
Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Isolate
2.2. Molecular Identification
2.3. Impact of Various Nutrient Media on the Growth Potential of Oscillatoria sancta
2.4. Modification of the Chemical Composition of Zarrouk’s Medium
2.5. Growth Assessment
2.6. Growth Rate
2.7. Biomass Harvesting
2.8. Biochemical Properties
2.9. Crude Extraction with Methylene Chloride/Methanol (1:1) v/v
2.10. Biological Activities of Oscillatoria sancta Crude Extract
2.10.1. Antibacterial Screening Assay
2.10.2. In Vitro Acetylcholinesterase Activity Test
2.10.3. Evaluation of Anticancer Activity
Cell Line
In Vitro MTT Assay
2.11. GC-MS Analysis
2.12. Statistical Analysis
3. Results
3.1. Phylogenetic Analysis and Placement of Oscillatoria sancta
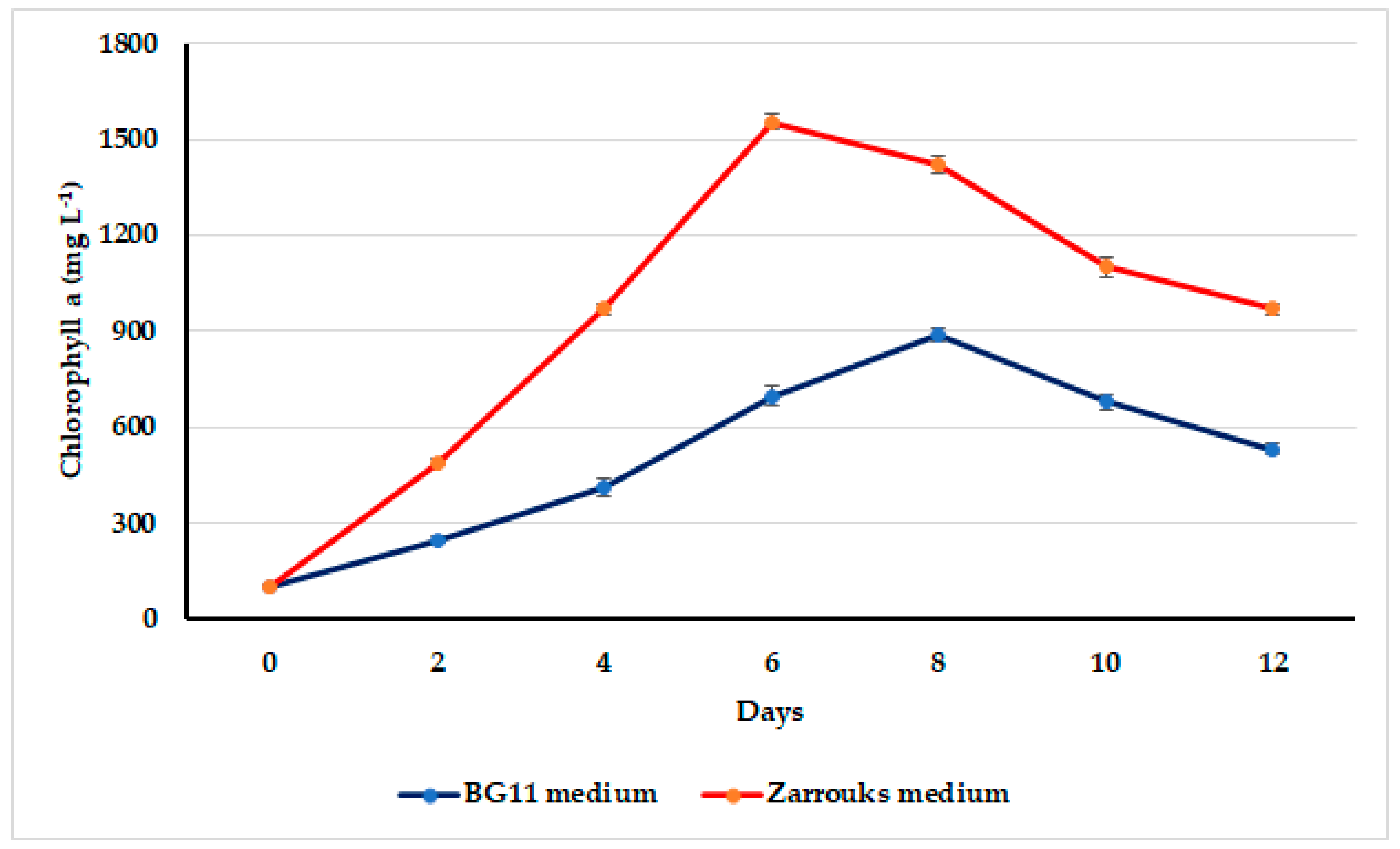
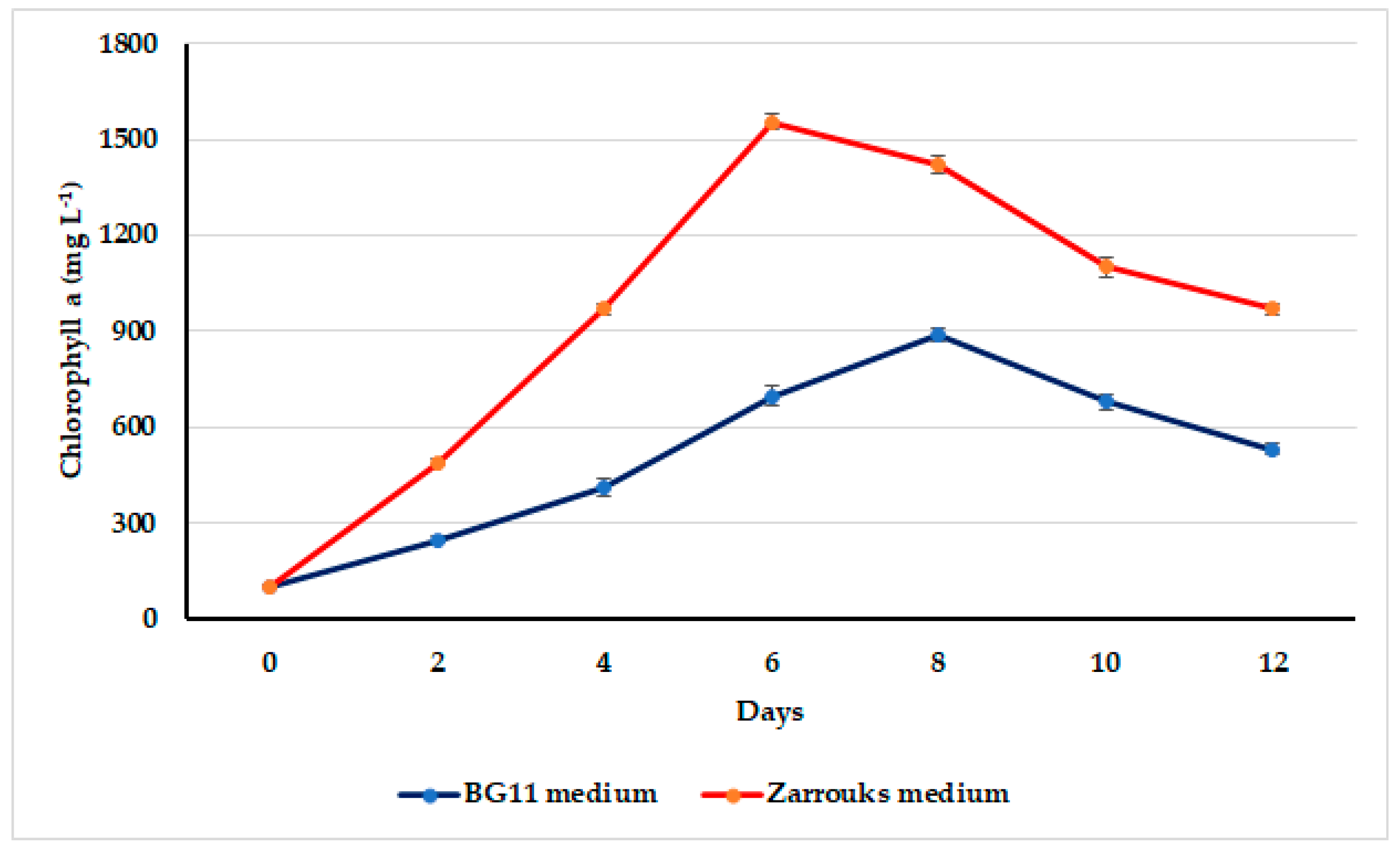
3.2. Growth Assessment of Oscillatoria sancta on Different Nutrient Media
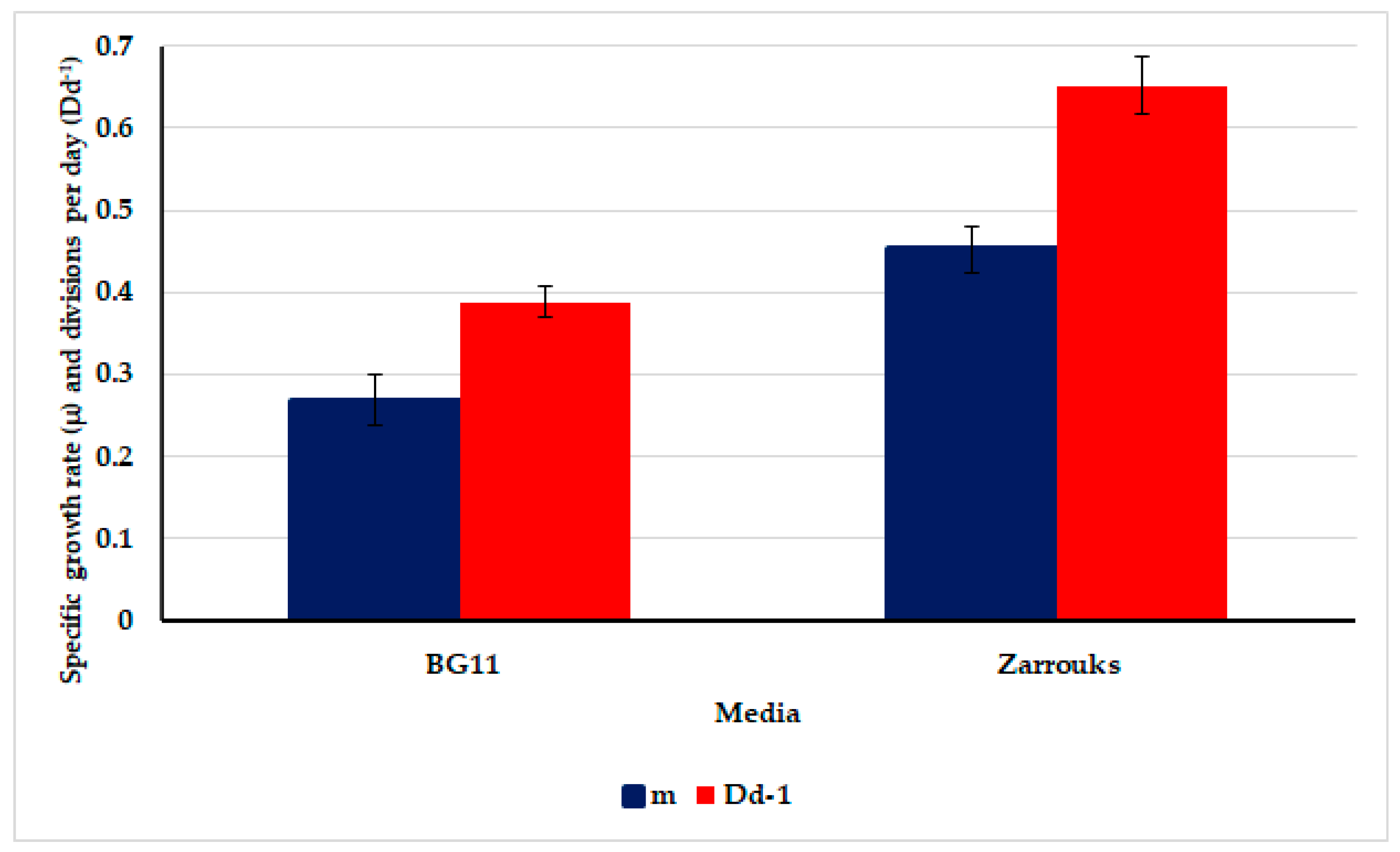
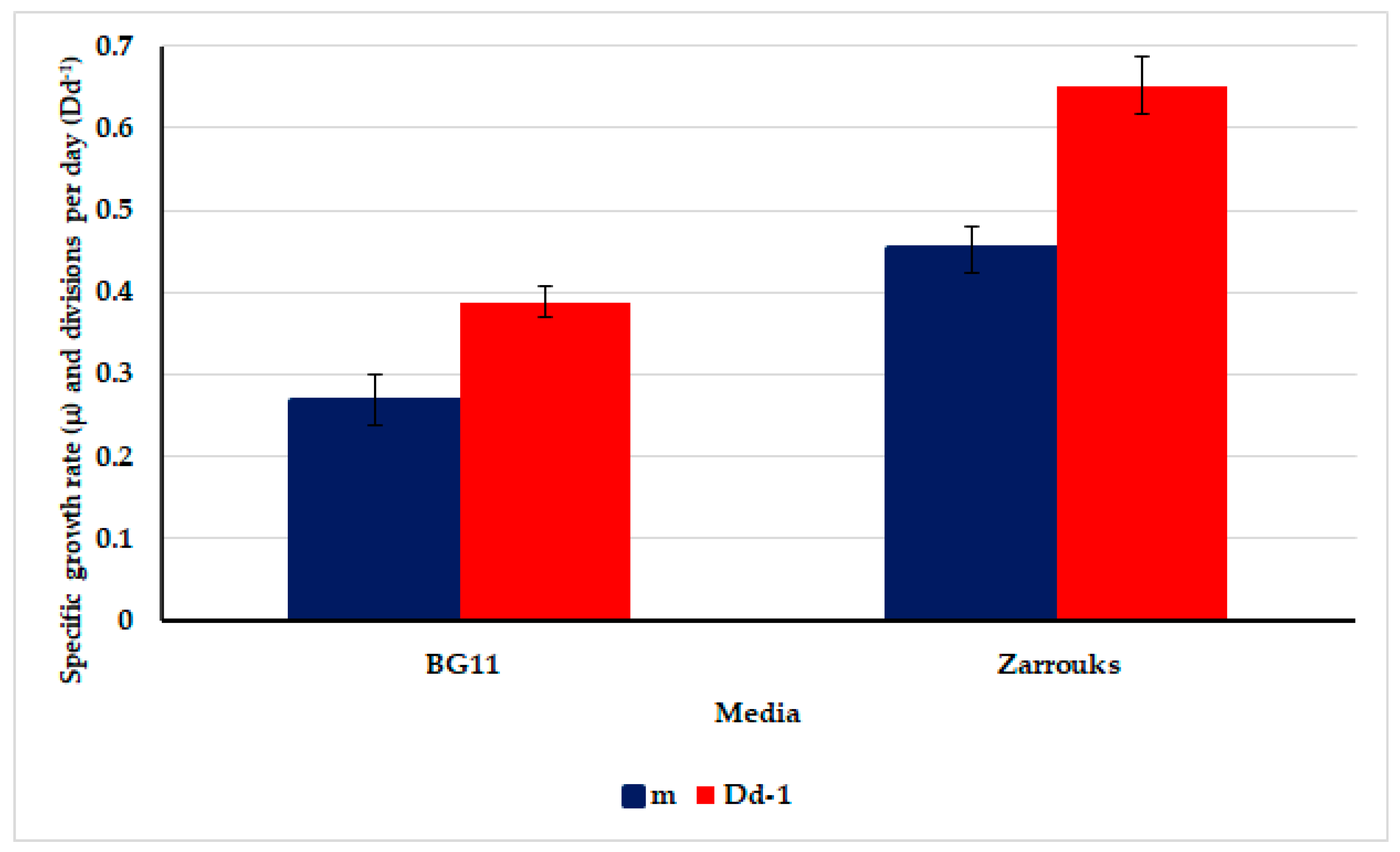
3.3. Growth Rates of the Test Cyanobacterium Oscillatoria sancta
3.4. Oscillatoria sancta Growth in Modified Zarrouk’s Medium at Varying Nutrient Concentrations
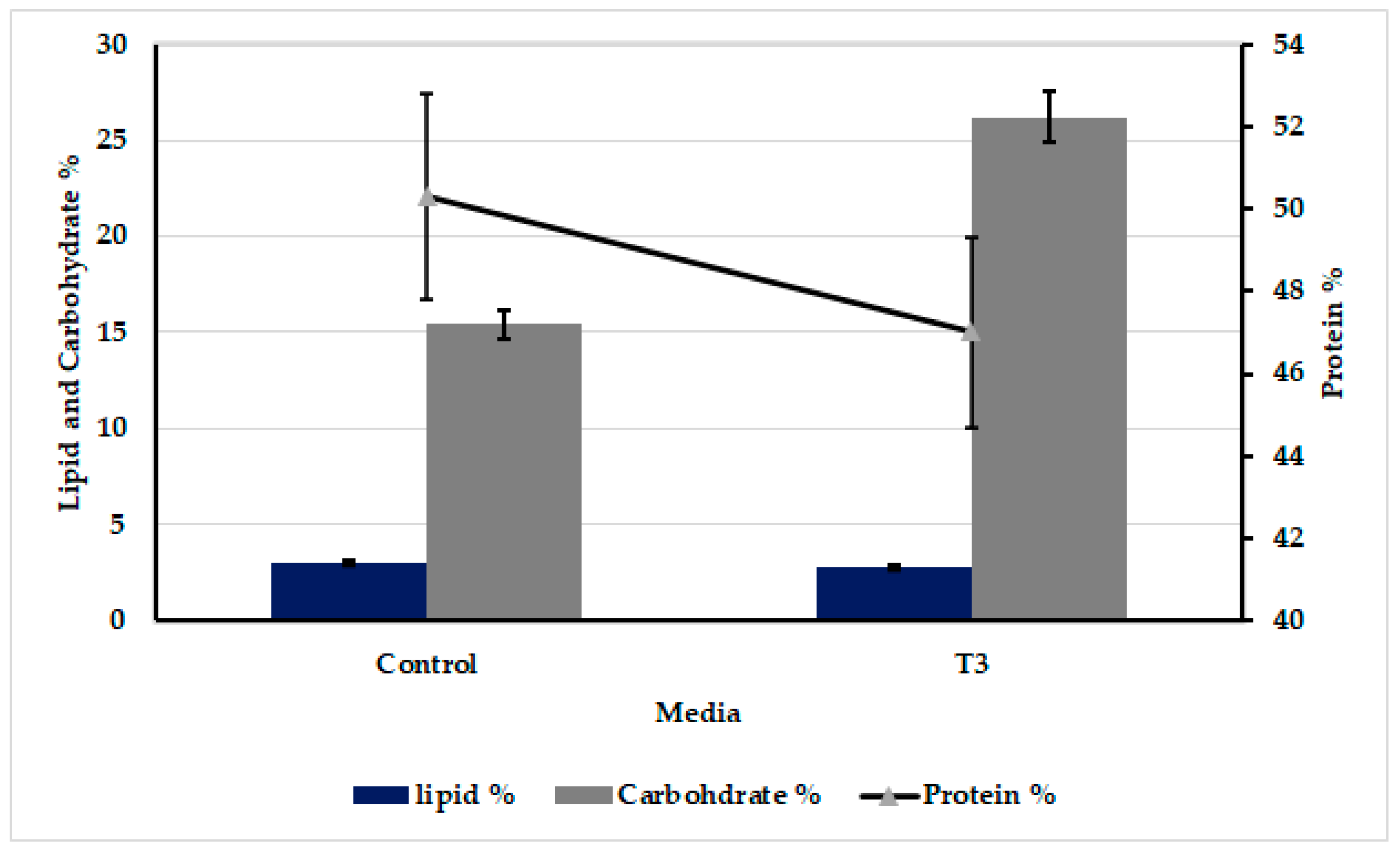
3.5. Biochemical Composition of Oscillatoria sancta Grown on Both 12.5% Modified Zarrouk’s Medium and Standard Zarrouk’s Medium
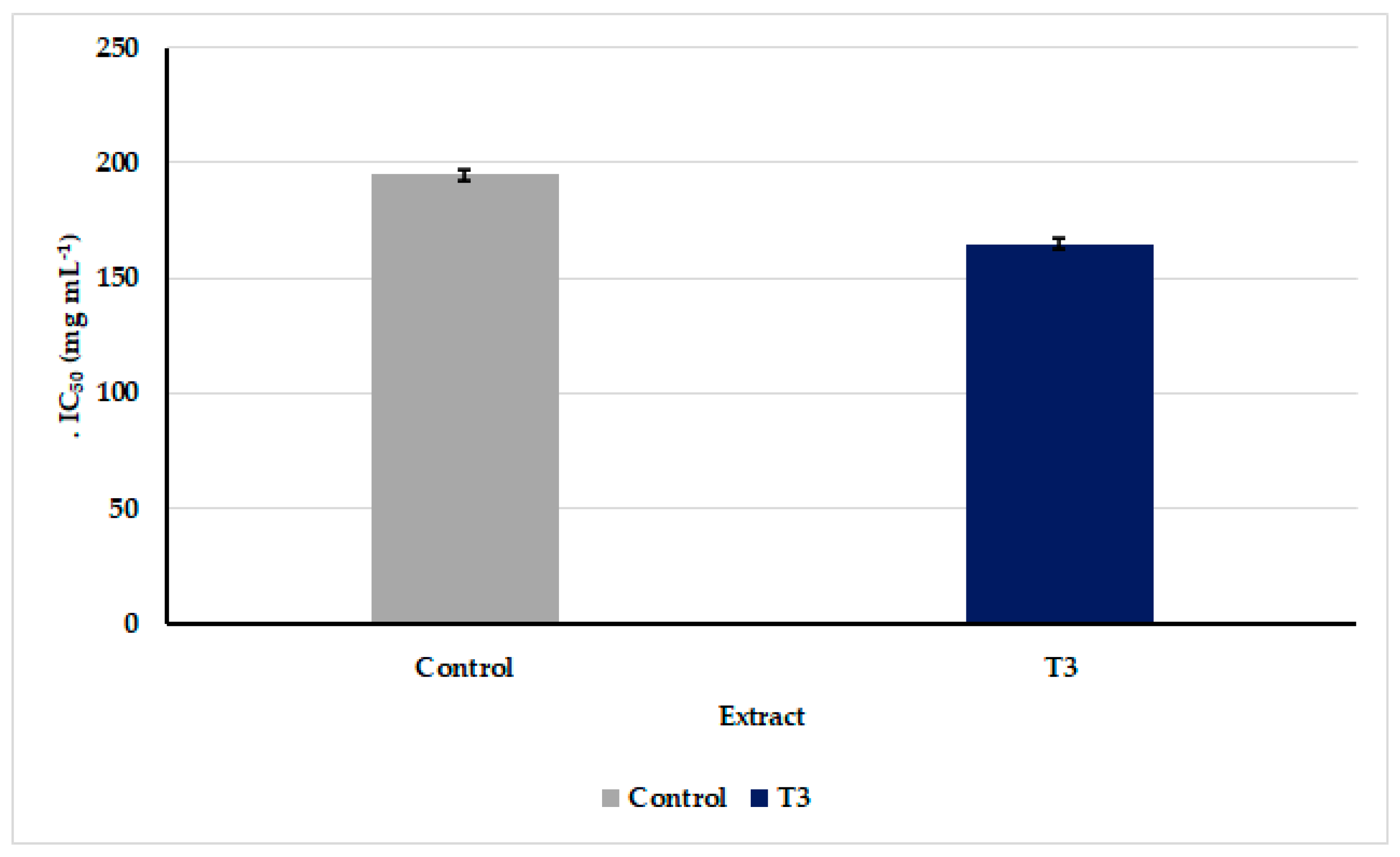
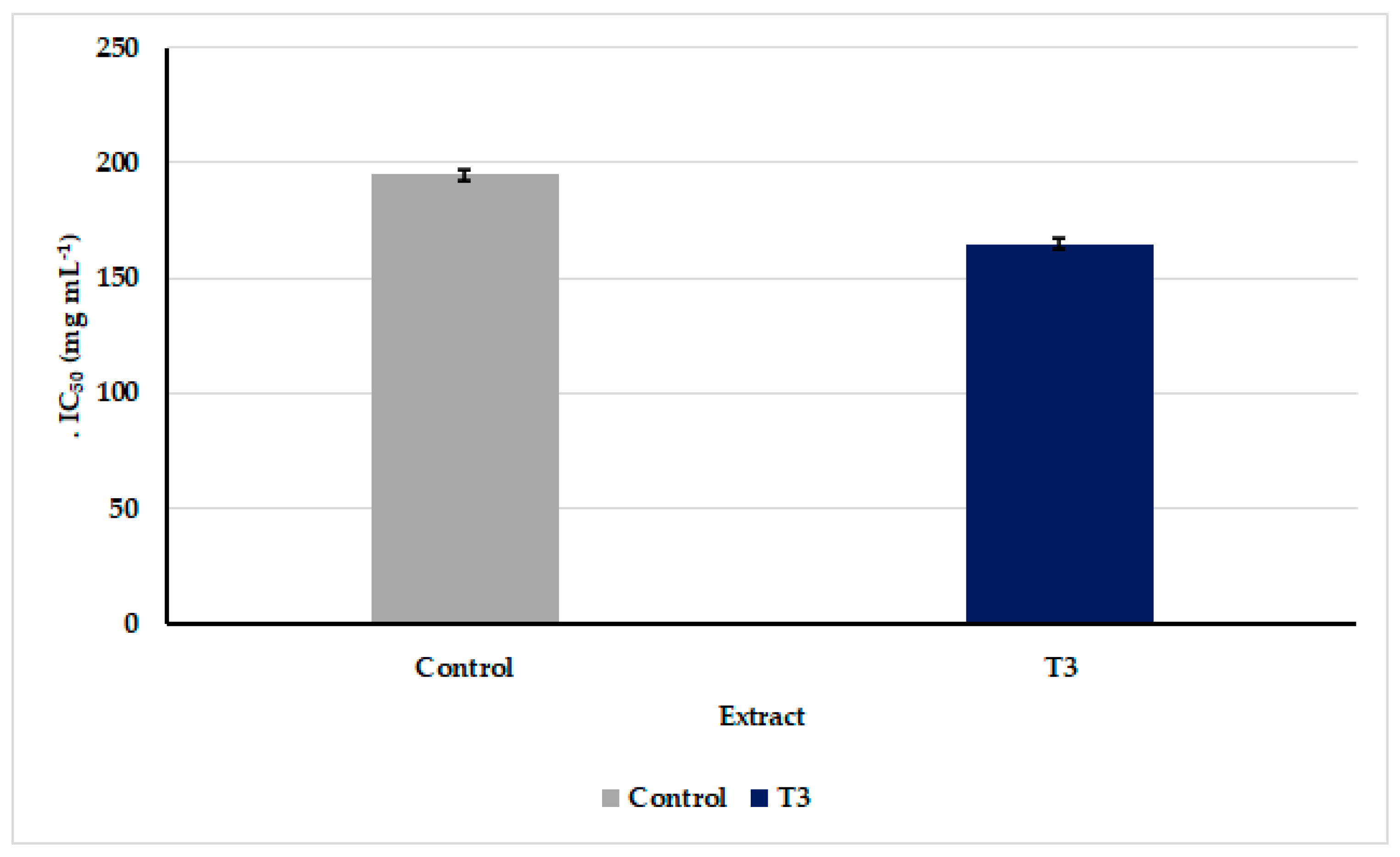
3.6. Anticancer Activity of Oscillatoria sancta Methylene Chloride/Methanol Crude Extracts against MDA-MB-231 Cell Line
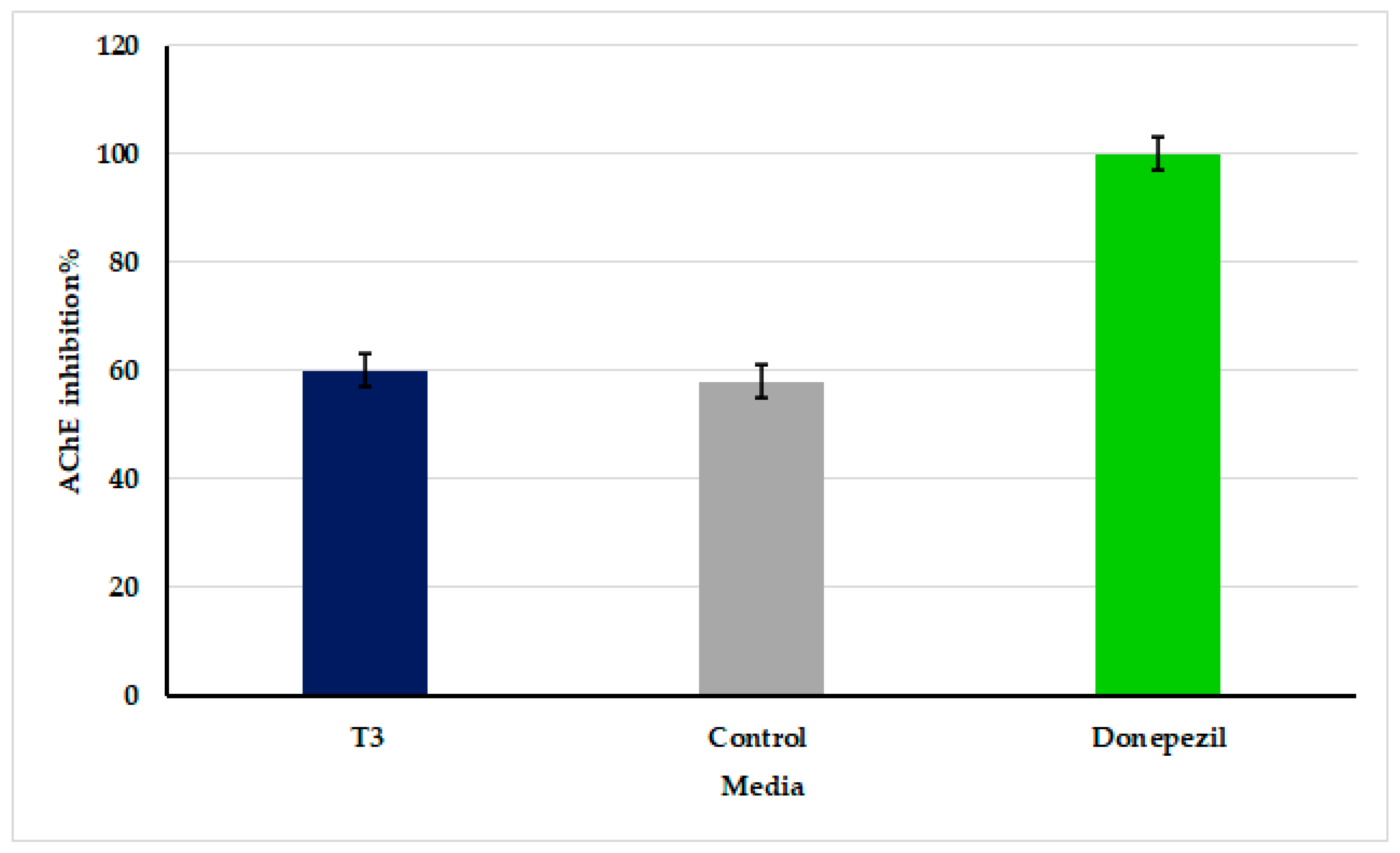
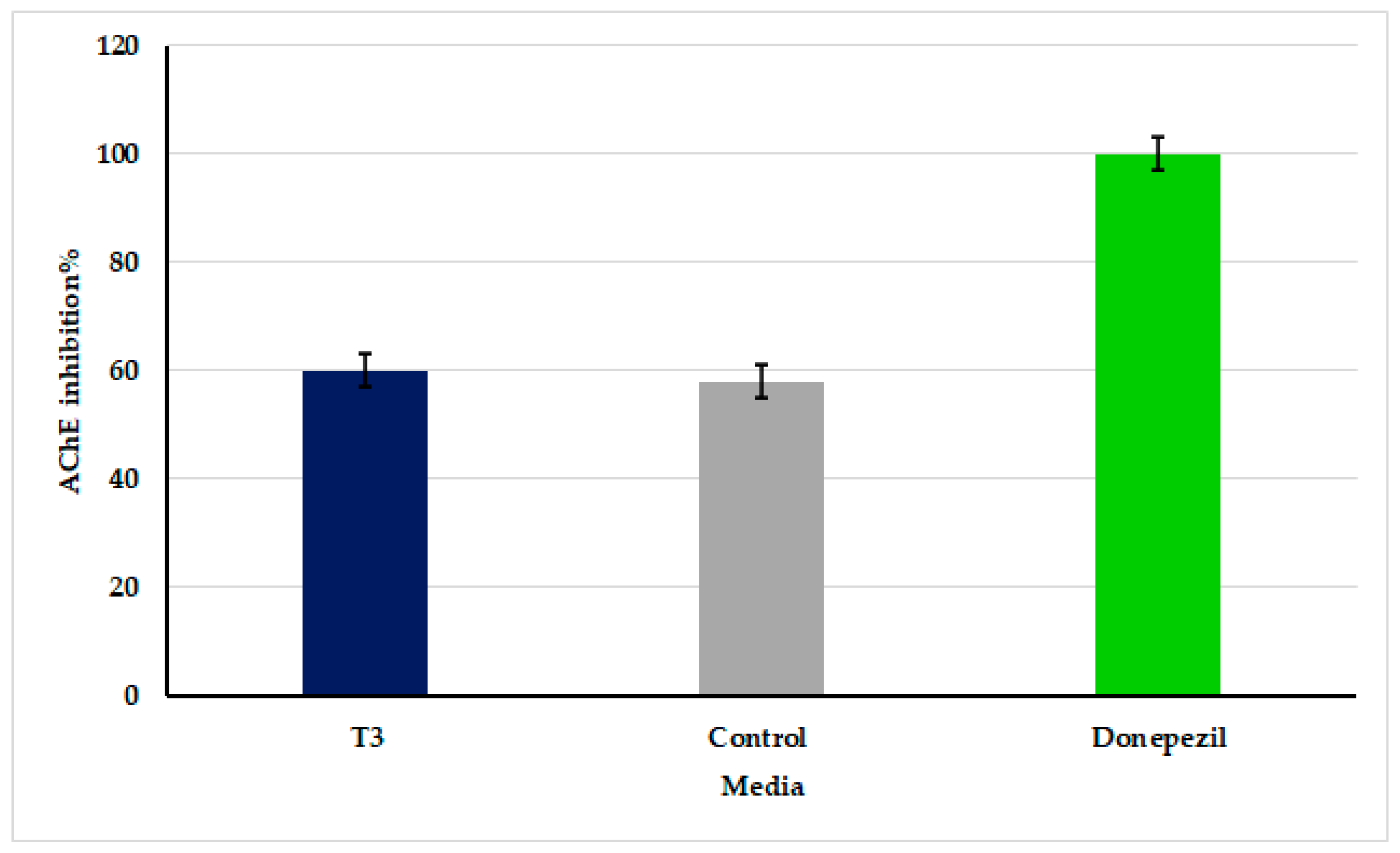
3.7. Inhibition of Acetylcholinesterase Activity by Oscillatoria sancta Crude Extracts
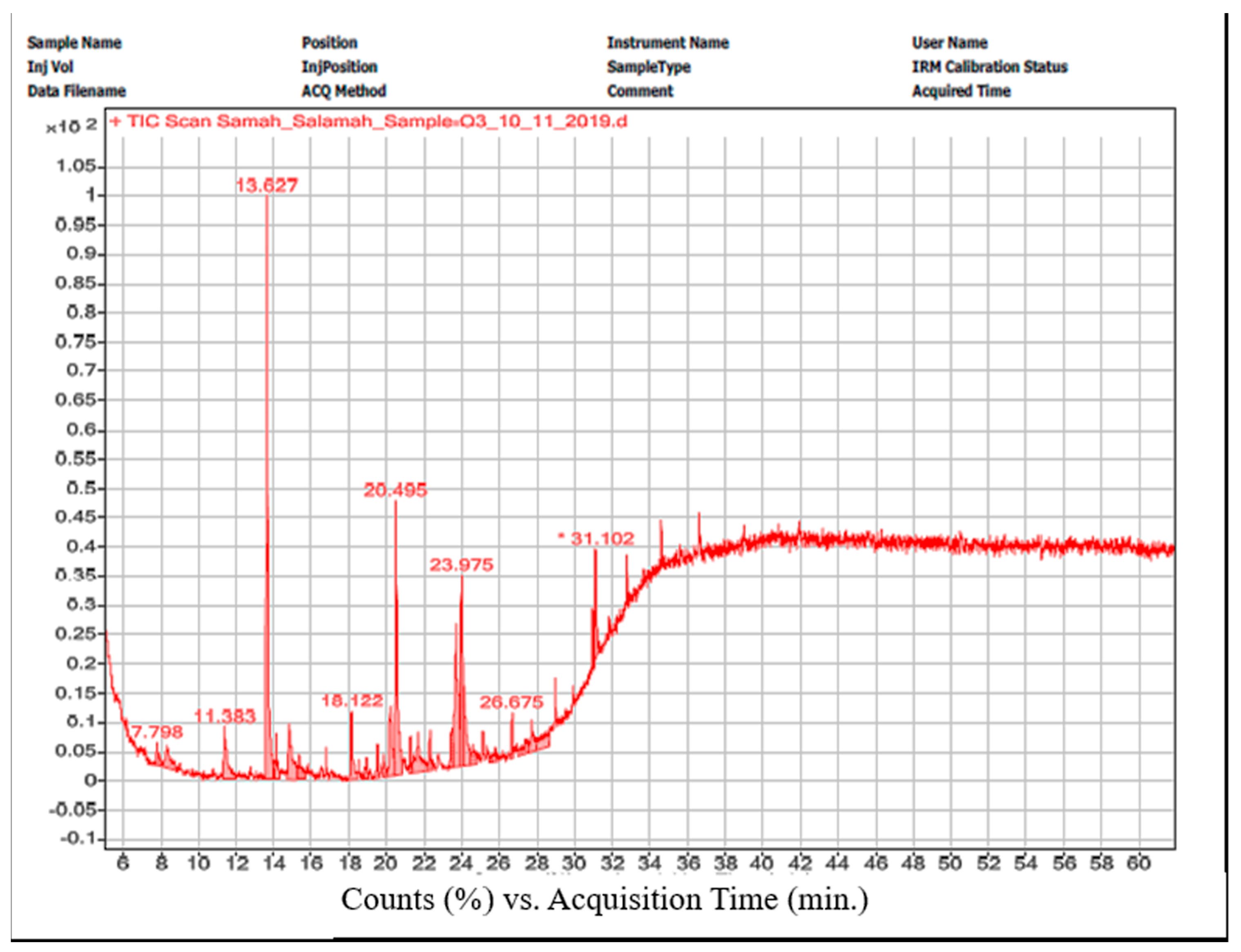
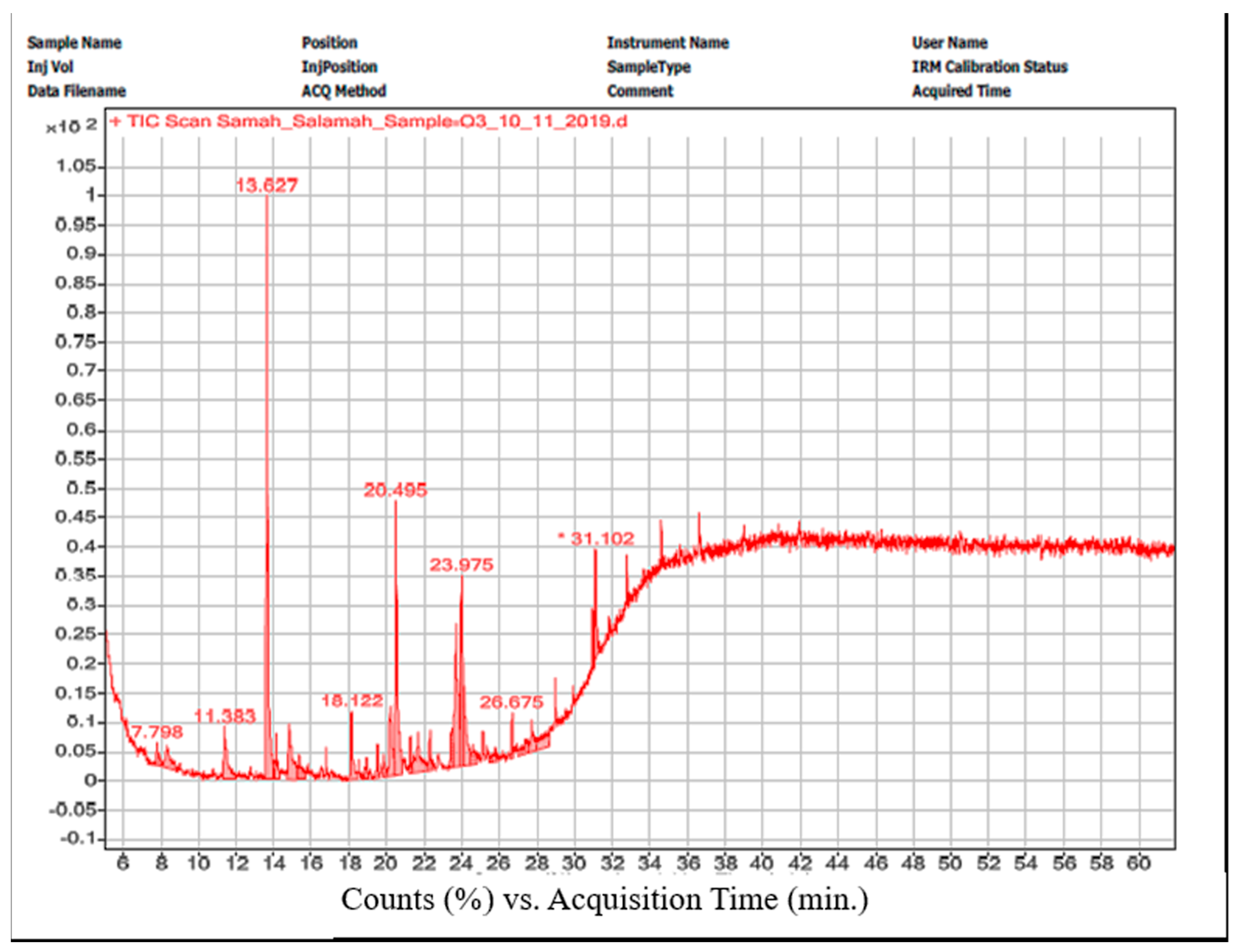
3.8. GC/MS
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sijil, P.V.; Cherita, C.; Jethani, H.; Chauhan, V.S. Chapter 1. Microalgae as a renewable and sustainable source of high value metabolites. In Microalgae for Sustainable Products; The Royal Society of Chemistry: London, UK, 2022; pp. 1–26. [Google Scholar]
- Burnap, R.L. Systems and photosystems: Cellular limits of autotrophic productivity in cyanobacteria. Front. Bioeng. Biotechnol. 2015, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hua, Q.; Shimizu, K. Metabolic flux analysis in Synechocystis using isotope distribution from 13c-labeled glucose. Metab. Eng. 2002, 4, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Ambrosi, M.; Reinehr, C.; Bertolin, T.; Costa, J.A.; Colla, L. Propriedades de saúde de Spirulina spp. Rev. Ciências Farm. Básica Apl. 2008, 29, 109–117. [Google Scholar]
- Schwarz, R.; Forchhammer, K. Acclimation of unicellular cyanobacteria to macronutrient deficiency: Emergence of a complex network of cellular responses. Microbiology 2005, 151, 2503–2514. [Google Scholar] [CrossRef]
