
Functional Traits Drive the Changes in Diversity and Composition of Benthic Invertebrate Communities in Response to Hydrological Regulation


Abstract
1. Introduction
2. Materials and Methods
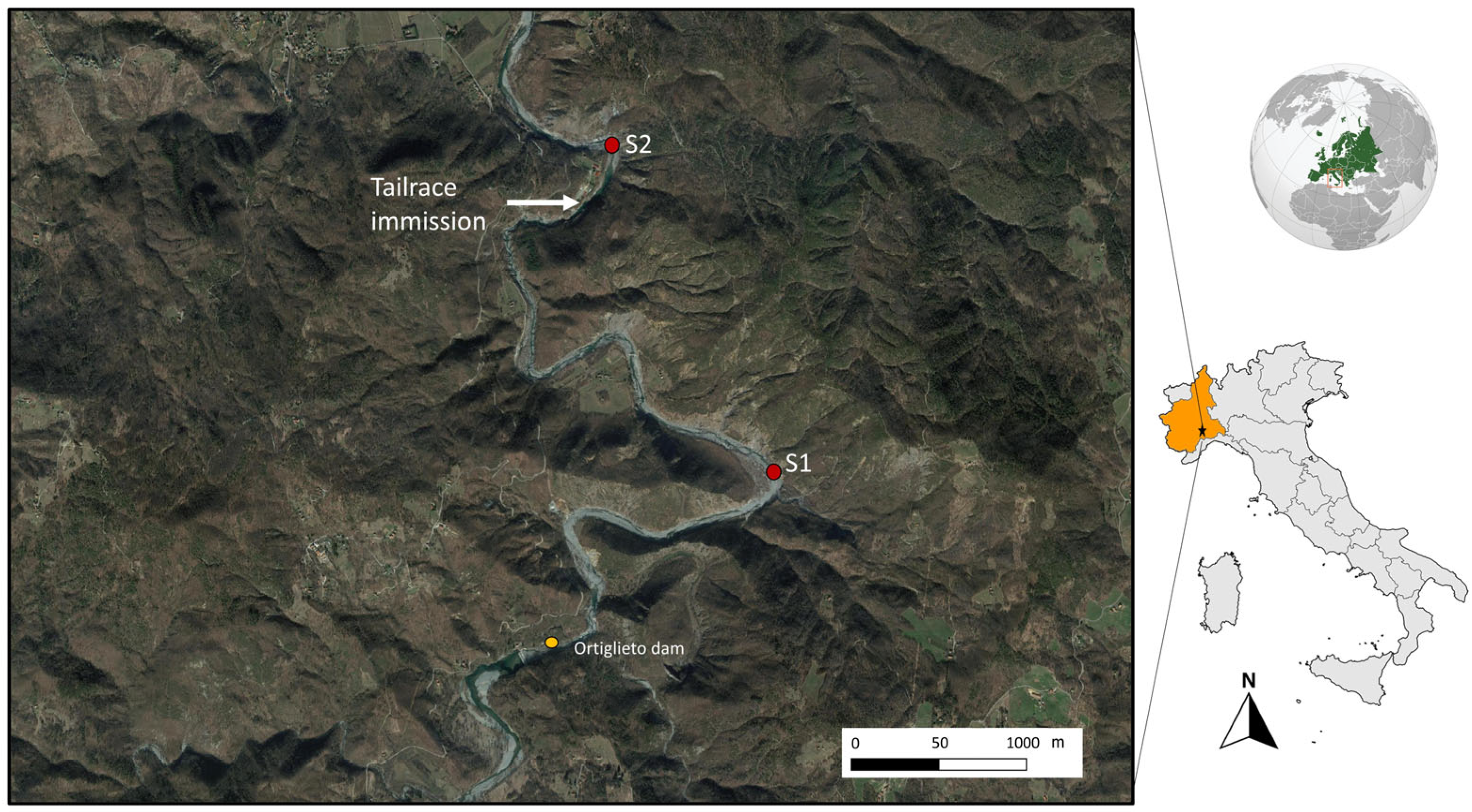
2.1. Area of Study
2.2. Macroinvertebrate Sampling and Processing
2.3. Statistical Analyses
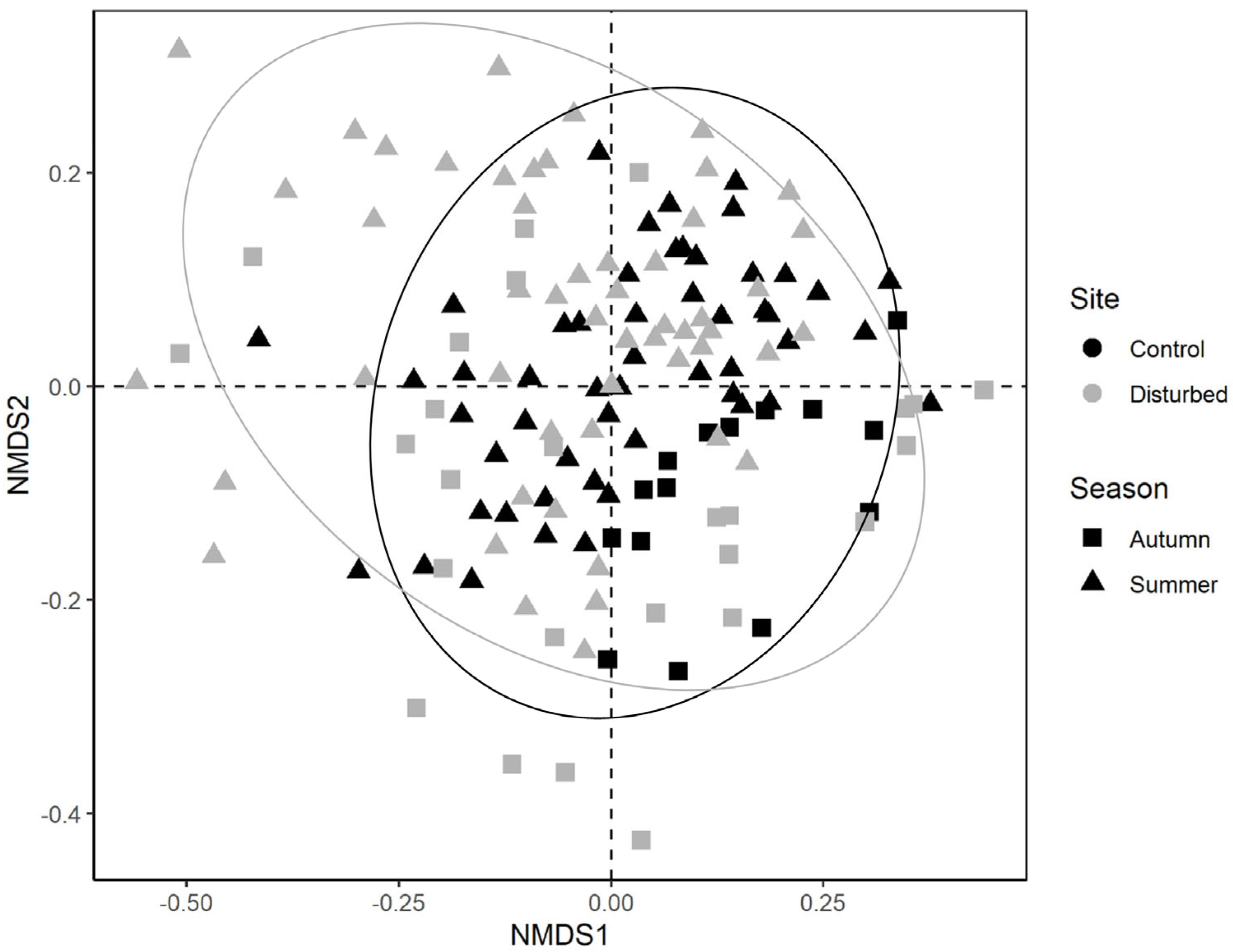
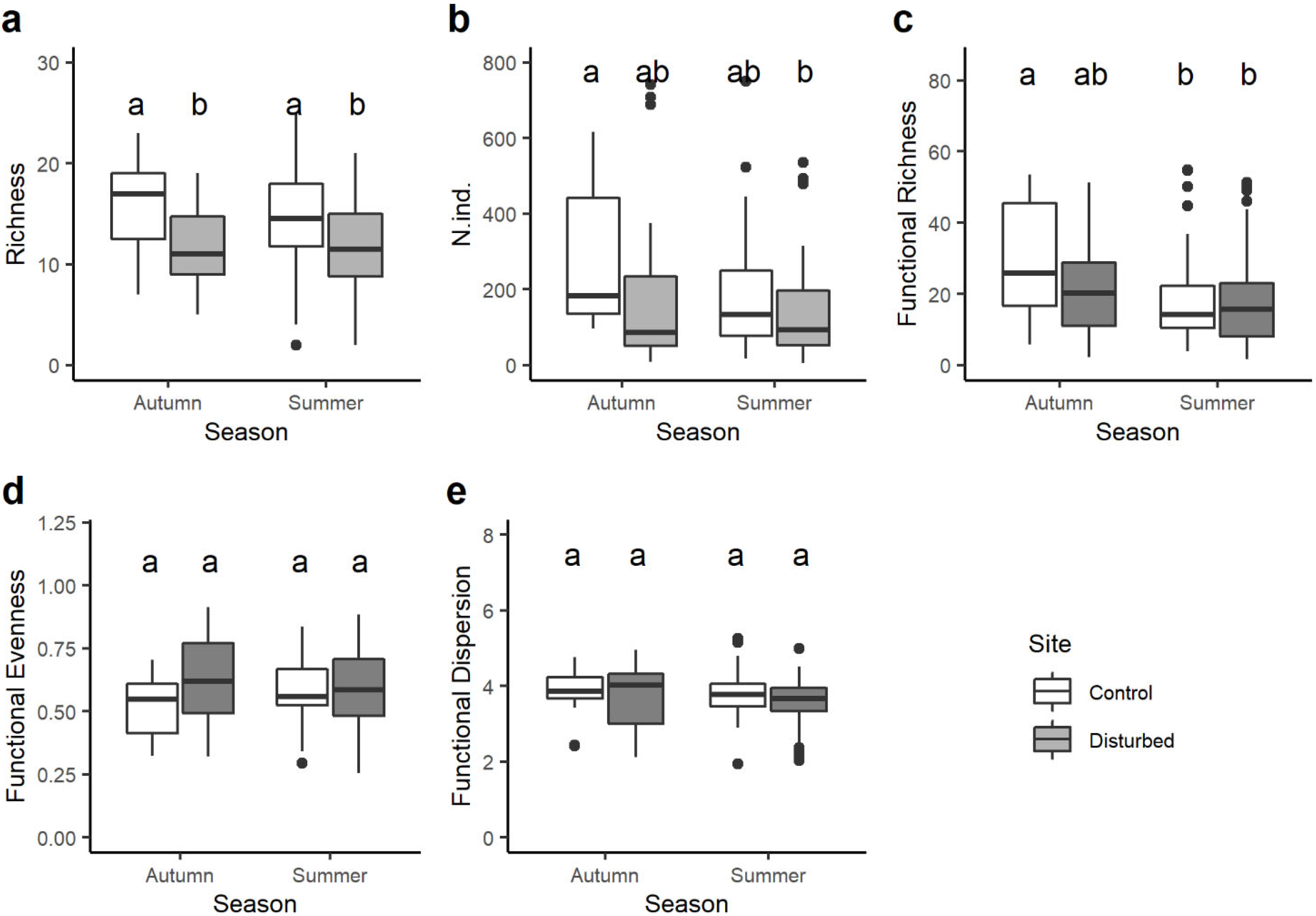
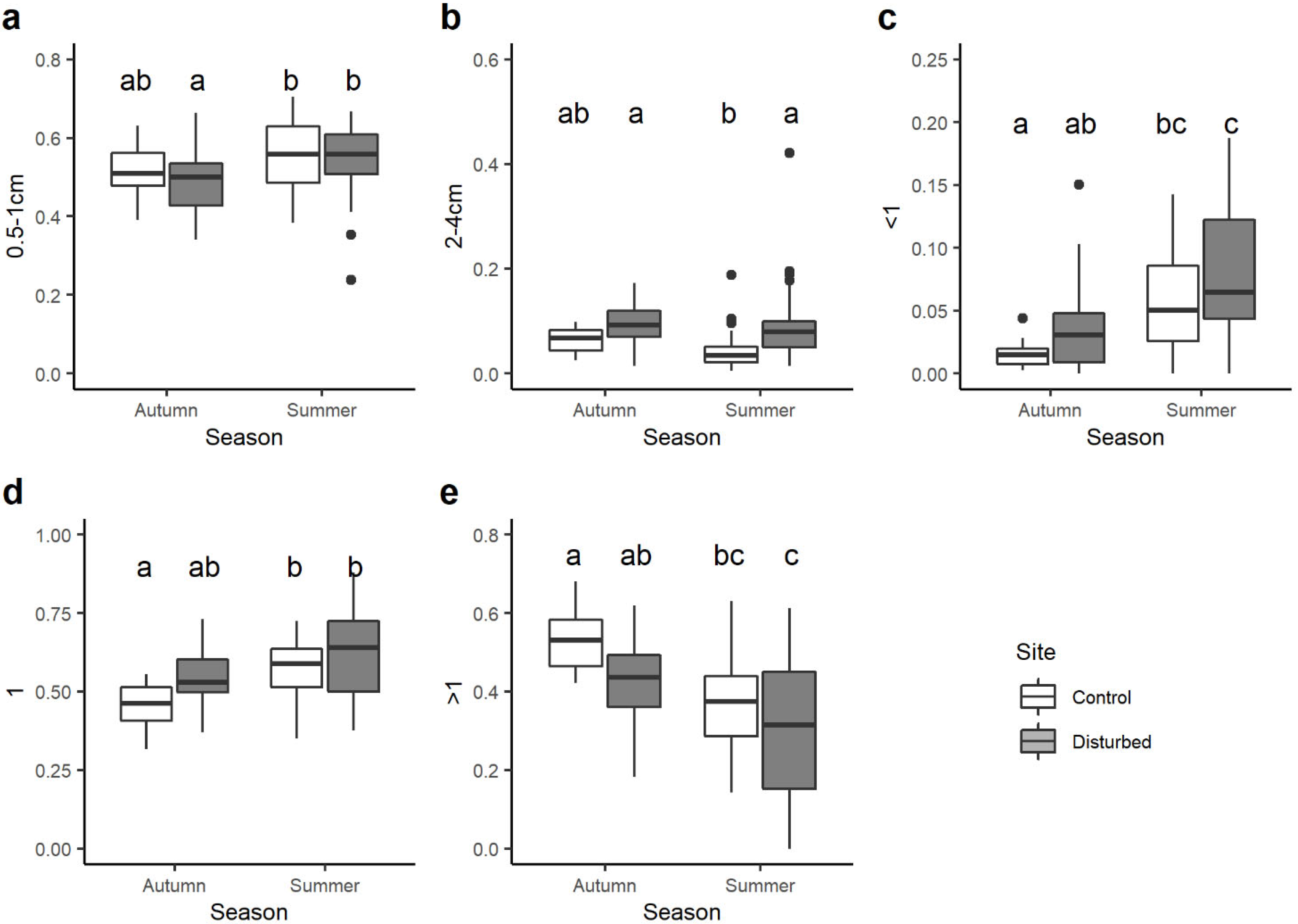
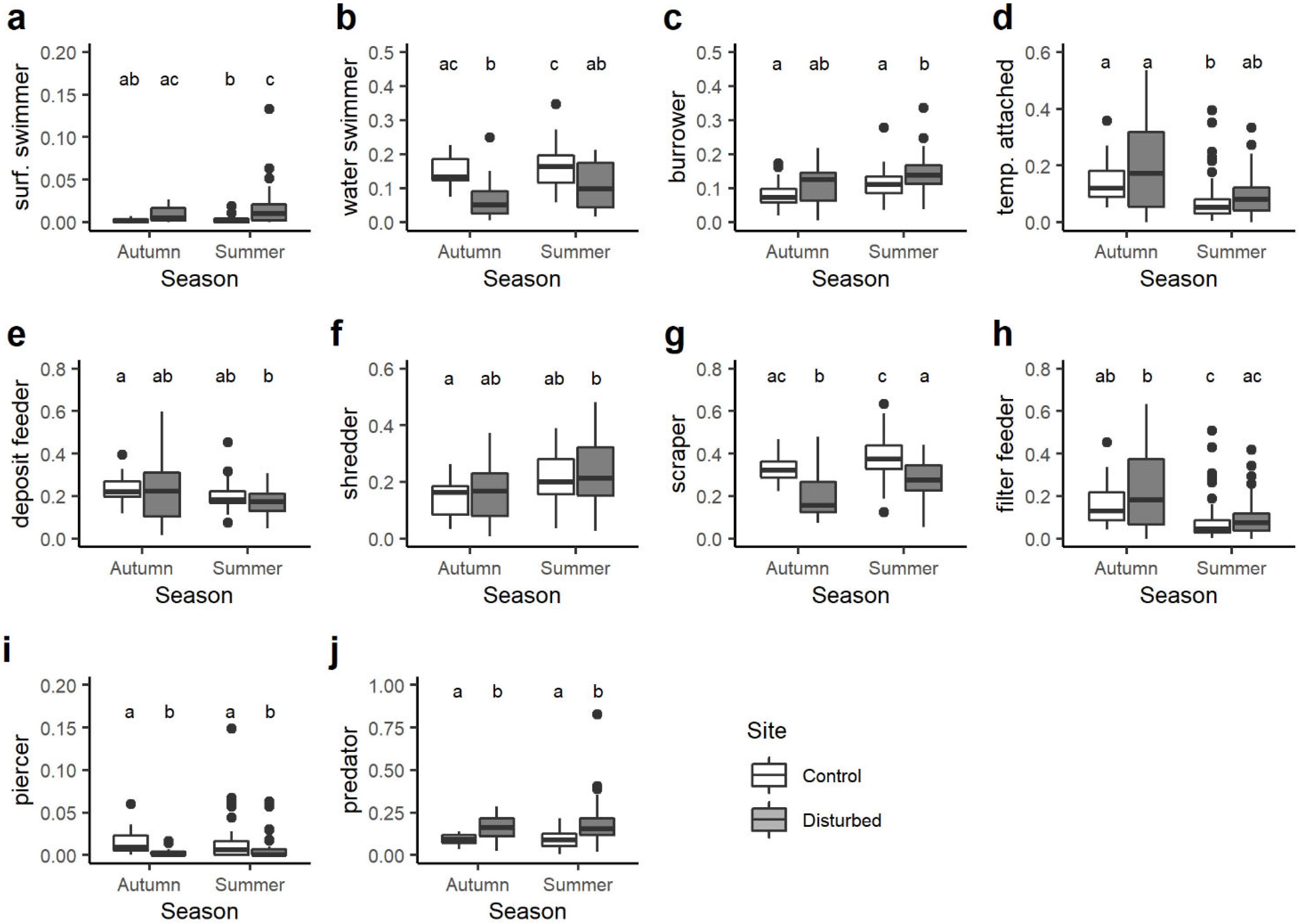
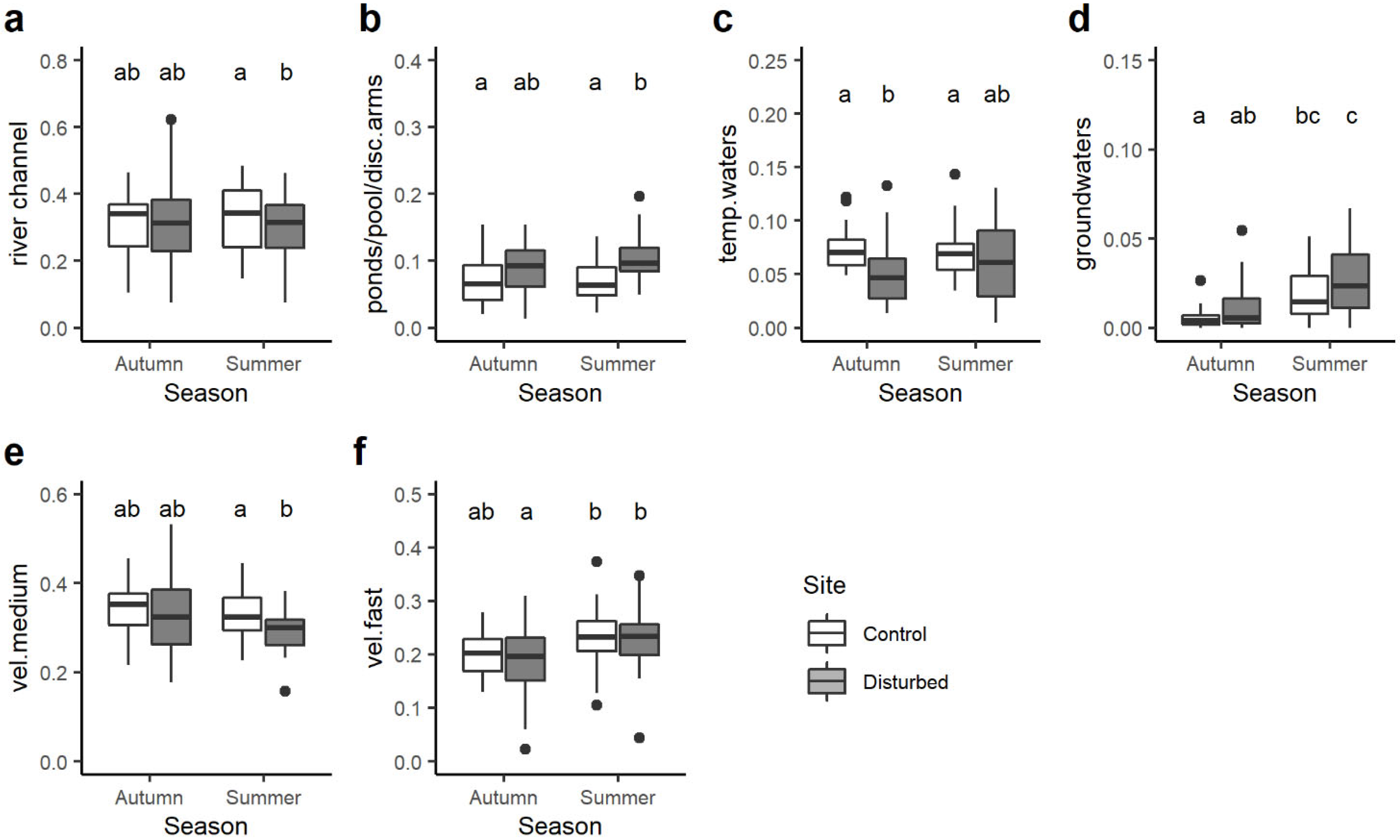
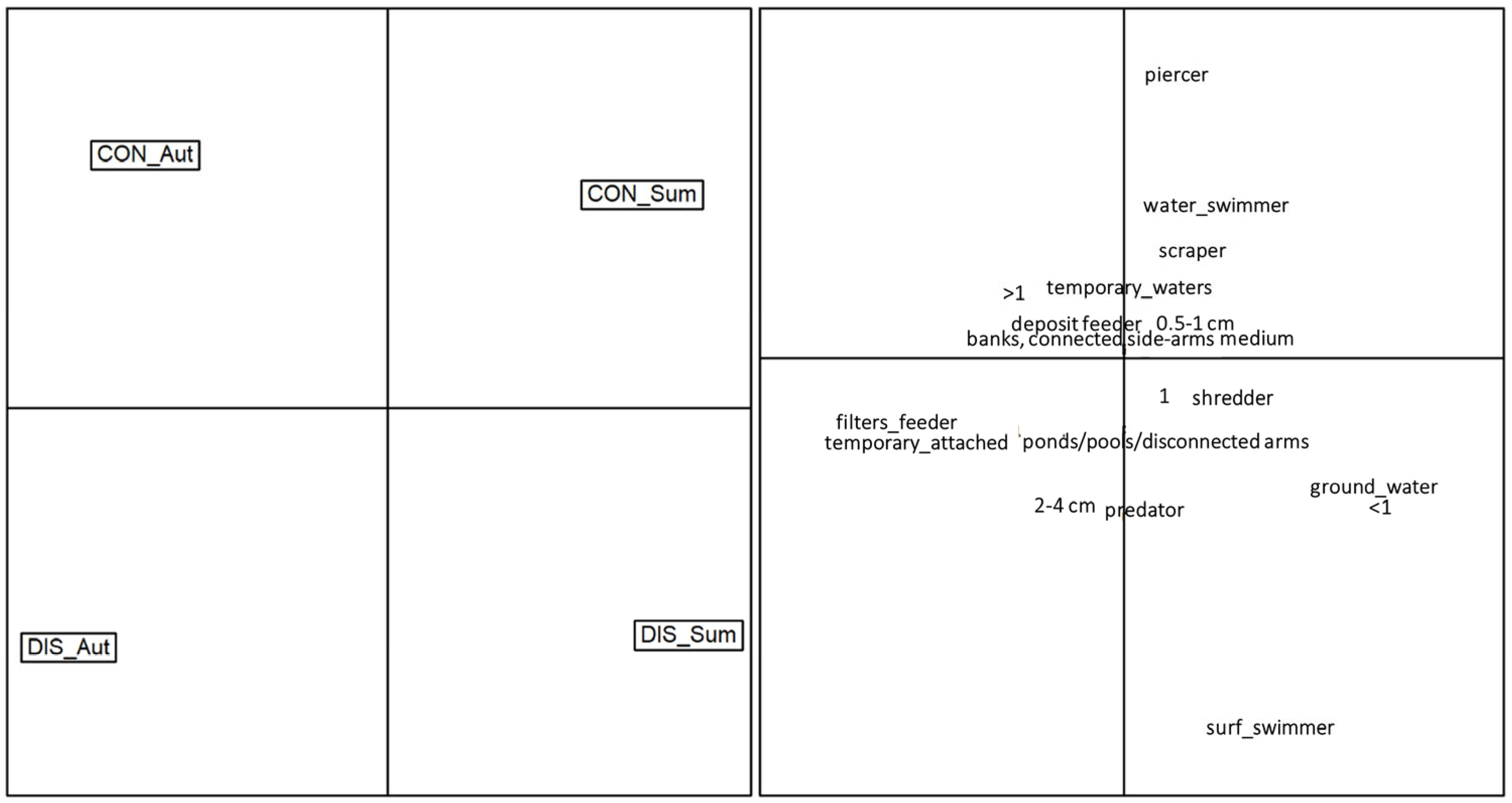
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gracey, E.O.; Verones, F. Impacts from hydropower production on biodiversity in an LCA framework—Review and recommendations. Int. J. LCA 2016, 21, 412–428. [Google Scholar] [CrossRef]
- Wolf, S.; Esser, V.; Schüttrumpf, H.; Lehmkuhl, F. Influence of 200 years of water resource management on a typical central European river. Does industrialization straighten a river? Environ. Sci. Eur. 2021, 33, 15. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M.; Capps, K.A. Stream Ecology: Structure and Function of Running Waters, 3rd ed.; Springer Nature: Dordrecht, The Netherlands, 2021; p. 485. [Google Scholar]
- Meybeck, M. River water quality global ranges, time and space variabilities, proposal for some redefinitions. Int. Ver. Theor. Angew. Limnol. Verh. 1996, 26, 81–96. [Google Scholar] [CrossRef]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R960–R967. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, A.; Hering, D.; Feld, C.K.; Rolauffs, P. A new method for assessing the impact of hydromorphological degradation on the macroinvertebrate fauna of five German stream types. Hydrobiologia 2004, 516, 107–127. [Google Scholar] [CrossRef]
