Z.; Deng, L.; Chang, X.F.; Shi, Z.H. Root channels to indicate the increase in soil matrix water infiltration capacity of arid reclaimed mine soils. J. Hydrol. 2017, 546, 133–139. [Google Scholar] [CrossRef]
- Bogner, C.; Gaul, D.; Kolb, A.; Schmiedinger, I.; Huwe, B. Investigating flow mechanisms in a forest soil by mixed-effects modelling. Eur. J. Soil Sci. 2010, 61, 1079–1090. [Google Scholar] [CrossRef]
- Peter, G.; Benjamin, L.; Peter, L. Preferential Flow Dynamics and Plant Rooting Systems. In Hydropedology: Synergistic Integration of Soil Science and Hydrology; Academic Press: Cambridge, MA, USA, 2012; pp. 121–141. [Google Scholar]
- Wang, P.P.; Liu, Q.; Zhou, Z.C.; Liu, J.; Cao, L.G.; Wang, N.; Cao, Y.Y. Distinct variations of soil infiltrability of contrasting root types in a temperate mosaic-pattern grassland in northern China. CATENA 2024, 243, 108174. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, S.Y.; Zhao, G.J.; Fan, J.J.; Yang, M.Y.; Tian, P.; Mu, X.M.; Geng, R. Variations of soil infiltration in response to vegetation restoration and its influencing factors on the Loess Plateau. Environ. Manag. 2024, 372, 123356. [Google Scholar] [CrossRef]
- Huang, Y.; Xiong, T.; Zhao, M.Q.; Deng, Y.S.; Yang, G.R.; Ban, Y.Y.; Lei, T.W.; Yu, X.X.; Huang, Y.H. Influence of soil properties and near-surface roots on soil infiltration process in short-rotation eucalyptus plantations in southern subtropical China. CATENA 2024, 234, 107606. [Google Scholar] [CrossRef]
- Zhang, H.J.; Fu, X.; Ma, S.W. Soil root distribution and water infiltration characteristics of different crop farmlands in Simian Mountain, Chongqing. Sci. Soil Water Conserv. 2023, 21, 137–144. (In Chinese) [Google Scholar]
- Assouline, S. Infiltration into soils: Conceptual approaches and solutions. Water Resour. Res. 2013, 49, 1755–1772. [Google Scholar] [CrossRef]
- Schweizer, S.A.; Bucka, F.; Graf-Rosenfellner, M.; Knabner, I.K. Soil microaggregate size composition and organic matter distribution as affected by clay content. Geoderma 2019, 355, 113901. [Google Scholar] [CrossRef]
- Zhao, Y.G.; Wu, P.T.; Zhao, S.W.; Feng, H. Variation of soil infiltrability across a 79-year chronosequence of naturally restored grassland on the Loess Plateau, China. J. Hydrol. 2013, 504, 94–103. [Google Scholar] [CrossRef]
- Lozano-Baez, S.E.; Cooper, M.; Meli, P. Land restoration by tree planting in the tropics and subtropics improves soil infiltration, but some critical gaps still hinder conclusive results. For. Ecol. Manag. 2019, 444, 89–95. [Google Scholar] [CrossRef]
