

Decreased Efficiency of Free Naked DNA Transformation by Chlorine and UV Disinfection and Its Detection Limitations
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Preparation
2.2. Disinfection of Plasmid DNA Using Chlorine and UV Irradiation
2.3. pUC19 Transformation into E. coli and Plate Colony-Forming Unit Counting
2.4. PCR and qPCR Detection of the Treated pUC19
3. Results
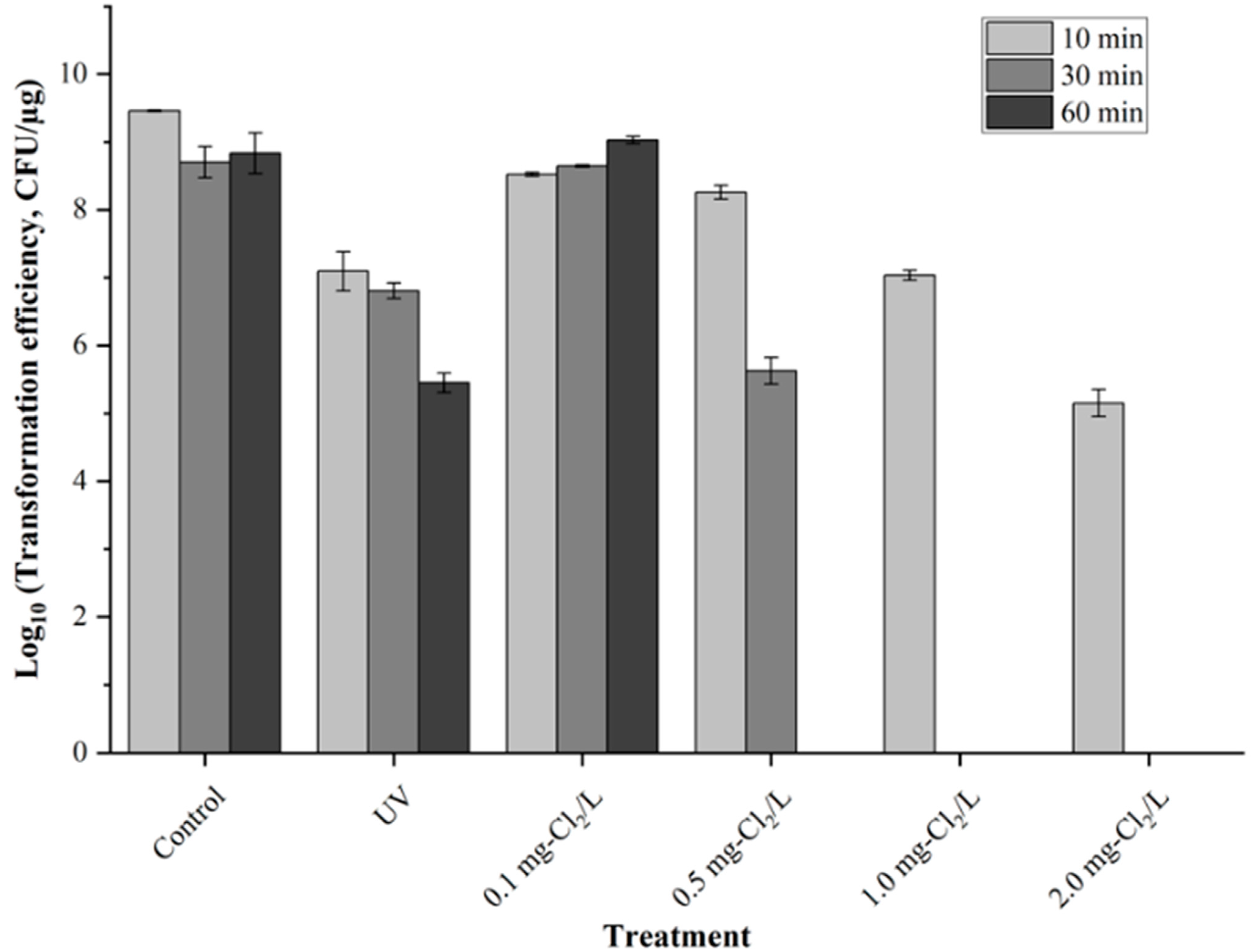
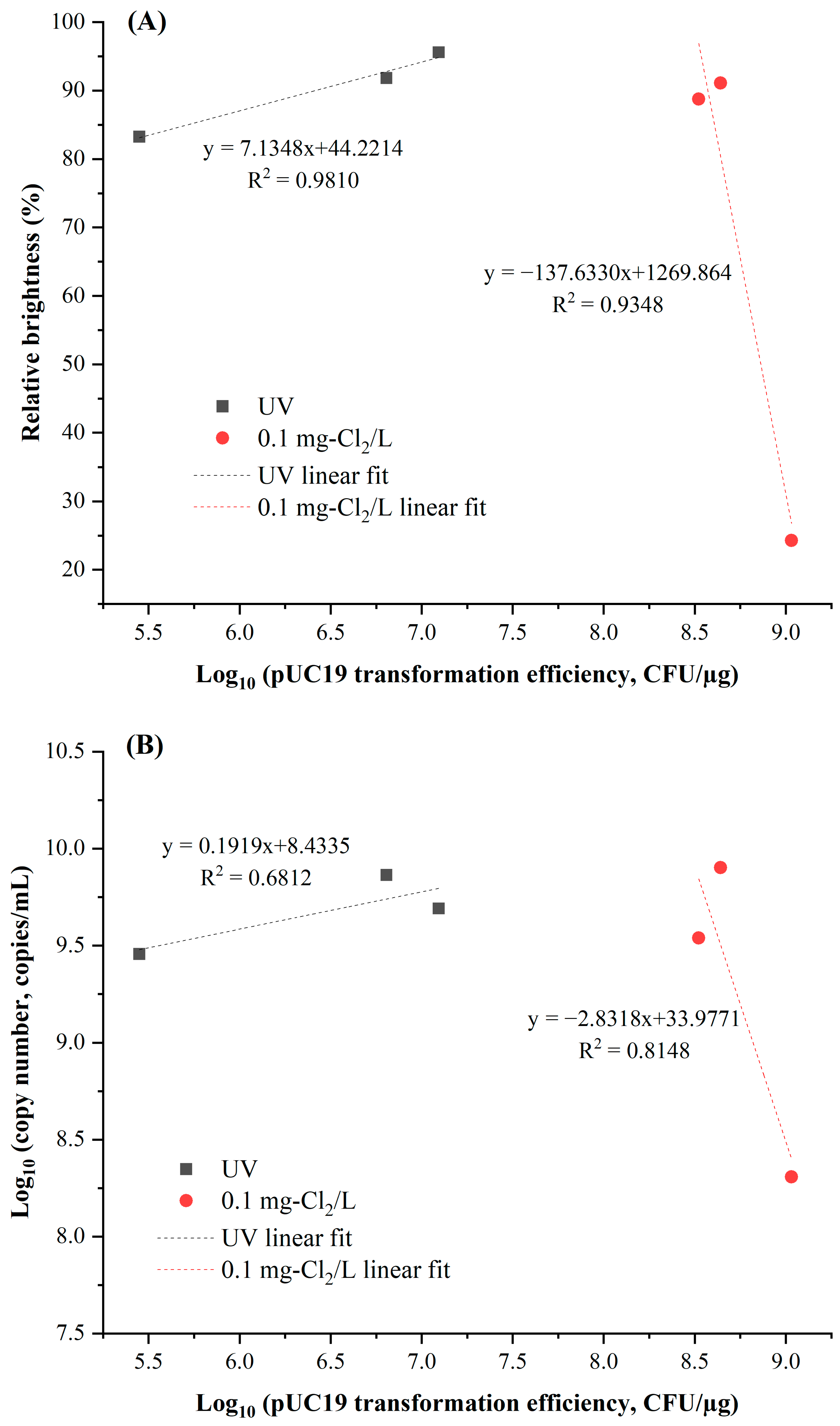
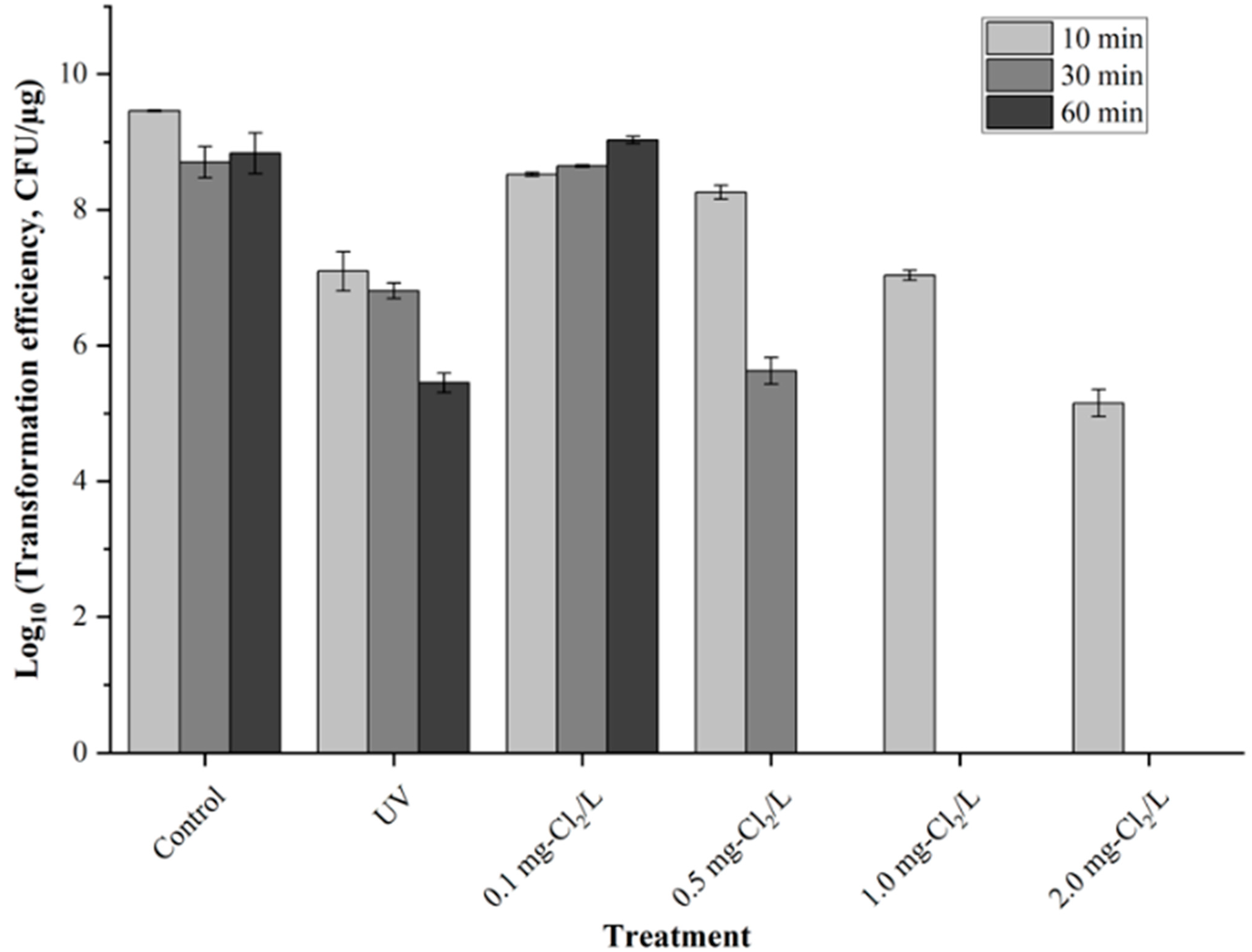
3.1. Reduction in Transformation Efficiency by Chlorine and UV Irradiation
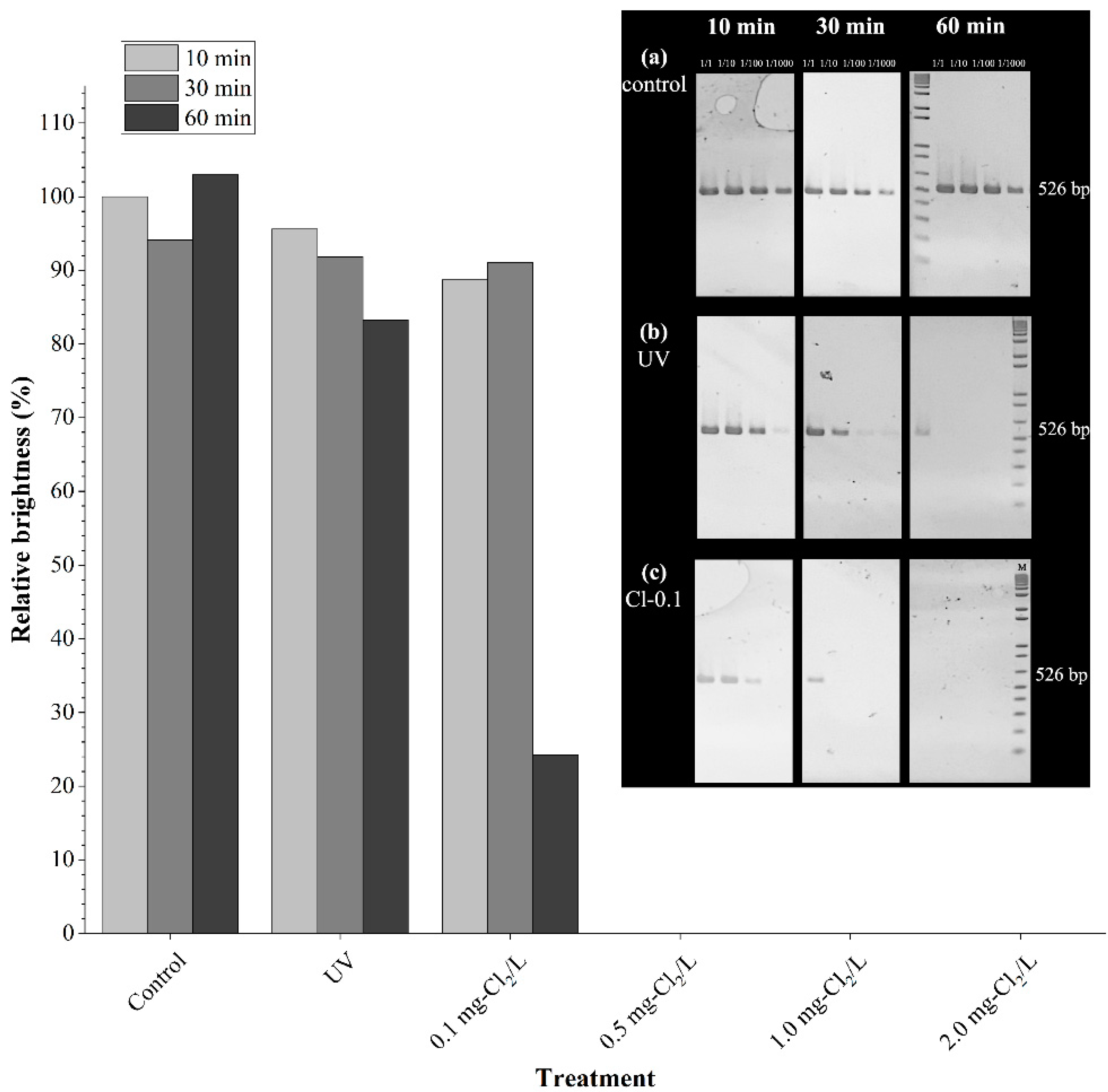
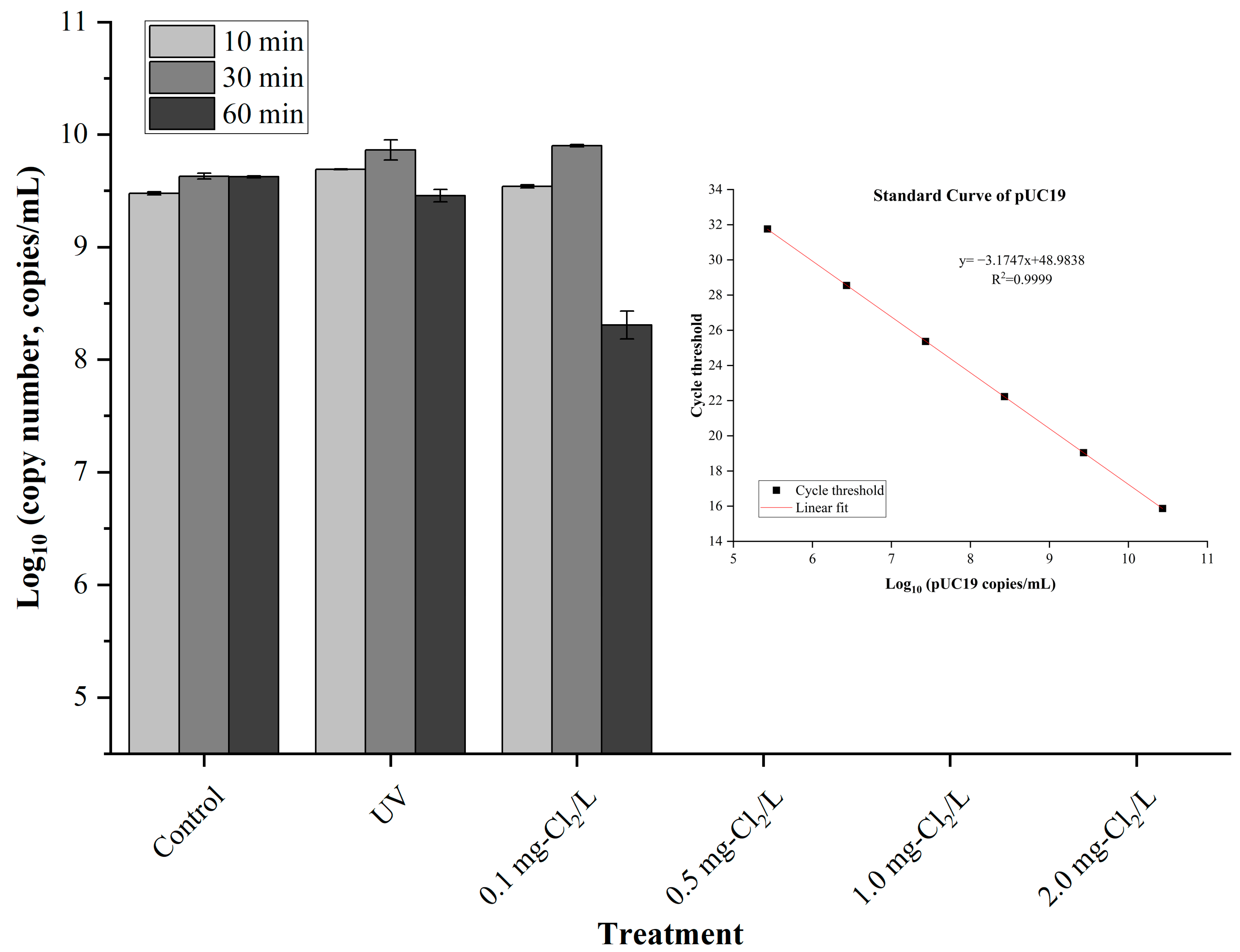
3.2. Detection of Treated pUC19 Using PCR and qPCR
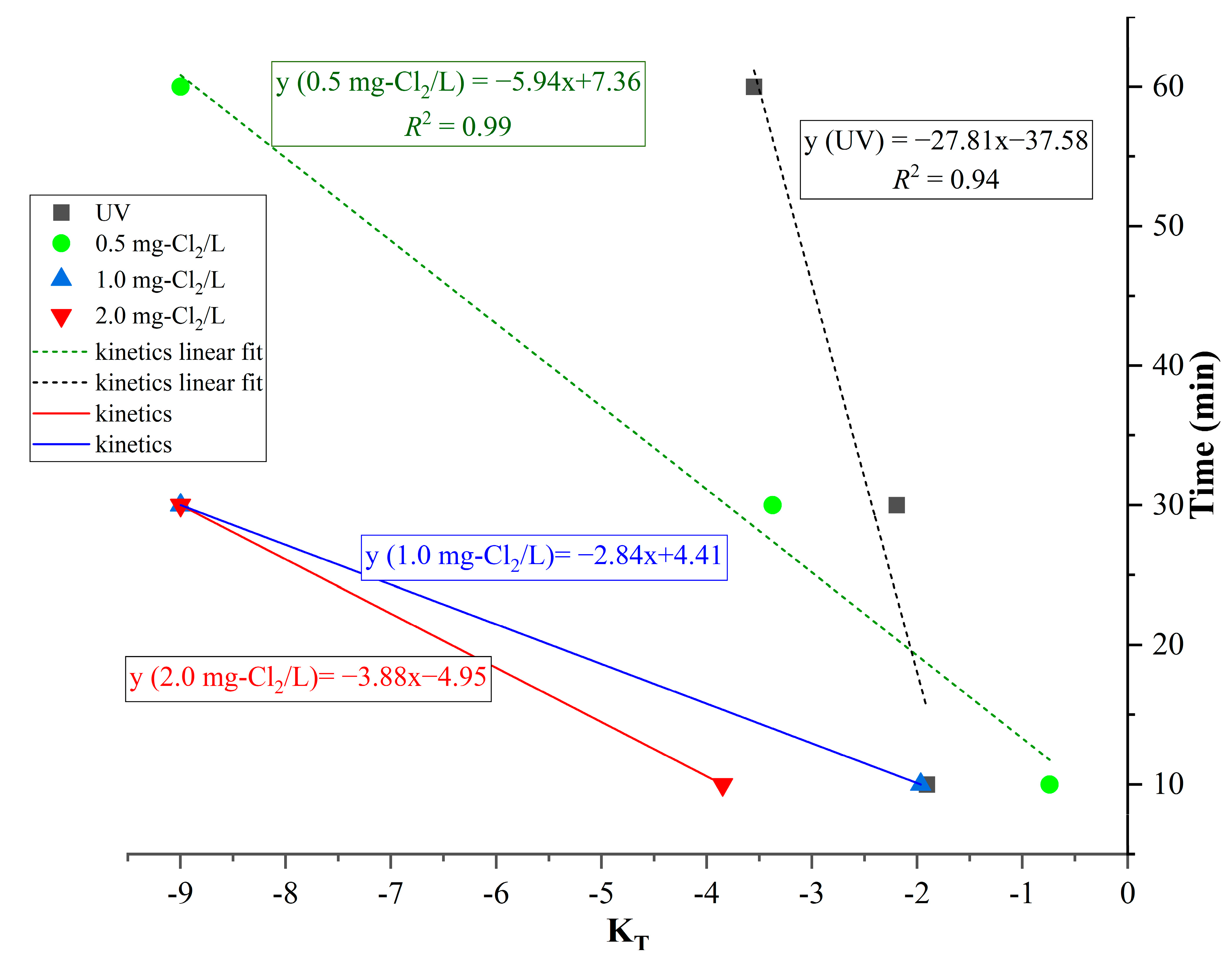
4. Discussion
- transformation efficiencyt: the transformation efficiency after t min of treatment
- transformation efficiency0: the no treatment transformation efficiency
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global antibiotic Consumption 2000–2010: An Analysis of National Pharmaceutical Sales Data. Lancet Infect. Dis. 2014, 14, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Terahara, F.; Nishiura, H. Fluoroquinolone consumption and Escherichia coli resistance in Japan: An ecological study. BMC Public Health 2019, 19, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Pazda, M.; Kumirska, J.; Stepnowski, P.; Mulkiewicz, E. Antibiotic resistance genes identified in wastewater treatment plant systems—A review. Sci. Total Environ. 