
Morphological Characteristics and Annual Population Dynamics of Microcystis (Cyanobacteria) in Nanwan Reservoir (Xinyang, China)

 ,
,
Abstract
1. Introduction
2. Materials and Methods
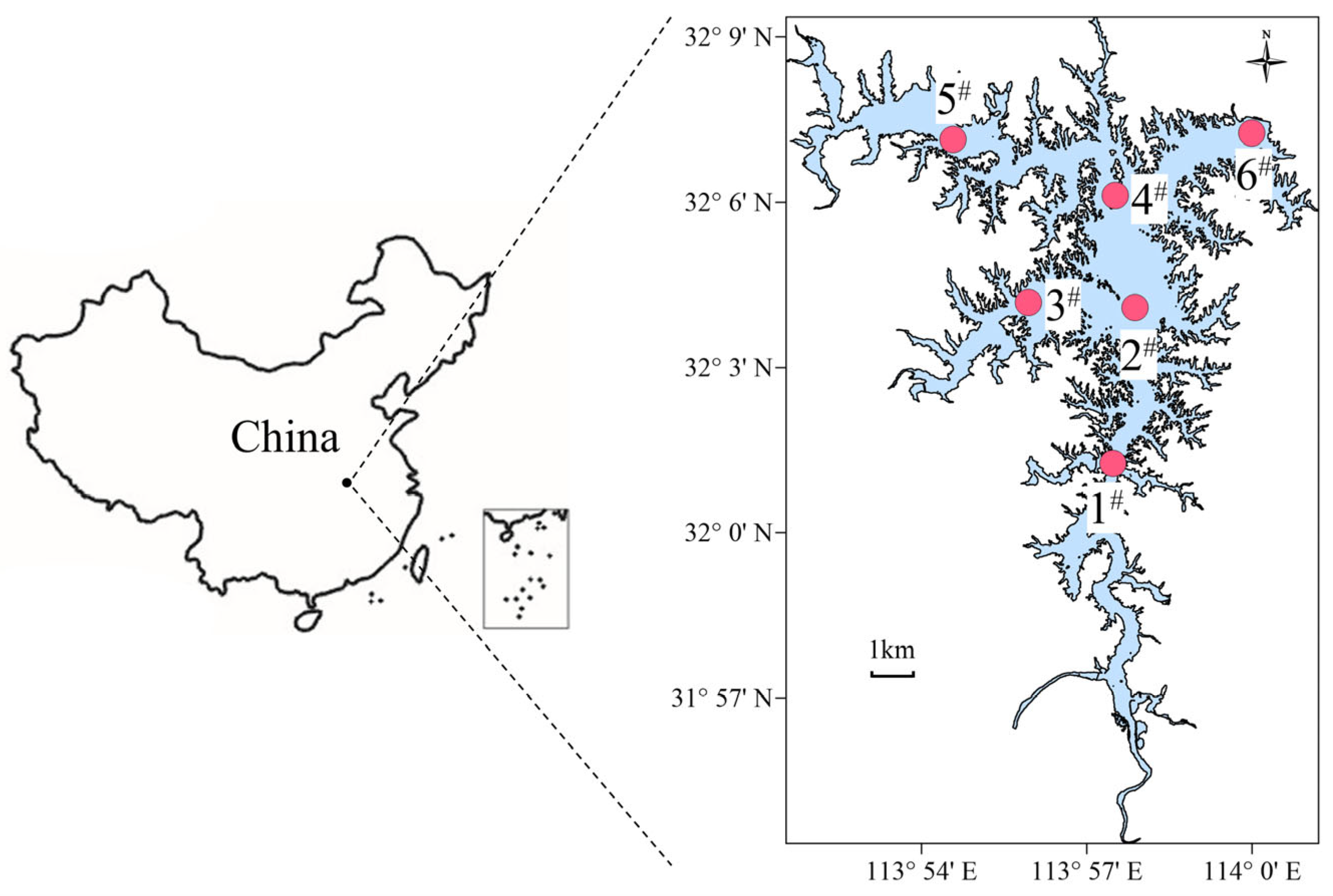
2.1. Study Area
2.2. Sampling Sites and Sample Collection
2.3. Nutrient Measurement
2.4. Species Identification and Population Density
2.5. Data Processing and Statistical Analysis
3. Results
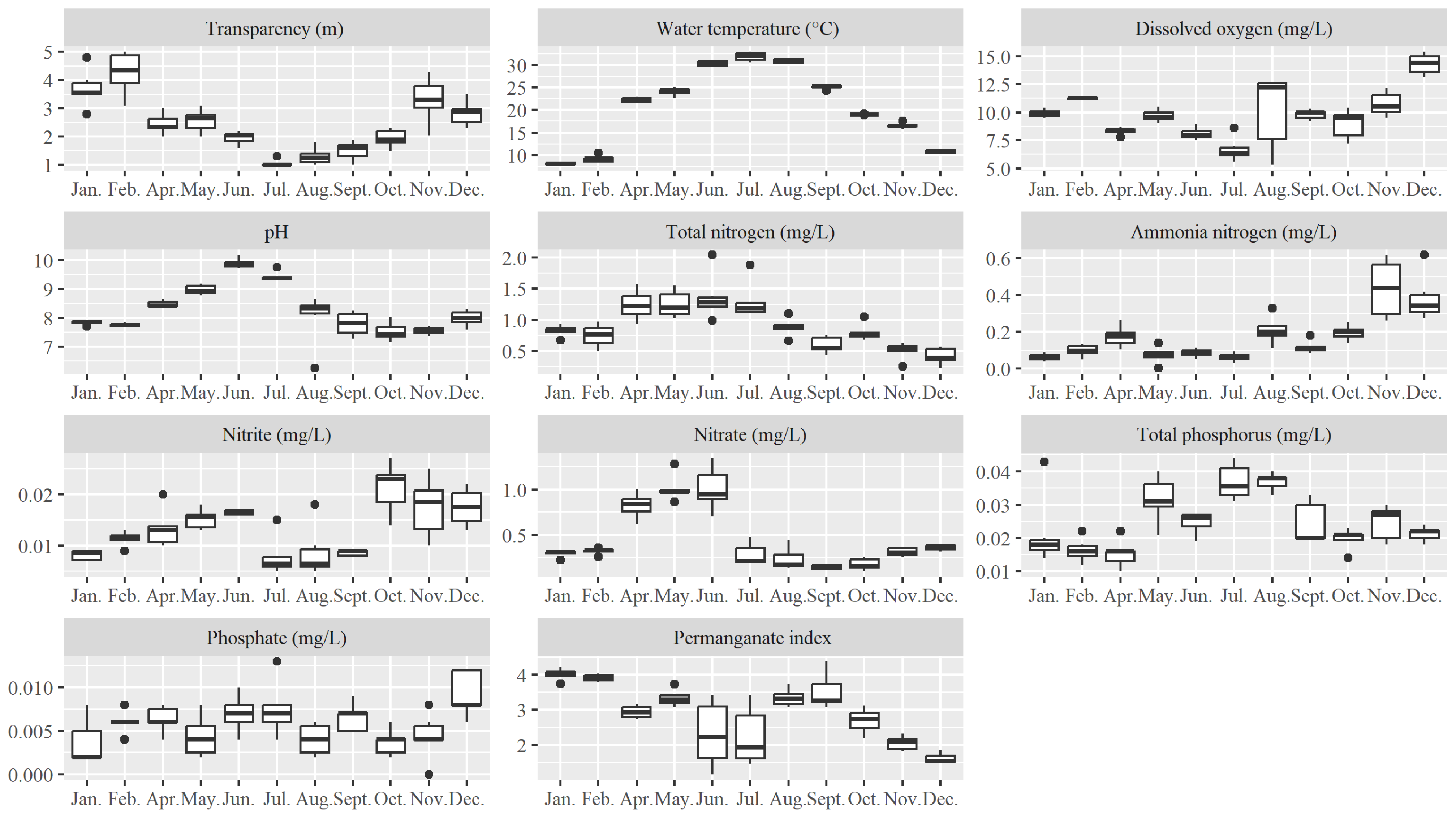
3.1. Monthly Variation of Physicochemical Factors
3.2. Characteristics of Microcystis Species
3.3. Variation in Microcystis Density
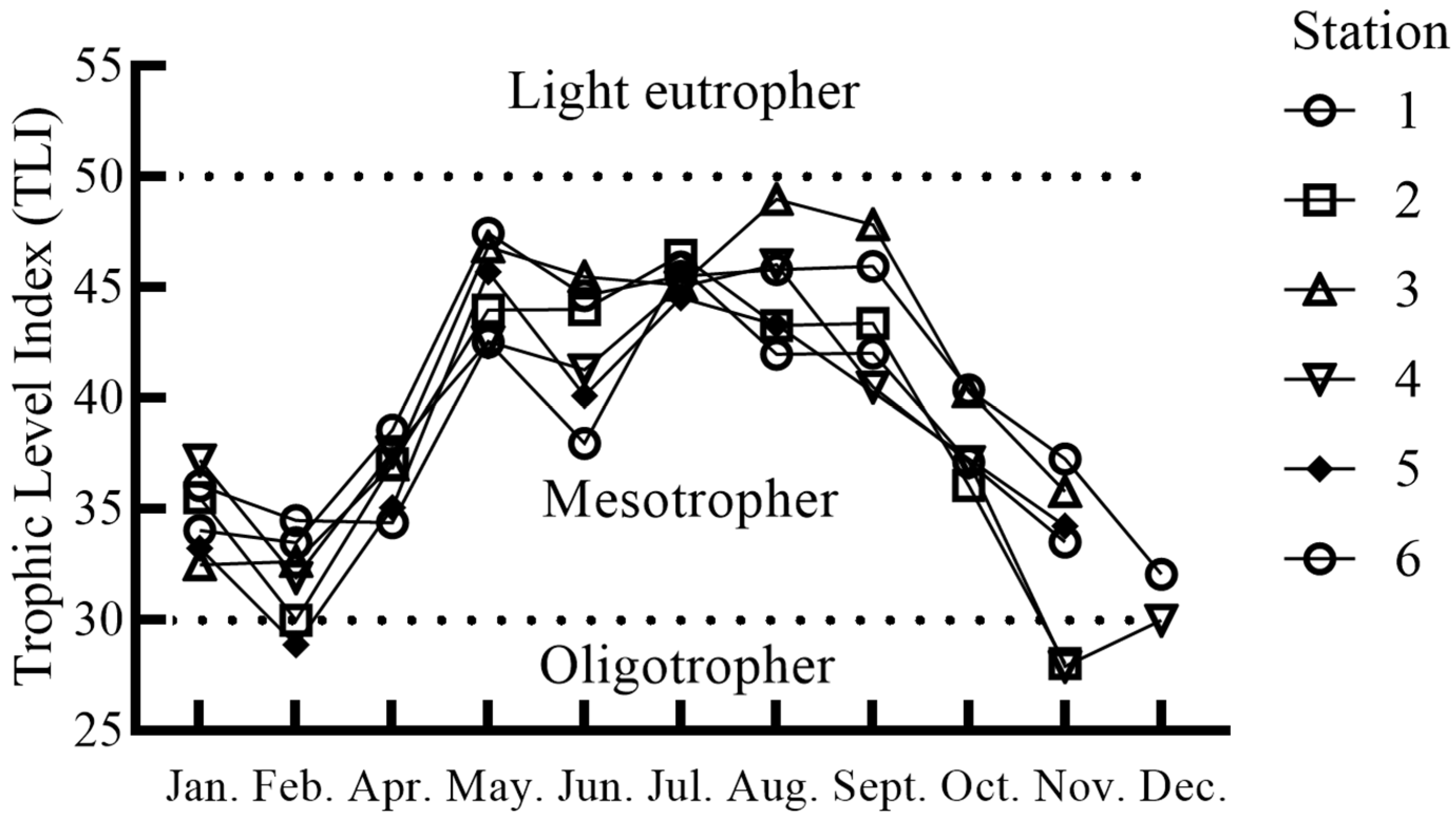
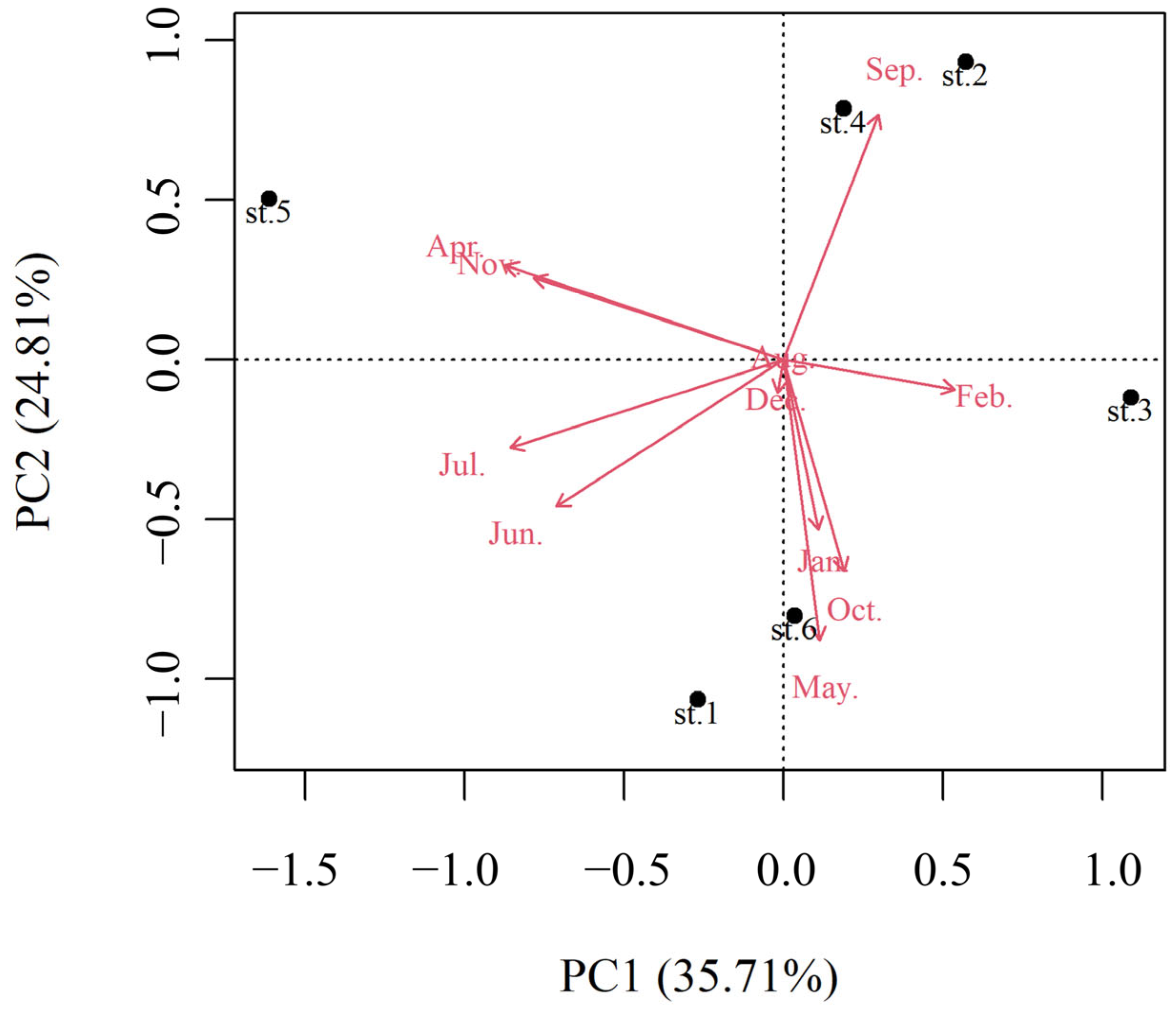
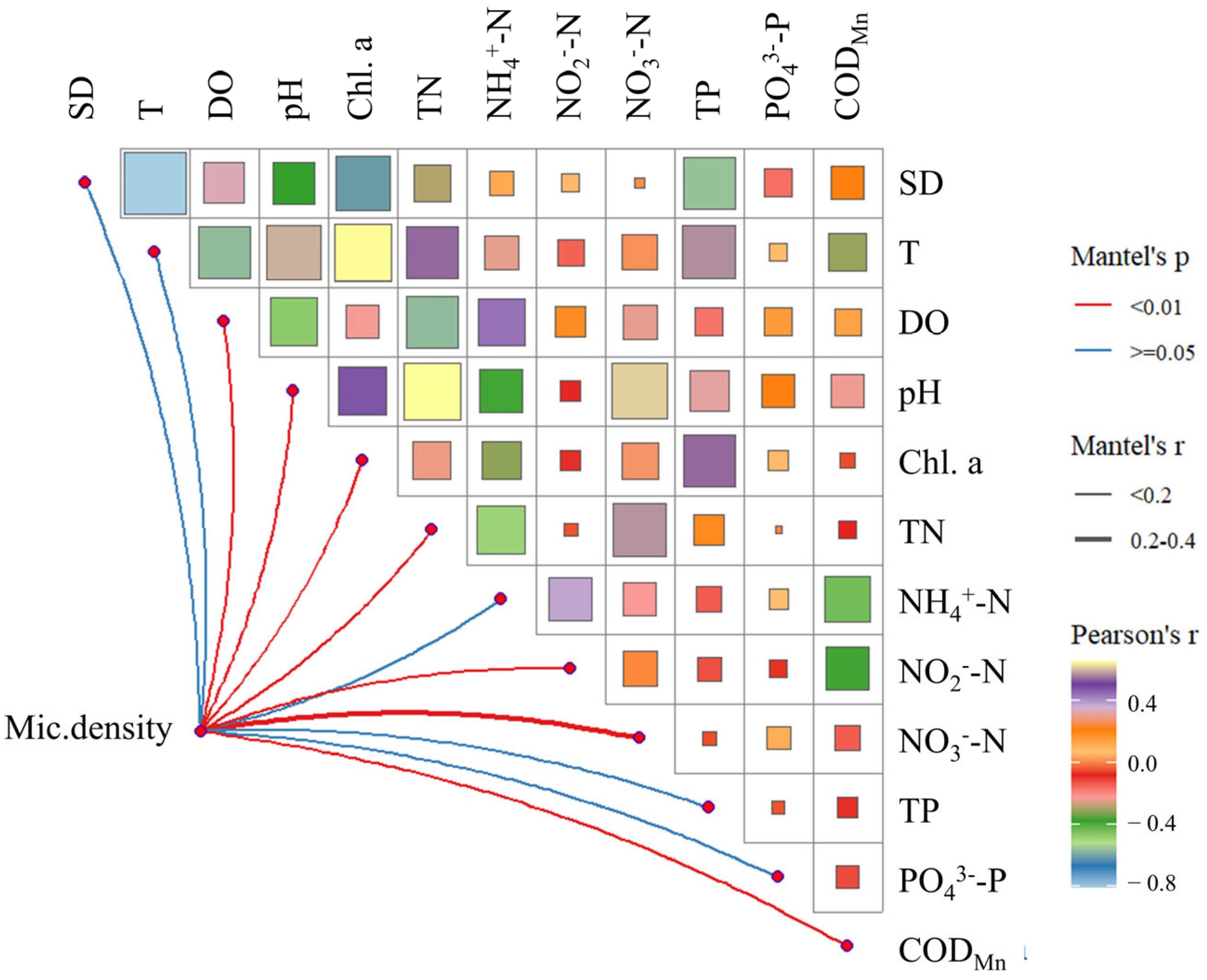
3.4. Relationship Between Microcystis Density and Physicochemical Factors
4. Discussion
4.1. Microcystis Cell Features and Diversity
4.2. Variation of Microcystis Density
4.3. Driving Factors of Microcystis Abundance Variation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Environ. Microbiolol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K.; Eggleton, M.A.; Lochmann, R.T. An environmentally friendly approach for mitigating cyanobacterial bloom and their toxins in hypereutrophic ponds: Potentiality of a newly developed granular hydrogen peroxide-based compound. Sci. Total Environ. 2018, 637–638, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Zhu, G.; Gao, G.; Zhang, Y.; Li, W.; Paerl, H.W.; Carmichael, W.W. A drinking water crisis in Lake Taihu, China: Linkage to climatic variability and lake management. Environ. Manag. 2010, 45, 105–112. [Google Scholar] [CrossRef]
- Alonso-Andicoberry, C.; García-Viliada, L.; Lopez-Rodas, V.; Costas, E. Catastrophic mortality of flamingos in a Spanish national park caused by cyanobacteria. Vet. Rec. 2002, 151, 706. [Google Scholar]
- Komárek, J.; Komárková, J. Review of the European Microcystis-morphospecies (Cyanoprokaryotes) from nature. Czech Phycol. 2002, 2, 1–24. [Google Scholar]
