
A Cold-Resistant Aerobic Denitrifying Bacterium Rhizobium pusense N7 for Aquaculture Wastewater Treatment and Its Denitrifying Characteristics

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Media
2.2. Isolation and Screening
2.3. Identification
2.4. Denitrification Characteristics
2.5. Application of Removing Nitrogen from Aquaculture Wastewater
2.6. Statistical Analysis
3. Results
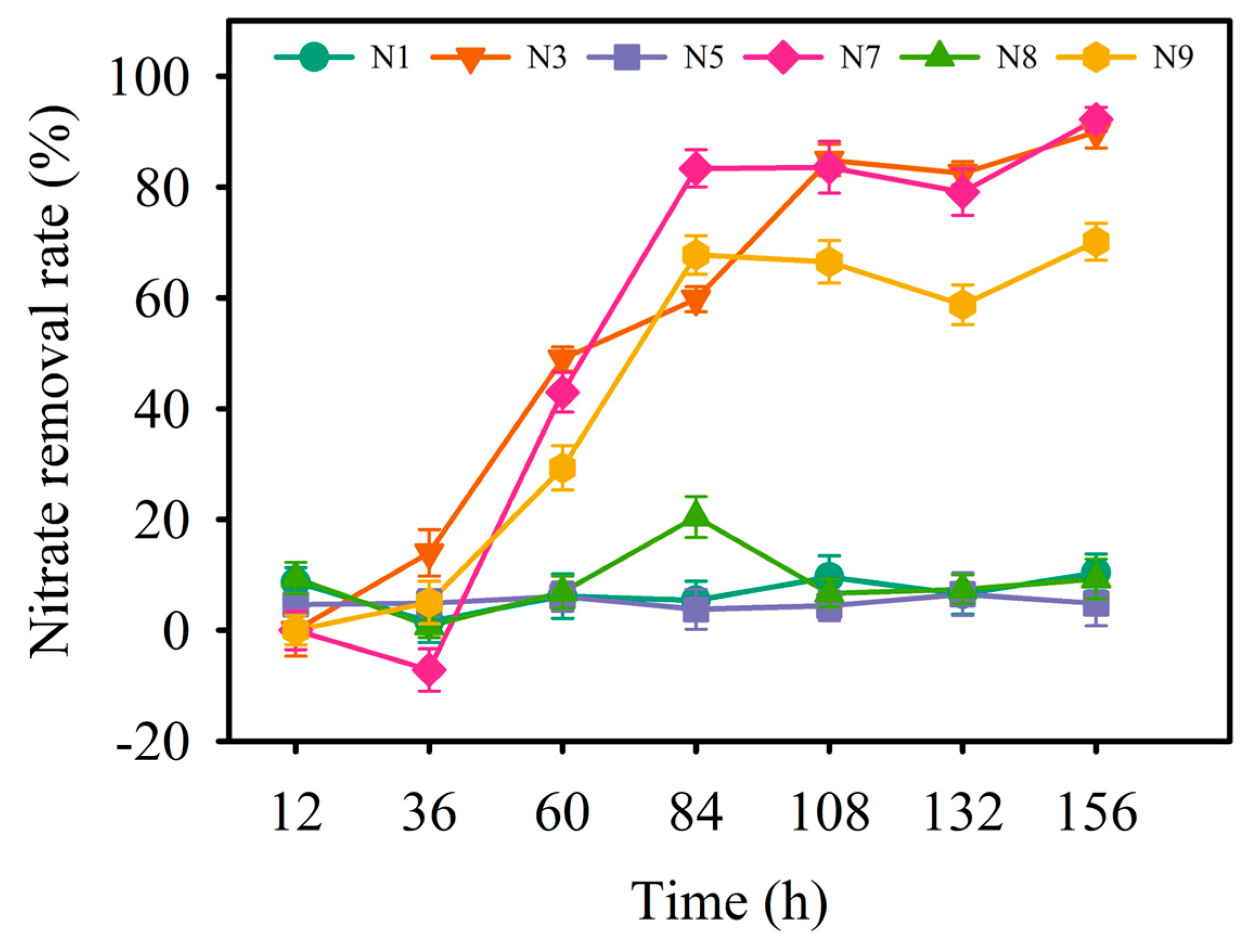
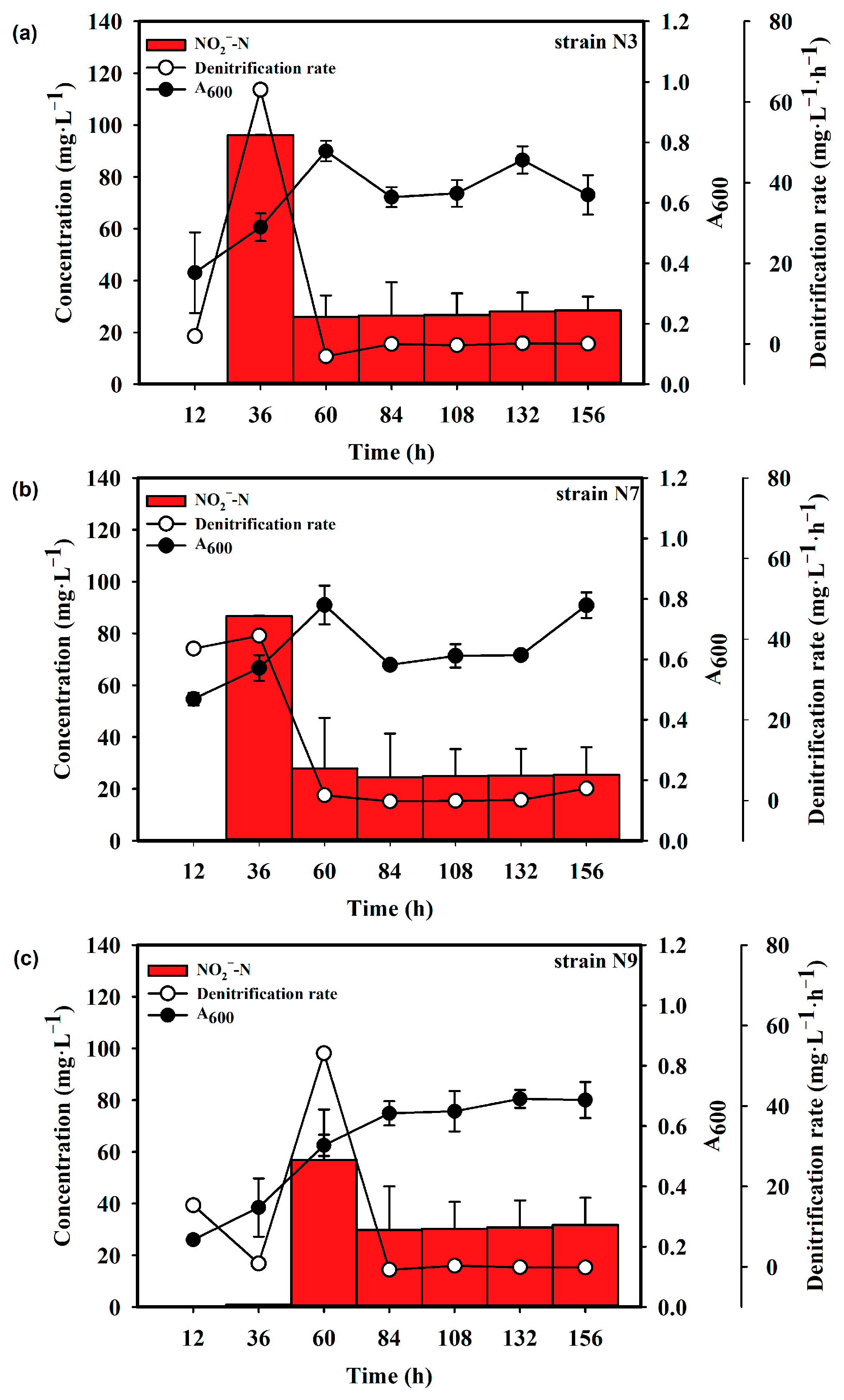
3.1. Isolation of Denitrifying Bacteria
3.2. Identification of Strain N7
3.3. Characteristics of Denitrification
3.3.1. Carbon Source
3.3.2. C/N Ratio
3.3.3. Temperature
3.3.4. Shaking Speed
3.3.5. pH

3.4. Effect of Removing Nitrogen from Aquaculture Wastewater
4. Discussion
4.1. Isolation of Bacteria
4.2. Effect of Environmental Conditions on Nitrate Removal
4.2.1. Carbon Source
4.2.2. C/N Ratio
4.2.3. Temperature
4.2.4. Shaking Speed
4.2.5. pH
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xue, S.; Xu, W.; Wei, J.; Sun, J. Impact of environmental bacterial communities on fish health in marine recirculating aquaculture systems. Vet. Microbiol. 2017, 203, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Jiang, R.; Qiu, J.; Liu, J.; Shao, L.; Zhang, J.; Liu, Q.; Jiang, Z.; Wang, H.; He, W.; et al. How to control pollution from tailwater in large scale aquaculture in China: A review. Aquaculture 2024, 590, 741085. [Google Scholar] [CrossRef]
- Quijano, G.; Arcila, J.S.; Buitrón, G. Microalgal-bacterial aggregates: Applications and perspectives for wastewater treatment. Biotechnol. Adv. 2017, 35, 772–781. [Google Scholar] [CrossRef] [PubMed]