2019, 697, 134023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, B. Occurrence, transformation, and fate of antibiotics in municipal wastewater treatment plants. Crit. Rev. Environ. Sci. Technol. 2011, 41, 951–998. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, A.; Dai, T.; Li, F.; Xie, H.; Chen, L.; Wen, D. Cell-free DNA: A neglected source for antibiotic resistance genes spreading from WWTPs. Environ. Sci. Technol. 2018, 52, 248–257. [Google Scholar] [CrossRef]
- Collivignarelli, M.C.; Abbà, A.; Benigna, I.; Sorlini, S.; Torretta, V. Overview of the Main Disinfection Processes for Wastewater and Drinking Water Treatment Plants. Sustainability 2018, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, T.; Jikumaru, A.; Ueno, T.; Sei, K. Inactivation effect of antibiotic-resistant gene using chlorine disinfection. Water 2017, 9, 547. [Google Scholar] [CrossRef]
- Wang, L.; Ye, C.; Guo, L.; Chen, C.; Kong, X.; Chen, Y.; Shu, L.; Wang, P.; Yu, X.; Fang, J. Assessment of the UV/chlorine process in the disinfection of Pseudomonas aeruginosa: Efficiency and mechanism. Environ. Sci. Technol. 2021, 55, 9221–9230. [Google Scholar] [CrossRef]
- Liu, L.; Xing, X.; Hu, C.; Wang, H.; Lyu, L. Effect of sequential UV/free chlorine disinfection on opportunistic pathogens and microbial community structure in simulated drinking water distribution systems. Chemosphere 2019, 219, 971–980. [Google Scholar] [CrossRef]
