

Interpreting Seasonal Patterns and Long-Term Changes of Zooplankton in a Deep Subalpine Lake Using Stable Isotope Analysis




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (1)
- to examine the seasonal and inter-annual variability of the isotopic signals of δ13C and δ15N in the different zooplankton groups and taxa.
- (2)
- to investigate the long-term trends of zooplankton isotopic signals of δ13C and δ15N and their relationships with environmental variables.
2. Study Site
3. Materials and Methods
3.1. Zooplankton Sampling and Isotope Analyses
3.2. Statistical Analyses
4. Results
4.1. Variations in δ13C‰ and δ15N‰ in Zooplankton Taxa
4.2. δ13C‰ and Environmental Variables
4.2.1. Temperature and δ13C‰ Fractionation
4.2.2. Chlorophyll a and Nitrates
5. Discussion
5.1. δ13C and Temperature
5.2. Carbon Isotopic Signature in Different Taxa/Groups as Niche Indicators
5.3. Zooplankton (Daphnia) δ13C and Chlorophyll a: Indication of Food Quality and Availability
5.4. δ13C and δ15N Time Series and Environmental Variables
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
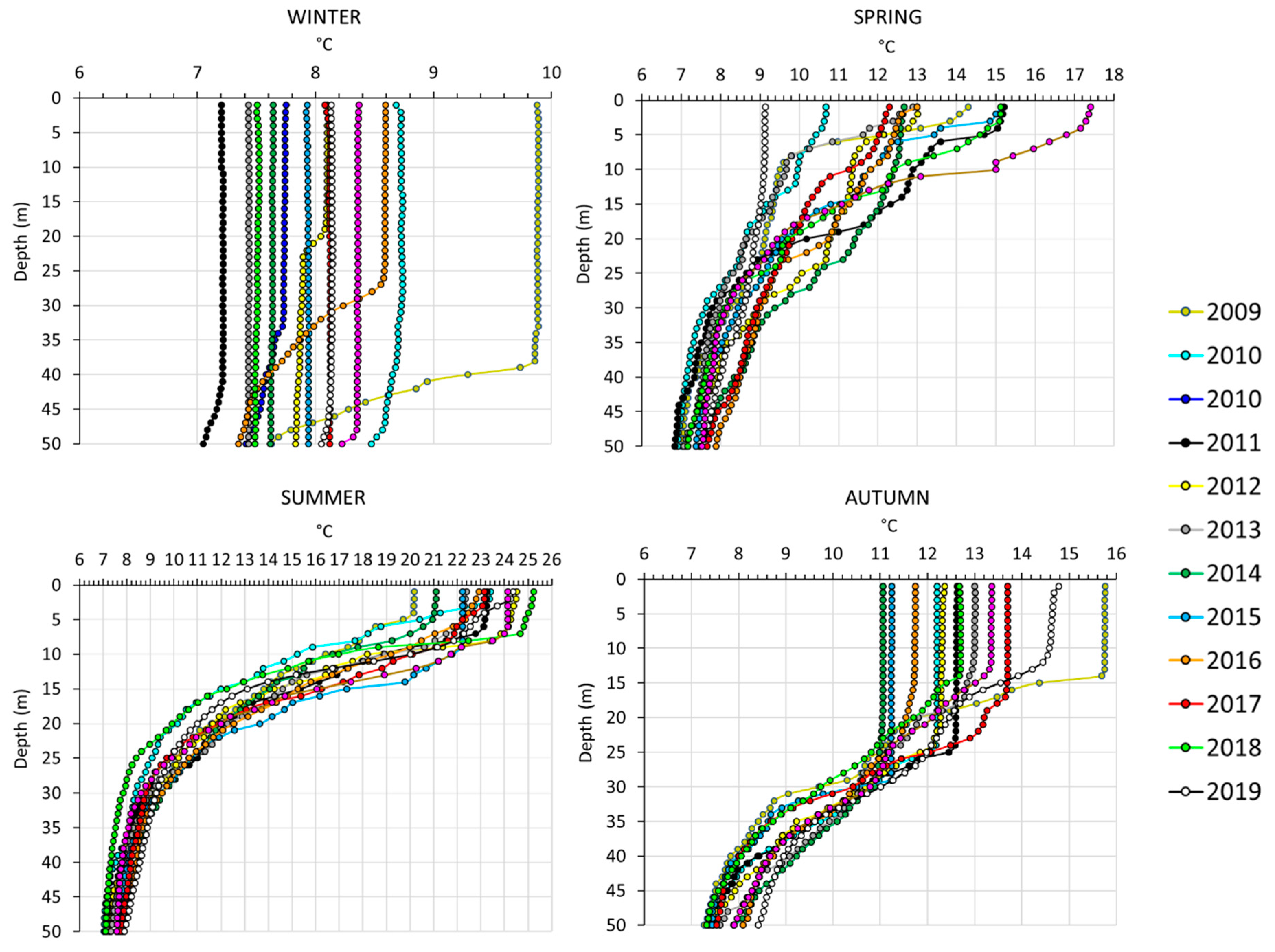

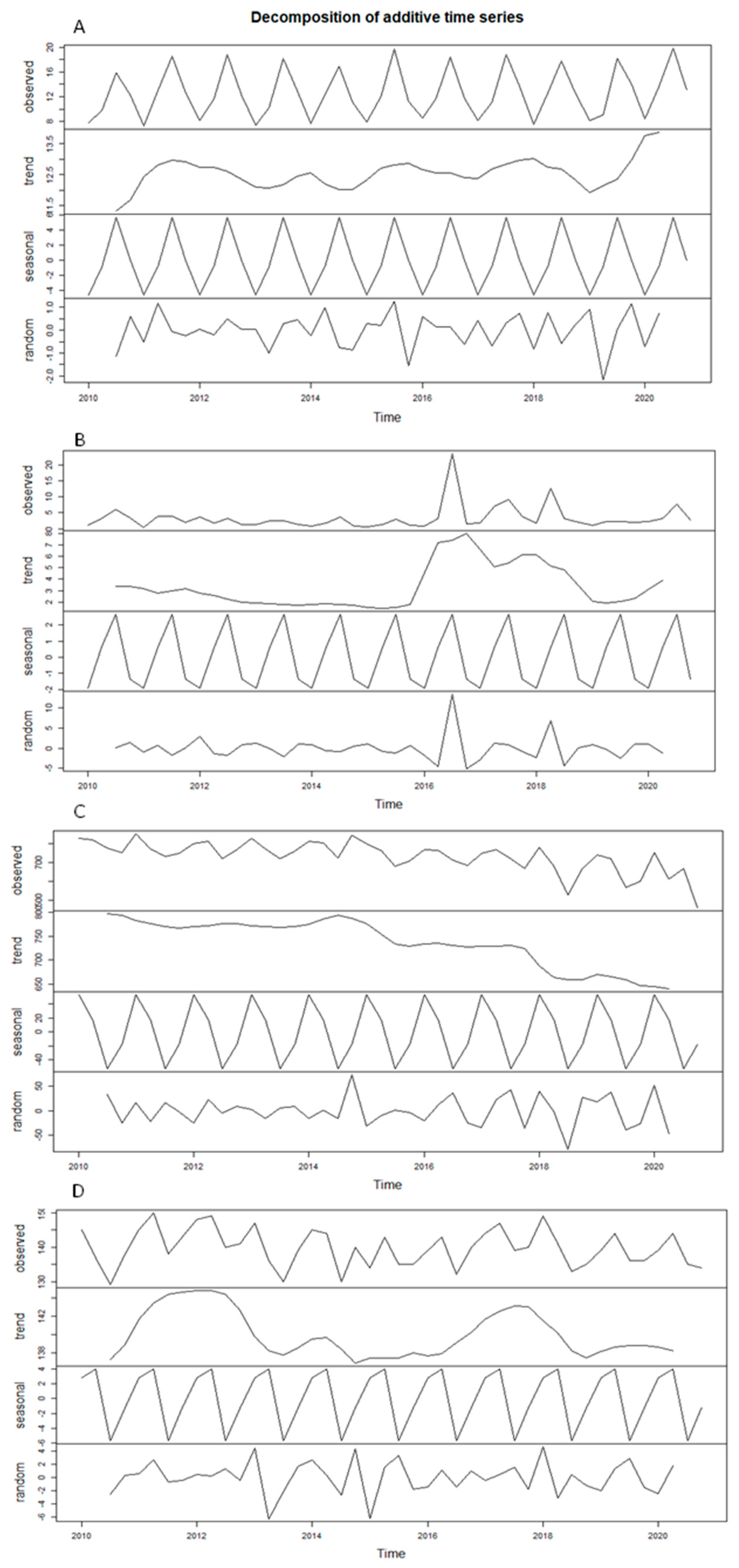
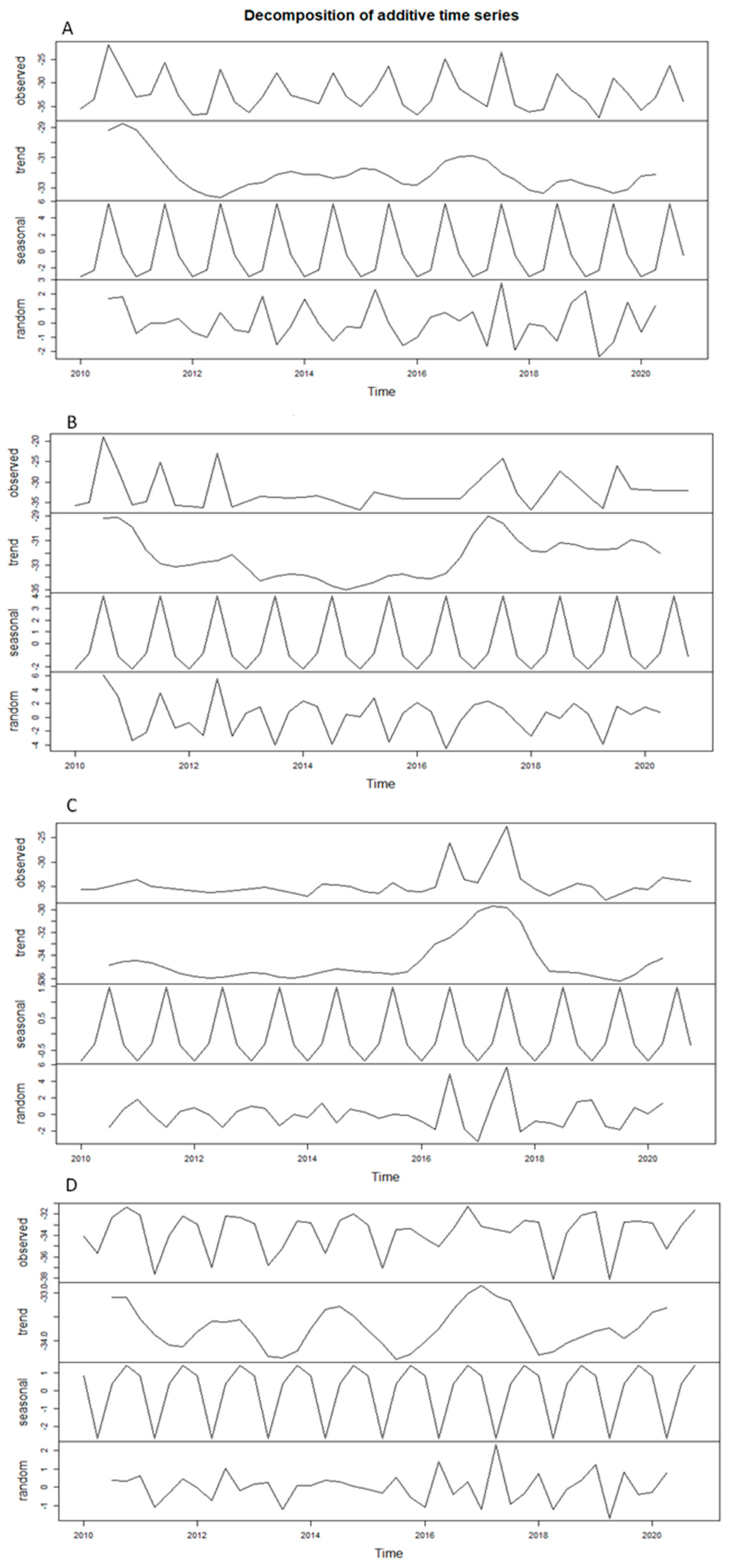
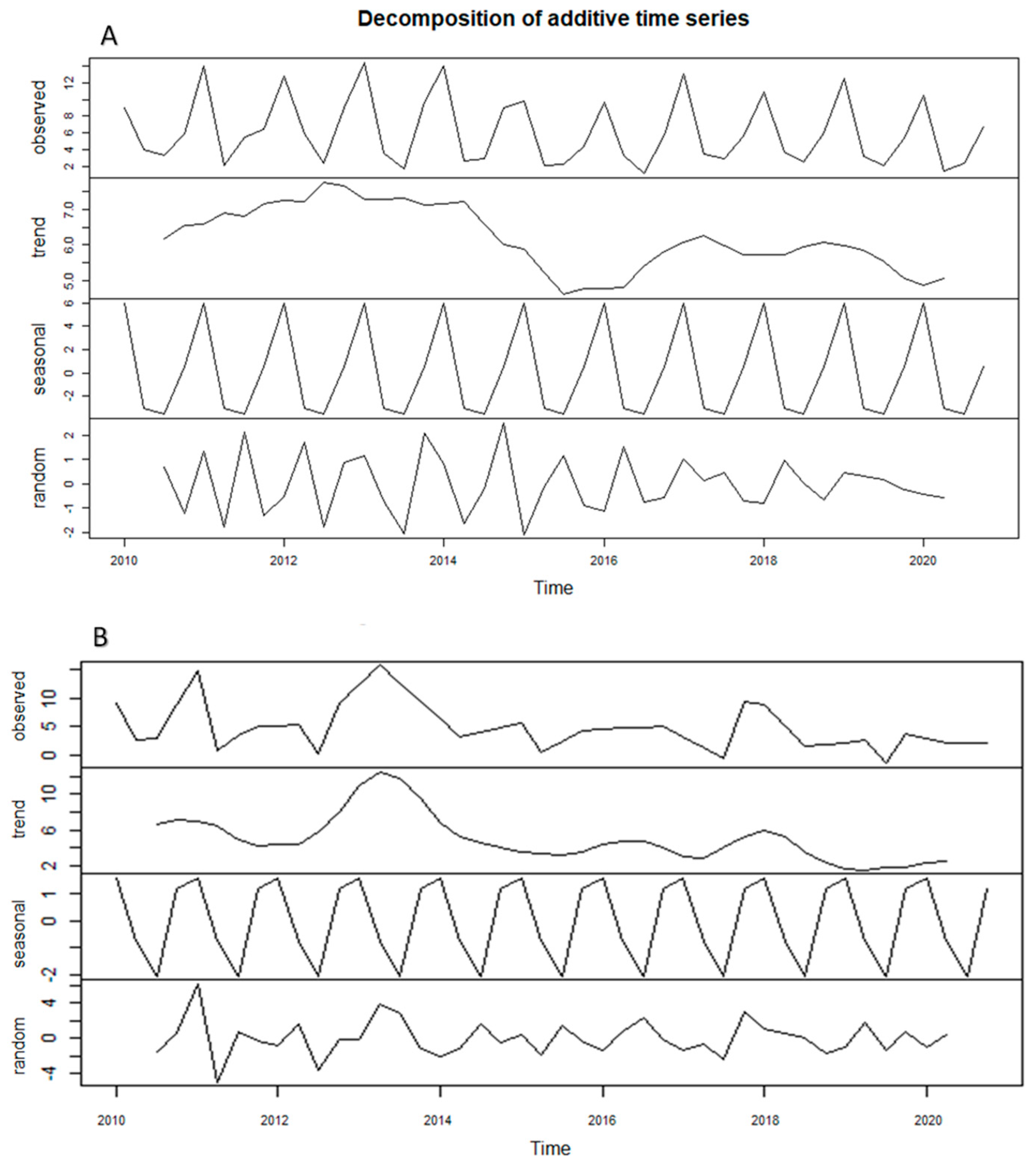
References
- Moss, B. Cogs in the endless machine: Lakes, climate change and nutrient cycles: A review. Sci. Total Environ. 2012, 434, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Kagalou, I.; Papastergiadou, E.; Leonardos, I. Long term changes in the eutrophication process in a shallow Mediterranean lake ecosystem of W. Greece: Response after the reduction of external load. J. Environ. Manag. 2008, 87, 497–506. [Google Scholar] [CrossRef]
- Foley, B.; Jones, I.D.; Maberly, S.C.; Rippey, B. Long-term changes in oxygen depletion in a small temperate lake: Effects of climate change and eutrophication. Freshw. Biol. 2012, 57, 278–289. [Google Scholar] [CrossRef]