- Romay, C.; González, R.; Ledón, N.; Remirez, D.; Rimbau, V. C-phycocyanin: A biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef]
- Mogany, T.; Swalaha, F.M.; Kumari, S.; Bux, F. Elucidating the role of nutrients in c-phycocyanin production by the halophilic cyanobacterium euhalothece sp. J. Appl. Phycol. 2018, 30, 2259–2271. [Google Scholar] [CrossRef]
- Tan, L.T. Filamentous tropical marine cyanobacteria: A rich source of natural products for anticancer drug discovery. J. Appl. Phycol. 2010, 22, 659–676. [Google Scholar] [CrossRef]
- Ahmed, W.A.; El-Semary, N.A.; Abd El-Hameed, O.M.; El Tawill, G.; Ibrahim, D.M. Bioactivity and cytotoxic effect of cyanobacterial toxin against hepatocellular carcinoma. J. Cancer Sci. Ther. 2017, 9, 505–511. [Google Scholar]
- Pelizer, L.H.; Danesi, E.D.G.; de O Rangel, C.; Sassano, C.E.; Carvalho, J.C.M.; Sato, S.; Moraes, I.O. Influence of inoculum age and concentration in Spirulina platensis cultivation. J. Food Eng. 2003, 56, 371–375. [Google Scholar] [CrossRef]
- Peter, P.; Sarma, A.P.; Azeem ul Hasan, M.; Murthy, S.D.D.; Attitalla, I.H. Studies on the impact of nitrogen sarma starvation on the photosynthetic pigments through spectral properties of the cyanobacterium, Spirulina platensis: Identification of target phycobiliprotein under nitrogen chlorosis. Bot. Res. Int. 2010, 3, 30–34. [Google Scholar]
- Hai, T.; Hein, S.; Steinbuchel, A. Multiple evidence for widespread and general occurrence of type-iii pha synthases in cyanobacteria and molecular characterization of the pha synthases from two thermophilic cyanobacteria: Chlorogloeopsis fritschii pcc 6912 and synechococcus sp. Strain ma19. Microbiology 2001, 147, 3047–3060. [Google Scholar] [CrossRef]
- El-Kassas, H.Y.; El-Sheekh, M.M. Cytotoxic activity of biosynthesized gold nanoparticles with an extract of the red seaweed corallina officinalis on the mcf-7 human breast cancer cell line. Asian Pac. J. Cancer Prev. 2014, 15, 4311–4317. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; El-Kassas, H.Y. Application of biosynthesized silver nanoparticles against a cancer promoter cyanobacterium, microcystis aeruginosa. Asian Pac. J. Cancer Prev. 2014, 15, 6773–6779. [Google Scholar] [CrossRef] [Green Version]
- El-Sheekh, M.M.; El-Kassas, H.Y. Algal production of nano-silver and gold: Their antimicrobial and cytotoxic activities: A review. J. Genet. Eng. Biotechnol. 2016, 14, 299–310. [Google Scholar] [CrossRef]
- Gheda, S.; El-Sheekh, M.; Abou-Zeid, A. In vitro anticancer activity of polysaccharide extracted from the red alga Jania rubens against breast and colon cancer cell lines. Asian Pac. J. Trop. Med. 2018, 11, 583–589. [Google Scholar]
