
Acute and Subchronic Exposure of the Common Carp (Cyprinus carpio) to Herbicide S-Metolachlor

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Chemicals
2.2. Experimental Design
2.3. Acute Toxicity Test
2.4. Subchronic Toxicity Test
2.5. Sampling
2.6. Blood Biochemistry Analyses
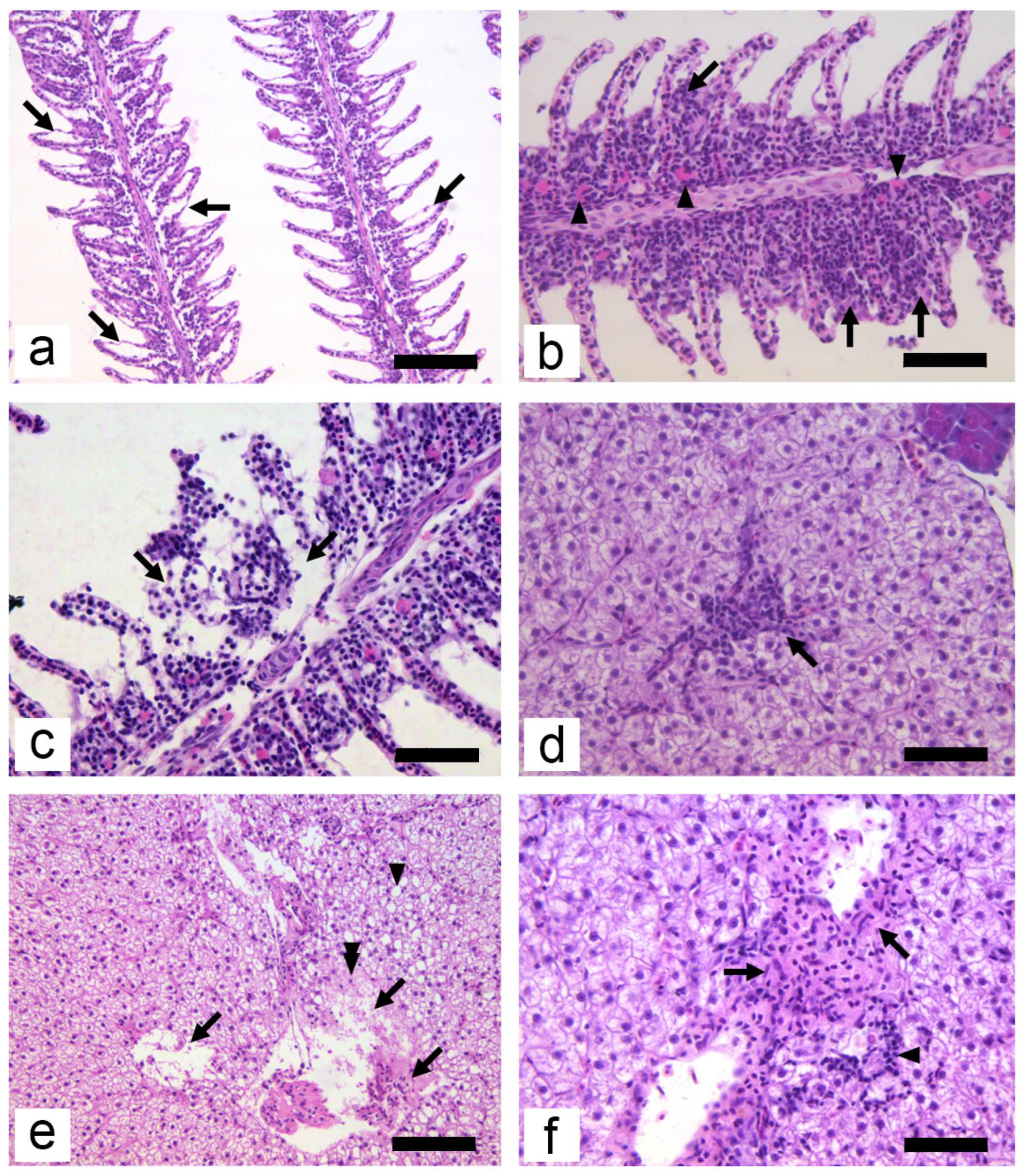
2.7. Histological Analyses
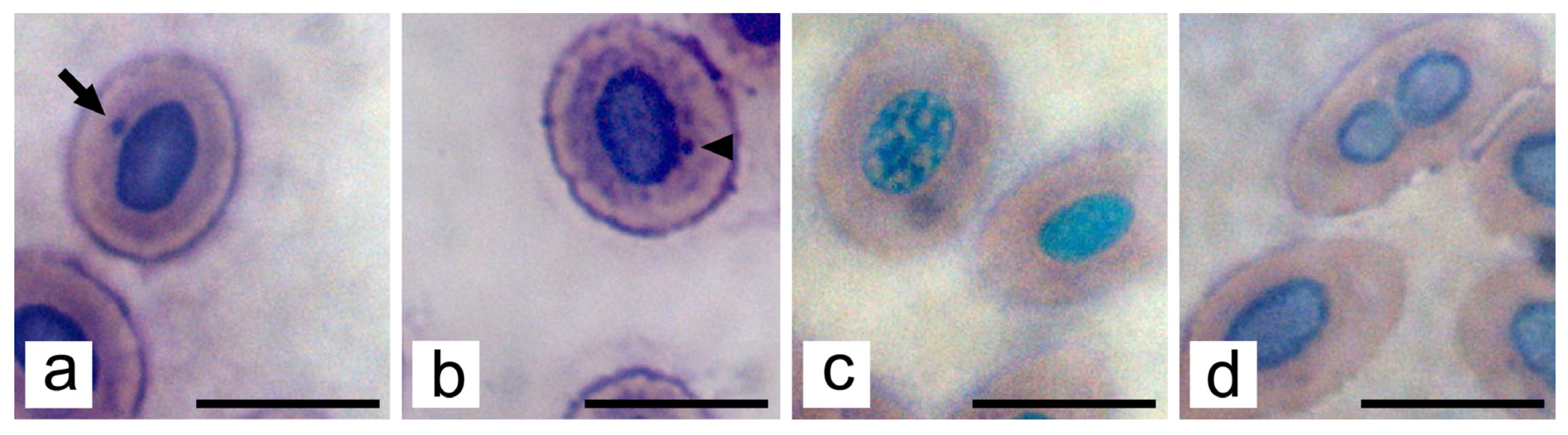
2.8. Nuclear Abnormalities of Erythrocytes
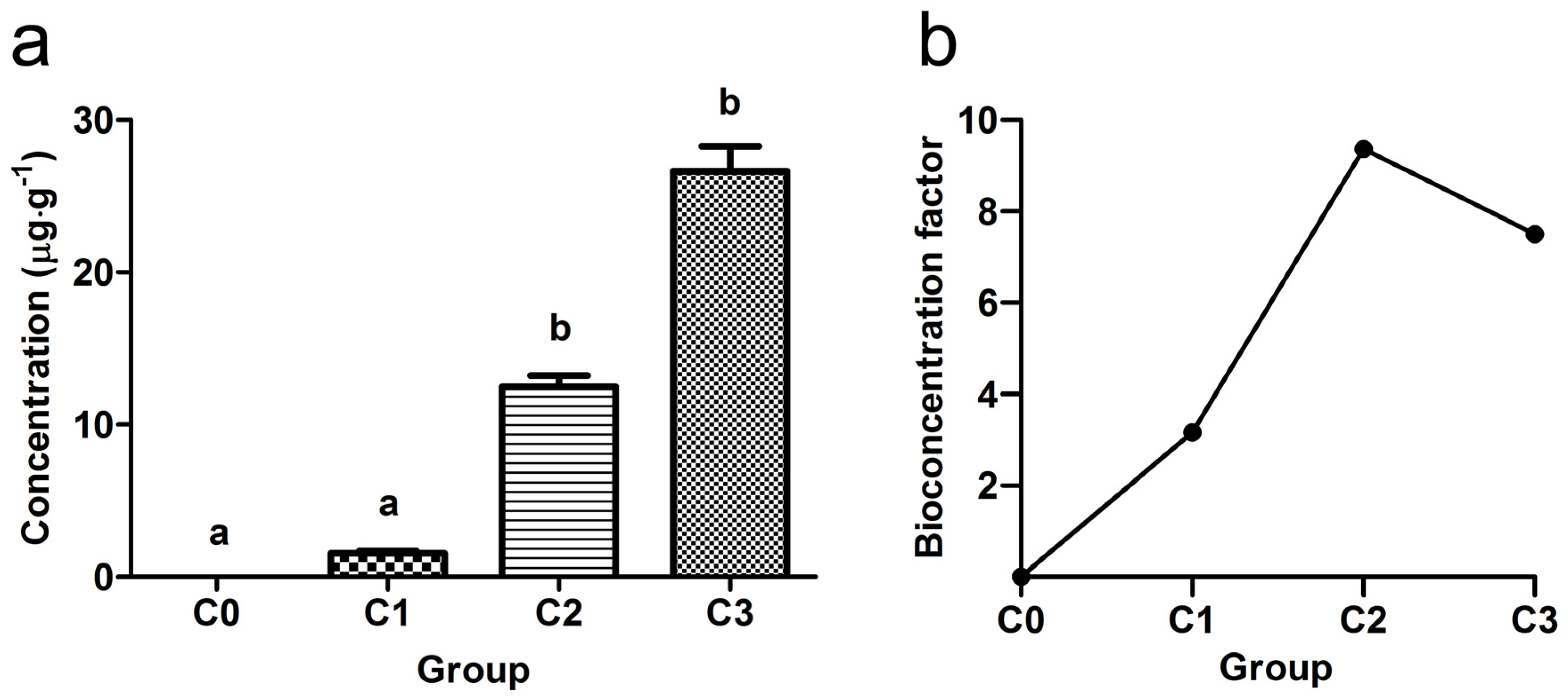
2.9. Determination of S-MET Concentrations in Water and Common Carp Muscles
2.9.1. Chemicals and Apparatus
2.9.2. Sample Preparation for LC-MS/MS Analyses
2.10. Bioconcentration Factor
2.11. Statistical Analyses
3. Results
3.1. Acute Toxicity Test
3.2. Subchronic Toxicity Test
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jorge, F.