
Accumulation of Nitrogen Species from Industrial Wastewater by Vetiver Grass (Chrysopogon zizanioides)


 ,
,  ,
, 
Abstract
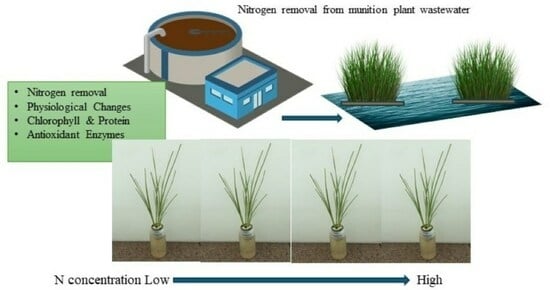
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Chlorophyll Content
2.3. Protein Extraction and Quantification
2.4. Antioxidant Enzyme Activity
2.5. Data Analysis
3. Results and Discussion
3.1. Removal of Nitrogen Species from Simulated Wastewater
3.1.1. Nitrate
3.1.2. Nitrite
3.1.3. Ammonium
3.2. Plant Growth
3.3. Chlorophyll
3.4. Analysis of Protein
3.5. Antioxidant Enzyme Response Due to Nitrogen Species Stress
3.5.1. Superoxide Dismutase
3.5.2. Catalase
3.5.3. Guaiacol Peroxidase
3.5.4. Glutathione S-Transferase
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IHEs | insensitive high explosives |
| ECs | energetic compounds |
| Chl | chlorophyll |
| ROS | reactive oxygen species |
| CAT | catalase |
| GST | glutathione S-transferase |
| SOD | superoxide dismutase |
| GPX | glutathione peroxidase |
References
- Panja, S.; Sarkar, D.; Datta, R. Vetiver grass (Chrysopogon zizanioides) is capable of removing insensitive high explosives from munition industry wastewater. Chemosphere 2018, 209, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Zoh, K.-D.; Stenstrom, M.K. Fenton oxidation of hexahydro-1,3,5-trinitro-1,3,5-triazine (RDX) and octahydro-1,3,5,7-tetranitro-1,3,5,7-tetrazocine (HMX). Water Res. 2002, 36, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Resende, J.D.; Nolasco, M.A.; Pacca, S.A. Life cycle assessment and costing of wastewater treatment systems coupled to constructed wetlands. Resour. Conserv. Recycl. 2019, 148, 170–177. [Google Scholar] [CrossRef]
- Karimi, H.; Mohammadi, F.; Rajabi, S.; Mahvi, A.H.; Ghanizadeh, G. Biological 2,4,6-trinitrotoluene removal by extended aeration activated sludge: Optimization using artificial neural network. Sci. Rep. 2023, 13, 9053. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chen, N.; Feng, Z.; Feng, C.; Deng, Y. Treatment of nitrate containing wastewater by adsorption process using polypyrrole-modified plastic-carbon: Characteristic and mechanism. Chemosphere 2022, 297, 134107. [Google Scholar] [CrossRef]
- Yan, A.; Wang, Y.; Tan, S.N.; Mohd Yusof, M.L.; Ghosh, S.; Chen, Z. Phytoremediation: A promising approach for revegetation of heavy metal-polluted land. Front. Plant Sci. 2020, 11, 359. [Google Scholar] [CrossRef]
- Andra, S.S.; Sarkar, D.; Saminathan, S.K.M.; Datta, R.; Andra, S.S.; Saminathan, S.K.M.; Sarkar, D.; Datta, R. Exchangeable lead from prediction models relates to vetiver lead uptake in different soil types. Environ. Monit. Assess. 2011, 183, 571–579. [Google Scholar] [CrossRef]
- Ketaubon, P.; Ritthikasem, N.; Tanheng, P.; Prapagdee, B. Enhancing heavy metal phytoremediation in landfill soil by Chrysopogon zizanioides (L.) roberty through the application of bacterial-biochar pellets. Environ. Technol. Innov. 2024, 35, 103738. [Google Scholar] [CrossRef]
- RoyChowdhury, A.; Datta, R.; Sarkar, D. Heavy Metal Pollution and Remediation. Green Chem. Incl. Approach 2018, 359–373. [Google Scholar] [CrossRef]
- Corredor, D.; Duchicela, J.; Flores, F.J.; Maya, M.; Guerron, E. Review of Explosive Contamination and Bioremediation: Insights from Microbial and Bio-Omic Approaches. Toxics 2024, 12, 249. [Google Scholar] [CrossRef]
- Worku, A.; Tefera, N.; Kloos, H.; Benor, S. Bioremediation of brewery wastewater using hydroponics planted with vetiver grass in Addis Ababa, Ethiopia. Bioresour. Bioprocess. 2018, 5, 1–12. [Google Scholar] [CrossRef]