- Yu, G.L.; Song, L.R.; Li, R.H. Taxonomic notes on water bloom forming Microcystis species (Cyanophyta) from China—An example from samples of the Dianchi Lake. Acta Phytotax. Sin. 2007, 45, 727–741, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zhang, J.Y.; Zhu, B.C.; Wu, Z.J.; Xu, T.; Lu, Z.h. Microcystis panniformis-A newly recorded species of Microcystis in China. J. Lake Sci. 2012, 24, 647–650, (In Chinese with English Abstract). [Google Scholar]
- Lv, X.J.; Cheng, Y.; Zhang, S.L.; Hu, Z.W.; Xiao, P.; Zhang, H.; Geng, R.Z.; Li, R.H. Polyphasic Characterization and Taxonomic Evaluation of a Bloom-Forming Strain Morphologically Resembling Radiocystis fernandoi (Chroococcales, Cyanobacteria) from Lake Erhai, China. Diversity 2022, 14, 816. [Google Scholar] [CrossRef]
- Wu, Z.X.; Gan, N.Q.; Song, L.R. Genetic diversity: Geographical distribution and toxin profiles of Microcystis strains (Cyanobacteria) in China. J. Integr. Plant Biol. 2007, 49, 262–269. [Google Scholar] [CrossRef]
- Bolch, C.J.; Blackburn, S.I. Isolation and purification of Australian isolates of the toxic cyanobacterium Microcystis aeruginosa Kütz. J. Appl. Phycol. 1996, 8, 5–13. [Google Scholar] [CrossRef]
- Xu, S.; Sun, Q.; Zhou, X.; Tan, X.; Xiao, M.; Zhu, W.; Li, M. Polysaccharide biosynthesis-related genes explain phenotype-genotype correlation of Microcystis colonies in Meiliang Bay of Lake Taihu, China. Sci. Rep. 2016, 6, 35551. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Q.; Li, M.; Hu, R.; Lei, L. Sequence analysis of gyrB Gene from Microcystis in Lake Taihu and Tangxi Reservoir (Guangdong Province). J. Lake Sci. 2010, 22, 221–226, (In Chinese with English Abstract). [Google Scholar]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef]
- Cellamare, M.; Duval, C.; Drelin, Y.; Djediat, C.; Touibi, N.; Agogué, H.; Leboulanger, C.; Ader, M.; Bernard, C. Characterization of phototrophic microorganisms and description of new cyanobacteria isolated from the saline-alkaline crater-lake Dziani Dzaha (Mayotte, Indian Ocean). FEMS Microbiol. Ecol. 2018, 94, fiy108. [Google Scholar] [CrossRef] [PubMed]
- Brunberg, A.-K.; Blomqvist, P. Recruitment of Microcystis (Cyanophyceae) from Lake Sediments: The importance of Littoral Inocula. J. Phycol. 2003, 39, 58–63. [Google Scholar] [CrossRef]
