
Trait-Based Research on Rotifera: The Holy Grail or Just Messy?

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
3. Results
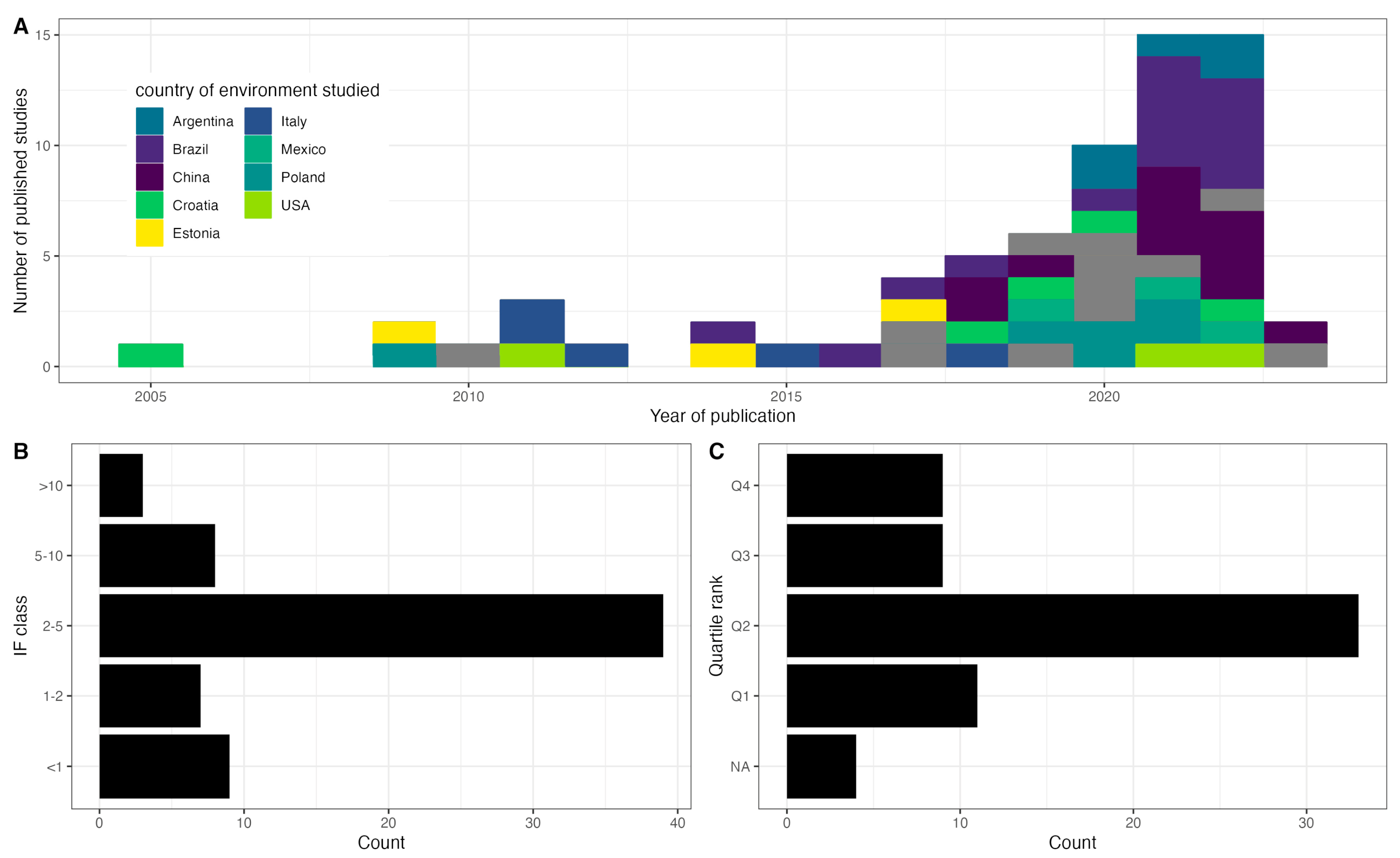
3.1. Analysis of Articles on Rotifer Functional Diversity
3.2. Rotifer Trait Matrix
3.3. Clustering of Morphological Rotifer Traits
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data availability Statement
Acknowledgments
Conflicts of Interest
References
- McGill, B.J.; Enquist, B.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Pomerleau, C.; Sastri, A.R.; Beisner, B. Evaluation of functional trait diversity for marine zooplankton communities in the Northeast subarctic Pacific Ocean. J. Plankton Res. 2015, 37, 712–726. [Google Scholar] [CrossRef]
- Dawson, S.K.; Carmona, C.P.; González-Suárez, M.; Jönsson, M.; Chichorro, F.; Mallen-Cooper, M.; Melero, Y.; Moor, H.; Simaika, J.P.; Duthie, A.B. The traits of “trait ecologists”: An analysis of the use of trait and functional trait terminology. Ecol. Evol. 2021, 11, 16434–16445. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Aleixo, I.; Norris, D.; Hemerik, L.; Barbosa, A.; Prata, E.; Costa, F.; Poorter, L. Amazonian rainforest tree mortality driven by climate and functional traits. Nat. Clim. Chang. 2019, 9, 384–388. [Google Scholar] [CrossRef]
- Mahaut, L.; Fort, F.; Violle, C.; Freschet, G.T.; Spasojevic, M. Multiple facets of diversity effects on plant productivity: Species richness, functional diversity, species identity and intraspecific competition. Funct. Ecol. 2019, 34, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Senzaki, M.; Barber, J.R.; Phillips, J.N.; Carter, N.H.; Cooper, C.B.; Ditmer, M.A.; Fristrup, K.M.; McClure, C.J.W.; Mennitt, D.J.; Tyrrell, L.P.; et al. Sensory pollutants alter bird phenology and fitness across a continent. Nature 2020, 587, 605–609. [Google Scholar] [CrossRef]
- Thakur, M.P.; Geisen, S. Trophic Regulations of the Soil Microbiome. Trends Microbiol. 2019, 27, 771–780. [Google Scholar] [CrossRef]
- Trindade, R.M.L.; Ribeiro, A.K.N.; Nabout, J.C.; Bortolini, J.C. The global scientific literature on applications and trends in the use of functional morphological groups in phytoplankton studies. Acta Limnol. Bras. 2021, 33, e12. [Google Scholar] [CrossRef]
- Reynolds, C.S. Resilience in aquatic ecosystems—hysteresis, homeostasis, and health. Aquat. Ecosyst. Heal. Manag. 2002, 5, 3–17. [Google Scholar] [CrossRef]
- Frost, T.M.; Fischer, J.M.; Klug, J.L.; Arnott, S.E.; Montz, P.K. Trajectories of Zooplankton Recovery in the Little Rock Lake Whole-Lake Acidification Experiment. Ecol. Appl. 2006, 16, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Bruel, R.; Marchetto, A.; Bernard, A.; Lami, A.; Sabatier, P.; Frossard, V.; Perga, M.-E. Seeking alternative stable states in a deep lake. Freshw. Biol. 2018, 63, 553–568. [Google Scholar] [CrossRef] [Green Version]
- Baruah, G.; Clements, C.F.; Guillaume, F.; Ozgul, A. When do shifts in trait dynamics precede population declines? Am. Nat. 2019, 193, 633–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shadrin, N.V. The alternative saline lake ecosystem states and adaptive environmental management. J. Oceanol. Limnol. 2018, 36, 2010–2017. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, M.; Liang, Z. Identification of regime shifts and their potential drivers in the shallow eutrophic Lake Yilong, Southwest China. Sustainability 2020, 9, 3704. [Google Scholar] [CrossRef]
- Pace, M.; Carpenter, S.R.; Johnson, R.; Kurtzweil, J.T. Zooplankton provide early warnings of a regime shift in a whole lake manipulation. Limnol. Oceanogr. 2013, 58, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Helenius, L.K.; Leskinen, E.; Lehtonen, H.; Nurminen, L. Spatial patterns of littoral zooplankton assemblages along a salinity gradient in a brackish sea: A functional diversity perspective. Estuar. Coast. Shelf Sci. 2017, 198, 400–412. [Google Scholar] [CrossRef]
- Barnett, A.J.; Finlay, K.; Beisner, B.E. Functional diversity of crustacean zooplankton communities: Towards a trait-based classification. Freshw. Biol. 2007, 52, 796–813. [Google Scholar] [CrossRef]
- Litchman, E.; Ohman, M.D.; Kiørboe, T. Trait-based approaches to zooplankton communities. J. Plankton Res. 2013, 35, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Obertegger, U.; Smith, H.A.; Flaim, G.; Wallace, R.L. Using the guild ratio to characterize pelagic rotifer communities. Hydrobiologia 2011, 662, 157–162. [Google Scholar] [CrossRef]