- Akbarzadeh, A.; Jamshidi, S.; Vakhshouri, M. Nutrient uptake rate and removal efficiency of Vetiveria zizanioides in contaminated waters. Pollution 2015, 1, 1–8. [Google Scholar]
- Goren, A.Y.; Yucel, A.; Sofuoglu, S.C.; Sofuoglu, A. Phytoremediation of olive mill wastewater with Vetiveria zizanioides (L.) Nash and Cyperus alternifolius L. Environ. Technol. Innov. 2021, 24, 102071. [Google Scholar] [CrossRef]
- Junio, C.J.; Ontar, P.A.; Rutaquio, V.J.; Manlapaz, J.M.; Braga, E.B.; Tugade, C.; Pescos, C.J. Horizontal-flow constructed wetlands by phytoremediation using vetiver grass, common reed, and canna lily as tertiary wastewater treatment for the reduction of pollutant concentrations of ammonia, phosphates, and nitrates. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2024; Volume 1372, p. 12045. [Google Scholar]
- Kangwannarakul, N.; Wantawin, C.; Noophan, P. Anammox bacteria with attached-growth media for nitrogen removal in wastewater. Clean Technol. Environ. Policy 2018, 20, 219–226. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Y.; Liu, C.; Wang, Z.; Dong, J.; Zhu, G.; Huang, X. Potential effect and accumulation of veterinary antibiotics in Phragmites australis under hydroponic conditions. Ecol. Eng. 2013, 53, 138–143. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Q.; Jinadasa, K.; Gersberg, R.M.; Liu, Y.; Ng, W.J.; Tan, S.K. Application of constructed wetlands for wastewater treatment in developing countries—A review of recent developments (2000–2013). J. Environ. Manag. 2014, 141, 116–131. [Google Scholar] [CrossRef]
- Effendi, H.; Utomo, B.A.; Pratiwi, N.T.M. Ammonia and orthophosphate removal of tilapia cultivation wastewater with Vetiveria zizanioides. J. King Saud Univ. 2020, 32, 207–212. [Google Scholar] [CrossRef]
- Norton, S. Removal Mechanisms in Constructed Wastewater Wetlands. 2014. Available online: https://home.engineering.iastate.edu/tge/ce421-521/stephen.pdf (accessed on 5 May 2025).
- Gholipour, M.; Mehrabanjoubani, P.; Abdolzadeh, A.; Raghimi, M.; Seyedkhademi, S.; Karimi, E.; Sadeghipour, H.R. Facilitated decrease of anions and cations in influent and effluent of sewage treatment plant by vetiver grass (Chrysopogon zizanioides): The uptake of nitrate, nitrite, ammonium, and phosphate. Environ. Sci. Pollut. Res. 2020, 27, 21506–21516. [Google Scholar] [CrossRef]
- Boonsong, K.; Chansiri, M. Domestic wastewater treatment using vetiver grass cultivated with floating platform technique. AU J. Technol. 2008, 12, 73–80. [Google Scholar]
- Panja, S.; Sarkar, D.; Li, K.; Datta, R. Uptake and transformation of ciprofloxacin by vetiver grass (Chrysopogon zizanioides). Int. Biodeterior. Biodegrad. 2019, 142, 200–210. [Google Scholar] [CrossRef]
- Panja, S.; Sarkar, D.; Zhang, Z.; Datta, R. Removal of antibiotics and nutrients by vetiver grass (Chrysopogon zizanioides) from a plug flow reactor based constructed wetland model. Toxics 2021, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- RoyChowdhury, A.; Mukherjee, P.; Panja, S.; Datta, R.; Christodoulatos, C.; Sarkar, D. Evidence for Phytoremediation and Phytoexcretion of NTO from Industrial Wastewater by Vetiver Grass. Molecules 2020, 26, 74. [Google Scholar] [CrossRef]
- Ahmed, M.; Rauf, M.; Akhtar, M.; Mukhtar, Z.; Saeed, N.A. Hazards of nitrogen fertilizers and ways to reduce nitrate accumulation in crop plants. Environ. Sci. Pollut. Res. 2020, 27, 17661–17670. [Google Scholar] [CrossRef]
- Hoque, M.A.; Okuma, E.; Banu, M.N.A.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Exogenous proline mitigates the detrimental effects of salt stress more than exogenous betaine by increasing antioxidant enzyme activities. J. Plant Physiol. 2007, 164, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Shilpha, J.; Song, J.; Jeong, B.R. Ammonium phytotoxicity and tolerance: An insight into ammonium nutrition to improve crop productivity. Agronomy 2023, 13, 1487. [Google Scholar] [CrossRef]