- Zhang, S.; Lin, T.; Chen, H.; Chen, W.; Xu, H.; Tao, H. DNA pyrimidine bases in water: Insights into relative reactivity, by-products formation and combined toxicity during chlorination. Sci. Total Environ. 2020, 717, 137205. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Xiao, R.; Dionysiou, D.D.; Spinney, R.; Fu, T.; Li, Q.; Wang, Z.; Wang, D.; Wei, Z. Kinetics and mechanisms of the formation of chlorinated and oxygenated polycyclic aromatic hydrocarbons during chlorination. Chem. Eng. J. 2018, 351, 248–257. [Google Scholar] [CrossRef]
- Spiliotopoulou, A.; Hansen, K.M.S.; Andersen, H.R. Secondary formation of disinfection by-products by UV treatment of swimming pool water. Sci. Total Environ. 2015, 520, 96–105. [Google Scholar] [CrossRef]
- Tran, J.H.; Jacoby, G.A. Mechanism of plasmid-mediated quinolone resistance. Proc. Natl. Acad. Sci. USA 2002, 99, 5638–5642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Chen, S.; Yu, X.; Vikesland, P.; Pruden, A. Degradation of extracellular genomic, plasmid DNA and specific antibiotic resistance genes by chlorination. Front. Environ. Sci. Eng. 2019, 13, 38. [Google Scholar] [CrossRef]
- Gopal, K.; Tripathy, S.S.; Bersillon, J.L.; Dubey, S.P. Chlorination byproducts, their toxicodynamics and removal from drinking water. J. Hazard. Mater. 2007, 140, 1–6. [Google Scholar] [CrossRef]