- David, F.S.; Proença, D.C.; Valenti, W.C. Nitrogen budget in integrated aquaculture systems with Nile tilapia and Amazon River prawn. Aquacult. Int. 2017, 25, 1733–1746. [Google Scholar] [CrossRef]
- Vargas-Albores, F.; Martínez-Córdova, L.R.; Hernández-Mendoza, A.; Cicala, F.; Lago-Lestón, A.; Martínez-Porchas, M. Therapeutic modulation of fish gut microbiota, a feasible strategy for aquaculture? Aquaculture 2021, 544, 737050. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, Q.; Sun, H.; Jia, L.; Zhao, L.; Wu, W. Metagenomic analyses of microbial structure and metabolic pathway in solid-phase denitrification systems for advanced nitrogen removal of wastewater treatment plant effluent: A pilot-scale study. Water Res. 2021, 196, 117067. [Google Scholar] [CrossRef] [PubMed]
- Giatsis, C.; Sipkema, D.; Smidt, H.; Heilig, H.; Benvenuti, G.; Verreth, J.; Verdegem, M. The impact of rearing environment on the development of gut microbiota in tilapia larvae. Sci. Rep. 2015, 5, 18206. [Google Scholar] [CrossRef]
- Castignetti, D.; Hollocher, T.C. Heterotrophic nitrification among denitrifiers. Appl. Environ. Microbiol. 1984, 47, 620–623. [Google Scholar] [CrossRef]
- Wei, B.; Luo, X.; Ma, W.; Lv, P. Biological nitrogen removal and metabolic characteristics of a novel cold-resistant heterotrophic nitrification and aerobic denitrification Rhizobium sp. WS7. Bioresour. Technol. 2022, 362, 127756. [Google Scholar] [CrossRef]
- Lin, Z.; Zhou, J.; He, L.; He, X.; Pan, Z.; Wang, Y.; He, Q. High-temperature biofilm system based on heterotrophic nitrification and aerobic denitrification treating high-strength ammonia wastewater: Nitrogen removal performances and temperature-regulated metabolic pathways. Bioresour. Technol. 2022, 344, 126184. [Google Scholar] [CrossRef]