- Gademann, K.; Portmann, C. Secondary metabolites from cyanobacteria: Complex structures and powerful bioactivities. Curr. Org. Chem. 2008, 12, 326–341. [Google Scholar] [CrossRef]
- LewisOscar, F.; Nithya, C.; Alharbi, S.A.; Alharbi, N.S.; Thajuddin, N. Microfouling inhibition of human nosocomial pathogen Pseudomonas aeruginosa using marine cyanobacteria. Microb. Pathog. 2018, 114, 107–115. [Google Scholar] [CrossRef]
- Volk, R.-B. Screening of microalgal culture media for the presence of algicidal compounds and isolation and identification of two bioactive metabolites, excreted by the cyanobacteria Nostoc insulare and Nodularia harveyana. J. Appl. Phycol. 2005, 17, 339–347. [Google Scholar] [CrossRef]
- Zelík, P.; Lukešová, A.; Čejka, J.; Buděšínský, M.; Havlíček, V.; Čegan, A.; Kopecký, J. Nostotrebin 6, a bis(cyclopentenedione) with cholinesterase inhibitory activity isolated from nostoc sp. Str. Lukešová 27/97. J. Enzym. Inhib. Med. Chem. 2010, 25, 414–420. [Google Scholar] [CrossRef]
- Becher, P.G.; Baumann, H.I.; Gademann, K.; Jüttner, F. The cyanobacterial alkaloid nostocarboline: An inhibitor of acetylcholinesterase and trypsin. J. Appl. Phycol. 2009, 21, 103–110. [Google Scholar] [CrossRef]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural product biosynthetiAc diversity and comparative genomics of the cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Krunic, A.; Chlipala, G.; Orjala, J. Antimicrobial ambiguine isonitriles from the cyanobacterium Fischerella ambigua. J. Nat. Prod. 2009, 72, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Karan, T.; Aydin, A. Anticancer potential and cytotoxic effect of some freshwater cyanobacteria. Trop. J. Pharm. Res. 2018, 17, 2183–2188. [Google Scholar] [CrossRef]
- Sithranga Boopathy, N.; Kathiresan, K. Anticancer drugs from marine flora: An overview. J. Oncol. 2010, 2010, 214186. [Google Scholar] [CrossRef]
- Robles-Banuelos, B.; Duran-Riveroll, L.M.; Rangel-Lopez, E.; Perez-Lopez, H.I.; Gonzalez-Maya, L. Marine cyanobacteria as sources of lead anticancer compounds: A review of families of metabolites with cytotoxic, antiproliferative, and antineoplastic effects. Molecules 2022, 27, 4814. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Tiwari, B.S. Cyanotherapeutics: An emerging field for future drug discovery. Appl. Phycol. 2020, 1, 44–57. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Smoker, J.A.; Susan, R.B. Rapid small-scale DNA isolation from filamentous cyanobacteria. FEMS Microbiol. Lett. 1988, 56, 119–122. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zarrouk, C. Contribution a l’etude d’une Cyanophycee. Influence de Divers Facteurs Physiques et Chimiques sur la Croissance et la Photosynthese de Spirulina maxima. Ph.D. Thesis, Université De Paris, Paris, France, 1966. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association, American Water Works Association and Water Environment Federation: Washington, DC, USA, 2008. [Google Scholar]