- Gibson, J.; Drake, J.; Karney, B. UV disinfection of wastewater and combined sewer overflows. Adv. Exp. Med. Biol. 2017, 996, 267–275. [Google Scholar] [CrossRef]
- McKinney, C.W.; Pruden, A. Ultraviolet disinfection of antibiotic resistant bacteria and their antibiotic resistance genes in water and wastewater. Environ. Sci. Technol. 2012, 46, 13393–13400. [Google Scholar] [CrossRef]
- Zhang, S.; Ye, C.; Lin, H.; Lv, L.; Yu, X. UV disinfection induces a Vbnc State in Escherichia coli and Pseudomonas aeruginosa. Environ. Sci. Technol. 2015, 49, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Umar, M.; Anglès D’auriac, M.; Wennberg, A.C. Application of UV-LEDs for antibiotic resistance genes inactivation—Efficiency monitoring with QPCR and transformation. J. Environ. Chem. Eng. 2021, 9, 105260. [Google Scholar] [CrossRef]
- Komiyama, Y.; Maeda, S. Natural transformation in Escherichia coli. In Horizontal Gene Transfer: Methods and Protocols; de la Cruz, F., Ed.; Springer: New York, NY, USA, 2020; pp. 179–187. ISBN 978-1-4939-9877-7. [Google Scholar] [CrossRef]
- Sun, D. Two different routes for double-stranded DNA transfer in natural and artificial transformation of Escherichia coli. Biochem. Biophys. Res. Commun. 2016, 471, 213–218. [Google Scholar] [CrossRef]