- Sabater, S.; Tockner, K. Effects of hydrologic alterations on the ecological quality of river ecosystems. In Water Scarcity in the Mediterranean. Perspectives Under Global Change; Sabater, S., Marcelo, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 15–39. [Google Scholar]
- Aquiloni, L.; Martín, M.P.; Gherardi, F.; Diéguez-Uribeondo, J. The North American crayfish Procambarus clarkii is the carrier of the oomycete Aphanomyces astaci in Italy. Biol. Invasions 2011, 13, 359–367. [Google Scholar] [CrossRef]
- Bo, T.; Fenoglio, S. Impacts of a micro-sewage effluent on the biota of a small Apennine creek. J. Freshw. Ecol. 2011, 4, 537–545. [Google Scholar] [CrossRef]
- Roberts, J.J.; Fausch, K.D.; Peterson, D.P.; Hooten, M.B. Fragmentation and thermal risks from climate change interact to affect persistence of native trout in the Colorado River basin. Glob. Chang. Biol. 2013, 19, 1383–1398. [Google Scholar] [CrossRef] [PubMed]
- Guareschi, S.; Laini, A.; Racchetti, E.; Bo, T.; Fenoglio, S.; Bartoli, M. How do hydromorphological constraints and regulated flows govern macroinvertebrate communities along an entire lowland river? Ecohydrology 2014, 7, 366–377. [Google Scholar] [CrossRef]
- Dewson, Z.S.; James, A.B.; Death, R.G. A review of the consequences of decreased flow for instream habitat and macroinvertebrates. J. N. Am. Benthol. Soc. 2007, 26, 401–415. [Google Scholar] [CrossRef]
- Carolli, M.; Vanzo, D.; Siviglia, A.; Zolezzi, G.; Bruno, M.C.; Alfredsen, K. A simple procedure for the assessment of hydropeaking flow alterations applied to several European streams. Aquat. Sci. 2015, 77, 639–653. [Google Scholar] [CrossRef]
