1. Introduction
Mountain regions provide water resources to downstream populations for agriculture, industrial activities, and human consumption [
1,
2]. In tropical mountain regions, water is mainly taken from surface and subsurface ecosystems. In mountain watersheds, water can be stored and released by a combination of different hydrological components including snowpack, glaciers, groundwater, and lakes [
3,
4]. Further, in the tropical high mountains, the páramo ecosystems [
5] are characterized by having special properties for storage and regulation of the hydrological cycle, since they have a high content of organic matter, high porosity, and low apparent density, which allows most of the water to be retained in the soil and slowly be delivered to the watercourses [
6], becoming the main regulator of the hydrological cycle in this type of ecosystems [
7].
In the tropical high mountains, human activities have strongly intensified in recent decades. Agricultural activities have moved upward to higher elevations, modifying forested areas into pasture and agricultural lands, resulting in a severe loss of biodiversity [
8] and increasing contamination by fertilizers and pesticides, which affect river water quality [
9]. The lack of sanitation systems in most tropical high mountain settlements contributes to the release of a high number of diverse contaminants, such as pathogens of fecal origin [
10].
In addition to the increasing contamination of tropical high mountain rivers, the modification of their channels also represents an alarming problem. Riverine vegetation is strongly affected or modified and, most of the time, eliminated [
11]. Uncontrolled construction of houses, roads, and/or other civil infrastructures up to the riverbanks is common in this region. Riverine vegetation is key for healthy biological communities and, as such, for ecosystem health [
12]. Different characteristics of riverine vegetation (i.e., percentage of vegetation cover, vegetation stratification, presence of native/introduced species, etc.) should be evaluated for ecosystem health assessment [
13,
14]. Further, the evaluation of fluvial habitat quality (i.e., occurrence of rheophile zones, sediment types, presence of shade, presence of natural elements and aquatic plants, etc.) also needs to be considered in biomonitoring programs and ecosystem health assessments [
15].
In addition to the above-mentioned anthropogenic impacts, climate change is likely to affect tropical high mountain river systems through the possible change of rainfall patterns, which would have an effect on water availability for human use [
16]. Cuenca, the third largest city of Ecuador with 505,585 inhabitants [
2], obtains its drinking water from the surface resources of seven surrounding catchments. Owing to the fast-increasing population and the present anthropogenic activities, this city will face drinking water availability problems by no later than the year 2050. The Cajas National Park (CNP) is the most important páramo ecosystem in the Cuenca canton and is responsible for a large part of the current and future water supply for the city [
5,
17].
Within the CNP, the main economic activities developed are tourism and fishing [
18]. However, while human intervention in the headwaters of the CNP rivers is relatively scarce and is quite controlled, the impact in the middle and lower sectors of the rivers is substantially greater, threatening the ecological integrity of the catchments [
2]. This is despite the fact that the sanitation control of the city of Cuenca has a combined sewage system and a network of sanitary interceptors and wastewater treatment plants, particularly the Ucubamba plant that treats about 95% of the wastewater of the city, and perimeter areas whose sewer networks are intercepted [
19,
20]. The remaining wastewater from the urban areas is treated in small plants. Nevertheless, there are important perimeter areas that have developed recently, the sewer networks of which are not intercepted and conveyed to treatment plants [
20]; thus, an additional treatment plant and supplementary sanitary interceptors are projected [
19].