- Wu, G.L.; Yang, Z.; Cui, Z.; Liu, Y..; Fang, N.-F.; Shi, Z.-H. Mixed artificial grasslands with more roots improved mine soil infiltration capacity. J. Hydrol. 2016, 535, 54–60. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Liu, Y.H.; Yang, M.Y.; Tian, P.; Mu, X.M.; Zhao, G.J. Impact of vegetation restoration on preferential flow and soil infiltration capacity in the hilly region of the Loess Plateau. J. Hydrol. Reg. Stud. 2025, 59, 102333. [Google Scholar] [CrossRef]
- Angers, D.A.; Caron, J. Plant-induced Changes in Soil Structure: Processes and Feedbacks. Biogeochemistry 1998, 42, 55–72. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, L.; Xu, X.; Wang, Y.; Liu, W.; Schwarzel, K. Impact of land-use changes on soil hydraulic properties of Calcaric Regosols on the Loess Plateau, NW China. Plant Nutr. Soil Sci. 2015, 178, 486–498. [Google Scholar] [CrossRef]
- Cui, Z.; Wu, G.L.; Huang, Z.; Liu, Y. Fine roots determine soil infiltration potential than soil water content in semi-arid grassland soils. J. Hydrol. 2019, 578, 124023. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, X.; Zakari, S.; Chen, C.; Liu, W.; Jiang, X.-J. Assessing the Effects of Plant Roots on Soil Water Infiltration Using Dyes and Hydrus-1D. Forests 2022, 13, 1095. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand | Altitude | Slope (°) | Aspect of Slope | Plant Density | Mean Tree Height (m) | Mean DBH (cm) | Canopy Density |
|---|---|---|---|---|---|---|---|
| PPF | 73 | 10° | West | 2100 | 16 | 12 | 0.75 |
| MPF | 75 | 10° | South-west | 1350 | 12 | 10 | 0.85 |
| PCF | 76 | 10° | South-west | 925 | 10 | 10 | 0.6 |
| Soil Depth (cm) | Forest Types | W | BD (g∙(100 cm−3)) | TP (%) | CP (%) | NCP (%) | SOC (g∙kg−1) | pH | TN (g∙kg−1) | Sandy (%) | Silty (%) | Clay (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0–10 | PPF | 0.23 ± 0.01 a | 1.11 ± 0.08 a | 49.11 ± 3.47 a | 38.64 ± 2.25 a | 10.47 ± 2.02 a | 29.02 ± 4.05 a | 4.91 ± 0.37 a | 2.46 ± 0.28 a | 69 ± 4.21 a | 19.94 ± 2.8 a | 11.07 ± 1.85 ab |
| MPF | 0.22 ± 0.01 a | 1.19 ± 0.03 a | 42.82 ± 4.71 b | 21.32 ± 14.25b | 8.72 ± 0.99 ab | 22.54 ± 2.03 a | 4.44 ± 0.46 ab | 1.71 ± 0.31 a | 65.03 ± 6.43 a | 21.29 ± 6.18 a | 13.69 ± 2.86 a | |
| PCF | 0.22 ± 0.01 a | 1.2 ± 0.05 a | 48.12 ± 3.26 ab | 40.78 ± 4.05 a | 7.34 ± 0.83 b | 28.22 ± 15.21 a | 4.28 ± 0.1 b | 1.64 ± 0.78 a | 69.66 ± 2.69 a | 20.47 ± 2.82 a | 9.88 ± 0.83 b | |