- Arfè, A.; Quatto, P.; Zambon, A.; MacIsaac, H.J.; Manca, M. Long-term changes in the zooplankton community of Lake Maggiore in response to multiple stressors: A functional principal components analysis. Water 2019, 11, 962. [Google Scholar] [CrossRef]
- Rogora, M.; Austoni, M.; Caroni, R.; Giacomotti, P.; Kamburska, L.; Marchetto, A.; Mosello, R.; Orrù, A.; Tartari, G.; Dresti, C. Temporal changes in nutrients in a deep oligomictic lake: The role of external loads versus climate change. J. Limnol. 2021, 80, 2051. [Google Scholar] [CrossRef]
- Chang, C.W.; Miki, T.; Ye, H.; Souissi, S.; Adrian, R.; Anneville, O.; Agasild, H.; Ban, S.; Be’eri-Shlevin, Y.; Chiang, Y.R.; et al. Causal networks of phytoplankton diversity and biomass are modulated by environmental context. Nat. Commun. 2022, 13, 1140. [Google Scholar] [CrossRef] [PubMed]
- Poff, N.L.; Brinson, M.M.; Day, J.W. Aquatic ecosystems and global climate change. Pew Cent. Glob. Clim. Change Arlingt. VA 2002, 44, 1–56. Available online: https://www.c2es.org/wp-content/uploads/2002/01/aquatic.pdf (accessed on 27 August 2023).