- Singh, P.K.; Tewari, R.K. Cadmium toxicity induced changes in plant water relations and oxidative metabolism of Brassica juncea L. plants. J. Environ. Biol. 2003, 24, 107–112. [Google Scholar] [PubMed]
- Wang, R.; Guegler, K.; LaBrie, S.T.; Crawford, N.M. Genomic analysis of a nutrient response in Arabidopsis reveals diverse expression patterns and novel metabolic and potential regulatory genes induced by nitrate. Plant Cell 2000, 12, 1491–1509. [Google Scholar] [CrossRef]
- Stitt, M. Nitrate regulation of metabolism and growth. Curr. Opin. Plant Biol. 1999, 2, 178–186. [Google Scholar] [CrossRef]
- Krapp, A.; David, L.C.; Chardin, C.; Girin, T.; Marmagne, A.; Leprince, A.-S.; Chaillou, S.; Ferrario-Méry, S.; Meyer, C.; Daniel-Vedele, F. Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 789–798. [Google Scholar] [CrossRef]
- Ng, Y.S.; Chan, D.J.C. Wastewater phytoremediation by Salvinia molesta. J. Water Process Eng. 2017, 15, 107–115. [Google Scholar] [CrossRef]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Navabpour, S.; Yamchi, A.; Bagherikia, S.; Kafi, H. Lead-induced oxidative stress and role of antioxidant defense in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2020, 26, 793–802. [Google Scholar] [CrossRef]
- Stephenie, S.; Chang, Y.P.; Gnanasekaran, A.; Esa, N.M.; Gnanaraj, C. An insight on superoxide dismutase (SOD) from plants for mammalian health enhancement. J. Funct. Foods 2020, 68, 103917. [Google Scholar] [CrossRef]
- Medici, L.O.; Azevedo, R.A.; Smith, R.J.; Lea, P.J. The influence of nitrogen supply on antioxidant enzymes in plant roots. Funct. Plant Biol. 2004, 31, 1–9. [Google Scholar] [CrossRef]
- Alfonso-Prieto, M.; Biarnés, X.; Vidossich, P.; Rovira, C. The molecular mechanism of the catalase reaction. J. Am. Chem. Soc. 2009, 131, 11751–11761. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.; Gomes, D.; Lima, M.; Bronze-da-Rocha, E.; Santos-Silva, A. Peroxiredoxin 2, glutathione peroxidase, and catalase in the cytosol and membrane of erythrocytes under H2O2-induced oxidative stress. Free Radic. Res. 2015, 49, 990–1003. [Google Scholar] [CrossRef]
- Kumar, S.; Trivedi, P.K. Glutathione S-transferases: Role in combating abiotic stresses including arsenic detoxification in plants. Front. Plant Sci. 2018, 9, 751. [Google Scholar] [CrossRef]
- Lin, D.; Huang, Y.; Zhao, J.; Wu, Z.; Liu, S.; Qin, W.; Wu, D.; Chen, H.; Zhang, Q. Evaluation of seed nitrate assimilation and stimulation of phenolic-linked antioxidant on pentose phosphate pathway and nitrate reduction in three feed-plant species. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Nimptsch, J.; Pflugmacher, S. Ammonia triggers the promotion of oxidative stress in the aquatic macrophyte Myriophyllum mattogrossense. Chemosphere 2007, 66, 708–714. [Google Scholar] [CrossRef]
- Skopelitis, D.S.; Paranychianakis, N.V.; Paschalidis, K.A.; Pliakonis, E.D.; Delis, I.D.; Yakoumakis, D.I.; Kouvarakis, A.; Papadakis, A.K.; Stephanou, E.G.; Roubelakis-Angelakis, K.A. Abiotic stress generates ROS that signal expression of anionic glutamate dehydrogenases to form glutamate for proline synthesis in tobacco and grapevine. Plant Cell 2006, 18, 2767–2781. [Google Scholar] [CrossRef] [PubMed]