| 10–20 | PPF | 0.21 ± 0.02 a | 1.28 ± 0.06 b | 42.43 ± 2.15 a | 35 ± 1.25 a | 7.43 ± 1.03 a | 17.7 ± 3.4 a | 4.78 ± 0.1 a | 1.32 ± 0.32 a | 62.81 ± 11.5 a | 23.31 ± 11.8 a | 13.88 ± 2.08 a |
| MPF | 0.2 ± 0.01 a | 1.41 ± 0.06 b | 44.04 ± 5.54 a | 37.66 ± 5.63b | 6.38 ± 0.09 b | 13.77 ± 3.35 a | 4.75 ± 0.18 a | 0.91 ± 0.21 a | 64.73 ± 2.55 a | 23.34 ± 2.15b | 11.92 ± 3.54 b | |
| PCF | 0.22 ± 0.01 a | 1.46 ± 0.02 a | 44.79 ± 2.09 a | 38.89 ± 2.03 a | 5.9 ± 0.06 c | 12.79 ± 0.6 b | 4.96 ± 0.23 a | 0.84 ± 0.04 b | 62.57 ± 3 b | 26.41 ± 4.64 ab | 11.03 ± 2.04 b | |
| 20–30 | PPF | 0.2 ± 0.02 a | 1.27 ± 0.01 b | 39.89 ± 4.1 a | 31.87 ± 2.97 a | 5.82 ± 1.87 a | 11.19 ± 1.32 a | 5.17 ± 0.27 a | 0.9 ± 0.25 a | 56.91 ± 8.27 a | 31.52 ± 6.78 a | 11.57 ± 3.44 a |
| MPF | 0.19 ± 0 a | 1.33 ± 0.02 b | 39.63 ± 2.84 a | 27.95 ± 11.2 b | 7.27 ± 0.8 b | 11.08 ± 1.76 a | 5.2 ± 0.18 b | 0.77 ± 0.19 b | 60.81 ± 6.69 b | 26.25 ± 8.54 b | 12.94 ± 2.12 b | |
| PCF | 0.21 ± 0 a | 1.47 ± 0.1 a | 44.19 ± 3.27 a | 37.52 ± 3.3 ab | 6.66 ± 0.11 b | 8.42 ± 4.51 a | 5.2 ± 0.1 b | 0.62 ± 0.21 b | 59.62 ± 5.02 b | 26.47 ± 5.42 b | 13.92 ± 0.83 b | |
| 30–40 | PPF | 0.19 ± 0.02 a | 1.37 ± 0.03 a | 42.14 ± 3.29 a | 37.59 ± 3.36 a | 4.56 ± 0.7 a | 7.35 ± 1.55 a | 5.43 ± 0.34 a | 0.53 ± 0.09 a | 40.89 ± 7.81 a | 49.3 ± 3.52 a | 9.81 ± 4.83 a |
| MPF | 0.18 ± 0.01b | 1.39 ± 0.02 b | 36.46 ± 2.71 a | 31.03 ± 2.42 ab | 5.43 ± 0.43 b | 6.68 ± 1.12 b | 5.24 ± 0.3 a | 0.48 ± 0.07 a | 44.05 ± 10.68 ab | 40.37 ± 10.56 b | 15.58 ± 1.2 b | |
| PCF | 0.2 ± 0.01 a | 1.45 ± 0.01 a | 39.69 ± 2.92 b | 34.43 ± 2.83 b | 5.26 ± 0.48 c | 7.65 ± 0.74 ab | 5.49 ± 0.18 a | 0.57 ± 0.07 a | 50.09 ± 1.06 b | 38.04 ± 2 b | 11.87 ± 2.1 b |
| Soil Depth (cm) | Root Traits | Soil Physical Index | Soil Chemical Index | Soil Texture | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W | BD (g∙(100 cm−3)) | TP (%) | CP (%) | NCP (%) | SOC (g∙kg−1) | pH | TN (g∙kg−1) | Sandy (%) | Silty (%) | Clay (%) | ||
| 0–10 cm | RBD (<2 mm) | 0.249 | −0.713 ** | 0.072 | 0.104 | 0.626 * | 0.028 | 0.568 * | 0.398 | −0.135 | 0.109 | 0.087 |
| RLD (<2 mm) | 0.196 | −0.841 ** | 0.185 | 0.284 | 0.671 ** | 0.074 | 0.601 * | 0.392 | −0.083 | 0.101 | −0.003 | |
| RSAD (<2 mm) | 0.144 | −0.734 ** | 0.197 | 0.219 | 0.734 ** | 0.065 | 0.468 | 0.37 | −0.144 | 0.134 | 0.064 | |
| RVD (<2 mm) | 0.178 | −0.818 ** | 0.002 | 0.189 | 0.616 * | −0.259 | 0.413 | 0.016 | 0.065 | 0.019 | −0.162 | |