- Mooij, W.M.; Hülsmann, S.; De Senerpont Domis, L.N.; Nolet, B.A.; Bodelier, P.L.; Boers, P.; Pires, L.M.D.; Gons, H.J.; Ibelings, B.W.; Noordhuis, R.; et al. The impact of climate change on lakes in the Netherlands: A review. Aquat. Ecol. 2005, 39, 381–400. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 2008, 11, 1351–1363. [Google Scholar] [CrossRef]
- Williamson, C.E.; Saros, J.E.; Vincent, W.F.; Smol, J.P. Lakes and reservoirs as sentinels, integrators, and regulators of climate change. Limnol. Oceanogr. 2009, 54, 2273–2282. [Google Scholar] [CrossRef]
- Fenocchi, A.; Rogora, M.; Sibilla, S.; Dresti, C. Relevance of inflows on the thermodynamic structure and on the modeling of a deep subalpine lake (Lake Maggiore, Northern Italy/Southern Switzerland). Limnologica 2017, 63, 42–56. [Google Scholar] [CrossRef]
- Tanentzap, A.J.; Morabito, G.; Volta, P.; Rogora, M.; Yan, N.D.; Manca, M. Climate warming restructures an aquatic food web over 28 years. Glob. Change Biol. 2020, 26, 6852–6866. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Holmgren, K.; González-Bergonzoni, I.; Teixeira-de Mello, F.; Declerck, S.A.; De Meester, L.; Søndergaard, M.; Lauridsen, T.L.; Bjerring, R.; et al. Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia 2010, 646, 73–90. [Google Scholar] [CrossRef]
- Jeppesen, E.; Brucet, S.; Naselli-Flores, L.; Papastergiadou, E.; Stefanidis, K.; Noges, T.; Noges, P.; Attayde, J.L.; Zohary, T.; Coppens, J.; et al. Ecological impacts of global warming and water abstraction on lakes and reservoirs due to changes in water level and related changes in salinity. Hydrobiologia 2015, 750, 201–227. [Google Scholar] [CrossRef]
- Lin, Q.; Xu, L.; Hou, J.; Liu, Z.; Jeppesen, E.; Han, B.P. Responses of trophic structure and zooplankton community to salinity and temperature in Tibetan lakes: Implication for the effect of climate warming. Water Res. 2017, 124, 618–629. [Google Scholar] [CrossRef]
- Rantala, M.V.; Luoto, T.P.; Weckström, J.; Rautio, M.; Nevalainen, L. Climate drivers of diatom distribution in shallow subarctic lakes. Freshw. Biol. 2017, 62, 1971–1985. [Google Scholar] [CrossRef]
- Woolway, R.I.; Merchant, C.J. Worldwide alteration of lake mixing regimes in response to climate change. Nat. Geosci. 2019, 12, 271–276. [Google Scholar] [CrossRef]
- Woolway, R.I.; Kraemer, B.M.; Lenters, J.D.; Merchant, C.J.; O’Reilly, C.M.; Sharma, S. Global lake responses to climate change. Nat. Rev. Earth Environ. 2020, 1, 388–403. [Google Scholar] [CrossRef]
- Sommer, U.; Adrian, R.; De Senerpont Domis, L.; Elser, J.J.; Gaedke, U.; Ibelings, B.; Jeppesen, E.; Lurling, M.; Molinero, J.C.; Mooij, W.M.; et al. Beyond the Plankton Ecology Group(PEG) Model: Mechanisms Driving Plankton Succession. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- Varpe, Ø. Fitness and phenology: Annual routines and zooplankton adaptations to seasonal cycles. J. Plankton Res. 2012, 34, 267–276. [Google Scholar] [CrossRef]
- Kürten, B.; Painting, S.J.; Struck, U.; Polunin, N.V.; Middelburg, J.J. Tracking seasonal changes in North Sea zooplankton trophic dynamics using stable isotopes. Biogeochemistry 2013, 113, 167–187. [Google Scholar] [CrossRef]
- Bănaru, D.; Carlotti, F.; Barani, A.; Grégori, G.; Neffati, N.; Harmelin-Vivien, M. Seasonal variation of stable isotope ratios of size-fractionated zooplankton in the Bay of Marseille (NW Mediterranean Sea). J. Plankton Res. 2014, 36, 145–156. [Google Scholar] [CrossRef]
- Moss, B.R. Ecology of Fresh Waters: Man and Medium, Past to Future; John Wiley & Sons Publishing: Hoboken, NJ, USA, 2009; p. 482. ISBN 1405113324. [Google Scholar]
- Visconti, A.; Caroni, R.; Rawcliffe, R.; Fadda, A.; Piscia, R.; Manca, M. Defining seasonal functional traits of a freshwater zooplankton community using δ13C and δ15N stable isotope analysis. Water 2018, 10, 108. [Google Scholar] [CrossRef]
- Cicala, D.; Polgar, G.; Mor, J.R.; Piscia, R.; Brignone, S.; Zaupa, S.; Volta, P. Trophic niches, trophic positions, and niche overlaps between non-native and native fish species in a subalpine lake. Water 2020, 12, 3475. [Google Scholar] [CrossRef]
- Pel, R.; Hoogveld, H.; Floris, V. Using the hidden isotopic heterogeneity in phyto-and zooplankton to unmask disparity in trophic carbon transfer. Limnol. Oceanogr. 2003, 48, 2200–2207. [Google Scholar] [CrossRef]
- Matthews, B.; Mazumder, A. Temporal variation in body composition (C:N) helps explain seasonal patterns of zooplankton δ13C. Freshwat. Biol. 2005, 50, 502–515. [Google Scholar] [CrossRef]
- Perga, M.E.; Gerdaux, D. Seasonal variability in the δ13C and δ15N values of the zooplankton taxa in two alpine lakes. Acta Oecol. 2006, 30, 69–77. [Google Scholar] [CrossRef]
- Goericke, R.; Fry, B. Variations of marine plankton δ13C with latitude, temperature, and dissolved CO2 in the world ocean. Glob. Biogeochem. Cycles 1994, 8, 85–90. [Google Scholar] [CrossRef]
- Rau, G.H.; Takahashi, T.; Marais, D.J.D. Latitudinal variations in plankton δ13C: Implications for CO2 and productivity in past oceans. Nature 1989, 341, 516–518. [Google Scholar] [CrossRef]