- Motamedi-Tehrani, J.; Peyghan, R.; Shahriari, A.; Razijalali, M.; Ebrahimi, E. The influence of ammonia-N and salinity levels on oxidative stress markers, hepatic enzymes, and acid phosphatase activity in Nile tilapia (Oreochromis niloticus). Sci. Rep. 2025, 15, 559. [Google Scholar] [CrossRef]
- Polesskaya, O.G.; Kashirina, E.I.; Alekhina, N.D. Changes in the activity of antioxidant enzymes in wheat leaves and roots as a function of nitrogen source and supply. Russ. J. Plant Physiol. 2004, 51, 615–620. [Google Scholar] [CrossRef]
- Abraham, J.; Lin, Y.; RoyChowdhury, A.; Christodoulatos, C.; Conway, M.; Smolinski, B.; Braida, W. Algae toxicological assessment and valorization of energetic-laden wastewater streams using Scenedesmus obliquus. J. Clean. Prod. 2018, 202, 838–845. [Google Scholar] [CrossRef]
- Brenner, A.; Ronen, Z.; Harel, Y.; Abeliovich, A. Use of Hexahydro-1,3,5-trinitro-1,3,5-triazine as a Nitrogen Source in Biological Treatment of Munitions Wastes. Water Environ. Res. 2000, 72, 469–475. [Google Scholar] [CrossRef]
- Huibregtse, K.R.; Fulk, R.; Zanoni, A.E.; Zenker, D. Feasibility Study Regarding Landfill of Nitrocellulose Lime Sludge and Oxidation of Nitroglycerin Wastewater Stream; Envirex, Inc.: Milwaukee, WI, USA, 1978. [Google Scholar]
- Oh, S.-Y.; Cha, D.K.; Chiu, P.-C.; Kim, B.J. Zero-valent iron treatment of RDX-containing and perchlorate-containing wastewaters from an ammunition-manufacturing plant at elevated temperatures. Water Sci. Technol. 2006, 54, 47–53. [Google Scholar] [CrossRef]
- Ronen, Z.; Brenner, A.; Abeliovich, A. Biodegradation of RDX-contaminated wastes in a nitrogen-deficient environment. Water Sci. Technol. 1998, 38, 219–224. [Google Scholar] [CrossRef]
- Terracciano, A.; Christodoulatos, C.; Koutsospyros, A.; Zheng, Z.; Su, T.-L.; Smolinski, B.; Arienti, P.; Meng, X. Degradation of 3-nitro-1,2,4-trizole-5-one (NTO) in wastewater with UV/H2O2 oxidation. Chem. Eng. J. 2018, 354, 81–491. [Google Scholar] [CrossRef]
- Terracciano, A.; Koutsospyros, A.; Christodoulatos, C.; Mai, A.; Meng, X.; Su, T.-L.; Smolinski, B. Oxidative degradation of nitroguanidine (NQ) by UV-C and oxidants: Hydrogen peroxide, persulfate and peroxymonosulfate. Chemosphere 2022, 292, 133357. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Removal Efficiency (%) | Effective Concentration Range | Chlorophyll Decline at High Conc. | Protein Trends | CAT Enzyme Activity (Root) | |
|---|---|---|---|---|---|
| Nitrate | 73% | L1–L4 | 70–95% decrease at L5–L8 | ↑ shoots, ↓ roots (L5–L8) | ↑ 22–77% |
| Nitrite | 75% | L1–L3 | Up to 75% decrease at L1 | ↑ shoots, ↓ roots | ↑ 40–155% |
| Ammonium | 83.60% | L1–L3 | >90% decrease from L4 onward | ↑ shoots (L4–L8), ↓ roots | ↑ 8–33% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aliasghar, A.; Zhang, Z.; Datta, R.; Christodoulatos, C.; Sarkar, D. Accumulation of Nitrogen Species from Industrial Wastewater by Vetiver Grass (Chrysopogon zizanioides). Water 2025, 17, 1464. https://doi.org/10.3390/w17101464
Aliasghar A, Zhang Z, Datta R, Christodoulatos C, Sarkar D. Accumulation of Nitrogen Species from Industrial Wastewater by Vetiver Grass (Chrysopogon zizanioides). Water. 2025; 17(10):1464. https://doi.org/10.3390/w17101464
Chicago/Turabian StyleAliasghar, Arash, Zhiming Zhang, Rupali Datta, Christos Christodoulatos, and Dibyendu Sarkar. 2025. "Accumulation of Nitrogen Species from Industrial Wastewater by Vetiver Grass (Chrysopogon zizanioides)" Water 17, no. 10: 1464. https://doi.org/10.3390/w17101464
APA StyleAliasghar, A., Zhang, Z., Datta, R., Christodoulatos, C., & Sarkar, D. (2025). Accumulation of Nitrogen Species from Industrial Wastewater by Vetiver Grass (Chrysopogon zizanioides). Water, 17(10), 1464. https://doi.org/10.3390/w17101464