- Zhu, L.; Ding, W.; Feng, L.-j.; Dai, X.; Xu, X.-Y. Characteristics of an aerobic denitrifier that utilizes ammonium and nitrate simultaneously under the oligotrophic niche. Environ. Sci. Pollut. Res. 2012, 19, 3185–3191. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Chen, X.; Bai, J.; Li, B.; Li, H.; Huang, X. Bioaugmentation performance for moving bed biofilm reactor (MBBR) treating mariculture wastewater by an isolated novel halophilic heterotrophic nitrification aerobic denitrification (HNAD) strain (Zobellella B307). J. Environ. Manag. 2022, 325, 116566. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Zhou, M.; Chen, Y.; Hu, Y.; Luo, J. Insight into short-cut of simultaneous nitrification and denitrification process in moving bed biofilm reactor: Effects of carbon to nitrogen ratio. Chem. Eng. J. 2020, 400, 125905. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, C.; Rong, H.; Zheng, G.; Zhao, L. The effect of dissolved oxygen concentration (DO) on oxygen diffusion and bacterial community structure in moving bed sequencing batch reactor (MBSBR). Water Res. 2017, 108, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.Y.A.; Gill, L.; Monleon, A.; Pronk, M.; van Loosdrecht, M.; Saikaly, P.E.; Ali, M. Genome-resolved metatranscriptomics unveils distinct microbial functionalities across aggregate sizes in aerobic granular sludge. Environ. Sci. Ecotechnol. 2025, 25, 100560. [Google Scholar] [CrossRef]
- Tian, L.; Feng, K.; Qin, G.; Tong, X.; Feng, X.; Xu, X.; Zhu, L. Enhancing nitrogen removal of aerobic granular sludge with optimization of dissolved oxygen spatial distribution. J. Water Process Eng. 2022, 49, 103005. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, F.; Huang, Z.; Liu, M.; Mu, X.; Zhang, X.; Giglio, G.L.; Peng, Z. The impact of organic loading rate on the cultivation and stability of aerobic granular sludge in continuous flow reactor. Int. Biodeterior. Biodegrad. 2025, 198, 106016. [Google Scholar] [CrossRef]