- Andersen, R.A. Algal Culturing Techniques; Elsevier Academic Press: Birlington, MA, USA; San Diego, CA, USA; London, UK, 2005. [Google Scholar]
- Stein, J.R.; Hellebust, J.A.; Craigie, J. Handbook of Phycological Methods: Culture Methods and Growth Measurements; Cambridge University Press: New York, NY, USA, 1973; 448p. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Stoscheck, C.M. [6] Quantitation of protein. In Methods in Enzymology; Deutscher, M.P., Ed.; Academic Press: Cambridge, MA, USA, 1990; Volume 182, pp. 50–68. [Google Scholar]
- Hedge, J.E.; Hofreiter, B.T. Carbohydrate Chemistry; Whistler, R.L., Be Miller, J.N., Eds.; Academic Press: New York, NY, USA, 1962; Volume 17. [Google Scholar]
- Sadasivam, S.; Manickam, A. Biochemical Methods; New Age International (P) Limited: New Delhi, India, 1996; Volume 2, pp. 124–126. [Google Scholar]
- Perez, C. Antibiotic assay by agar-well diffusion method. Acta Biol. Med. Exp. 1990, 15, 113–115. [Google Scholar]
- Refaay, D.A.; Abdel-Hamid, M.I.; Alyamani, A.A.; Abdel Mougib, M.; Ahmed, D.M.; Negm, A.; Mowafy, A.M.; Ibrahim, A.A.; Mahmoud, R.M. Growth Optimization and Secondary Metabolites Evaluation of Anabaena variabilis for Acetylcholinesterase Inhibition Activity. Plants 2022, 11, 735. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hong, S.-J.; Lee, C.G. Statistical optimization of culture media for production of phycobiliprotein by Synechocystis sp. Pcc 6701. Biotechnol. Bioprocess Eng. 2008, 13, 491–498. [Google Scholar] [CrossRef]
- Grobbelaar, J. Factors governing algal growth in photobioreactors: The “open” versus “closed” debate. J. Appl. Phycol. 2009, 21, 489–492. [Google Scholar] [CrossRef]
- Schwartz, W.; Fogg, G.E.; Stewart, W.D.P.; Fay, P.; Walsby, A.E. The blue-green algae. Vii, 459 s., 170 abb. Und 8 farbige abb., 33 tab., 1 farbtafel. London-new york 1973: Academic press, £ 8.50. Z. Allg. Mikrobiol. 1975, 15, 68. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. Cell Mol. Biol. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Ferrell, J.; Sarisky-Reed, V. National Algal Biofuels Technology Roadmap; EERE Publication and Product Library: Washington, DC, USA, 2010. [Google Scholar]
- Borowitzka, M. Culturing Microalgae in Outdoor Ponds; Academic Press: Boston, MA, USA, 2005; pp. 205–221. [Google Scholar]
- Mehla, S.; Kukade, S.; Kumar, P.; Rao, P.; Sriganesh, G.; Ravishankar, R. Fine tuning h-transfer and β-scission reactions in vgo fcc using metal promoted dual functional zsm-5. Fuel 2019, 242, 487–495. [Google Scholar] [CrossRef]