- Zohary, T.; Erez, J.; Gophen, M.; Berman-Frank, I.; Stiller, M. Seasonality of stable carbon isotopes within the pelagic food web of Lake Kinneret. Limnol. Oceanogr. 1994, 39, 1030–1043. [Google Scholar] [CrossRef]
- Schindler, D.W. The cumulative effects of climate warming and other human stresses on Canadian freshwaters in the new millennium. Can. J. Fish. Aquat. Sci. 2001, 58, 18–29. [Google Scholar] [CrossRef]
- Visconti, A.; Manca, M.; de Bernardi, R. Eutrophication-like response to climate warming: An analysis of Lago Maggiore (N. Italy) zooplankton in contrasting years. J. Limnol. 2008, 67, 87–92. [Google Scholar] [CrossRef]
- Takahashi, K.; Yoshioka, T.; Wada, E.; Sakamoto, M. Temporal variations in carbon isotope ratio of phytoplankton in a eutrophic lake. J. Plankton Res. 1990, 12, 799–808. [Google Scholar] [CrossRef]
- Ruggiu, D.; Morabito, G.; Panzani, P.; Pugnetti, A. Trends and relations among basic phytoplankton characteristics in the course of the long-term oligotrophication of Lake Maggiore (Italy). Hydrobiologia 1998, 369/370, 243–257. [Google Scholar] [CrossRef]
- Salmaso, N.; Mosello, R. Limnological research in the deep southern subalpine lakes: Synthesis, directions and perspectives. Adv. Oceanogr. Limnol. 2010, 1, 29–66. [Google Scholar] [CrossRef]
- Tapolczai, K.; Anneville, O.; Padisák, J.; Salmaso, N.; Morabito, G.; Zohary, T.; Tadonléké, R.D.; Rimet, F. Occurrence and mass development of Mougeotia spp. (Zygnemataceae) in large, deep lakes. Hydrobiologia 2015, 745, 17–29. [Google Scholar] [CrossRef]
- Bresciani, M.; Cazzaniga, I.; Austoni, M.; Sforzi, T.; Buzzi, F.; Morabito, G.; Giardino, C. Mapping phytoplankton blooms in deep subalpine lakes from Sentinel-2A and Landsat-8. Hydrobiologia 2018, 824, 197–214. [Google Scholar] [CrossRef]
- Morabito, G.; Oggioni, A.; Austoni, M. Resource ratio and human impact: How diatom assemblages in Lake Maggiore responded to oligotrophication and climatic variability. In Phytoplankton Responses to Human Impacts at Different Scales; Salmaso, N., Naselli-Flores, L., Cerasino, L., Flaim, G., Tolotti, M., Padisák, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 221, pp. 47–60. [Google Scholar] [CrossRef]
- Tanentzap, A.J.; Fitch, A.; Orland, C.; Emilson, E.J.; Yakimovich, K.M.; Osterholz, H.; Dittmar, T. Chemical and microbial diversity covary in fresh water to influence ecosystem functioning. Proc. Natl. Acad. Sci. USA 2019, 116, 24689–24695. [Google Scholar] [CrossRef]
- Fenocchi, A.; Rogora, M.; Sibilla, S.; Ciampittiello, M.; Dresti, C. Forecasting the evolution in the mixing regime of a deep subalpine lake under climate change scenarios through numerical modelling (Lake Maggiore, Northern Italy/Southern Switzerland). Clim. Dyn. 2018, 51, 3521–3536. [Google Scholar] [CrossRef]
- Rogora, M.; Buzzi, F.; Dresti, C.; Leoni, B.; Lepori, F.; Mosello, R.; Patelli, M.; Salmaso, N. Climatic effects on vertical mixing and deep-water oxygen content in the subalpine lakes in Italy. Hydrobiologia 2018, 824, 33–50. [Google Scholar] [CrossRef]
- Gorham, E.; Boyce, F.M. Influence of lake surface area and depth upon thermal stratification and the depth of the summer thermocline. J. Great Lakes Res. 1989, 15, 233–245. [Google Scholar] [CrossRef]
- Piscia, R.; Manca, M.; Caroni, R.; Guilizzoni, P.; Bettinetti, R. Zooplankton Taxa Repository of DDTtot and sumPCB14: Seasonal and Decadal Variations in Lake Maggiore. Available at SSRN 4485232. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=4485232 (accessed on 27 August 2023).
- Tonolli, L. Holomixy and oligomixy in Lake Maggiore: Inference on the vertical distribution of zooplankton. Int. Ver. Theor. Angew. Limnol. Verhandlungen 1969, 17, 231–236. [Google Scholar] [CrossRef]
- Visconti, A.; Manca, M. Seasonal changes in the δ13C and δ15N signatures of the Lago Maggiore pelagic food web. J. Limnol. 2011, 70, 263–271. [Google Scholar] [CrossRef]
- Matthews, B.; Mazumder, A. Compositional and interlake variability of zooplankton affect baseline stable isotope signatures. Limnol. Oceanogr. 2003, 48, 1977–1987. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnology: Lake and River Ecosystems; Academic Press: London, UK, 2001; p. 1024. ISBN 9780080574394. [Google Scholar]
- Post, D.M.; Pace, M.L.; Hairston, N.G., Jr. Ecosystem size determines food-chain length in lakes. Nature 2000, 405, 1047–1049. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- Ambrosetti, W.; Barbanti, L.; Carrara, E.A. Mechanisms of hypolimnion erosion in a deep lake (Lago Maggiore, N. Italy). J. Limnol. 2010, 69, 3–14. [Google Scholar] [CrossRef]
- APHA; AWWA; WEF. Standard Methods for the Examination of Water & Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- IRSA-CNR, A.P.A.T. Metodi Analitici per le Acque; Manuali e Linee Guida; IRSA-CNR: Brugherio, Italy, 2003; Volume 29, p. 1153. ISBN 88-448-0083-7. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer Cham Publishing: Berlin/Heidelberg, Germany, 2006; p. 260. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (Version 3.6.1); R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 27 August 2023).