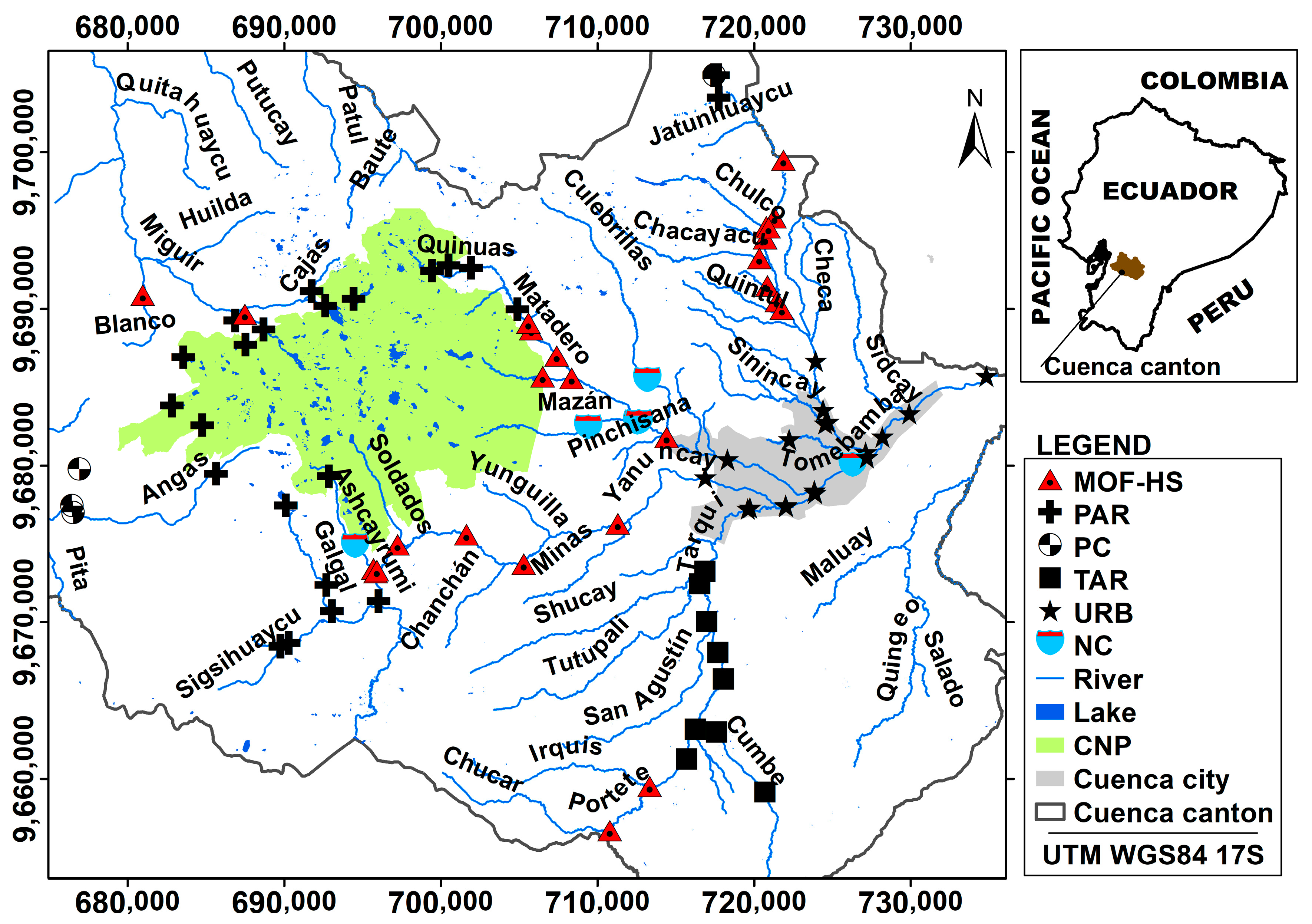
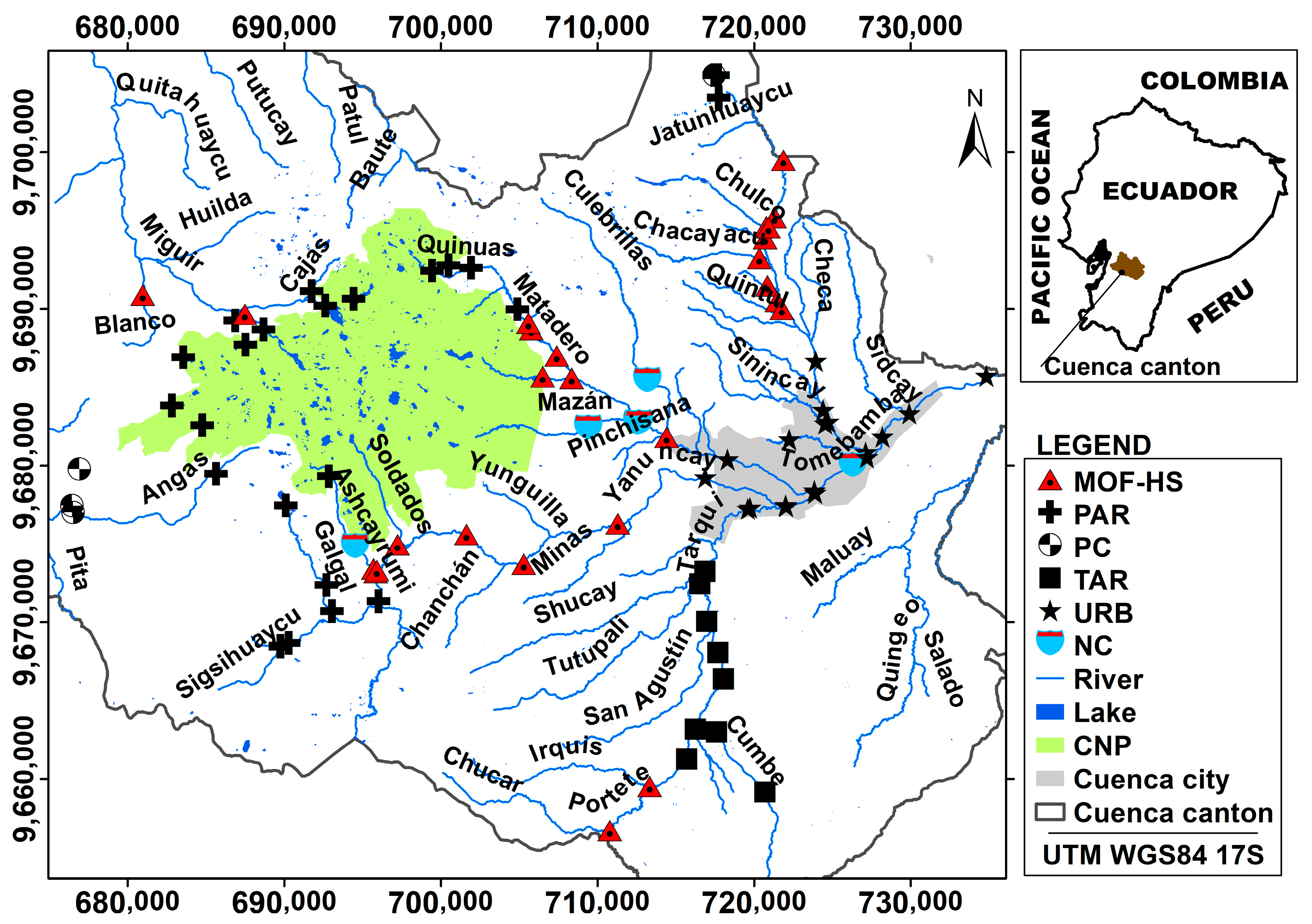
Therefore, the assessment of ecosystem health through an extensive biomonitoring network, which characterizes the anthropogenic pressure gradient and natural variability of the ecosystems, is a fundamental tool in integrated watershed management. This is particularly relevant in the seven catchments supplying water to the city of Cuenca, where natural variability is linked to elevation and topographical characteristics. The use of the community of benthic macroinvertebrates present in water bodies is a fundamental tool to recognize natural or anthropogenic changes. Macroinvertebrates are widely used in various integrated management plans applied in other parts of the world, such as the Water Framework Directive (WFD) of the European Union, due to the demonstrated sensitivity of many species to different types of pollution [
21,
22,
23]. Biological monitoring is based on the possibility of using structural and functional characteristics of the different levels of biological organization to comparatively evaluate the state of the biota, whose condition reflects the ecological state of the water body [
24].
In view of the increasing anthropogenic activities currently occurring in the Andean region and the envisaged future water resources scarcity, evaluation of river ecosystems health is necessary to ensure the sustainable management of water resources in this region. Hereafter, to contribute to the achievement of this goal, the general objective of the study was to evaluate the ecological status of high mountain fluvial networks of tropical Andean catchments in southern Ecuador. The specific objectives were as follows: (i) defining the river types in the study catchments by means of multivariate methods; (ii) validating the previously defined river typology through the analyses of representative macroinvertebrate communities; (iii) assessing the anthropogenic pressure gradient on the different river types; and (iv) identifying the key environmental variables for benthic communities present in the different study river types. The novelty of the study included the development of a river typology in a highly heterogeneous (i.e., encompassing natural as well as human-induced variability) high mountain region using hydro-geomorphological characteristics. Further, this typology was refined by considering biological characteristics of the sampled rivers; an approach that has not been used commonly. The study was executed in a strongly understudied region using an extensive sampling network for the evaluation of river ecosystem health.