- Ma, J.R.; Qin, B.Q.; Wu, P.; Zhou, J.; Niu, C.; Deng, J.M.; Niu, H.L. Controlling cyanobacterial blooms by managing nutrient ratio and limitation in a large hyper-eutrophic lake: Lake Taihu, China. J. Environ. Sci. 2015, 27, 80–86. [Google Scholar] [CrossRef]
- An, K.G.; Jones, J.R. Factors regulating bluegreen dominance in a reservoir directly influenced by the Asian monsoon. Hydrobiologia 2000, 432, 37–48. [Google Scholar] [CrossRef]
- Ma, J.; Brookes, J.D.; Qin, B.; Paerl, H.W.; Gao, G.; Wu, P.; Zhang, W.; Deng, J.; Zhu, G.; Zhang, Y. Environmental factors controlling colony formation in blooms of the cyanobacteria Microcystis spp. in Lake Taihu, China. Harmful Algae 2014, 31, 136–142. [Google Scholar] [CrossRef]
- Yu, G.Z.; Su, H.; Liu, X.C.; Zhao, C.M.; Gao, H.; Huang, K. Effects of function changes of the large reservoirs in Xinyang of Henan Province on water qualities—Usig Nanwan Reservoir as an example. Areal Res. Dev. 2009, 28, 115–119, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Fu, M.; Wang, Z.; Liang, X. Floristic composition and plant community diversity of water-level fluctuation zone of Nanwan Reservoir. Desalination Water Treat. 2023, 291, 233–241. [Google Scholar] [CrossRef]
- Hu, J.; Chi, S.; Hu, J. Studies on stress response of phytoplankton community in Nanwan Reservoir based on ABC curves. Trans. Oceanol. Limnol. 2022, 44, 64–72, (In Chinese with English Abstract). [Google Scholar]
- Sun, G.; Duan, J.; Zhao, J. Water quality evaluation and influencing factor analysis of Nanwan Reservoir with a bayesian model based on AHM-CRITIC combination weighting. Pearl River 2023, 44, 84–93+103, (In Chinese with English Abstract). [Google Scholar]
- Zuo, J.; Yang, S.; Grossart, H.-P.; Xiao, P.; Zhang, H.; Sun, R.; Li, G.; Jiang, H.; Zhao, Q.; Jiao, M.; et al. Sequential decline in cyanobacterial, total prokaryotic, and eukaryotic responses to backward flow in a river connected to Lake Taihu. Water Res. 2024, 269, 122784. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.S. Methods for Monitoring and Analyzing Water and Wastewater, 4th ed.; China Environmental Science Press: Beijing, China, 2002. (In Chinese) [Google Scholar]
- Hu, H.J.; Wei, Y.X. The Freshwarter Algae of China-Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006; pp. 62–68. (In Chinese) [Google Scholar]