- Obertegger, U.; Flaim, G. Community assembly of rotifers based on morphological traits. Hydrobiologia 2015, 753, 31–45. [Google Scholar] [CrossRef]
- Obertegger, U.; Flaim, G. Taxonomic and functional diversity of rotifers, what do they tell us about community assembly? Hydrobiologia 2018, 823, 79–91. [Google Scholar] [CrossRef]
- Kakouei, K.; Kraemer, B.M.; Adrian, R. Variation in the predictability of lake plankton metric types. Limnol. Oceanogr. 2022, 67, 608–620. [Google Scholar] [CrossRef]
- Fintelman-Oliveira, E.; Kruk, C.; Lacerot, G.; Klippel, G.; Branco, C.W.C. Zooplankton functional groups in tropical reservoirs: Discriminating traits and environmental drivers. Hydrobiologia 2022, 850, 365–384. [Google Scholar] [CrossRef]
- Ehrlich, E.; Gaedke, U. Coupled changes in traits and biomasses cascading through a tritrophic plankton food web. Limnol. Oceanogr. 2020, 65, 2502–2514. [Google Scholar] [CrossRef]
- Gilbert, J.J. Food niches of planktonic rotifers: Diversification and implications. Limnol. Oceanogr. 2022, 67, 2218–2251. [Google Scholar] [CrossRef]
- Farley, S.S.; Dawson, A.; Goring, S.J.; Williams, J.W. Situating Ecology as a Big-Data Science: Current Advances, Challenges, and Solutions. Bioscience 2018, 68, 563–576. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.S.; Ceríaco, L.M.; Daniel, G.M.; Dellapé, P.M.; Löbl, I.; Marinov, M.; Reis, R.E.; Young, M.T.; Dubois, A.; Agarwal, I.; et al. The taxonomic impediment: A shortage of taxonomists, not the lack of technical approaches. Zool. J. Linn. Soc. 2021, 193, 381–387. [Google Scholar] [CrossRef]
- Stemberger, R.S.; Gilbert, J.J. Body Size, Food Concentration, and Population Growth in Planktonic Rotifers. Ecology 1985, 66, 1151–1159. [Google Scholar] [CrossRef]
- Wallace, R.L.; Snell, T.W.; Ricci, C.; Nogrady, T. Rotifera: Volume 1: Biology, Ecology and Systematics, 2nd ed.; Backhuys Publishers: Leiden, The Netherlands, 2006; p. 299. [Google Scholar]
- Fontaneto, D.; De Smet, W.H. Rotifera. In Handbook of Zoology: Gastrotricha, Cycloneuralia and Gnathifera, Volume 3: Gastrotricha and Gnathifera; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2015; pp. 217–300. [Google Scholar]
- Örstan, A. An extraordinary new fluvial bdelloid rotifer, Coronistomus impossibilis gen. nov., sp. nov., with adaptations for turbulent flow (Rotifera: Bdelloidea: Coronistomidae fam. nov.). Zootaxa 2021, 4966, 16–28. [Google Scholar] [CrossRef]
- Jersabek, C.D.; Leitner, M.F.; The Rotifer World Catalog. World Wide Web Electronic Publication. 2013. Available online: http://www.rotifera.hausdernatur.at/ (accessed on 1 February 2023).
- Smith, H.A.; Ejsmont-Karabin, J.; Hess, T.M.; Wallace, R.L. Paradox of planktonic rotifers: Similar structure but unique trajectories in communities of the Great Masurian Lakes (Poland). Verh. Int. Ver. Theor. Angew. Limnol. 2009, 30, 951–956. [Google Scholar] [CrossRef]
- Karabin, A. Pelagic zooplankton (Rotatoria + Crustacea) variation in the process of lake eutrophication II. Modifying effect of biotic agents. Ekol. Pol. 1985, 33, 617–644. [Google Scholar]
- Monakov, A.V. Feeding of Freshwater Zooplankton Invertebrates; Kenobi Productions: Ghent, Belgium, 2003; p. 273. [Google Scholar]
- Palazzo, F.; Bomfim, F.F.; Dias, J.D.; Simões, N.R.; Lansac-Tôha, F.A.; Bonecker, C.C. Temporal dynamics of rotifers’ feeding guilds shaped by chlorophyll-a, nitrate, and environmental heterogeneity in subtropical floodplain lakes. Int. Rev. Hydrobiol. 2021, 106, 95–105. [Google Scholar] [CrossRef]
- Gavrilko, D.E.; Shurganova, G.V.; Kudrin, I.A.; Yakimov, B.N. Identification of Freshwater Zooplankton Functional Groups Based on the Functional Traits of Species. Biol. Bull. 2021, 48, 1849–1856. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. cluster: Cluster Analysis Basics and Extensions. R Package Version 2.1.4; The R Project: Vienna, Austria, 2022. [Google Scholar]
- Hennig, C. fpc: Flexible Procedures for Clustering; R Package Version 2.2-10; The R Project: Vienna, Austria, 2023; Available online: https://CRAN.R-project.org/package=fpc (accessed on 5 March 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2-book.org (accessed on 3 February 2023).