- Velleman, P.F. Data Desk: Handbook; Data Description, Inc.: New York, NY, USA, 1989; Volume 1. [Google Scholar]
- Piscia, R.; Mazzoni, M.; Bettinetti, R.; Caroni, R.; Cicala, D.; Manca, M.M. Stable Isotope Analysis and persistent organic pollutants in crustacean zooplankton: The role of size and seasonality. Water 2019, 11, 1490. [Google Scholar] [CrossRef]
- Lehmann, M.F.; Bernasconi, S.M.; McKenzie, J.A. Seasonal variation of the δ13C and δ15N of particulate and dissolved carbon and nitrogen in Lake Lugano: Constraints on biogeochemical cycling in a eutrophic lake. Limnol. Oceanogr. 2004, 49, 415–429. [Google Scholar] [CrossRef]
- Grey, J.; Jones, R.I. Seasonal changes in the importance of the source of organic matter to the diet of zooplankton in Loch Ness, as indicated by stable isotope analysis. Limnol. Oceanogr. 2001, 46, 505–513. [Google Scholar] [CrossRef]
- Caroni, R.; Free, G.; Visconti, A.; Manca, M. Phytoplankton functional traits and seston stable isotopes signature: A functional-based approach in a deep, subalpine lake, Lake Maggiore (N. Italy). J. Limnol. 2012, 71, 84–94. [Google Scholar] [CrossRef]
- Gearing, J.N.; Gearing, P.J.; Rudnick, D.T.; Requejo, A.G.; Hutchins, M.J. Isotopic variability of organic carbon in a phytoplankton-based, temperate estuary. Geochim. Cosmochim. Acta 1984, 48, 1089–1098. [Google Scholar] [CrossRef]
- Miller, T.W.; Brodeur, R.D.; Rau, G.H. Carbon stable isotopes reveal relative contribution of shelf-slope production to the Northern California Current pelagic community. Limnol. Oceanogr. 2008, 53, 1493–1503. [Google Scholar] [CrossRef]
- Lara, R.J.; Alder, V.; Franzosi, C.A.; Kattner, G. Characteristics of suspended particulate organic matter in the southwestern Atlantic: Influence of temperature, nutrient and phytoplankton features on the stable isotope signature. J. Mar. Syst. 2010, 79, 199–209. [Google Scholar] [CrossRef]
- Papiol, V.; Cartes, J.E.; Fanelli, E.; Rumolo, P. Food web structure and seasonality of slope megafauna in the NW Mediterranean elucidated by stable isotopes: Relationship with available food sources. J. Sea Res. 2013, 77, 53–69. [Google Scholar] [CrossRef]
- Kürten, B.; Al-Aidaroos, A.M.; Struck, U.; Khomayis, H.S.; Gharbawi, W.Y.; Sommer, U. Influence of environmental gradients on C and N stable isotope ratios in coral reef biota of the Red Sea, Saudi Arabia. J. Sea Res. 2014, 85, 379–394. [Google Scholar] [CrossRef]
- Gu, B.; Schelske, C.L.; Waters, M.N. Patterns and controls of seasonal variability of carbon stable isotopes of particulate organic matter in lakes. Oecologia 2011, 165, 1083–1094. [Google Scholar] [CrossRef]
- Hou, W.; Gu, B.; Zhang, H.; Gu, J.; Han, B.P. The relationship between carbon and nitrogen stable isotopes of zooplankton and select environmental variables in low-latitude reservoirs. Limnology 2013, 14, 97–104. [Google Scholar] [CrossRef]
- Manca, M.; DeMott, W.R. Response of the invertebrate predator Bythotrephes to a climate-linked increase in the duration of a refuge from fish predation. Limnol. Oceanogr. 2009, 54, 2506–2512. [Google Scholar] [CrossRef]
- Manca, M.; Portogallo, M.; Brown, M.E. Shifts in phenology of Bythotrephes longimanus and its modern success in Lake Maggiore as a result of changes in climate and trophy. J. Plankton Res. 2007, 29, 515–525. [Google Scholar] [CrossRef]
- Manca, M.; Cavicchioni, N.; Morabito, G. First observations on the effect of a complete, exceptional overturn of Lake Maggiore on plankton and primary productivity. Int. Rev. Hydrobiol. 2000, 85, 209–222. [Google Scholar] [CrossRef]
- Fadda, A.; Rawcliffe, R.; Padedda, B.M.; Lugliè, A.; Sechi, N.; Camin, F.; Ziller, L.; Manca, M. Spatiotemporal dynamics of C and N isotopic signature of zooplankton: A seasonal study on a man-made lake in the Mediterranean region. Ann. Limnol. Int. J. Limnol. 2014, 50, 279–287. [Google Scholar] [CrossRef]
- Rau, G.H.; Takahashi, T.; Des Marais, D.J.; Repeta, D.J.; Martin, J.L. The relationship between δ13C of organic matter and [CO2 (aq)] in ocean surface water: Data from a JGOFS site in the northeast Atlantic Ocean and a model. Geochim. Cosmochim. Acta 1992, 56, 1413–1419. [Google Scholar] [CrossRef]
- Yoshioka, T.; Wada, E.; Hayashi, H. A stable isotope study on seasonal food web dynamics in a eutrophic lake. Ecology 1994, 75, 835–846. [Google Scholar] [CrossRef]
- Gu, B.; Alexander, V.; Schell, D.M. Seasonal and interannual variability of plankton carbon isotope ratios in a subarctic lake. Freshwat. Biol. 1999, 42, 417–426. [Google Scholar] [CrossRef]
- Gu, B.; Schelske, D.M. Temporal and spatial variations in phytoplankton carbon isotopes in a polymictic subtropical lake. J. Plankton Res. 1996, 18, 2081–2092. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Rasmussen, J.B. Primary consumer δ13C and δ15N and the trophic position of aquatic consumers. Ecology 1999, 1395–1404. [Google Scholar] [CrossRef]
- Grey, J.; Jones, R.I.; Sleep, D. Stable isotope analysis of the origins of zooplankton carbon in lakes of differing trophic state. Oecologia 2000, 123, 232–240. [Google Scholar] [CrossRef]