- Zhao, W. Hydrobiology, 2nd ed.; Agricultural Press of China: Beijing, China, 2005; pp. 16–31. (In Chinese) [Google Scholar]
- Liu, H.; He, B.; Zhou, Y.; Kutser, T.; Toming, K.; Feng, Q.; Yang, X.; Fu, C.; Yang, F.; Li, W. Trophic state assessment of optically diverse lakes using Sentinel-3-derived trophic level index. Int. J. Appl. Earth Obs. Geoinform. 2022, 114, 103026. [Google Scholar] [CrossRef]
- Li, R.; Xiao, K.; Zhao, G.; Huang, X.; Li, Z.; Wu, H.; Huang, X.; Pan, Y.; Liang, L. Comprehensive assessment of eutrophication and the mechanisms driving phytoplankton blooms in multifunctional reservoirs. Water 2024, 16, 1752. [Google Scholar] [CrossRef]
- Cook, K.V.; Li, C.; Cai, H.; Krumholz, L.R.; Hambright, K.D.; Paerl, H.W.; Steffen, M.M.; Wilson, A.E.; Burford, M.A.; Grossart, H.P. The global Microcystis interactome. Limnol. Oceanogr. 2020, 65, S194–S207. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, X.; Chen, H.; Gao, L.; Xiao, M.; Li, M. High nutrient concentration and temperature alleviated formation of large colonies of Microcystis: Evidence from field investigations and laboratory experiments. Water Res. 2016, 101, 167–175. [Google Scholar] [CrossRef]
- Sun, Q.; Zhu, W.; Li, M.; Tan, X. Morphological changes of Microcystis aeruginosa colonies in culture. J. Limnol. 2016, 75, 14–23. [Google Scholar] [CrossRef]
- Otsuka, S.; Suda, S.; Li, R.; Matsumoto, S.; Watanabe, M.M. Morphological variability of colonies of Microcystis morphospecies in culture. J. Gen. Appl. Microbiolol. 2000, 46, 39–50. [Google Scholar] [CrossRef]
- Carrillo, E.; Ferrero, L.M.; Alonso-Andicoberry, C.; Basanta, A.; Martín, A.; López-Rodas, V.; Costas, E. Interstrain variability in toxin production in populations of the cyanobacterium Microcystis aeruginosa from water-supply reservoirs of Andalusia and lagoons of Doñana National Park (southern Spain). Phycologia 2003, 42, 269–274. [Google Scholar] [CrossRef]
- Neilan, B.A. Detection and identification of cyanobacteria associated with toxic blooms: DNA amplification protocols. Phycologia 1996, 35, 147–155. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, Y.; Cen, J.; Lü, S.; Hu, H. Taxonomic study on Microcystis (Cyanophyta) from some scenic lakes in Guangzhou. J. Trop. Subtrop. Bot. 2010, 18, 405–414, (In Chinese with English Abstract). [Google Scholar]