| RBD (>2 mm) | 0.101 | −0.067 | −0.275 | −0.017 | 0.204 | −0.117 | −0.049 | −0.037 | −0.022 | −0.178 | 0.339 | |
| RLD (>2 mm) | 0.116 | −0.358 | −0.297 | −0.19 | 0.475 | −0.161 | 0.114 | −0.003 | −0.083 | −0.08 | 0.299 | |
| RSAD (>2 mm) | 0.115 | −0.182 | −0.261 | −0.023 | 0.296 | −0.122 | 0.003 | −0.026 | −0.029 | −0.147 | 0.301 | |
| RVD (>2 mm) | 0.108 | −0.04 | −0.247 | 0.044 | 0.149 | −0.104 | −0.053 | −0.032 | −0.008 | −0.182 | 0.318 | |
| 10–20 cm | RBD (<2 mm) | −0.209 | −0.610 * | 0.11 | 0.068 | 0.075 | 0.656 ** | 0.052 | 0.547 * | −0.329 | 0.288 | 0.247 |
| RLD (<2 mm) | 0.035 | −0.563 * | 0.136 | 0.173 | −0.191 | 0.646 ** | 0.168 | 0.556 * | −0.457 | 0.412 | 0.309 | |
| RSAD (<2 mm) | −0.22 | −0.496 | 0.082 | 0.04 | 0.091 | 0.650 ** | −0.088 | 0.5 | −0.174 | 0.093 | 0.305 | |
| RVD (<2 mm) | −0.099 | −0.136 | 0.527 * | 0.517 * | −0.242 | 0.248 | 0.301 | 0.22 | −0.336 | 0.296 | 0.248 | |
| RBD (>2 mm) | −0.301 | −0.283 | −0.232 | −0.404 | 0.669 ** | 0.611 * | −0.203 | 0.3 | 0.29 | −0.263 | −0.193 | |
| RLD (>2 mm) | −0.084 | −0.05 | −0.124 | −0.272 | 0.536 * | 0.527 * | −0.207 | 0.131 | 0.25 | −0.2 | −0.242 | |
| RSAD (>2 mm) | −0.259 | −0.364 | −0.101 | −0.237 | 0.486 | 0.653 ** | −0.129 | 0.385 | 0.072 | −0.067 | −0.045 | |
| RVD (>2 mm) | −0.246 | −0.279 | −0.123 | −0.269 | 0.530 * | 0.642 ** | −0.156 | 0.335 | 0.154 | −0.14 | −0.101 | |
| 20–30 cm | RBD (<2 mm) | −0.273 | −0.075 | −0.113 | −0.234 | 0.222 | 0.127 | −0.019 | 0.123 | 0.509 | −0.455 | 0.205 |
| RLD (<2 mm) | −0.052 | −0.158 | 0.395 | 0.131 | 0.318 | 0.229 | −0.218 | 0.344 | 0.345 | −0.236 | −0.067 | |
| RSAD (<2 mm) | −0.146 | −0.197 | −0.136 | −0.034 | −0.09 | 0.107 | −0.188 | 0.145 | 0.156 | −0.145 | 0.077 | |
| RVD (<2 mm) | 0.08 | −0.351 | 0.194 | 0.338 | −0.306 | 0.262 | −0.408 | 0.442 | 0.035 | 0.113 | −0.391 | |
| RBD (>2 mm) | −0.244 | 0.067 | 0.004 | −0.184 | 0.439 | −0.368 | 0.441 | −0.394 | 0.495 | −0.577 * | 0.578 * | |
| RLD (>2 mm) | −0.307 | −0.078 | −0.073 | −0.204 | 0.267 | −0.058 | 0.135 | −0.046 | 0.484 | −0.479 | 0.327 | |
| RSAD (>2 mm) | −0.3 | −0.01 | −0.05 | −0.208 | 0.356 | −0.209 | 0.295 | −0.22 | 0.505 | −0.544 * | 0.464 | |
| RVD (>2 mm) | −0.23 | 0.092 | 0.022 | −0.151 | 0.426 | −0.373 | 0.454 | −0.411 | 0.482 | −0.571 * | 0.589 * | |
| Principal Component | PC-1 | PC-2 | PC-3 | PC-4 | PC-5 |
|---|---|---|---|---|---|
| Eigenvalue | 9.254 | 4.764 | 2.764 | 1.72 | 1.602 |