- Garnier, S.; Ross, N.; Rudis, R.; Camargo, A.P.; Sciaini, M.; Scherer, C. Rvision—Colorblind-Friendly Color Maps for R, R package version 0.6.2; The R Project: Vienna, Austria, 2021. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2022. Available online: https://www.R-project.org/ (accessed on 3 February 2023).
- Obertegger, U.; Manca, M. Response of rotifer functional groups to changing trophic state and crustacean community. J. Limnol. 2011, 70, 231–238. [Google Scholar] [CrossRef]
- Pourriot, R. Sur la nutrition des Rotifères à partir des Algues d‘eau douce. Hydrobiologia 1957, 9, 50–59. [Google Scholar] [CrossRef]
- Pourriot, R. Food and feeding habits of Rotifera. Arch. Für Hydrobiol. 1977, 8, 243–260. [Google Scholar]
- Gu, Y.; Cai, Q.; Tan, L.; Li, B.; Ju, S.; Ye, L. Taxonomic and functional diversity of planktonic rotifers along a phosphorus gradient in the Three Gorges Reservoir, China. Fresenius Environ. Bull. 2021, 30, 1687–1695. [Google Scholar]
- Goździejewska, A.M.; Koszałka, J.; Tandyrak, R.; Grochowska, J.; Parszuto, K. Functional responses of zooplankton communities to depth, trophic status, and ion content in mine pit lakes. Hydrobiologia 2021, 848, 2699–2719. [Google Scholar] [CrossRef]
- Wang, Q.; Feng, K.; Du, X.; Yuan, J.; Liu, J.; Li, Z. Effects of land use and environmental gradients on the taxonomic and functional diversity of rotifer assemblages in lakes along the Yangtze River, China. Ecol. Indic. 2022, 142, 109199. [Google Scholar] [CrossRef]
- Tavsanoglu, U.N.; Akbulut, N.E. Seasonal dynamics of riverine zooplankton functional groups in Turkey: Kocaçay Delta as a case study. Turk. J. Fish. Aquat. Sci. 2020, 20, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Jansson, A.; Klais-Peets, R.; Grinienė, E.; Rubene, G.; Semenova, A.; Lewandowska, A.; Engström-Öst, J. Functional shifts in estuarine zooplankton in response to climate variability. Ecol. Evol. 2020, 10, 11591–11606. [Google Scholar] [CrossRef]
- O’Brien, D.A.; Gal, G.; Thackeray, S.J.; Matsuzaki, S.S.; Clements, C.F. Planktonic functional diversity changes in synchrony with lake ecosystem state. Glob. Chang. Biol. 2023, 29, 686–701. [Google Scholar] [CrossRef] [PubMed]
- Chevene, F.; Doleadec, S.; Chessel, D. A fuzzy coding approach for the analysis of long-term ecological data. Freshw. Biol. 1994, 31, 295–309. [Google Scholar] [CrossRef]
- Neury-Ormanni, J.; Vedrenne, J.; Wagner, M.; Jan, G.; Morin, S. Micro-meiofauna morphofunctional traits linked to trophic activity. Hydrobiologia 2020, 847, 2725–2736. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.; Špoljar, M.; Zhang, C.; Pronin, M. Zooplankton functional traits as a tool to assess latitudinal variation in the northern-southern temperate European regions during spring and autumn seasons. Ecol. Indic. 2020, 117, 106629. [Google Scholar] [CrossRef]
- Stamenković, O.; Stojković Piperac, M.; Čerba, D.; Milošević, D.; Ostojić, A.; Đorđević, N.B.; Simić, S.B.; Cvijanović, D.; Buzhdygan, O.Y. Taxonomic and functional aspects of diversity and composition of plankton communities in shallow lentic ecosystems along the human impact and environmental gradients. Aquat. Sci. 2022, 84, 57. [Google Scholar] [CrossRef]
- Pothoven, S.A.; Vanderploeg, H.A. Factors affecting rotifer assemblages along a nearshore to offshore transect in southeastern Lake Michigan. J. Great Lakes Res. 2022, 48, 1230–1238. [Google Scholar] [CrossRef]
- Wallace, R.L.; Snell, T.W.; Walsh, E.J.; Sarma, S.S.S.; Segers, H. Phylum Rotifera. In Thorp and Covich’s Freshwater Invertebrates: Keys to the Palaearctic Fauna; Rogers, D.C., Thorpe, J., Eds.; Academic Press: London, UK, 2019; Volume IV, pp. 219–267. [Google Scholar]
- Koste, W. Rotatoria. Die Rädertiere Mitteleuropas; 2 volumes; Gebrüder Borntraeger: Stuttgart, Germany, 1978. [Google Scholar]
- Koste, W.; Shiel, R.J. Rotifera from Australian Inland Waters. II. Epiphanidae and Brachionidae (Rotifera: Monogononta). Invertebr. Taxon. 1987, 7, 949–1021. [Google Scholar] [CrossRef]
- Ricci, C.; Melone, G.; Walsh, E.J. A carnivorous bdelloid rotifer, Abrochtha carnivora n.sp. Invertebr. Biol. 2001, 120, 136–141. [Google Scholar] [CrossRef]
- Devetter, M. Soil rotifers (Rotifera) of the Kokořínsko protected landscape area. Biologia 2007, 62, 222–224. [Google Scholar] [CrossRef]
- Melone, G.; Ricci, C. Rotatory apparatus in Bdelloids. Hydrobiologia 1995, 313, 91–98. [Google Scholar] [CrossRef]
- Hochberg, R.; Yang, H.; Hochberg, A.; Walsh, E.J.; Wallace, R.L. When heads are not homologous: The coronae of larval and adult collothecid rotifers (Rotifera: Monogononta: Collothecaceae). Hydrobiologia 2019, 844, 191–207. [Google Scholar] [CrossRef]
- Naeem, S.; Wright, J.P. Disentangling biodiversity effects on ecosystem functioning: Deriving solutions to a seemingly insurmountable problem. Ecol. Lett. 2003, 6, 567–579. [Google Scholar] [CrossRef] [Green Version]
- Stemberger, R.S.; Gilbert, J.J. Defenses of planktonic rotifers against predators. In Predation: Direct and Indirect Impacts on Aquatic Communities; Kerfoot, W.C., Sih, A., Eds.; University Press of New England: Hanover, NH, USA, 1987; pp. 227–239. [Google Scholar]
- Roche, K. Post-encounter vulnerability of some rotifer prey types to predation by the copepod Acanthocyclops robustus. Hydrobiologia 1987, 147, 229–233. [Google Scholar] [CrossRef]
- Gilbert, J.J.; McPeek, M.A. Maternal age and spine development in a rotifer: Ecological implications and evolution. Ecology 2013, 94, 2166–2172. [Google Scholar] [CrossRef] [Green Version]
- Salt, G.W. The components of feeding behavior in rotifers. Hydrobiologia 1987, 147, 271–281. [Google Scholar] [CrossRef]