- Shameem, M.R.; Sonali, J.M.I.; Kumar, P.S.; Rangasamy, G.; Gayathri, K.V.; Parthasarathy, V. Rhizobium mayense sp. Nov., an efficient plant growth-promoting nitrogen-fixing bacteria isolated from rhizosphere soil. Environ. Res. 2023, 220, 115200. [Google Scholar] [CrossRef]
- Safonov, A.V.; Babich, T.L.; Sokolova, D.S.; Grouzdev, D.S.; Tourova, T.P.; Poltaraus, A.B.; Zakharova, E.V.; Merkel, A.Y.; Novikov, A.P.; Nazina, T.N. Microbial community and in situ bioremediation of groundwater by nitrate removal in the zone of a radioactive waste surface repository. Front Microbiol. 2018, 9, 1985. [Google Scholar] [CrossRef]
- Hidalgo-Garcia, A.; Tortosa, G.; Pacheco, P.J.; Gates, A.J.; Richardson, D.J.; Bedmar, E.J.; Girard, L.; Torres, M.J.; Delgado, M.J. Rhizobium etli is able to emit nitrous oxide by connecting assimilatory nitrate reduction with nitrite respiration in the bacteroids of common bean nodules. J. Plant Interact. 2023, 18, 2251511. [Google Scholar] [CrossRef]
- Zou, Y.Y.; Zhang, Y.; Li, M.Z.; Mei, R.W.; Wei, Y.F.; Ding, L.X. Isolation and identification of a heterotropgic nitrification-aerobic denitrification bacterium and its denitrification ability. China Environ. Sci. 2016, 36, 887–893. Available online: https://kns.cnki.net/kcms2/article/abstract?v=fSCzX0TVvUjLExBLO09sX70OU-mEbMnTeMXXOwHGabPSFcPyQYKG-BlQtfiHWytFWuu3z06m5g82Tpr1kBF9keweI7Nv4zALoELMEoeidZupzGppSxYoH0YS39kSLrdi5QAmoT-1YxlnPuSbqExNfs6ouZ9WkMyI2Pdnox77GgwkcOv3BEBxIA==&uniplatform=NZKPT&language=CHS (accessed on 7 August 2015). (In Chinese).