- Sorinolu, A.J.; Munir, M. Degradation and horizontal gene transfer analysis of plasmid-encoded antibiotic resistance genes during UV254, hydroxyl radical and sulphate radical treatments. Chem. Eng. J. 2022, 450, 138380. [Google Scholar] [CrossRef]
- Yadav, V.K.; Awasthi, P.; Kumar, A. Detection of UV-induced thymine dimers. In Genotoxicity Assessment: Methods and Protocols; Dhawan, A., Bajpayee, M., Eds.; Springer: New York, NY, USA, 2019; pp. 313–322. ISBN 978-1-4939-9646-9. [Google Scholar] [CrossRef]
- Ding, W.; Jin, W.; Cao, S.; Zhou, X.; Wang, C.; Jiang, Q.; Huang, H.; Tu, R.; Han, S.F.; Wang, Q. Ozone disinfection of chlorine-resistant bacteria in drinking water. Water Res. 2019, 160, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Venieri, D.; Mantzavinos, D. Disinfection of waters/wastewaters by solar photocatalysis. In Advances in Photocatalytic Disinfection; An, T., Zhao, H., Wong, P.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 177–198. ISBN 978-3-662-53496-0. [Google Scholar] [CrossRef]
- Wan, K.; Zheng, S.; Ye, C.; Hu, D.; Zhang, Y.; Dao, H.; Chen, S.; Yu, X. Ancient oriental wisdom still works: Removing ARGs in drinking water by boiling as compared to chlorination. Water Res. 2021, 209, 117902. [Google Scholar] [CrossRef]
- Van Aken, B.; Lin, L.S. Effect of the disinfection agents chlorine, UV irradiation, silver ions, and TiO2 nanoparticles/near-UV on DNA molecules. Water Sci. Technol. 