4. Discussion
In the high mountains, millions of people depend on the use of surface water for human consumption [
42]. Hence, ensuring river ecosystem health is a fundamental requirement to provide the vital liquid [
43]. Establishing an extensive biomonitoring network and a regular ecosystem health assessment is an important tool of integrated watershed management. To evaluate ecosystem health, it is important to consider the anthropogenic pressure gradient but also the natural variability of the ecosystems; hence, classification of rivers is necessary. River classification is a long-established practice developed for a range of applications [
44]. In Europe, the classification of rivers was elaborated at different spatial scale, such as national [
45] or European [
46]. In most cases, the development of these typologies needs extensive datasets of hydrological regimes [
47] or geomorphology [
48]. In Ecuador, Villamarín et al. [
49] identified two different elevation bioregions, but a more detailed classification does not exist, mainly due to the complex topography and climatology [
14]. Owing to the lack of detailed hydrological datasets, a condition that importantly differs from what is observed, for instance, in northern countries, simple morphological/climatological/geological characteristics were used, which proved to be a successful approach for identifying river types; an information that may be useful for optimizing local biomonitoring costs and efforts.
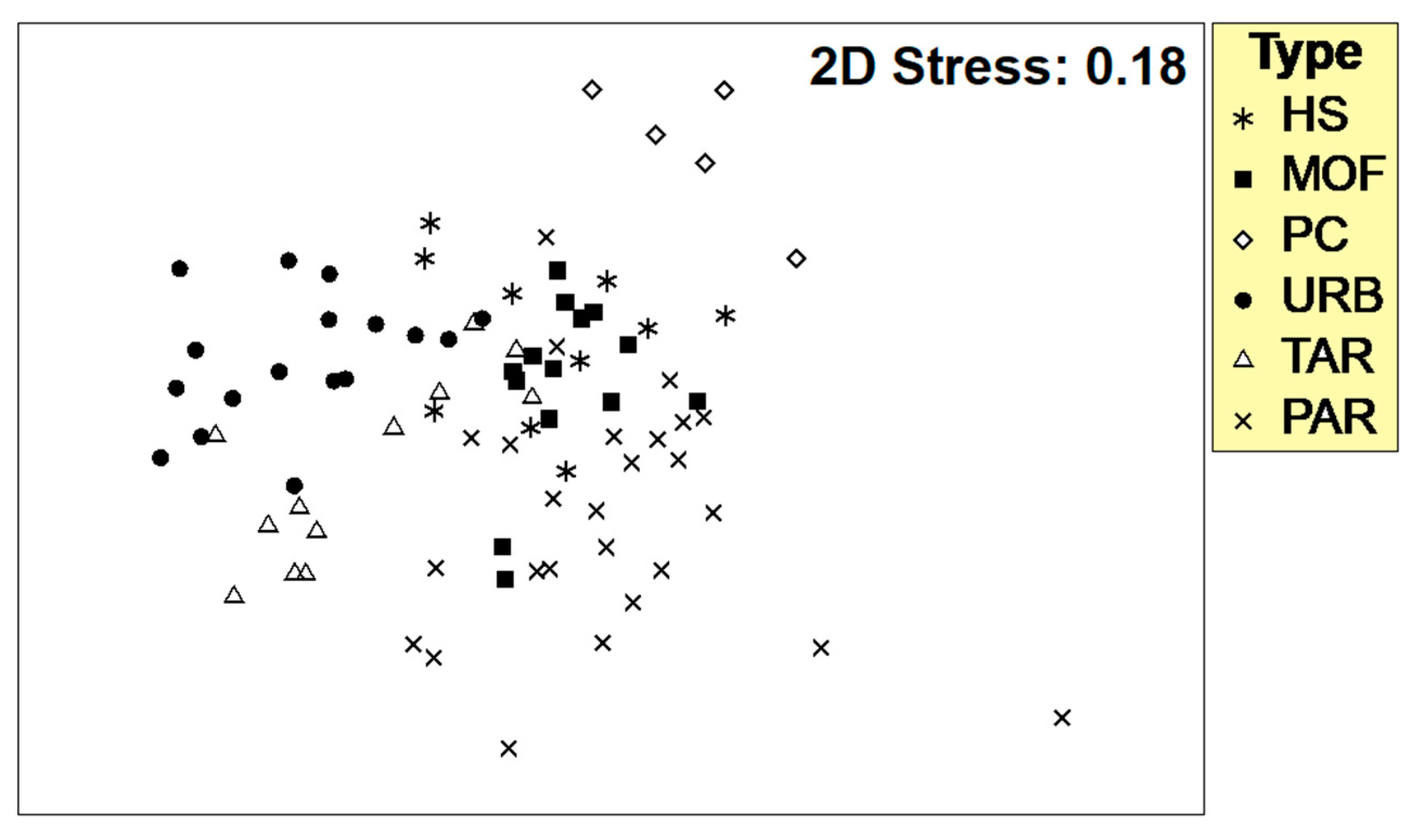
Benthic macroinvertebrates are demonstrated to be the most useful biological assessment methods for freshwater ecosystem health monitoring (i.e., Álvarez-Cabria et al. [
50], Liu et al. [
51]). Thereby, many indices have been developed using macroinvertebrates for evaluating the ecological status of lotic systems [
52]. However, macroinvertebrate communities generally are not used to define river typology. Nevertheless, this approach was successful in the present study to refine the previously developed river typology that was based exclusively on morphological/climatological/geological characteristics. This approach could be also replicated in similar studies.
In the defined river types, inorganic, organic, and microbial pollution and the state of the riparian vegetation seem to be the most important anthropogenic pressures. Turbidity and electric conductivity strongly correlate with the presence of human activities related to industrial discharges [
53]. Turbidity increases with mining activities [
54] or cattle raising [