- Hidalgo-Garcia, A.; Torres, M.J.; Sales, A.; Bedmar, E.J.; Girard, L.; Delgado, M.J. Rhizobium etli produces nitrous oxide by coupling the assimilatory and denitrification pathways. Front. Microbiol. 2019, 10, 980. [Google Scholar] [CrossRef] [PubMed]
- Vercellino, M.; Anahi Gomez, M. Denitrifying capacity of rhizobial strains of Argentine soils and herbicide sensitivity. Ann. Microbiol. 2013, 63, 1563–1570. [Google Scholar] [CrossRef]
- Lu, Y.; Kronzucker, H.J.; Shi, W. Stigmasterol root exudation arising from Pseudomonas inoculation of the duckweed rhizosphere enhances nitrogen removal from polluted waters. Environ. Pollut. 2021, 287, 117587. [Google Scholar] [CrossRef]
- Hu, N.; Li, Y.; Yin, J.; Ren, Z.; Li, J.; Zhao, J.; Wang, L.; Wu, L. A novel Zobellella endophytica W14 strain for nitrogen removal from hypersaline wastewater through simultaneous nitrification and denitrification. J. Environ. Manag. 2024, 371, 123171. [Google Scholar] [CrossRef]
- Chen, C.; Song, M.; Huang, G.; Li, R. Foam FeSO₄ modified limestone sulfur concrete for non-stink and high-rate nitrogen and phosphorus removal from wastewater. Water Res. 2025, 271, 122996. [Google Scholar] [CrossRef]
- Xu, M.J.; Cui, Y.W. Simultaneous aerobic nitrogen and phosphorus removal by novel halotolerant fungus Mucor circinelloides SNDM1: Function and metabolism pathway. Bioresour. Technol. 2024, 410, 131257. [Google Scholar] [CrossRef]
- Idenyi, J.N.; Abdallah, H.; Adeyemi, A.D.; Huber, D.H.; Gannam, A.; Sealey, W.; Igwe, D.O.; Eya, J.C. Optimizing growth and mitochondrial function in rainbow trout, Oncorhynchus mykiss through eco-friendly dietary and changes in water temperature regimen strategies. Aquaculture 2025, 595, 741591. [Google Scholar] [CrossRef]
- Korus, J.; Filgueira, R.; Grant, J. Influence of temperature on the behaviour and physiology of Atlantic salmon (Salmo salar) on a commercial farm. Aquaculture 2024, 589, 740978. [Google Scholar] [CrossRef]
- Kang, X.; Zhao, X.; Song, X.; Wang, D.; Shi, G.; Duan, X.; Chen, X.; Shen, G. Nitrogen removal by a novel strain Priestia aryabhattai KX-3 from East Antarctica under alkaline pH and low-temperature conditions. Process Biochem. 2023, 130, 674–684. [Google Scholar] [CrossRef]
- Lara-Moreno, A.; Vargas-Ordóñez, A.; Villaverde, J.; Madrid, F.; Carlier, J.D.; Santos, J.L.; Alonso, E.; Morillo, E. Bacterial bioaugmentation for paracetamol removal from water and sewage sludge. Genomic approaches to elucidate biodegradation pathway. J. Hazard. Mater. 2024, 480, 136128. [Google Scholar] [CrossRef]
- Zarei, M.; Yousefvand, A.; Maktabi, S.; Pourmahdi Borujeni, M.; Mohammadpour, H. Identification, phylogenetic characterisation and proteolytic activity quantification of high biofilm-forming Pseudomonas fluorescens group bacterial strains isolated from cold raw milk. Int. Dairy J. 2020, 109, 104787. [Google Scholar] [CrossRef]
- Tamminen, M.; Spaak, J.; Tlili, A.; Eggen, R.; Stamm, C.; Raesaenen, K. Wastewater constituents impact biofilm microbial community in receiving streams. Sci. Total Environ. 2022, 807, 151080. [Google Scholar] [CrossRef]
- Xu, W.; Xu, Y.; Su, H.; Hu, X.; Yang, K.; Wen, G.; Cao, Y. Characteristics of ammonia removal and nitrifying microbial communities in a hybrid biofloc-RAS for intensive Litopenaeus vannamei culture: A Pilot-Scale Study. Water 2020, 12, 3000. [Google Scholar] [CrossRef]
- Tang, B.; Yu, C.; Bin, L.; Zhao, Y.; Feng, X.; Huang, S.; Fu, F.; Ding, J.; Chen, C.; Li, P.; et al. Essential factors of an integrated moving bed biofilm reactor–membrane bioreactor: Adhesion characteristics and microbial community of the biofilm. Bioresour. Technol. 2016, 211, 574–583. [Google Scholar] [CrossRef]