2011, 64, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, A.M.; Garcia, L.A.T.; Silva, K.J.S.; Sabogal-Paz, L.P.; Hincapié, M.M.; Montoya, L.J.; Galeano, L.; Galdos-Balzategui, A.; Reygadas, F.; Herrera, C.; et al. Chlorination for low-cost household water disinfection—A critical review and status in three Latin American countries. Int. J. Hyg. Environ. Health 2022, 244, 114004. [Google Scholar] [CrossRef]
- Ibrahim, M.A.S.; MacAdam, J.; Autin, O.; Jefferson, B. Evaluating the impact of LED bulb development on the economic viability of ultraviolet technology for disinfection. Environ. Technol. 2014, 35, 400–406. [Google Scholar] [CrossRef]
- Kim, S.; Masai, S.; Murakami, K.; Azuma, M.; Kataoka, K.; Sebe, M.; Shimizu, K.; Itayama, T.; Whangchai, N.; Whangchai, K.; et al. Characteristics of antibiotic resistance and tolerance of environmentally endemic Pseudomonas aeruginosa. Antibiotics 2022, 11, 1120. [Google Scholar] [CrossRef] [PubMed]
- Haaken, D.; Schmalz, V.; Dittmar, T.; Worch, E. Limits of UV disinfection: UV/electrolysis hybrid technology as a promising alternative for direct reuse of biologically treated wastewater. J. Water Supply Res. Technol. 2013, 62, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Chen, Z.; Chen, X.; Huang, P.; Chen, X.; Ding, R.; Liu, J.; Chen, R. The drinking water disinfection performances and mechanisms of UVA-LEDs promoted by electrolysis. J. Hazard. Mater. 2022, 435, 129099. [Google Scholar] [CrossRef]