55]. Average turbidity increased by more than 90% due to the activity of cattle in a British lowland river [
55]. In the current study, the two less impacted river types, namely, PAR and MOF-HS, had lower values of conductivity and turbidity: 70.5 µS cm
−1 and 1.5 NTU in PAR rivers and 81.1 µS cm
−1 and 3.2 NTU in the MOF-HS rivers, respectively. The catchment of the Tarqui river type is strongly dominated by cattle raising, having importantly higher values of conductivity and turbidity (average 131.4 µS cm
−1 and 8.8 NTU, respectively). Urban rivers (URB) receive important industrial discharge (average electric conductivity 127.5 µS cm
−1), and upstream deforestation resulted in the highest turbidity value (in average, 12.8 NTU) among the studied river types.
Fecal coliforms is one of the most important microbial contaminants derived, among others, from urban sewage and/or animal husbandry [
56]. Wilson and Everard [
55] reported that cattle raising doubled the level of fecal coliforms in the British lowland river. The páramo and mountain forest–humid shrub rivers had very low fecal coliforms concentrations (on average, 49 and 1318 MPN (100 mL)
−1, respectively). However, a few sampling stations, which had slightly higher concentrations compared to the rest of the stations, caused fecal coliforms to become important variables in these two river types. These stations were affected by the presence of cattle, which shows the advancement of agricultural/cattle raising frontiers toward higher altitudes. In the TAR rivers, the concentration of fecal coliforms was the most important anthropogenic pressure variable, reflecting the very active cattle raising activity occurring in this catchment. Similarly, Jayakod et al. [
57] reported that livestock operations and failing septic systems are the two main sources of fecal coliforms in the Pelahatchie watershed in Mississippi. The present study confirmed elevated values of fecal coliforms in the urban river type, which most probably originate from urban septic systems. The presence of fecal coliforms result in the increase of BOD5 as reported already several decades ago [
58]. In the present study, relatively low BOD5 values were recorded with highly elevated fecal coliforms concentrations, which is in line with the results of local studies [
59,
60]. BOD5 levels at a sampling location with slower, deeper water might be higher than the respective BOD5 levels for a similar site in aerated waters [
34]. This might be the case in the sampled rivers, which are high mountain water courses with more pronounced slopes, faster flow and more turbulence, features that facilitate stream water aeration.