- Möllers, K.B.; Cannella, D.; Jørgensen, H.; Frigaard, N.-U. Cyanobacterial biomass as carbohydrate and nutrient feedstock for bioethanol production by yeast fermentation. Biotechnol. Biofuels 2014, 7, 64. [Google Scholar] [CrossRef]
- Babushkina, E.; Belokopytova, L.; Grachev, A.; Meko, D.; Vaganov, E.A. Variation of the hydrological regime of bele-shira closed basin in southern siberia and its reflection in the radial growth of Larix sibirica. Reg. Environ. Chang. 2017, 17, 1725–1737. [Google Scholar] [CrossRef]
- Dean, A.P.; Nicholson, J.M.; Sigee, D.C. Impact of phosphorus quota and growth phase on carbon allocation in Chlamydomonas reinhardtii: An ftir microspectroscopy study. Eur. J. Phycol. 2008, 43, 345–354. [Google Scholar] [CrossRef]
- Cade-Menun, B.J.; Paytan, A. Nutrient temperature and light stress alter phosphorus and carbon forms in culture-grown algae. Mar. Chem. 2010, 121, 27–36. [Google Scholar] [CrossRef]
- Felczykowska, A.; Pawlik, A.; Mazur-Marzec, H.; Toruńska-Sitarz, A.; Narajczyk, M.; Richert, M.; Węgrzyn, G.; Herman-Antosiewicz, A. Selective inhibition of cancer cells’ proliferation by compounds included in extracts from Baltic sea cyanobacteria. Toxicon 2015, 108, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides B and C, cancer cell toxins from a Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef]
- Yu, F.R.; Lian, X.Z.; Guo, H.Y.; McGuire, P.M.; Li, R.D.; Wang, R.; Yu, F.H. Isolation and characterization of methyl esters and derivatives from Euphorbia kansui (euphorbiaceae) and their inhibitory effects on the human sgc-7901 cells. J. Pharm. Pharm. Sci. 2005, 8, 528–535. [Google Scholar]
- Altameme, H.; Hameed, I.; Idan, S.; Hadi, M. Biochemical analysis of Origanum vulgare seeds by fourier-transform infrared (ft-ir) spectroscopy and gas chromatography-mass spectrometry (gc-ms). J. Pharmacogn. Phytother. Res. 2015, 7, 221–237. [Google Scholar]
- Ibraheam, I.A.; Hadi, M.Y.; Hameed, I.H. Analysis of bioactive compounds of methanolic leaves extract of mentha pulegium using gas chromatography-mass spectrometry (gc-ms) technique. Int. J. Pharm. Qual. Assur. 2017, 8, 174–182. [Google Scholar] [CrossRef]
- Nair, R.V.R.; Jayasree, D.V.; Biju, P.G.; Baby, S. Anti-inflammatory and anticancer activities of erythrodiol-3-acetate and 2,4-di-tert-butylphenol isolated from humboldtia unijuga. Nat. Prod. Res. 2020, 34, 2319–2322. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, P.; Lucardi, R.; Su, Z.; Li, S. Natural sources and bioactivities of 2,4-di-tert-butylphenol and its analogs. Toxins 2020, 12, 35. [Google Scholar] [CrossRef]
- Costa, M.; Costa-Rodrigues, J.; Fernandes, M.H.; Barros, P.; Vasconcelos, V.; Martins, R. Marine cyanobacteria compounds with anticancer properties: A review on the implication of apoptosis. Mar. Drugs 2012, 10, 2181–2207. [Google Scholar] [CrossRef]