- Kutikova, L.A. Parallelism in the evolution of rotifers. Hydrobiologia 1983, 104, 3–7. [Google Scholar] [CrossRef]
- Walz, N.; Schoell, K.; Wallace, R.L.; Gilbert, J.J.; Clement, P.; Rougier, C. Walter Koste (1912–2007) In memoriam. Hydrobiologia 2011, 662, 5–9. [Google Scholar] [CrossRef]
- Hochberg, R.; Wallace, R.L.; Walsh, E. Soft Bodies, Hard Jaws: An Introduction to the Symposium, with Rotifers as Models of Jaw Diversity. Integr. Comp. Biol. 2019, 55, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Bērzinš, B.; Pejler, B. Rotifer occurrence in relation to pH. Hydrobiologia 1987, 147, 107–116. [Google Scholar] [CrossRef]
- Sládecĕk, V. Rotifers as indicators of water quality. Hydrobiologia 1983, 100, 169–201. [Google Scholar] [CrossRef]
- Segers, H. Contribution to a revision of Floscularia Cuvier, 1798 (Rotifera: Monogononta): Notes on some Neotropical taxa. Hydrobiologia 1997, 354, 165–175. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Amarasekare, P.; Araújo, M.S.; Bürger, R.; Levine, J.M.; Novak, M.; Rudolf, V.H.; Schreiber, S.J.; Urban, M.C.; Vasseur, D.A. Why intraspecific trait variation matters in community ecology. Trends Ecol. Evol. 2011, 26, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Ricci, C. Culturing of some bdelloid rotifers. Hydrobiologia 1984, 112, 45–51. [Google Scholar] [CrossRef]
- Padisák, J.; Crossetti, L.O.; Naselli-Flores, L. Use and misuse in the application of the phytoplankton functional classification: A critical review with updates. Hydrobiologia 2009, 621, 1–19. [Google Scholar] [CrossRef]
- Zhang, M.; Lv, X.; Dong, J.; Gao, Y.; Zhang, J.; Li, M.; Gao, X.; Li, X. Multiple habitat templates for phytoplankton indicators within the functional group system. Hydrobiologia 2023, 850, 5–19. [Google Scholar] [CrossRef]
- Chaparro, G.; O'Farrell, I.; Hein, T. Multi-scale analysis of functional plankton diversity in floodplain wetlands: Effects of river regulation. Sci. Total Environ. 2019, 667, 338–347. [Google Scholar] [CrossRef]
- Litchman, E.; Edwards, K.F.; Boyd, P.W. Toward trait-based food webs: Universal traits and trait matching in planktonic predator–prey and host–parasite relationships. Limnol. Oceanogr. 2021, 66, 3857–3872. [Google Scholar] [CrossRef]
- Colina, M.; Calliari, D.; Carballo, C.; Kruk, C. A trait-based approach to summarize zooplankton–phytoplankton interactions in freshwaters. Hydrobiologia 2016, 767, 221–233. [Google Scholar] [CrossRef]
- Lansac-Tôha, F.M.; Heino, J.; Bini, L.M.; Peláez, O.; Baumgartner, M.T.; Quirino, B.A.; Pineda, A.; Meira, B.R.; Florêncio, F.M.; Oliveira, F.R.; et al. Cross-Taxon Congruence of Taxonomic and Functional Beta-Diversity Facets Across Spatial and Temporal Scales. Front. Environ. Sci. 2022, 10, 903074. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J. The usefulness of zooplankton as lake ecosystem indicators: Rotifer trophic state index. Pol. J. Ecol. 2012, 60, 339–350. [Google Scholar]
- Kremer, C.T.; Williams, A.K.; Finiguerra, M.; Fong, A.A.; Kellerman, A.; Paver, S.F.; Tolar, B.B.; Toscano, B.J. Realizing the potential of trait-based aquatic ecology: New tools and collaborative approaches. Limnol. Oceanogr. 2017, 62, 253–271. [Google Scholar] [CrossRef]
- Chust, G.; Vogt, M.; Benedetti, F.; Nakov, T.; Villéger, S.; Aubert, A.; Vallina, S.M.; Righetti, D.; Not, F.; Biard, T. Mare Incognitum: A glimpse into future plankton diversity and ecology research. Front. Mar. Sci. 2017, 4, 68. [Google Scholar]
- Winder, M.; Schindler, D.E. Climate change uncouples trophic interactions in an aquatic ecosystem. Ecology 2004, 85, 2100–2106. [Google Scholar] [CrossRef]
- Weithoff, G. On the ecology of the rotifer Cephalodella hoodi from an extremely acidic lake. Freshw. Biol. 2005, 50, 1464–1473. [Google Scholar] [CrossRef]
- Gonzalez, M.J.; Frost, T.M. Comparisons of laboratory bioassays and a whole-lake experiment: Rotifer responses to experimental acidification. Ecol. Appl. 1994, 4, 69–80. [Google Scholar] [CrossRef]
- Halse, S.A.; Shiel, R.J.; Williams, W.D. Aquatic invertebrates of Lake Gregory, northwestern Australia, in relation to salinity and ionic composition. Hydrobiologia 1998, 381, 15–29. [Google Scholar] [CrossRef]
- Tiffany, M.A.; Swan, B.K.; Watts, J.M.; Hurlbert, S.H. Metazooplankton dynamics in the Salton Sea, California, 1997–1999. Hydrobiologia 2002, 473, 103–120. [Google Scholar] [CrossRef]
- Snell, T.W.; Johnston, R.K.; Matthews, A.B.; Park, N.; Berry, S.; Brashear, J. Using Proales similis (Rotifera) for toxicity assessment in marine waters. Environ. Toxicol. 2019, 34, 634–644. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Won, E.J.; Hwang, U.K.; Kim, I.C.; Yim, J.H.; Lee, J.S. Triclosan (TCS) and Triclocarban (TCC) cause lifespan reduction and reproductive impairment through oxidative stress-mediated expression of the defensome in the monogonont rotifer (Brachionus koreanus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 185, 131–137. [Google Scholar] [CrossRef]
- Rivera-Dávila, O.L.; Sánchez-Martínez, G.; Rico-Martínez, R. Toxicity tests, bioaccumulation and residuality of pyrethroid insecticides commonly used to control conifer bark beetles in Mexico. Ecotoxicology 2022, 31, 782–796. [Google Scholar] [CrossRef]
- Su, G.; Xu, J.; Akasaka, M.; Molinos, J.G.; Shin-ichiro, S.M. Human impacts on functional and taxonomic homogenization of plateau fish assemblages in Yunnan, China. Glob. Ecol. Conserv. 2015, 4, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Sui, Y.; Wang, S.; Mohsen, M.; Zhang, L.; Shen, M.; Liu, Z.; Nguyen, H.; Zhang, S.; Li, K.; Lv, L.; et al. The combined effect of plastic particles size and concentration on rotifers’ (Brachionus plicatilis) performance. J. Ocean. Univ. China 2022, 21, 509–519. [Google Scholar] [CrossRef]