Strongly reduced River Vegetation Index and lack of vegetation naturalness characterized the Tarqui and urban rivers. Cattle raising eliminated or reduced riverine vegetation in the Tarqui river, and city development had the same effect in the urban rivers. Despite several studies that argue about the upmost importance of riverine vegetation to enhance the ecological status [
61] and its impact on important ecosystem processes [
62,
63], the currently existing national regulation, which prohibits the construction of civil infrastructures within a 50 m buffer zone from the riversides, is rarely fulfilled. Sotomayor et al. [
37] stated for the whole Paute river basin, which the seven study catchments belong to, that the presence of native vegetation has the potential, along the river courses, to form buffer systems for the enhancement of riparian ecosystems and, finally, of improving the downstream water quality.
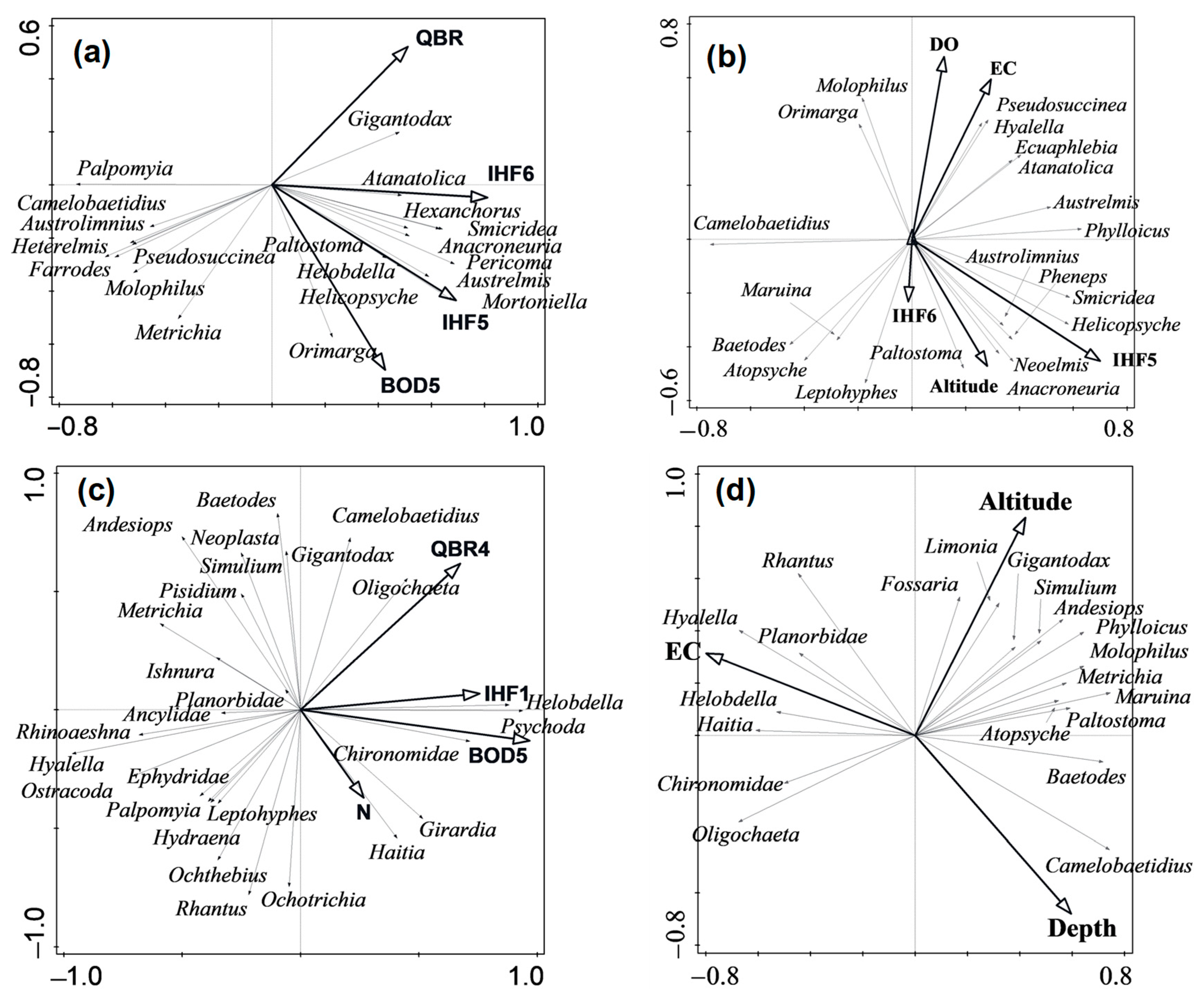
Macroinvertebrate communities responded to different environmental variables in the four river types. Species of the caddisfly (
Atanatolica,
Mortoniella, and
Helicopsyche), stoneflies (
Anacroneuria), and diptera (
Paltostoma) are present in the neotropical high mountains that normally inhabit little-impacted sites, with a good water quality and heterogeneous fluvial habitat [
64,
65]. Similarly, in the present study, these were related to high River Habitat Index in the higher elevations, páramo sites.