- Stedtfeld, R.D.; Williams, M.R.; Fakher, U.; Johnson, T.A.; Stedtfeld, T.M.; Wang, F.; Khalife, W.T.; Hughes, M.; Etchebarne, B.E.; Tiedje, J.M.; et al. Antimicrobial resistance dashboard application for mapping environmental occurrence and resistant pathogens. FEMS Microbiol. Ecol. 2016, 92, fiw020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamata, K.; Tokuda, Y.; Gu, Y.; Ohmagari, N.; Yanagihara, K. Public knowledge and perception about antimicrobials and antimicrobial resistance in Japan: A national questionnaire survey in 2017. PLoS ONE 2018, 13, e0207017. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Reference |
|---|---|---|
| pUC19amp121F | 5′-CGAGTGGGTTACATCGAACTG-3′ | This study |
| pUC19amp646R | 5′-CTGCAACTTTATCCGCCTCC-3′ | This study |
| pUC19amp587F | 5′-ACTTACTCTAGCTTCCCGGC-3′ | This study |
| Disinfection Method | Processing Time Required for Transformation Efficiency Reduction 2-log10 |
|---|---|
| UV (40 µW/cm2) | 18.04 min |
| 0.1 mg-Cl2/L | NA |
| 0.5 mg-Cl2/L | 19.25 min |
| 1.0 mg-Cl2/L | 10.10 min |
| 2.0 mg-Cl2/L | 2.82 min |
| Amplicon or DNA | Dimers (Counts) | Dimers/Amplicon Length (%) | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | CT | TC | TT | Total | CC | CT | TC | TT | Total | ||
| 60 bp * | 5 | 5 | 4 | 4 | 18 | 8.3 | 8.3 | 6.7 | 6.7 | 30.0 | This study |
| tetA (210 bp) * | 37 | 26 | 30 | 12 | 105 | 17.6 | 12.4 | 14.3 | 5.7 | 50.0 | [21] |