- Doretto, A.; Bo, T.; Bona, F.; Apostolo, M.; Bonetto, D.; Fenoglio, S. Effectiveness of artificial floods for benthic community recovery after sediment flushing from a dam. Environ. Monit. Assess 2019, 191, 88. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.C.; Doretto, A.; Boano, F.; Ridolfi, L.; Fenoglio, S. Role of the hyporheic zone in increasing the resilience of mountain streams progressively shifting from naturally perennial to intermittent systems. Water 2020, 12, 2034. [Google Scholar] [CrossRef]
- Bruno, M.C.; Vallefuoco, F.; Casari, A.; Larsen, S.; Dallafior, V.; Zolezzi, G. Moving waters to mitigate hydropeaking: A case study from the Italian Alps. River. Res. Appl. 2023, 39, 570–587. [Google Scholar] [CrossRef]
- Reindl, R.; Neuner, J.; Schletterer, M. Increased hydropower production and hydropeaking mitigation along the upper Inn River (Tyrol, Austria) with a combination of buffer reservoirs, diversion hydropower plants and retention basins. River Res. Appl. 2023, 39, 602–609. [Google Scholar] [CrossRef]
- Bipa, N.J.; Stradiotti, G.; Righetti, M.; Pisaturo, G.R. Impacts of hydropeaking: A systematic review. Sci. Total Environ. 2023, 912, 169251. [Google Scholar] [CrossRef] [PubMed]
- Leitner, P.; Borgwardt, F.; Birk, S.; Graf, W. Multiple stressor effects on benthic macroinvertebrates in very large European rivers–A typology-based evaluation of faunal responses as a basis for future bioassessment. Sci. Total Environ. 2021, 756, 143472. [Google Scholar] [CrossRef] [PubMed]