- Wang, J.; Shi, Y.; Liu, Q.; Li, Z.; Zhang, M.; Xie, S. Taxonomic and molecular phylogenetics of bloom-forming algae from the Taiyuan section of Fenhe River, China. J. Lake Sci. 2018, 30, 1332–1342, (In Chinese with English abstract). [Google Scholar]
- Loudiki, M.; Oudra, B.; Sabour, B.; Sbiyyaa, B.; Vasconcelos, V. Taxonomy and geographic distribution of potential toxic cyanobacterial strains in Morocco. Ann. Limnol. 2002, 38, 101–108. [Google Scholar] [CrossRef]
- You, B. Assessment on present situation for sites of drinking water sources at Nanwan Reservoir in Xinyang city and countermeasures for protection. Water Resour. Prot. 2001, 4, 45–47+73, (In Chinese with English Abstract). [Google Scholar]
- DB44/T 2261-2020; Technical Specification for Classification and Monitoring of Algal Blooms. Guangdong Provincial Local Standard. Guangdong Market Supervision Administration: Guangzhou, China, 2020. (In Chinese)
- Zhu, C.; Yang, X.; Zhao, B.; Zhang, M.; Zhang, F. Spatiotemporal dynamics of phytoplankton and cyanotoxins in Chaohu Lake during summer cyanobacterial blooms of 2017. Environ. Monit. China 2018, 34, 103–112, (In Chinese with English Abstract). [Google Scholar]
- Su, Y.; Lin, J.; Lin, W.; Lan, R.; Cheng, Y.; Lin, Q.; Zhang, Y. Spatial and temporal dynamics of cyanobacteria Microcystis in Shanzi Reservoir of Fujian Province. J. Fujian Norm. Univ. (Nat. Sci. Ed.) 2016, 32, 63–69, (In Chinese with English Abstract). [Google Scholar]
- Liu, X.; Lu, X.; Chen, Y. The effects of temperature and nutrient ratios on Microcystis blooms in Lake Taihu, China: An 11-year investigation. Harmful Algae 2011, 10, 337–343. [Google Scholar] [CrossRef]
- Wu, Y.; Li, L.; Gan, N.; Zheng, L.; Ma, H.; Shan, K.; Liu, J.; Xiao, B.; Song, L. Seasonal dynamics of water bloom-forming Microcystis morphospecies and the associated extracellular microcystin concentrations in large, shallow, eutrophic Dianchi Lake. J. Environ. Sci. 2014, 26, 1921–1929. [Google Scholar] [CrossRef]