- Finan, T.M.; Wood, J.M.; Jordan, D.C. Succinate transport in Rhizobium leguminosarum. J. Bacteriol. 1981, 148, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Liu, Y.; Sun, G.; Gao, X.; Zhang, Q.; Liu, Z. Denitrification characteristics of a marine origin psychrophilic aerobic denitrifying bacterium. J. Environ. Sci. 2011, 23, 1888–1893. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.Y.; Liu, Y.; Gao, X.Y.; Ai, G.M.; Miao, L.L.; Liu, Z.P. Characterization of a marine origin aerobic nitrifying–denitrifying bacterium. J Biosci. Bioeng. 2012, 114, 33–37. [Google Scholar] [CrossRef]
- Shao, S.; Zhong, J.; Wang, C.; Pan, D.; Wu, X. Performance of simultaneous nitrification–denitrification and denitrifying phosphorus and manganese removal by driving a single-stage moving bed biofilm reactor based on manganese redox cycling. Bioresour. Technol. 2022, 362, 127846. [Google Scholar] [CrossRef]
- Lei, Y.; Wang, Y.; Liu, H.; Xi, C.; Song, L. A novel heterotrophic nitrifying and aerobic denitrifying bacterium, Zobellella taiwanensis DN-7, can remove high-strength ammonium. Appl. Microbiol. Biotechnol. 2016, 100, 4219–4229. [Google Scholar] [CrossRef]
- Shu, H.; Sun, H.; Huang, W.; Zhao, Y.; Ma, Y.; Chen, W.; Sun, Y.; Chen, X.; Zhong, P.; Yang, H.; et al. Nitrogen removal characteristics and potential application of the heterotrophic nitrifying-aerobic denitrifying bacteria Pseudomonas mendocina S16 and Enterobacter cloacae DS’5 isolated from aquaculture wastewater ponds. Bioresour. Technol. 2022, 345, 126541. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Lu, J.; Qin, Y.; Zhou, M.; Tan, Y.; Wu, P.; Zhao, J. A critical review of heterotrophic nitrification and aerobic denitrification process: Influencing factors and mechanisms. J. Water Process Eng. 2023, 54, 103995. [Google Scholar] [CrossRef]
- Hu, J.; Yang, X.; Deng, X.; Liu, X.; Yu, J.; Chi, R.; Xiao, C. Isolation and nitrogen removal efficiency of the heterotrophic nitrifying-aerobic denitrifying strain K17 from a rare earth element leaching site. Front. Microbiol. 2022, 13, 905409. [Google Scholar] [CrossRef]
- Ren, Y.X.; Yang, L.; Liang, X. The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium, Acinetobacter junii YB. Bioresour. Technol. 2014, 171, 1–9. [Google Scholar] [CrossRef]
- Sun, L.; Ge, Q.L.; Cao, W.P.; Zhang, H.F.; Zhang, J.K.; Zhang, H.; Dong, Y.W.; Li, Q.N.; Wang, Y.H. Screening and characterization study on an aerobic denitrifying bacteria. J. Henan Polytech. Univ. (Nat. Sci.) 2017, 36, 79–85. (In Chinese) [Google Scholar] [CrossRef]
- Volokita, M.; Belkin, S.; Abeliovich, A.; Soares, M.I.M. Biological denitrification of drinking water using newspaper. Water Res. 1996, 30, 965–971. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Liu, M.; Liu, C.; Yin, Z. Simultaneous nitrogen and phosphorus removal from black water: Effects of dissolved oxygen level and sludge concentration on full-scale performance and bacterial community dynamics. J. Water Process Eng. 2024, 57, 104735. [Google Scholar] [CrossRef]
- Davies, K.J.P.; Lloyd, D.; Boddy, L. The effect of oxygen on denitrification in Paracoccus denitrificans and Pseudomonas aeruginosa. Microbiology 1989, 135, 2445–2451. [Google Scholar] [CrossRef] [PubMed]
- Li, W.F.; Du, L.B.; Liu, S.Y.; Weng, M.S.; Shu, H.; Chen, Q.H. Isolation and identification of an efficient aerobic denitrifying bacterium. Biotechnol. Bull. 2019, 35, 202–209. Available online: https://biotech.aiijournal.com/EN/10.13560/j.cnki.biotech.bull.1985.2019-0339 (accessed on 19 April 2019). (In Chinese).