- Quadroni, S.; Salmaso, F.; Gentili, G.; Crosa, G.; Espa, P. Response of benthic macroinvertebrates to different hydropower off-stream diversion schemes. Ecohydrology 2021, 14, e2267. [Google Scholar] [CrossRef]
- Salmaso, F.; Servanzi, L.; Crosa, G.; Quadroni, S.; Espa, P. Assessing the impacts of hydropeaking on river benthic macroinvertebrates: A state-of-the-art methodological overview. Environments 2021, 8, 67. [Google Scholar] [CrossRef]
- Schülting, L.; Dossi, F.; Graf, W.; Tonolla, D. Flow amplitude or up-ramping rate? Quantifying single and combined effects on macroinvertebrate drift during hydropeaking simulations, considering sensitive traits. River Res. Appl. 2023, 39, 412–426. [Google Scholar] [CrossRef]
- Smokorowski, K.E. The ups and downs of hydropeaking: A Canadian perspective on the need for, and ecological costs of, peaking hydropower production. Hydrobiologia 2022, 849, 421–441. [Google Scholar] [CrossRef]
- Tonolla, D.; Dossi, F.; Kastenhofer, O.; Doering, M.; Hauer, C.; Graf, W.; Schülting, L. Effects of hydropeaking on drift, stranding and community composition of macroinvertebrates: A field experimental approach in three regulated Swiss rivers. River Res. Appl. 2023, 39, 427–443. [Google Scholar] [CrossRef]
- Tiemann, J.S.; Gillette, D.P.; Wildhaber, M.L.; Edds, D.R. Effects of lowhead dams on riffle-dwelling fishes and macroinvertebrates in a midwestern river. Trans. Am. Fish. Soc. 2004, 133, 705–717. [Google Scholar] [CrossRef]
- Carter, J.L.; Resh, V.H.; Hannaford, M.J. Macroinvertebrates as Biotic Indicators of Environmental Quality. In Methods in Stream Ecology, 3rd ed.; Academic Press: London, UK, 2017; pp. 293–318. [Google Scholar]