- Ger, K.A.; Urrutia-Cordero, P.; Frost, P.C.; Hansson, L.-A.; Sarnelle, O.; Wilson, A.E.; Lürling, M. The interaction between cyanobacteria and zooplankton in a more eutrophic world. Harmful Algae 2016, 54, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Gobler, C.J.; Burkholder, J.M.; Davis, T.W.; Harke, M.J.; Johengen, T.; Stow, C.A.; Van de Waal, D.B. The dual role of nitrogen supply in controlling the growth and toxicity of cyanobacterial blooms. Harmful Algae 2016, 54, 87–97. [Google Scholar] [CrossRef]
- Kitchens, C.M.; Johengen, T.H.; Davis, T.W. Establishing spatial and temporal patterns in Microcystis sediment seed stock viability and their relationship to subsequent bloom development in Western Lake Erie. PLoS ONE 2018, 13, e0206821. [Google Scholar] [CrossRef]
- Dick, G.J.; Duhaime, M.B.; Evans, J.T.; Errera, R.M.; Godwin, C.M.; Kharbush, J.J.; Nitschky, H.S.; Powers, M.K.A.; Vanderploeg, H.A.; Schmidt, K.C. The genetic and ecophysiological diversity of Microcystis. Environ. Microbiolol. 2021, 23, 7278–7313. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Yao, Y.; Li, D.; Sun, T.; Leng, X. The influence of water quality on spatial-temporal pattern of phytoplankton diversity in Chaohu Lake. Wetl. Sci. Manag. 2024, 20, 50–54, (In Chinese with English Abstract). [Google Scholar]
- Sun, J.; Wang, J.; Yang, Y.; Chen, J.; Zhou, Q.; Zhong, W. Temporal and spatial characteristics and abnormal changes of nitrogen and phosphorus in Poyang lake and the impact pathways. Jiangxi Hydraul. Sci. Technol. 2024, 50, 199–205, (In Chinese with English Abstract). [Google Scholar]
- Chaffin, J.D.; Bridgeman, T.B.; Heckathorn, S.A.; Mishra, S. Assessment of Microcystis growth rate potential and nutrient status across a trophic gradient in western Lake Erie. J. Great Lakes Res. 2011, 37, 92–100. [Google Scholar] [CrossRef]
- Kromkamp, J.; van den Heuvel, A.; Mur, L.R. Phosphorus uptake and photosynthesis by phosphate-limited cultures of the cyanobacterium Microcystis aeruginosa. Br. Phycol. J. 1989, 24, 347–355. [Google Scholar] [CrossRef]
- Duhamel, S.; Nogaro, G.; Steinman, A.D. Effects of water level fluctuation and sediment–water nutrient exchange on phosphorus biogeochemistry in two coastal wetlands. Aquat. Sci. 2017, 79, 57–72. [Google Scholar] [CrossRef]