- Zhang, Y.H.; Dong, X.B.; Liu, X.Y.; Xu, J.Q.; Xu, Z.L. Isolation of a novel heterotrophic nitrification-aerobic denitrification bacterium Paracoccus sp. QD-19 and its characterization of removing nitrogen. Biotechnol. Bull. 2023, 39, 301–310. Available online: https://biotech.aiijournal.com/EN/abstract/abstract13790.shtml (accessed on 6 July 2022). (In Chinese).
- AbdelGawwad, M.R.; Mahmutović, E.; Al Farraj, D.A.; Elshikh, M.S. In silico prediction of silver nitrate nanoparticles and nitrate reductase A (NAR A) interaction in the treatment of infectious disease causing clinical strains of E. coli. J. Infect. Public Health 2020, 13, 1580–1585. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Qiao, S.; Zhou, J.; Bhatti, Z. Effects of redox mediators on nitrogen removal performance by denitrifying biomass and the activity of Nar and Nir. Chem. Eng. J. 2014, 257, 90–97. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Huang, H.K.; Tseng, S.K. Nitrate reduction by Citrobacter diversus under aerobic environment. Appl. Microbiol. Biotechnol. 2001, 55, 90–94. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Size/cm | Morphology | Color | Surface | Bulge | Transparency | Texture |
|---|---|---|---|---|---|---|---|
| N1 | 3–5 | circle | white | rough | bulge | non-transparent | dry |
| N2 | 3–28 | circle | black | rough | bulge | non-transparent | dry |
| N3 | 1–3 | circle | none | smooth | bulge | transparent | wet |
| N4 | 5 | water-drop | white | smooth | flat | translucent | wet |
| N5 | 2–3 | circle | yellow | smooth | bulge | non-transparent | dry |
| N6 | 27 | circle | green | rough | bulge | non-transparent | dry |
| N7 | 1–3 | circle | white | smooth | bulge | non-transparent | wet |
| N8 | 0.5 | circle | yellow | rough | bulge | non-transparent | dry |
| N9 | 1–3 | circle | none | smooth | bulge | transparent | wet |
| Test | Results | Test | Results |
|---|---|---|---|
| Glucose | - | Citrate | - |
| Lactose | - | Ornithine | + |
| Mannitol | + | Arginine dehydrogenase | - |
| Hydrogen sulfide | - | lysine | + |
| Urease | + | Salicin | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, S.; Song, M.; Kan, Z.; Fu, Y.; Liu, Y.; Zhang, Y.; Zhang, R. A Cold-Resistant Aerobic Denitrifying Bacterium Rhizobium pusense N7 for Aquaculture Wastewater Treatment and Its Denitrifying Characteristics. Water 2025, 17, 1592. https://doi.org/10.3390/w17111592
Lyu S, Song M, Kan Z, Fu Y, Liu Y, Zhang Y, Zhang R. A Cold-Resistant Aerobic Denitrifying Bacterium Rhizobium pusense N7 for Aquaculture Wastewater Treatment and Its Denitrifying Characteristics. Water. 2025; 17(11):1592. https://doi.org/10.3390/w17111592
Chicago/Turabian StyleLyu, Siwei, Meiying Song, Zhiying Kan, Yuhao Fu, Yang Liu, Yixiang Zhang, and Rongfei Zhang. 2025. "A Cold-Resistant Aerobic Denitrifying Bacterium Rhizobium pusense N7 for Aquaculture Wastewater Treatment and Its Denitrifying Characteristics" Water 17, no. 11: 1592. https://doi.org/10.3390/w17111592
APA StyleLyu, S., Song, M., Kan, Z., Fu, Y., Liu, Y., Zhang, Y., & Zhang, R. (2025). A Cold-Resistant Aerobic Denitrifying Bacterium Rhizobium pusense N7 for Aquaculture Wastewater Treatment and Its Denitrifying Characteristics. Water, 17(11), 1592. https://doi.org/10.3390/w17111592