- Woodcock, T.S.; Huryn, A.D. The response of macroinvertebrate production to a pollution gradient in a headwater stream. Freshw. Biol. 2007, 52, 177–196. [Google Scholar] [CrossRef]
- Erba, S.; Cazzola, M.; Belfiore, C.; Buffagni, A. Macroinvertebrate metrics responses to morphological alteration in Italian rivers. Hydrobiologia 2020, 847, 2169–2191. [Google Scholar] [CrossRef]
- Verberk, W.C.; Siepel, H.; Esselink, H. Life-history strategies in freshwater macroinvertebrates. Freshw. Biol. 2008, 53, 1722–1738. [Google Scholar] [CrossRef]
- Merritt, R.W.; Cummins, K.W.; Berg, M.B. Trophic relationships of macroinvertebrates. In Methods in Stream Ecology; Academic Press: Cambridge, MA, USA, 2017; Volume 1, pp. 413–433. [Google Scholar]
- Townsend, C.R.; Hildrew, A.G. Species traits in relation to habitat templet for river systems. Freshw. Biol. 1994, 31, 265–275. [Google Scholar] [CrossRef]
- Statzner, B.; Hildrew, A.G.; Resh, V.H. Species traits and environmental constraints: Entomological research and the history of ecological theory. Annu. Rev. Entomol. 2001, 46, 291–316. [Google Scholar] [CrossRef] [PubMed]
- Cummins, K.W. Invertebrates. In Rivers Handbook; Blackwell Scientific: Oxford, UK, 1995; pp. 234–250. [Google Scholar]
- Englund, G.; Malmqvist, B. Effects of flow regulations, habitat area and isolation on the macroinvertebrates fauna of rapids in north Swedish Rivers. River Res. Appl. 1996, 12, 433–445. [Google Scholar] [CrossRef]
- Fleituch, T. Structure and functional Organization of benthic Invertebrates in a Regulated Stream. Int. Rev. Hydrobiol. 2003, 88, 332–344. [Google Scholar] [CrossRef]