- Cattaneo, A.; Manca, M.; Rasmussen, J.B. Peculiarities in the stable isotope composition of organisms from an alpine lake. Aquat. Sci. 2004, 66, 440–445. [Google Scholar] [CrossRef]
- Leoni, B. Zooplankton predators and preys: Body size and stable isotope to investigate the pelagic food in a deep lake (Lake Iseo, Northern Italy). J. Limnol. 2017, 76, 85–93. [Google Scholar] [CrossRef]
- Makino, W.; Yoshida, T.; Sakano, H.; Ban, S. Stay cool: Habitat selection of a cyclopoid copepod in a north temperate oligotrophic lake. Freshw. Biol. 2003, 48, 1551–1562. [Google Scholar] [CrossRef]
- Helland, I.P.; Freyhof, J.; Kasprzak, P.; Mehner, T. Temperature sensitivity of vertical distributions of zooplankton and planktivorous fish in a stratified lake. Oecologia 2007, 151, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Thackeray, S.J.; Elliott, J.A.; Fielding, R.F.; Swinburne, K. Effects of onset of thermal stratification on vertical distribution of phytoplankton and zooplankton species. Int. Ver. Theor. Angew. Limnol. Verhandlungen 2005, 29, 555–559. [Google Scholar] [CrossRef]
- Smyntek, P.M.; Maberly, S.C.; Grey, J. Dissolved carbon dioxide concentration controls baseline stable carbon isotope signatures of a lake food web. Limnol. Oceanogr. 2012, 57, 1292–1302. [Google Scholar] [CrossRef]
- Smyntek, P.M.; Teece, M.A.; Schulz, K.L.; Storch, A.J. Taxonomic differences in the essential fatty acid composition of groups of freshwater zooplankton relate to reproductive demands and generation time. Freshwat. Biol. 2008, 53, 1768–1782. [Google Scholar] [CrossRef]
- France, R.L. Differentiation between littoral and pelagic food webs in lakes using stable carbon isotopes. Limnol. Oceanogr. 1995, 40, 1310–1313. [Google Scholar] [CrossRef]
- Croteau, M.N.; Luoma, S.N.; Stewart, A.R. Trophic transfer of metals along freshwater food webs: Evidence of cadmium biomagnification in nature. Limnol. Oceanogr. 2005, 50, 1511–1519. [Google Scholar] [CrossRef]
- Visconti, A.; Volta, P.; Fadda, A.; Manca, M. Roach in Lake Maggiore: A Peaceful Invasion Detected with C, N Stable Isotope Analysis. Glob. J. Sci. Front. Res. Agric. Vet. 2013, 13, 1–8. [Google Scholar]
- Buhan, E.; Kaymak, N.; Akin, S.; Turan, H. Trophic Pathways from Pelagic and Littoral Sources Supports Food Web in A Eutrophic Natural Lake (Lake Zinav, Turkey). Turk. J. Fish. Aquat. Sci. 2018, 19, 99–107. [Google Scholar] [CrossRef]
- Geller, W.; Müller, H. The filtration apparatus of Cladocera: Filter mesh-sizes and their implications on food selectivity. Oecologia 1981, 49, 316–321. [Google Scholar] [CrossRef] [PubMed]
- DeMott, W.R.; Kerfoot, C.W. Competition among cladocerans: Nature of the interaction between Bosmina and Daphnia. Ecology 1982, 63, 1949–1966. [Google Scholar] [CrossRef]
- Deng, D.; Xie, P.; Zhou, O.; Yang, H.; Guo, L.; Geng, H. Field and experimental studies on the combined impacts of cyanobacterial blooms and small algae on crustacean zooplankton in a large, eutrophic, subtropical, Chinese lake. Limnology 2008, 9, 1–11. [Google Scholar] [CrossRef]
- Carpenter, E.J.; Montoya, J.P.; Burns, J.; Mulholland, M.R.; Subramaniam, A.; Capone, D.G. Extensive bloom of a N2-fixing diatom/cyanobacterial association in the tropical Atlantic Ocean. Mar. Ecol. Progr. Ser. 1999, 185, 273–283. [Google Scholar] [CrossRef]
- Bauersachs, T.; Schouten, S.; Compaoré, J.; Wollenzien, U.; Stal, L.J.; Damsteé, J.N.S. Nitrogen isotopic fractionation associated with growth on dinitrogen gas and nitrate by cyanobacteria. Limnol Oceanog. 2009, 54, 1403–1411. [Google Scholar] [CrossRef]
- Gu, B.; Chapman, A.D.; Schelske, C.L. Factors controlling seasonal variations in stable isotope composition of particulate organic matter in a softwater eutrophic lake. Limnol. Oceanogr. 2006, 51, 2837–2848. [Google Scholar] [CrossRef]
- Jones, R.I.; Grey, J.; Quarmby, C.; Sleep, D. An assessment using stable isotopes of the importance of allochthonous organic carbon sources to the pelagic food web in Loch Ness. Proc. R. Soc. Lond. B 1998, 265, 105–111. [Google Scholar] [CrossRef]
- Winder, M.; Hunter, D.A. Temporal organization of phytoplankton communities linked to physical forcing. Oecologia 2008, 156, 179–192. [Google Scholar] [CrossRef]
- Bertoni, R.; Callieri, C. Organic carbon trend during the oligotrophication of Lago Maggiore. Mem. Ist. Ital. Idrobiol. 1993, 52, 191–205. [Google Scholar]
- Smith, V.H. Nutrient dependence of primary productivity in lakes 1. Limnol. Oceanogr. 1979, 24, 1051–1064. [Google Scholar] [CrossRef]
- Beaver, J.R.; Crisman, T.L. Temporal variability in algal biomass and primary productivity in Florida lakes relative to latitudinal gradients, organic color and trophic state. Hydrobiologia 1991, 224, 89–97. [Google Scholar] [CrossRef]
- Cifuentes, L.A.; Sharp, J.H.; Fogel, M.L. Stable carbon and nitrogen isotope biogeochemistry in the Delaware estuary. Limnol. Oceanogr. 1998, 33, 1102–1115. [Google Scholar] [CrossRef]
- Nakatsuka, T.; Handa, N.; Wada, E.; Wong, C.S. The dynamic changes of stable isotopic ratios of carbon and nitrogen in suspended and sedimented particulate organic matter during a phytoplankton bloom. J. Mar. Res. 1992, 50, 267–296. [Google Scholar] [CrossRef]