- Oftedal, L.; Myhren, L.; Jokela, J. The lipopeptide toxins anabaenolysin a and b target biological membranes in a cholesterol-dependent manner. Biochim. Biophys. Acta Biomembr. 2012, 1818, 3000–3009. [Google Scholar] [CrossRef]
- Catassi, A.; Cesario, A.; Arzani, D.; Menichini, P.; Alama, A.; Bruzzo, C.; Imperatori, A.; Rotolo, N.; Granone, P.; Russo, P. Characterization of apoptosis induced by marine natural products in non small cell lung cancer a549 cells. Cell. Mol. Life Sci. 2006, 63, 2377–2386. [Google Scholar] [CrossRef]
- Sato, M.; Sagawa, M.; Nakazato, T.; Ikeda, Y.; Kizaki, M. A natural peptide, dolastatin 15, induces g2/m cell cycle arrest and apoptosis of human multiple myeloma cells. Int. J. Oncol. 2007, 30, 1453–1459. [Google Scholar] [CrossRef]
- Sato, S.; Murata, A.; Orihara, T.; Shirakawa, T.; Suenaga, K.; Kigoshi, H.; Uesugi, M. Marine natural product aurilide activates the opa1-mediated apoptosis by binding to prohibitin. Chem. Biol. 2011, 18, 131–139. [Google Scholar] [CrossRef]
- Paradies, G.; Petrosillo, G.; Paradies, V.; Ruggiero, F. Mitochondrial dysfunction in brain aging: Role of oxidative stress and cardiolipin. Neurochem. Int. 2011, 58, 447–457. [Google Scholar] [CrossRef]
- Oftedal, L.; Selheim, F.; Wahlsten, M.; Sivonen, K.; Døskeland, S.; Herfindal, L. Marine benthic cyanobacteria contain apoptosis-inducing activity synergizing with daunorubicin to kill leukemia cells, but not cardiomyocytes. Mar. Drugs 2010, 8, 2659–2672. [Google Scholar] [CrossRef]
- Yonezawa, T.; Mase, N.; Sasaki, H.; Teruya, T.; Hasegawa, S.-I.; Cha, B.-Y.; Yagasaki, K.; Suenaga, K.; Nagai, K.; Woo, J.-T. Biselyngbyaside, isolated from marine cyanobacteria, inhibits osteoclastogenesis and induces apoptosis in mature osteoclasts. J. Cell. Biochem. 2012, 113, 440–448. [Google Scholar] [CrossRef]
- Pereira, A.; Cao, Z.; Murray, T.F.; Gerwick, W.H. Hoiamide a, a sodium channel activator of unusual architecture from a consortium of two papua new guinea cyanobacteria. Chem. Biol. 2009, 16, 893–906. [Google Scholar] [CrossRef]
- Choi, H.; Pereira, A.; Cao, Z.; Shuman, C.; Engene, N.; Byrum, T.; Matainaho, L.; Murray, T.; Mangoni, A.; Gerwick, W. The hoiamides, structurally intriguing neurotoxic lipopeptides from papua new guinea marine cyanobacteria. J. Nat. Prod. 2010, 73, 1411–1421. [Google Scholar] [CrossRef]
- Acuña, U.M.; Mo, S.; Zi, J.; Orjala, J.; de Blanco, E.J.C. Hapalindole h induces apoptosis as an inhibitor of nf-kb and affects the intrinsic mitochondrial pathway in pc-3 androgen-insensitive prostate cancer cells. Anticancer Res. 2018, 38, 3299–3307. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, G.E.; Md Akim, A.; Sung, Y.Y.; Sifzizul, T.M.T. Cancer and apoptosis: The apoptotic activity of plant and marine natural products and their potential as targeted cancer therapeutics. Front. Pharmacol. 2022, 13, 842376. [Google Scholar] [CrossRef]