- Merritt, R.W.; Fenoglio, S.; Cummins, K.W. Promoting a functional macroinvertebrate approach in the biomonitoring of Italian lotic systems. J. Limnol. 2017, 76, 5–8. [Google Scholar] [CrossRef]
- Minshall, G.W.; Thomas, S.A.; Newbold, J.D.; Monaghan, M.T.; Cushing, C.E. Physical factors influencing fine organic particle transport and deposition in streams. J. N. Am. Benthol. Soc. 2000, 19, 1–16. [Google Scholar] [CrossRef]
- Bo, T.; Cammarata, M.; López-Rodríguez, M.J.; De Figueroa, J.M.T.; Baltieri, M.; Varese, P.; Fenoglio, S. The influence of water quality and macroinvertebrate colonization on the breakdown process of native and exotic leaf types in sub-alpine stream. J. Freshw. Ecol. 2014, 29, 159–169. [Google Scholar] [CrossRef]
- Erdozain, M.; Kidd, K.; Kreutzweiser, D.; Sibley, P. Increased reliance of stream macroinvertebrates on terrestrial food sources linked to forest management intensity. Ecol. Appl. 2019, 29, e01889. [Google Scholar] [CrossRef]
- Vallania, A.; Corigliano, M.D.C. The effect of regulation caused by a dam on the distribution of the functional feeding groups of the benthos in the sub basin of the Grande River (San Luis, Argentina). Environ. Monit. Assess. 2007, 124, 201–209. [Google Scholar] [CrossRef]
- Cereghino, R.; Cugny, P.; Lavandier, P. Influence of intermittent hydropeaking on the longitudinal zonation patterns of benthic invertebrates in a mountain stream. Int. Rev. Hydrobiol. 2002, 87, 47–60. [Google Scholar] [CrossRef]
- Meile, T.; Boillat, J.L.; Schleiss, A. Hydropeaking indicators for characterization of the upper-rhone river in Switzerland. Aquat. Sci. 2011, 73, 171–182. [Google Scholar] [CrossRef]