- Takahashi, A.; Takeda, K.; Ohnishi, T. Light-induced anthocyanin reduces the extent of damage to DNA in UV-irradiated Centaurea cyanus cells in culture. Plant Cell Physiol. 1991, 32, 541–547. [Google Scholar]
- Laws, E.A.; Popp, B.N.; Bidigare, R.R.; Kennicutt, M.C.; Macko, S.A. Dependence of phytoplankton carbon isotopic composition on growth rate and [CO2) aq: Theoretical considerations and experimental results. Geochim. Cosmochim. Acta 1995, 59, 1131–1138. [Google Scholar] [CrossRef]
- Francois, R.; Altabet, M.A.; Goericke, R.; McCorkle, D.C.; Brunet, C.; Poisson, A. Changes in the δ13C of surface water particulate organic matter across the subtropical convergence in the SW Indian Ocean. Glob. Biogeochem. Cycles 1993, 7, 627–644. [Google Scholar] [CrossRef]
- France, R.L.; Del Giorgio, P.A.; Westcott, K.A. Productivity and heterotrophy influences on zooplankton delta13C in northern temperate lakes. Aquat. Microb. Ecol. 1997, 12, 85–93. [Google Scholar] [CrossRef]
- Leggett, M.F.; Servos, M.R.; Hesslein, R.; Johannsson, O.; Millard, E.S.; Dixon, D.G. Biogeochemical influences on the carbon isotope signatures of Lake Ontario biota. Can. J. Fish. Aquat. Sci. 1999, 56, 2211–2218. [Google Scholar] [CrossRef]
- Leggett, M.F.; Johannsson, O.; Hesslein, R.; Dixon, D.G.; Taylor, W.D.; Servos, M.R. Influence of inorganic nitrogen cycling on the δ15N of Lake Ontario biota. Can. J. Fish. Aquat. Sci. 2000, 57, 1489–1496. [Google Scholar] [CrossRef]
- Vuorio, K.; Meili, M.; Sarvala, J. Taxon-specific variation in the stable isotopic signatures (δ13C and δ15N) of lake phytoplankton. Freshwat. Biol. 2006, 51, 807–822. [Google Scholar] [CrossRef]
- Free, G.; Bresciani, M.; Pinardi, M.; Ghirardi, N.; Luciani, G.; Caroni, R.; Giardino, C. Detecting climate driven changes in chlorophyll-a in deep subalpine lakes using long term satellite data. Water 2021, 13, 866. [Google Scholar] [CrossRef]
- Gu, B.; Ma, L.; Smoak, D.; Ewe, S.; Zhu, Y.; Irick, D.; Ross, M.; Li, Y. Mercury and Sulfur Environmental Assessment for the Everglades. South Florida Environmental Report. South Florida Water Management District, West Palm Beach. 2013, pp. 3B18–3B20. Available online: https://www.researchgate.net/profile/Donald-Axelrad/publication/265107992_Chapter_3B_Mercury_and_Sulfur_Monitoring_Research_and_Environmental_Assessment_in_South_Florida/links/5404b7a30cf2bba34c1d006d/Chapter-3B-Mercury-and-Sulfur-Monitoring-Research-and-Environmental-Assessment-in-South-Florida.pdf (accessed on 27 August 2023).
- Gu, B.; Schell, D.M.; Alexander, V. Stable carbon and nitrogen isotopic analysis of the plankton food web in a subarctic lake. Can. J. Fish. Aquat. Sci. 1994, 51, 1338–1344. [Google Scholar] [CrossRef]
- Gulati, R.D.; Bronkhorst, M.; Van Donk, E. Feeding in Daphnia galeata on Oscillatoria limnetica and on detritus derived from it. J. Plankton Res. 2001, 23, 705–718. [Google Scholar] [CrossRef]
- Fry, B.; Wainright, S.C. Diatom sources of 13C-rich carbon in marine food webs. Mar. Ecol. Progr. Ser. 1991, 76, 149–157. [Google Scholar] [CrossRef]
- Rogora, M.; Steingruber, S.; Marchetto, A.; Mosello, R.; Giacomotti, P.; Orru, A.; Tartari, G.A.; Tiberti, R. Response of atmospheric deposition and surface water chemistry to the COVID-19 lockdown in an alpine area. Environ. Sci. Pollut. Res. 2022, 29, 62312–62329. [Google Scholar] [CrossRef]
- Webb, B.W.; Hannah, D.M.; Moore, R.D.; Brown, L.E.; Nobilis, F. Recent advances in stream and river temperature research. Hydrol. Process. 2008, 22, 902–918. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2093–2106. [Google Scholar] [CrossRef]
- McQueen, D.J.; Johannes, M.R.S.; Post, J.R.; Stewart, T.J.; Lean, D.R.S. Bottom-up and top-down impacts on freshwater pelagic community structure. Ecol. Monogr. 1989, 59, 289–309. [Google Scholar] [CrossRef]
- Brett, M.T.; Goldman, C.R. A meta-analysis of the freshwater trophic cascade. Proc. Natl. Acad. Sci. USA 1996, 93, 7723–7726. [Google Scholar] [CrossRef]
- Brett, M.T.; Goldman, C.R. Consumer versus resource control in freshwater pelagic food webs. Science 1997, 275, 384–386. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caroni, R.; Piscia, R.; Free, G.; Manca, M. Interpreting Seasonal Patterns and Long-Term Changes of Zooplankton in a Deep Subalpine Lake Using Stable Isotope Analysis. Water 2023, 15, 3143. https://doi.org/10.3390/w15173143
Caroni R, Piscia R, Free G, Manca M. Interpreting Seasonal Patterns and Long-Term Changes of Zooplankton in a Deep Subalpine Lake Using Stable Isotope Analysis. Water. 2023; 15(17):3143. https://doi.org/10.3390/w15173143
Chicago/Turabian StyleCaroni, Rossana, Roberta Piscia, Gary Free, and Marina Manca. 2023. "Interpreting Seasonal Patterns and Long-Term Changes of Zooplankton in a Deep Subalpine Lake Using Stable Isotope Analysis" Water 15, no. 17: 3143. https://doi.org/10.3390/w15173143
APA StyleCaroni, R., Piscia, R., Free, G., & Manca, M. (2023). Interpreting Seasonal Patterns and Long-Term Changes of Zooplankton in a Deep Subalpine Lake Using Stable Isotope Analysis. Water, 15(17), 3143. https://doi.org/10.3390/w15173143