- Arnold, W.R.; Diamond, R.L.; Smith, D.S. The effects of salinity, pH, and dissolved organic matter on acute copper toxicity to the rotifer, Brachionus plicatilis (“L” strain). Arch. Environ. Contam. Toxicol. 2010, 59, 225–234. [Google Scholar] [CrossRef]
- Kim, J.; Haque, N.; Lee, S.; Lee, D.-H.; Rhee, J.-S. Exposure to Environmentally Relevant Concentrations of Polystyrene Microplastics Increases Hexavalent Chromium Toxicity in Aquatic Animals. Toxics 2022, 10, 563. [Google Scholar] [CrossRef]
- Ríos-Arana, J.V.; Gardea-Torresday, J.L.; Webb, R.; Walsh, E.J. Heat Shock Protein 60 (HSP60) Response of Plationus patulus (Rotifera: Monogononta) to Combined Exposures of Arsenic and Heavy Metals. Hydrobiologia 2005, 546, 577–585. [Google Scholar] [CrossRef]
- Angeler, D.G.; Goedkoop, W. Biological responses to liming in boreal lakes: An assessment using plankton, macroinvertebrate and fish communities. J. Appl. Ecol. 2010, 47, 478–486. [Google Scholar] [CrossRef]
- Arcifa, M.S.; de Souza, B.B.; de Morais-Junior, C.S.; Bruno, C.G.C. Functional groups of rotifers and an exotic species in a tropical shallow lake. Sci. Rep. 2020, 10, 14698. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.J.; Vaschetto, P.A.; Marchese, M.; Regaldo, L.; Gagneten, A.M. Benthic Macroinvertebrates and Zooplankton Communities as Ecological Indicators in Urban Wetlands of Argentina. Sustainability 2022, 14, 4045. [Google Scholar] [CrossRef]
- Arruda, G.d.A.; Diniz, L.P.; Almeida, V.L.D.S.; Neumann-Leitão, S.; de Melo Junior, M. Rotifer community structure in fish-farming systems associated with a Neotropical semiarid reservoir in north-eastern Brazil. Aquac. Res. 2017, 48, 4910–4922. [Google Scholar] [CrossRef]
- Bertani, I.; Ferrari, I.; Rossetti, G. Role of intra-community biotic interactions in structuring riverine zooplankton under low-flow, summer conditions. J. Plankton Res. 2012, 34, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Braghin, L.d.S.M.; Almeida, B.D.A.; Amaral, D.C.; Canella, T.F.; Gimenez, B.C.G.; Bonecker, C.C. Effects of dams decrease zooplankton functional β-diversity in river-associated lakes. Freshw. Biol. 2018, 63, 721–730. [Google Scholar] [CrossRef]
- Brown, P.D.; Schröder, T.; Ríos-Arana, J.V.; Rico-Martinez, R.; Silva-Briano, M.; Wallace, R.L.; Walsh, E.J. Patterns of rotifer diversity in the Chihuahuan desert. Diversity 2020, 12, 393. [Google Scholar] [CrossRef]
- Chen, Q.; Sun, X.; Yu, H.X. Relationship between environmental factors and metazooplankton community structure from Zhalong national nature reserve in Heilong Jiang province, NorthEastern China. Appl. Ecol. Environ. Res. 2021, 19, 2843–2858. [Google Scholar] [CrossRef]
- Chen, X.; Huang, K.; Zhang, J.; Huang, Q.; Liu, P.; Han, B. Spring community structure of pelagic rotifers in (sub)tropical reservoirs of China: Taxonomic and functional trait-based analysis. J. Lake Sci. 2021, 33, 1490–1499. [Google Scholar] [CrossRef]
- Coelho, P.N.; Henry, R. Is the littoral zone taxonomically and functionally more diverse? Investigating the rotifer community of a tropical shallow lake. Limnology 2022, 23, 429–440. [Google Scholar] [CrossRef]
- de Paggi, S.B.J.; Devercelli, M.; Molina, F.R. Zooplankton and their driving factors in a large subtropical river during low water periods. Fundam. Appl. Limnol. Schweiz. Verl. 2014, 184, 125–139. [Google Scholar] [CrossRef]
- Deosti, S.; de Fátima Bomfim, F.; Lansac-Tôha, F.M.; Quirino, B.A.; Bonecker, C.C.; Lansac-Tôha, F.A. Zooplankton taxonomic and functional structure is determined by macrophytes and fish predation in a Neotropical river. Hydrobiologia 2021, 848, 1475–1490. [Google Scholar] [CrossRef]
- Duré, G.A.V.; Simões, N.R.; Braghin, L.D.S.M.; Ribeiro, S.M.M.S. Effect of eutrophication on the functional diversity of zooplankton in shallow ponds in Northeast Brazil. J. Plankton Res. 2021, 43, 894–907. [Google Scholar] [CrossRef]
- Espinosa-Rodríguez, C.A.; Sarma, S.S.S.; Nandini, S. Zooplankton community changes in relation to different macrophyte species: Effects of Egeria densa removal. Ecohydrol. Hydrobiol. 2021, 21, 153–163. [Google Scholar] [CrossRef]
- Frau, D.; Gutierrez, M.F.; Molina, F.R.; de Mello, F.T. Drivers assessment of zooplankton grazing on phytoplankton under different scenarios of fish predation and turbidity in an in situ mesocosm experiment. Hydrobiologia 2021, 848, 485–498. [Google Scholar] [CrossRef]
- Gadelha, E.S.; Dunck, B.; SimoeS, N.R.; Paes, E.T.; Akama, A. High taxonomic turnover and functional homogenization of rotifer communities in an amazonian river. An. Acad. Bras. Ciências 2023, 94, e20201894. [Google Scholar] [CrossRef]
- Galir Balkić, A.; Ternjej, I.; Špoljar, M. Hydrology driven changes in the rotifer trophic structure and implications for food web interactions. Ecohydrology 2018, 11, e1917. [Google Scholar] [CrossRef]
- Galir Balkić, A.G. The importance of environmental differences in the structuring of rotifer functional diversity. J. Limnol. 2019, 78, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Goździejewska, A.M.; Gwoździk, M.; Kulesza, S.; Bramowicz, M.; Koszałka, J. Effects of suspended micro-and nanoscale particles on zooplankton functional diversity of drainage system reservoirs at an open-pit mine. Sci. Rep. 2019, 9, 16113. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, M.F.; Simões, N.R.; Frau, D.; Saigo, M.; Licursi, M. Responses of stream zooplankton diversity metrics to eutrophication and temporal environmental variability in agricultural catchments. Environ. Monit. Assess. 2020, 192, 1–17. [Google Scholar] [CrossRef]