- Bo, T.; Cammarata, M.; Candiotto, A.; Fenoglio, S.; López-Rodríguez, M.J.; Tierno de Figueroa, J.M.; Malacarne, G. Feeding habits of Padogobius bonelli (Bonaparte, 1846) (Osteichthyes, Gobiidae): The importance of fish dimensions and hydrological conditions. Ital. J. Zool. 2011, 78, 390–397. [Google Scholar] [CrossRef]
- Fenoglio, S.; Bo, T.; Cammarata, M.; López-Rodríguez, M.J.; Tierno De Figueroa, J.M. Seasonal variation of allochthonous and autochthonous energy inputs in an alpine stream. J. Limnol. 2015, 74, 272–277. [Google Scholar] [CrossRef]
- Campaioli, S.; Ghetti, P.F.; Minelli, A.; Ruffo, S. Manuale per il Riconoscimento dei Macroinvertebrati Delle Acque Dolci Italiane (Vol. I); Museo di Storia Naturale di Trento: Trento, Italy, 1994; p. 357. [Google Scholar]
- Campaioli, S.; Ghetti, P.F.; Minelli, A.; Ruffo, S. Manuale per il Riconoscimento dei Macroinvertebrati Delle Acque Dolci Italiane (Vol. II); Museo di Storia Naturale di Trento: Trento, Italy, 1994; p. 120. [Google Scholar]
- Tachet, H.; Bournaud, M.; Richoux, P.; Usseglio-Polatera, P. Invertébrés d’Eau Douce: Systématique, Biologie, Écologie; CNRS Editions: Paris, France, 2002; p. 588. [Google Scholar]
- Villéger, S.; Mason, N.W.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Laliberté, E.; Legendre, P.; Shipley, B.; Laliberté, M.E. Package ‘FD’. Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology; R Package Version 1.0-12. 2014. Available online: https://CRAN.R-project.org/package=FD (accessed on 1 October 2023).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package, R Package Version 2.2-1. 2015. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 1 October 2023).