- Ramamurthy, V.; Raveendran, S.; Thirumeni, S.; Krishnaveni, S. Antimicrobial activity of heterocytic cyanobacteria. Int. J. Adv. Life Sci. 2012, 1, 32–39. [Google Scholar]
- Khairy, H.M.; El-Kassas, H.Y. Active substance from some blue green algal species used as antimicrobial agents. Afr. J. Biotechnol. 2010, 9, 2789–2800. [Google Scholar]
- El-Sheekh, M.; Osman, M.; Dyab, M.; Amer, M. Production and characterization of antimicrobial active substance from the cyanobacterium Nostoc muscorum. Environ. Toxicol. Pharmacol. 2006, 21, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Mian, P.; Heilmann, J.; Bürgi, H.R.; Sticher, O. Biological screening of terrestrial and freshwater cyanobacteria for antimicrobial activity, brine shrimp lethality, and cytotoxicity. Pharm. Biol. 2008, 41, 243–247. [Google Scholar] [CrossRef]
- Huang, C.B.; George, B.; Ebersole, J.L. Antimicrobial activity of n-6, n-7 and n-9 fatty acids and their esters for oral microorganisms. Arch. Oral Biol. 2010, 55, 555–560. [Google Scholar] [CrossRef]
- Amaral, A.C.; Gomes, L.A.; Silva, J.R.; Ferreira, J.L.; Ramos Ade, S.; Rosa Mdo, S.; Vermelho, A.B.; Rodrigues, I.A. Liposomal formulation of turmerone-rich hexane fractions from Curcuma longa enhances their antileishmanial activity. BioMed Res. Int. 2014, 2014, 694934. [Google Scholar] [CrossRef]
- Sundar, S.; Justin-Koil Pillai, Y. Phytochemical screening and gas chromatograph-mass spectrometer profiling in the leaves of Solanum incanum. Asian J. Pharm. Clin. Res. 2015, 8, 179–188. [Google Scholar]
- Mehta, M.; Adem, A.; Sabbagh, M. New acetylcholinesterase inhibitors for alzheimer’s disease. Int. J. Alzheimer’s Dis. 2012, 2012, 728983. [Google Scholar] [CrossRef] [Green Version]
- Konrath, E.L.; Neves, B.M.; Lunardi, P.S.; Passos, C.d.S.; Simões-Pires, A.; Ortega, M.G.; Gonçalves, C.A.; Cabrera, J.L.; Moreira, J.C.F.; Henriques, A.T. Investigation of the in vitro and ex vivo acetylcholinesterase and antioxidant activities of traditionally used lycopodium species from south america on alkaloid extracts. J. Ethnopharmacol. 2012, 139, 58–67. [Google Scholar] [CrossRef]
- Ferreira, A.; Proença, C.; Serralheiro, M.L.M.; Araújo, M.E.M. The in vitro screening for acetylcholinesterase inhibition and antioxidant activity of medicinal plants from portugal. J. Ethnopharmacol. 2006, 108, 31–37. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Kumar, V.; Mal, M.; Houghton, P.J. Acetylcholinesterase inhibitors from plants. Phytomedicine 2007, 14, 289–300. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constituents | Zarrouk’s Medium (g L−1) * | Modified Zarrouk’s Medium (g L−1) | ||