- Gutierrez, M.F.; Epele, L.B.; Mayora, G.; Aquino, D.; Mora, C.; Quintana, R.; Mesa, L. Hydro-climatic changes promote shifts in zooplankton composition and diversity in wetlands of the Lower Paraná River Delta. Hydrobiologia 2022, 849, 3463–3480. [Google Scholar] [CrossRef]
- Gutierrez, M.F.; Tavşanoğlu, Ü.N.; Vidal, N.; Yu, J.; de Mello, F.T.; Çakiroglu, A.I.; He, H.; Liu, Z.; Jeppesen, E. Salinity shapes zooplankton communities and functional diversity and has complex effects on size structure in lakes. Hydrobiologia 2018, 813, 237–255. [Google Scholar] [CrossRef]
- Işkın, U.; Filiz, N.; Cao, Y.; Neif, É.M.; Öğlü, B.; Lauridsen, T.L.; Davidson, T.A.; Søndergaard, M.; Tavşanoğlu, Ü.N.; Beklioğlu, M. Impact of nutrients, temperatures, and a heat wave on zooplankton community structure: An experimental approach. Water 2020, 12, 3416. [Google Scholar] [CrossRef]
- Josué, I.I.; Sodré, E.O.; Setubal, R.B.; Cardoso, S.J.; Roland, F.; Figueiredo-Barros, M.P.; Bozelli, R.L. Zooplankton functional diversity as an indicator of a long-term aquatic restoration in an Amazonian lake. Restor. Ecol. 2021, 29, e13365. [Google Scholar] [CrossRef]
- Krztoń, W.; Kosiba, J.; Wilk-Woźniak, E. Features that matter: Studying how phytoplankton drives zooplankton community functional traits. Hydrobiologia 2022, 849, 2647–2662. [Google Scholar] [CrossRef]
- Krztoń, W.; Kosiba, J. Variations in zooplankton functional groups density in freshwater ecosystems exposed to cyanobacterial blooms. Sci. Total Environ. 2020, 730, 139044. [Google Scholar] [CrossRef]
- Labuce, A.; Ikauniece, A.; Jurgensone, I.; Aigars, J. Environmental Impacts on Zooplankton Functional Diversity in Brackish Semi-Enclosed Gulf. Water 2021, 13, 1881. [Google Scholar] [CrossRef]
- Lokko, K.; Virro, T. The structure of psammic rotifer communities in two boreal lakes with different trophic conditions: Lake Võrtsjärv and Lake Saadjärv (Estonia). Oceanol. Hydrobiol. Stud. 2014, 43, 49–55. [Google Scholar] [CrossRef]
- Lokko, K.; Virro, T.; Kotta, J. Seasonal variability in the structure and functional diversity of psammic rotifer communities: Role of environmental parameters. Hydrobiologia 2017, 96, 287–307. [Google Scholar] [CrossRef]
- Moi, D.A.; Romero, G.Q.; Jeppesen, E.; Kratina, P.; Alves, D.C.; Antiqueira, P.A.; de Mello, F.T.; Figueiredo, B.R.; Bonecker, C.C.; Pires, A.P. Regime shifts in a shallow lake over 12 years: Consequences for taxonomic and functional diversities, and ecosystem multifunctionality. J. Anim. Ecol. 2022, 91, 551–565. [Google Scholar] [CrossRef]
- Moreira, F.W.A.; Leite, M.G.P.; Fujaco, M.A.G.; Mendonça, F.P.C.; Campos, L.P.; Eskinazi-Sant’Anna, E.M. Assessing the impacts of mining activities on zooplankton functional diversity. Acta Limnol. Bras. 2016, 28, e7. [Google Scholar] [CrossRef]
- Mwagona, P.C.; Chengxue, M.; Hongxian, Y. Seasonal dynamics of Zooplankton functional groups in relation to environmental variables in Xiquanyan Reservoir, Northeast China. Ann. De Limnol.-Int. J. Limnol. 2018, 54, 33. [Google Scholar] [CrossRef]
- Oh, H.-J.; Jeong, H.-G.; Nam, G.-S.; Oda, Y.; Dai, W.; Lee, E.-H.; Kong, D.; Hwang, S.-J.; Chang, K.-H. Comparison of taxon-based and trophi-based response patterns of rotifer community to water quality: Applicability of the rotifer functional group as an indicator of water quality. Anim. Cells Syst. 2017, 21, 133–140. [Google Scholar] [CrossRef]
- Pinel-Alloul, B.; Chemli, A.; Taranu, Z.E.; Bertolo, A. Using the Diversity, Taxonomic and Functional Attributes of a Zooplankton Community to Determine Lake Environmental Typology in the Natural Southern Boreal Lakes (Québec, Canada). Water 2022, 14, 578. [Google Scholar] [CrossRef]
- Porcel, S.; Chaparro, G.; Marinone, M.C.; Saad, J.F.; Lancelotti, J.; Izaguirre, I. The role of environmental, geographical, morphometric and spatial variables on plankton communities in lakes of the arid Patagonian plateaus. J. Plankton Res. 2020, 42, 173–187. [Google Scholar] [CrossRef]
- Qian, F.-P.; Wen, X.-L.; Xi, Y.-L. Seasonal variations of rotifer communities in three climatic zones: Effects of environmental parameters on changes of functional groups. Limnology 2022, 23, 165–180. [Google Scholar] [CrossRef]
- Ribeiro, B.I.O.; Braghin, L.D.S.M.; Lansac-Tôha, F.M.; Bomfim, F.F.; Almeida, B.A.; Bonecker, C.C.; Lansac-Tôha, F.A. Environmental heterogeneity increases dissimilarity in zooplankton functional traits along a large Neotropical river. Hydrobiologia 2022, 849, 3135–3147. [Google Scholar] [CrossRef]
- Ríos-Arana, J.V.; del Carmen Agüero-Reyes, L.; Wallace, R.L.; Walsh, E.J. Limnological characteristics and rotifer community composition of Northern Mexico Chihuahuan Desert Springs. J. Arid Environ. 2019, 160, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Špoljar, M.; Habdija, I.; Primc-Habdija, B.; Sipos, L. Impact of environmental variables and food availability on rotifer assemblage in the karstic barrage Lake Visovac (Krka River, Croatia). Int. Rev. Hydrobiol. 2005, 90, 555–579. [Google Scholar] [CrossRef]
- Sun, X.; Chai, F.Y.; Mwagona, P.C.; Shabani, I.E.; Hou, W.J.; Li, X.Y.; Ma, C.X.; Pan, H.F.; Li, S.; Yu, H.X. Seasonal variations of zooplankton functional groups and relationship with environmental factors in a eutrophic reservoir from cold region. Appl. Ecol. Environ. Res. 2019, 17, 7727–7740. [Google Scholar] [CrossRef]