- Chen, M.; Ding, S.; Chen, X.; Sun, Q.; Fan, X.; Lin, J.; Ren, M.; Yang, L.; Zhang, C. Mechanisms driving phosphorus release during algal blooms based on hourly changes in iron and phosphorus concentrations in sediments. Water Res. 2018, 133, 153–164. [Google Scholar] [CrossRef]
- Chaffin, J.D.; Bridgeman, T.B.; Bade, D.L. Nitrogen constrains the growth of late summer cyanobacterial blooms in Lake Erie. Adv. Microbiol. 2013, 3, 16–26. [Google Scholar] [CrossRef]
- Chaffin, J.D.; Davis, T.W.; Smith, D.J.; Baer, M.M.; Dick, G.J. Interactions between nitrogen form, loading rate, and light intensity on Microcystis and Planktothrix growth and microcystin production. Harmful Algae 2018, 73, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Schampera, C.; Hellweger, F.L. Nitrogen availability controls response of microcystin concentration to phosphorus reduction: Evidence from model application to multiple lakes. Harmful Algae 2024, 139, 102711. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, J.; Yin, W.; Jia, H.; Xu, J.; Hao, R.; Zhong, Z.; Shi, Z. Linking water environmental factors and the local watershed landscape to the chlorophyll a concentration in reservoir bays. Sci. Total Environ. 2020, 758, 143617. [Google Scholar] [CrossRef]
- Qin, Q.; Shen, J. Physical transport processes affect the origins of harmful algal blooms in estuaries. Harmful Algae 2019, 84, 210–221. [Google Scholar] [CrossRef]
- Burford, M.A.; Johnson, S.A.; Cook, A.J.; Packer, T.V.; Taylor, B.M.; Townsley, E.R. Correlations between watershed and reservoir characteristics, and algal blooms in subtropical reservoirs. Water Res. 2007, 41, 4105–4114. [Google Scholar] [CrossRef]
- Jeong, K.-S.; Kim, D.-K.; Joo, G.-J. Delayed influence of dam storage and discharge on the determination of seasonal proliferations of Microcystis aeruginosa and Stephanodiscus hantzschii in a regulated river system of the lower Nakdong River (South Korea). Water Res. 2007, 41, 1269–1279. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model, R2 = 0.2 | ||||||
|---|---|---|---|---|---|---|
| Variables | n | Coefficient | SE | Standardized Coefficient | t | p |