| Variance | 40.235 | 20.714 | 12.019 | 7.477 | 6.967 |
| Cumulative variance | 40.235 | 60.949 | 72.968 | 80.446 | 87.412 |
| Eigenvectors | |||||
| W | −0.22 | 0.168 | 0.511 | −0.42 | 0.047 |
| BD | −0.818 | −0.109 | 0.341 | −0.194 | 0.013 |
| TP | −0.042 | 0.843 | 0.093 | 0.371 | −0.13 |
| CP | −0.065 | 0.858 | 0.324 | 0.172 | 0.032 |
| NCP | 0.688 | −0.434 | −0.121 | 0.489 | −0.177 |
| SOC | 0.269 | 0.276 | 0.04 | 0.494 | 0.741 |
| pH | 0.422 | 0.675 | −0.101 | −0.134 | −0.363 |
| TN | 0.572 | 0.518 | −0.302 | 0.21 | 0.479 |
| Sand | −0.271 | −0.75 | 0.206 | 0.496 | −0.202 |
| Silt | 0.121 | 0.819 | −0.008 | −0.465 | 0.072 |
| Clay | 0.41 | 0.08 | −0.494 | −0.221 | 0.345 |
| RBD1 | 0.955 | 0.14 | 0.159 | −0.015 | −0.058 |
| RLD1 | 0.904 | 0.28 | 0.232 | 0.06 | −0.083 |
| RSAD1 | 0.898 | 0.07 | 0.301 | 0.189 | −0.039 |
| RVD1 | 0.656 | 0.241 | 0.37 | 0.076 | −0.461 |
| RBD2 | 0.759 | −0.455 | 0.332 | −0.2 | 0.151 |
| RLD2 | 0.783 | −0.376 | 0.449 | 0.014 | 0.11 |
| RSAD2 | 0.823 | −0.331 | 0.367 | −0.198 | 0.134 |
| RVD2 | 0.702 | −0.326 | 0.431 | −0.299 | 0.257 |
| IIR | 0.256 | −0.481 | −0.646 | −0.192 | 0.018 |
| SIR | 0.869 | 0.057 | −0.378 | −0.015 | −0.15 |
| AIR | 0.761 | −0.234 | −0.499 | −0.17 | −0.071 |
| CIA | 0.705 | 0.189 | −0.348 | −0.043 | −0.325 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Wan, C.; Qi, S.; Ma, S.; Zhang, L.; Cheng, G.; Fan, C.; Zheng, X.; Zhao, T. Soil Infiltration Characteristics and Driving Mechanisms of Three Typical Forest Types in Southern Subtropical China. Water 2025, 17, 1720. https://doi.org/10.3390/w17121720
Guo Y, Wan C, Qi S, Ma S, Zhang L, Cheng G, Fan C, Zheng X, Zhao T. Soil Infiltration Characteristics and Driving Mechanisms of Three Typical Forest Types in Southern Subtropical China. Water. 2025; 17(12):1720. https://doi.org/10.3390/w17121720
Chicago/Turabian StyleGuo, Yanrui, Chongshan Wan, Shi Qi, Shuangshuang Ma, Lin Zhang, Gong Cheng, Changjiang Fan, Xiangcheng Zheng, and Tianheng Zhao. 2025. "Soil Infiltration Characteristics and Driving Mechanisms of Three Typical Forest Types in Southern Subtropical China" Water 17, no. 12: 1720. https://doi.org/10.3390/w17121720
APA StyleGuo, Y., Wan, C., Qi, S., Ma, S., Zhang, L., Cheng, G., Fan, C., Zheng, X., & Zhao, T. (2025). Soil Infiltration Characteristics and Driving Mechanisms of Three Typical Forest Types in Southern Subtropical China. Water, 17(12), 1720. https://doi.org/10.3390/w17121720