- Tasevska, O.; Špoljar, M.; Gušeska, D.; Kostoski, G.; Patcheva, S.; Sarafiloska, E.V. Zooplankton in ancient and oligotrophic Lake Ohrid (Europe) in association with environmental variables. Croat. J. Fish. 2017, 75, 95–103. [Google Scholar] [CrossRef]
- Tian, X.; Yuan, Y.; Zou, Y.; Qin, L.; Zhu, X.; Zhu, Y.; Zhao, Y.; Jiang, M.; Jiang, M. Cyanobacterial Blooms Increase Functional Diversity of Metazooplankton in a Shallow Eutrophic Lake. Water 2023, 15, 953. [Google Scholar] [CrossRef]
- Virro, T.; Haberman, J.; Haldna, M.; Blank, K. Diversity and structure of the winter rotifer assemblage in a shallow eutrophic northern temperate Lake Võrtsjärv. Aquat. Ecol. 2009, 43, 755–764. [Google Scholar] [CrossRef]
- Wang, H.; Huo, T.; Du, X.; Wang, L.; Song, D.; Huang, X.; Zhao, C. Zooplankton community and its environmental driving factors in Ulungur Lake, China. J. Freshw. Ecol. 2022, 37, 387–403. [Google Scholar] [CrossRef]
- Wu, L.; Ji, L.; Chen, X.; Ni, J.; Zhang, Y.; Geng, M. Distribution of Zooplankton Functional Groups in the Chaohu Lake Basin, China. Water 2022, 14, 2106. [Google Scholar] [CrossRef]
- Zhao, K.; Wang, L.; You, Q.; Zhang, J.; Pang, W.; Wang, Q. Impact of cyanobacterial bloom intensity on plankton ecosystem functioning measured by eukaryotic phytoplankton and zooplankton indicators. Ecol. Indic. 2022, 140, 109028. [Google Scholar] [CrossRef]
- Berzins, B. Some rotifers from Cambodia. Hydrobiologia 1973, 41, 453–459. [Google Scholar] [CrossRef]
- Bevington, D.J.; White, C.; Wallace, R.L. Predatory behavior of Cupelopagis vorax (Rotifera; Collothecacea; Atrochidae) on protozoan prey. Hydrobiologia 1993, 313, 213–217. [Google Scholar] [CrossRef]
- Castro, J.; Castro, G.; Castro, A.E.; Ramírez, J. Laboratory controlled production of Lepadella sp. (Bory of St. Vincent, 1826), using three different microalgae and their combination, enriched with yeast. Int. J. Fish. Aquat. Stud. 2018, 6, 372–376. [Google Scholar]
- De Smet, W.H. Rotifera. Volume 4: The Proalidae (Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; No. 9; SPB Academic Publishing: Amsterdam, The Netherlands, 1996. [Google Scholar]
- De Smet, W.H. Rotifera. Volume 5: The Dicranophoridae (Monogononta) and The Ituridae (Monogononta) (GUIDES # 12); SPB Academic Publishing: Amsterdam, The Netherlands, 1997. [Google Scholar]
- Harring, H.K.; Myers, F.J. The rotifera fauna of Wisconsin. IV. Dicranophorinae. Trans. Wis. Acad. Sci. Arts Lett. 1928, 23, 667–808. [Google Scholar]
- De Smet, W.H. Paradicranophorus sinus sp. nov. (Dicranophoridae, Monogononta) a new rotifer from Belgium, with remarks on some other species of the genus Paradicranophorus Wiszniewski, 1929 and description of Donneria gen. nov. Belg. J. Zool. 2003, 133, 181–188. [Google Scholar]
- De Smet, W.H. Asciaporrectidae, a new family of Rotifera (Monogononta: Ploima) with description of Asciaporrecta arcellicola gen. et sp. nov. and A. difflugicola gen. et sp. nov. inhabiting shells of testate amoebae (Protozoa). Zootaxa 2006, 1339, 31–49. [Google Scholar]
- De Smet, W.H. Cotylegaleatidae, a new family of Ploima (Rotifera: Monogononta), for Cotylegaleata perplexa gen. et sp. nov., from freshwater benthos of Belgium. Zootaxa 2007, 1425, 35–43. [Google Scholar] [CrossRef]
- De Smet, W.H. Rotifera from the Mediterranean Sea, with description of ten new species. Zootaxa 2015, 4028, 151–196. [Google Scholar] [CrossRef] [Green Version]
- De Smet, W.H.; Bozkurt, A. Cotylegaleata iskenderunensis n. sp., the second known species of the rotifer family Cotylegaleatidae (Monogononta: Ploima). Zootaxa 2016, 4193, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Dhanapathi, M.V.S.S.S. New species of rotifer from India belonging to the family Brachionidae. Zool. J. Linn. Soc. 1978, 62, 305–308. [Google Scholar] [CrossRef]
- Donner, J. Ordnung Bdelloidea (Rotatoria, Rädertiere); Akademie-Verlag: Berlin, Germany, 1965. [Google Scholar]
- Edmondson, W.T. Rotifera. In Freshwater Biology, 2nd ed.; Habrotrochidae-Type—Corona of Small Trochi, Gut a Syncytial Mass of Food Vacuoles; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1959; p. 484. [Google Scholar]
- Espinosa-Rodríguez, C.A.; Sarma, S.S.S.; Nandini, S.; Wallacer, R.L. Substratum selection and feeding responses influence the demography of the sessile rotifer Cupelopagis vorax (Collothecacea: Atrochidae). Int. Rev. Hydrobiol. 2021, 106, 18–28. [Google Scholar] [CrossRef]
- Flesch, F.; Berger, P.; Robles-Vargas, D.; Santos-Medrano, G.E.; Rico-Martínez, R. Characterization and determination of the toxicological risk of Biochar using invertebrate toxicity tests in the State of Aguascalientes, México. Appl. Sci. 2019, 9, 1706. [Google Scholar] [CrossRef] [Green Version]
- Ganf, G.G.; Shiel, R.J.; Merrick, C.J. Parasitism: The possible cause of the collapse of a Volvox population in Mount Bold Reservoir, South Australia. Aust. J. Mar. Freshw. Res. 1983, 34, 489–494. [Google Scholar] [CrossRef]
- Gilbert, J.J. Selective cannibalism in the rotifer Asplanchna sieboldii: Contact recognition of morphotype and clone. Proc. Natl. Acad. Sci. USA 1976, 73, 3233–3237. [Google Scholar] [CrossRef] [Green Version]
- Edmondson, W.T. Freshwater Biology, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1959; p. 482. [Google Scholar]
- Gulati, R.D.; Ejsmont-Karabin, J.; Rooth, J.; Siewertsen, K. A laboratory study of feeding and assimilation in Euchlanis dilatata lucksiana. Hydrobiologia 1987, 186–187, 346–354. [Google Scholar] [CrossRef]
- King, C.E. Food, age, and the dynamics of a laboratory population of rotifers. Ecology 1967, 48, 111–128. [Google Scholar] [CrossRef]
- Harring, H.K.; Myers, F.J. The rotifer fauna of Wisconsin. Trans. Wis. Acad. Sci. Arts Lett. 1922, 20, 553–662, (Habitat: p. 610). [Google Scholar]