| Intercept | 58 | −1,139,422.0 | 1,954,975.8 | −0.583 | 0.562 | |
| -N | 58 | 11,298,762.1 | 3,348,257.5 | 0.411 | 3.375 | <0.01 |
| Microcystis Species | Nanwan Reservoir (This Study) | Dianchi (Yunnan, China) [7] | Lakes in Guangzhou (Guangdong, China) [36] | Europe [6] |
|---|---|---|---|---|
| M. botrys | 5.22 ± 0.65 4.00 to 6.54 | 5.4 ± 0.51 4.3 to 6.5 | 4.0 to 6.0 | 4.9 to 6 |
| M. novacekii | 3.62 ± 0.47 2.90 to 5.09 | 4.9 3.7 to 5.8 | 3.5 to 6.2 | 2.4 to 6 |
| M. smithii | 4.65 ± 0.61 3.67 to 5.89 | 4.3 ± 0.73 2.5 to 6.0 | 2.2 to 5.5 | 3.2 to 5.6 |
| M. wesenbergii | 6.47 ± 0.80 4.67 to 8.34 | 6.4 ± 0.74 4.5 to 8.1 | 4.5 to 7.5 | 4 to 8.5 |
| M. flos-aquae | 6.32 ± 0.90 4.56 to 7.85 | 4.6 3.7 to 4.9 | 3.0 to 5.8 | 3.5 to 4.8 |
| M. aeruginosa | 4.71 ± 0.62 3.39 to 6.20 | 5.3 ± 0.59 3.8 to 6.3 | 3.5 to 6.0 | 4 to 6.5 |
| M. firma | 1.97 ± 0.56 1.26 to 3.36 | 3.0 ± 0.35 2.0 to 3.7 | 2.0 to 3.5 | 2 to 3.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Ju, C.; Wu, K.; Liu, X.; Zhang, H.; Guan, J.; Ge, Y.; Zhao, L.; Su, C.; Yang, T.; et al. Morphological Characteristics and Annual Population Dynamics of Microcystis (Cyanobacteria) in Nanwan Reservoir (Xinyang, China). Water 2024, 16, 3569. https://doi.org/10.3390/w16243569
Tian Y, Ju C, Wu K, Liu X, Zhang H, Guan J, Ge Y, Zhao L, Su C, Yang T, et al. Morphological Characteristics and Annual Population Dynamics of Microcystis (Cyanobacteria) in Nanwan Reservoir (Xinyang, China). Water. 2024; 16(24):3569. https://doi.org/10.3390/w16243569
Chicago/Turabian StyleTian, Yuan, Chenxi Ju, Kunjie Wu, Xin Liu, Haojun Zhang, Jiabao Guan, Yilin Ge, Liangjie Zhao, Chaoqun Su, Tiezhu Yang, and et al. 2024. "Morphological Characteristics and Annual Population Dynamics of Microcystis (Cyanobacteria) in Nanwan Reservoir (Xinyang, China)" Water 16, no. 24: 3569. https://doi.org/10.3390/w16243569
APA StyleTian, Y., Ju, C., Wu, K., Liu, X., Zhang, H., Guan, J., Ge, Y., Zhao, L., Su, C., Yang, T., & Ma, Y. (2024). Morphological Characteristics and Annual Population Dynamics of Microcystis (Cyanobacteria) in Nanwan Reservoir (Xinyang, China). Water, 16(24), 3569. https://doi.org/10.3390/w16243569