| tetA (1054 bp) * | 185 | 104 | 135 | 81 | 505 | 17.6 | 9.9 | 12.8 | 7.7 | 47.9 | |
| ampR (192 bp) * | 22 | 21 | 26 | 42 | 111 | 11.5 | 10.9 | 13.5 | 21.9 | 57.8 | |
| ampR (851 bp) * | 87 | 104 | 105 | 118 | 414 | 10.2 | 12.2 | 12.3 | 13.9 | 48.6 | |
| 80 bp * | 9 | 8 | 14 | 6 | 37 | 11.3 | 10.0 | 17.5 | 7.5 | 46.3 | [18] |
| 601 bp * | 61 | 74 | 84 | 97 | 316 | 10.2 | 12.3 | 14.0 | 16.1 | 52.6 | |
| 526 bp ** | 24 | 32 | 43 | 36 | 135 | 4.6 | 6.1 | 8.2 | 6.8 | 25.7 | This study |
| pUC19 (2686 bp) | 130 | 163 | 179 | 139 | 611 | 4.8 | 6.1 | 6.7 | 5.2 | 22.7 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Miao, H.; Lei, Z.; Yuan, T.; Zhang, Z.; Ihara, I.; Maseda, H.; Shimizu, K. Decreased Efficiency of Free Naked DNA Transformation by Chlorine and UV Disinfection and Its Detection Limitations. Water 2023, 15, 1232. https://doi.org/10.3390/w15061232
Zhang C, Miao H, Lei Z, Yuan T, Zhang Z, Ihara I, Maseda H, Shimizu K. Decreased Efficiency of Free Naked DNA Transformation by Chlorine and UV Disinfection and Its Detection Limitations. Water. 2023; 15(6):1232. https://doi.org/10.3390/w15061232
Chicago/Turabian StyleZhang, Chi, Hanchen Miao, Zhongfang Lei, Tian Yuan, Zhenya Zhang, Ikko Ihara, Hideaki Maseda, and Kazuya Shimizu. 2023. "Decreased Efficiency of Free Naked DNA Transformation by Chlorine and UV Disinfection and Its Detection Limitations" Water 15, no. 6: 1232. https://doi.org/10.3390/w15061232