- Dray, S.; Siberchicot, M.A. Package ‘ade4’; Université de Lyon: Lyon, France, 2020. [Google Scholar]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed]
- Takao, A.; Kawaguchi, Y.; Minagawa, T.; Kayaba, Y.; Morimoto, Y. The relationships between benthic macroinvertebrates and biotic and abiotic environmental characteristics downstream of the Yahagi dam, central Japan, and the state change caused by inflow from a tributary. River. Res. Applic. 2008, 24, 580–597. [Google Scholar] [CrossRef]
- Laini, A.; Viaroli, P.; Bolpagni, R.; Cancellario, T.; Racchetti, E.; Guareschi, S. Taxonomic and functional responses of benthic macroinvertebrate communities to hydrological and water quality variations in a heavily regulated river. Water 2019, 11, 1478. [Google Scholar] [CrossRef]
- Fornaroli, R.; White, J.C.; Boggero, A.; Laini, A. Spatial and temporal patterns of macroinvertebrate assemblages in the River Po Catchment (Northern Italy). Water 2020, 12, 2452. [Google Scholar] [CrossRef]
- Ding, N.; Yang, W.; Zhou, Y.; Gonzalez-Bergonzoni, I.; Zhang, J.; Chen, K.; Vidal, N.; Jeppesen, E.; Liu, Z.; Wang, B. Different responses of functional traits and diversity of stream macroinvertebrates to environmental and spatial factors in the Xishuangbanna watershed of the upper Mekong River Basin, China. Sci. Total Environ. 2017, 574, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Rajper, A.R.; Hughes, R.M.; Olson, J.R.; Wei, H.; Wang, B. Incorporating functional traits to enhance multimetric index performance and assess land use gradients. Sci. Total Environ. 2019, 691, 1005–1015. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Leiva, M.; Marchese, M.; Lorenz, G.; Diodato, L. Functional diversity of benthic macroinvertebrates regarding hydrological and land use disturbances in a heavily impaired lowland river. Limnologica 2022, 92, 125940. [Google Scholar] [CrossRef]
- Usseglio-Polatera, P.; Bournaud, P.M.; Richoux, H. Tachet. Biological and ecological traits of benthic freshwater macroinvertebrates: Relationships and definition of groups with similar traits. Freshw. Biol. 2000, 43, 175–205. [Google Scholar] [CrossRef]
- Mouillot, S.; Spatharis, S.; Reizopoulou, T.; Laugier, L.; Sabetta, A.; Basset, T.; Chi, T. Alternatives to taxonomic-based approaches to assess changes in transitional water communities. Aquat. Conserv. Mar. Freshw. Ecosyst. 2006, 16, 469–482. [Google Scholar] [CrossRef]
- Rabení, C.F.; Doisy, K.E.; Zweig, L.D. Stream invertebrate community functional responses to deposited sediment. Aquat. Sci. 2005, 67, 395–402. [Google Scholar] [CrossRef]
- Yap, N.I.R.; Kamarudin, K.R.; Rehan, A.M.; Badrulhisham, N.S.; Zakaria, M.Z.; Kemalok, J. Aquatic Insects as Bio-Indicators of Water Quality–A Study on Sungai Kawal, Johor National Park of Endau-Rompin, Peninsular Malaysia. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 736, No. 2, p. 012072. [Google Scholar]
- Céréghino, R.; Oertli, B.; Bazzanti, M.; Coccia, C.; Compin, A.; Biggs, J.; Bressi, N.; Grillas, P.; Hull, A.; Kalettka, T.; et al. Biological traits of European pond macroinvertebrates. Hydrobiologia 2012, 689, 51–61. [Google Scholar] [CrossRef]
- Owusu, A.; Mul, M.; Strauch, M.; van der Zaag, P.; Volk, M.; Slinger, J. The clam and the dam: A Bayesian belief network approach to environmental flow assessment in a data scarce region. Sci. Total Environ. 2022, 810, 151315. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control Site | Disturbed Site |
|---|---|---|
| Conductivity (µS/cm) | 185 ± 37.4 | 170 ± 21.8 |
| pH | 7.41 ± 0.5 | 7.50 ± 0.2 |
| BOD5 (mg/L) | 2.50 ± 0.2 | 3.00 ± 0.2 |
| COD (mg/L) | 6.30 ± 0.1 | 7.60 ± 0.1 |
| Total phosphorous (mg/L) mg P/L | <0.05 | <0.05 |
| Ammonia nitrogen (mg/L) mg N/L | <0.05 | <0.05 |
| Daphnia magna (acute toxicity) | N.T. | N.T. |
| Vibrio fischeri (acute toxicity) | N.T. | N.T. |
| % Natural areas | 62% | 71% |
| % Agricultural areas | 26% | 23% |
| % Urbanized areas | 12% | 6% |
| Category | Trait |
|---|---|
| Maximal potential body size | ≤0.25 cm |
| 0.25–0.5 cm | |
| 0.5–1 cm | |
| 1–2 cm | |
| 2–4 cm | |
| 4–8 cm | |
| 8 cm | |
| Potential number of cycles per year | <1 |
| 1 | |
| >1 | |
| Locomotion and substrate relation | Flier |
| surface swimmer | |
| full water swimmer | |
| crawler | |
| burrower | |
| interstitial | |
| temporarily attached | |
| permanently attached | |
| Feeding habits | absorber |
| deposit feeder | |
| shredder | |
| scraper | |
| filter-feeder | |
| piercer | |
| predator | |
| parasite | |
| Transversal distribution | river channel |
| banks, connected side-arms | |
| ponds, pools, disconnected side-arms | |
| marshes, peat bogs | |
| temporary waters | |
| lakes | |
| groundwaters | |
| Current velocity (preferendum) | null |
| slow | |
| medium | |
| fast |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marino, A.; Bona, F.; Fenoglio, S.; Bo, T. Functional Traits Drive the Changes in Diversity and Composition of Benthic Invertebrate Communities in Response to Hydrological Regulation. Water 2024, 16, 989. https://doi.org/10.3390/w16070989
Marino A, Bona F, Fenoglio S, Bo T. Functional Traits Drive the Changes in Diversity and Composition of Benthic Invertebrate Communities in Response to Hydrological Regulation. Water. 2024; 16(7):989. https://doi.org/10.3390/w16070989
Chicago/Turabian StyleMarino, Anna, Francesca Bona, Stefano Fenoglio, and Tiziano Bo. 2024. "Functional Traits Drive the Changes in Diversity and Composition of Benthic Invertebrate Communities in Response to Hydrological Regulation" Water 16, no. 7: 989. https://doi.org/10.3390/w16070989
APA StyleMarino, A., Bona, F., Fenoglio, S., & Bo, T. (2024). Functional Traits Drive the Changes in Diversity and Composition of Benthic Invertebrate Communities in Response to Hydrological Regulation. Water, 16(7), 989. https://doi.org/10.3390/w16070989