Orimarga, another genus from the páramo rivers, are dipterans that usually live associated with fine substrates, such as sand or silt [
66], where oxygen concentrations are lower and there may be accumulations of decomposing debris, which may explain the association of this genus with the higher BOD5 in the present study. Further, genera such as
Anacroneuria,
Helicopsyche,
Paltostoma,
Atopsyche,
Pheneps, and
Maruina are associated with heterogeneous fluvial habitats and high altitude rivers [
67,
68,
69]. In the current study, these genera were related to higher elevation sampling stations of mountain forest and humid shrub river types, where more shade and riverbed heterogeneity were present. Chironomidae and
Psychoda (Psychodidae) dipteran, present in lower elevation rivers, are usually resistant to conditions with little oxygen and high organic load [
70]. In the current study, these taxa were associated with higher BOD5 in the lower elevation Tarqui rivers. In the same rivers, the presence of snail
Haitia was linked to elevated nitrate concentrations similarly to other studies, which found this snail in eutrophic environments with excesses of nutrients [
71].
Genera of rheophilic mayflies (
Baetodes and
Camelobaetidius) that usually inhabit wide and deep river channels [
72] were detected in urban rivers where river channels were much deeper and wider than the rivers of the other types. Some genera with a preference for highlands (
Paltostoma,
Gigantodax,
Andesiops, and
Atopsyche) [
73,
74,
75] were still observed in some upper reaches of urban rivers (such as the Machángara), although with low abundances. Some of them may appear in these rivers as a consequence of dragging from the headwaters immediately after rainy periods. High conductivity, which in turn is highly correlated with other variables such as lower oxygen concentration, high fecal coliforms, and total dissolved solids, was associated with taxa such as Chironomidae, Oligochaeta, and
Haitia. This result could be due to the fact that Chironomidae and Oligochaeta were not identified further to genus or species levels, which was the case of the study of Rosa et al. [
76], who reported that the presence/absence of Chironomidae and Oligochaeta was able to detect the river’s pollution gradient. Further, Scheibler et al. [
77] reported that elevated conductivity levels and increased river discharge produced low chironomid density values in the Andes region of western–central Argentina.
5. Conclusions
Evaluations of ecological status are especially important in rivers where water is taken from surface ecosystems for drinking purposes. This study revealed that evaluations should include, in addition to physical–chemical variables, the river habitat and riparian vegetation quality, as these characteristics were important to identify anthropogenic pressures and seemed to strongly influence macroinvertebrate communities. The 90 sampling stations were grouped into five different river types, in which anthropogenic pressure gradients were inorganic (i.e., conductivity, turbidity), organic (i.e., fecal coliform), river habitat, and riparian vegetation quality. The macroinvertebrate communities were strongly influenced by different aspects of river habitat and riparian vegetation quality such as the presence of shade, riverbed heterogeneity, substrate inclusion, degree of vegetation naturalness, and Riparian Vegetation Quality Index. BOD5, representing organic pollution, was also an important variable for macroinvertebrates in two river types. Electric conductivity indicated inorganic pollution and was also correlated with biological pollution such as fecal coliforms and influenced macroinvertebrates in the mountain forest and humid shrub river types. The novelty of the study included the development of river typology on a strongly understudied region using an extensive sampling network for evaluation of river ecosystems health. Further, in addition to the physical and hydro-geomorphological characteristics, which are usually used to define river types, in the current study, analyses of macroinvertebrate communities helped to refine the river typology, which may lead to a more optimal local biomonitoring.



{kind=link}
{kind=link}
{kind=link}