|---|---|---|---|---|
| 100% (T1) | 50% (T2) | 12.5% (T3) | ||
| NaNO3 | 2.5 | - | - | - |
| Urea | - | 0.8 | 0.4 | 0.1 |
| CaCl2·2H2O | 0.04 | - | - | - |
| NaCl | 1.0 | - | - | - |
| FeSO4·7H2O | 0.01 | 0.01 | 0.005 | 0.00125 |
| EDTA (Na) | 0.08 | - | - | - |
| K2SO4 | 1.0 | 1.0 | 0.5 | 0.125 |
| MgSO4·7H2O | 0.2 | 0.2 | 0.1 | 0.025 |
| NaHCO3 | 16.8 | 16.8 | 8.4 | 2.1 |
| K2HPO4 | 0.5 | 0.5 | 0.25 | 0.0625 |
| Micronutrients (H3BO3, MnCl2·4H2O, ZnSO4·4H2O Na2MoO4, CuSO4·5H2O) | 1.0 mL | - | - | - |
| Treatments | Dry wt. (g L−1) |
|---|---|
| Control (standard Zarrouk’s medium) | 0.552 a ± 0.027 * |
| T1 | 0.539 ab ± 0.025 |
| T2 | 0.53 ab ± 0.022 |
| T3 | 0.509 ab ± 0.022 |
| Bacteria | Diameter of Inhibition Zone (mm) | ||
|---|---|---|---|
| Zarrouk’s Medium (Control) | T3 a | Solvent b | |
| Pseudomonas aeruginosa | N | N | N c |
| Escherichia coli | N | N | N |
| Bacillus cereus | 16.3 ± 0.5 | 17.2 ± 0.44 | N |
| Staphylococcus aureus | 12.3 ± 0.7 | 15.3 ± 0.58 | N |
| Peak No. | Compound | Chemical Group | Retention Time (min.) | Formula | Area% |
|---|---|---|---|---|---|
| 1 | Oleic acid, eicosyl ester | Oleic acid ester | 11.383 | C38H74O2 | 5.5 |
| 2 | Phenol, 2,4-bis(1,1-dimethylethyl)- | Phenol | 13.652 | C14H22O | 25 |
| 3 | Pentatriacont-17-ene | Alkene | 14.808 | C35H70 | 6.2 |
| 4 | 9-Hexadecenoic acid, 9-octadecenyl ester, (Z,Z)- | Ester | 18.134 | C34H64O2 | 3.8 |
| 5 | 7-Hexadecenoic acid, methyl ester, (Z)- | Methyl ester | 20.175 | C17H32O2 | 5.7 |
| 6 | Hexadecanoic acid, methyl ester | Methyl ester | 20.532 | C17H34O2 | 16 |
| 7 | Dasycarpidan-1-methanol, acetate (ester) | Indole alkaloid | 23.477 | C20H26N2O2 | 13.1 |
| 8 | 9,12-Octadecadienoyl chloride, (Z, Z)- | Linoleic acid chloride | 23.668 | C18H31ClO | 10.5 |
| 9 | Cholestan-3-ol, 2-methylene-, (3β,5α)- | Steroid | 23.975 | C28H48O | 7.1 |
| 10 | 3′,8,8′-Trimethoxy-3-piperidyl-2,2′-binaphthalene-1,1′,4,4′-tetrone | Terpene | 31.095 | C28H25NO7 | 7.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Touliabah, H.E.; Refaay, D.A. Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions. Water 2023, 15, 664. https://doi.org/10.3390/w15040664
Touliabah HE, Refaay DA. Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions. Water. 2023; 15(4):664. https://doi.org/10.3390/w15040664
Chicago/Turabian StyleTouliabah, Hussein E., and Dina A. Refaay. 2023. "Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions" Water 15, no. 4: 664. https://doi.org/10.3390/w15040664
APA StyleTouliabah, H. E., & Refaay, D. A. (2023). Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions. Water, 15(4), 664. https://doi.org/10.3390/w15040664