- Harring, H.K.; Myers, F.J. The rotifer fauna of Wisconsin II—A revision of the notommatid rotifers exclusive of Dicranophorinae. Trans. Wis. Acad. Sci. Arts Lett. 1924, 21, 415–549. [Google Scholar]
- Hochberg, R.; O’Brien, S.; Puleo, A. Behavior, metamorphosis, and muscular organization of the predatory rotifer Acyclus inquietus (Rotifera, Monogononta). Invertebr. Biol. 2010, 129, 210–219. [Google Scholar] [CrossRef]
- Jennings, H.S. A list of the Rotatoria of the Great Lakes and of some of the inland lakes of Michigan. Bull. Mich. Fish Commisson 1894, 3, 1–34. [Google Scholar]
- Jersabek, C.D. Encentrum (Parencentrum) walterkostei n. sp., a new dicranophorid rotifer (Rotatoria: Monogononta) from the high alpine zone of the Central Alps (Austria). Hydrobiologia 1994, 281, 51–66. [Google Scholar] [CrossRef]
- Koste, W.; Shiel, R.J. Rotifera from Australian inland waters. VII. Notommatidae (Rotifera: Monogononta). Trans. R. Soc. South Aust. 1991, 115, 111–159. [Google Scholar]
- Harring, H.K. A list of the Rotatoria of Washington and vicinity, with descroptions of a new genus and ten new species. Proc. United States Natl. Nuseum 1913, 46, 387–405. [Google Scholar] [CrossRef] [Green Version]
- Leasi, F.; Neves, R.C.; Worsaae, K.; Sørensen, M.V. Musculature of Seison nebaliae Grube, 1861 and Paraseison annulatus (Claus, 1876) revealed with CLSM: A comparative study of the gnathiferan key taxon Seisonacea (Rotifera). Zoomorphology 2012, 131, 185–195. [Google Scholar] [CrossRef]
- Luo, Y.; Segers, H. On Pulchritia new genus, with a reappraisal of the genera of Trichotriidae (Rotifera, Monogononta). Zookeys 2013, 342, 1–12. [Google Scholar] [CrossRef]
- Madhu, K.; Madhu, R.; Mohandas, M.P.; Vijayan, M.T. Isolation, identification and culture of the marine rotifer Colurella adriatica Ehrenberg, 1831 (Family: Lepadellidae) from Andaman & Nicobar Islands: A promising live feed for larval rearing of high value shellfishes and finfishes. J. Mar. Biol. Assoc. India 2016, 58, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Nandini, S.; Sarma, S.S.S. Life history characteristics of Asplanchnopus multiceps (Rotifera) fed rotifer and cladoceran Prey. Hydrobiologia 2005, 546, 491–501. [Google Scholar] [CrossRef]
- Nogrady, T.; Segers, H. Rotifera. Volume 6: Asplanchnidae, Gastropodidae, Lindiidae, Microcodidae, Synchaetidae, Trochosphaeridae and Filinia. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; No. 18; SPB Academic Publishers: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Nandini, S.; Sarma, S.S.S. Population growth, demography and competition studies on Dipleuchlanis propatula (Gosse, 1886) (Rotifera: Euchlanidae). Aquat. Ecol. 2021, 55, 1305–1316. [Google Scholar] [CrossRef]
- Nogrady, T.; Pourriot, R. Rotifera. Volume 3: The Notommatidae and The Scaridiidae. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; No. 8.; SPB Academic Publishers: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Ooms-Wilms, A.L. Are bacteria an important food source for rotifers in eutrophic lakes? J. Plankton Res. 1997, 19, 1125–1141. [Google Scholar] [CrossRef]
- Pejler, B.; Berzins, B. On the ecology of Cephalodella. Hydrobiologia 1993, 259, 125–128. [Google Scholar] [CrossRef]
- Pourriot, R. Rechérches sur l’ecologie des Rotiféres. Vie Et Milieu 1965, 21, 1–224. [Google Scholar]
- Ricci, C.; Melone, G. Key to the identification of the genera of bdelloid rotifers. Hydrobiologia 2000, 418, 73–80. [Google Scholar] [CrossRef]
- Ricci, C.; Melone, G. The Philodinavidae (Rotifera Bdelloidea): A special family. Hydrobiologia 1998, 385, 77–85. [Google Scholar] [CrossRef]
- Sharma, S.S.S. Zooplankton diversity: Freshwater planktonic and semi-planktonic Rotifera. Water Qual. Assess. Biomonit. Zooplankton Divers. 2001, 2001, 190–215. [Google Scholar]
- Sarma, S.S.S.; Lara Resendiz, R.A.R.A.; Nandini, S. Morphometric and demographic responses of brachionid prey (Brachionus calyciflorus Pallas and Plationus macracanthus (Daday) in the presence of different densities of the predator Asplanchna brightwellii (Rotifera: Asplanchnidae. Hydrobiologia 2011, 662, 179–187. [Google Scholar] [CrossRef]
- Schöll, K.; Dózsa-Farkas, K. A new record of an interesting parasitic rotifer Balatro calvus CLAPARÉDE, 1867 in Hungary. Opusc. Zool. 2006, 35, 85–87. [Google Scholar]
- Segers, H.; Shiel, R.J. Diversity of cryptic Metazoa in Australian freshwaters: A new genus and two new species of sessile rotifer (Rotifera, Monogononta, Gnesiotrocha, Flosculariidae. Zootaxa 2008, 1750, 19–31. [Google Scholar] [CrossRef]
- Van Damme, K.; Segers, H. Anomopus telphusae Piovanelli, 1903, an epizoic bdelloid (Rotifera: Bdelloidea) on the Socotran endemic crab Socotrapotamon socotrensis (Hilgendorf, 1883). Fauna Saudi Arab. 2004, 20, 169–175. [Google Scholar]
- Wallace, R.L. Coloniality in the phylum Rotifera. Hydrobiologia 1987, 147, 141–155. [Google Scholar] [CrossRef]
- Wallace, R.L.; Starkweather, P.L. Clearance rates of sessile rotifers: In vitro determinations. Hydrobiologia 1985, 121, 139–144. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obertegger, U.; Wallace, R.L. Trait-Based Research on Rotifera: The Holy Grail or Just Messy? Water 2023, 15, 1459. https://doi.org/10.3390/w15081459
Obertegger U, Wallace RL. Trait-Based Research on Rotifera: The Holy Grail or Just Messy? Water. 2023; 15(8):1459. https://doi.org/10.3390/w15081459
Chicago/Turabian StyleObertegger, Ulrike, and Robert Lee Wallace. 2023. "Trait-Based Research on Rotifera: The Holy Grail or Just Messy?" Water 15, no. 8: 1459. https://doi.org/10.3390/w15081459