-C.; Nehring, R.; Osteen, C.; Wechsler, S.; Martin, A.; Vialou, A. Pesticide Use in U.S. Agriculture: 21 Selected Crops, 1960–2008; U.S. Department of Agriculture, Economic Research Service: Washington, DC, USA, 2014. [Google Scholar]
- O’Connell, P.J.; Harms, C.T.; Allen, J.R.F. Metolachlor, S-metolachlor and their role within sustainable weed-management. Crop Prot. 1998, 17, 207–212. [Google Scholar] [CrossRef]
- Eurostat. The Use of Plant Protection Products in the European Union (Data 1992–2003); European Communities: Luxembourg, 2007; p. 222. [Google Scholar]
- Fenner, K.; Canonica, S.; Wackett, L.P.; Elsner, M. Evaluating Pesticide Degradation in the Environment: Blind Spots and Emerging Opportunities. Science 2013, 341, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Vallotton, N.; Moser, D.; Eggen, R.I.; Junghans, M.; Chevre, N. S-metolachlor pulse exposure on the alga Scenedesmus vacuolatus: Effects during exposure and the subsequent recovery. Chemosphere 2008, 73, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Lanasa, S.; Niedzwiecki, M.; Reber, K.P.; East, A.; Sivey, J.D.; Salice, C.J. Comparative Toxicity of Herbicide Active Ingredients, Safener Additives, and Commercial Formulations to the Nontarget Alga Raphidocelis subcapitata. Environ. Toxicol. Chem. 2022, 41, 1466–1476. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Q.; Su, W.; Sun, L.; Xu, H.; Xue, F.; Lu, C.; Wu, R. The mechanism of exogenous gibberellin A3 protecting sorghum shoots from S-metolachlor Phytotoxicity. Pest Manag. Sci. 2022, 78, 4497–4506. [Google Scholar] [CrossRef]
- Götz, T.; Böger, P. The Very-Long-Chain Fatty Acid Synthase Is Inhibited by Chloroacetamides. Z. Naturforsch. C 2004, 59, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Schmalfuß, J.; Matthes, B.; Mayer, P.; Böger, P. Chloroacetamide Mode of Action, I: Inhibition of Very Long Chain Fatty Acid Synthesis in Scenedesmus acutus. Z. Naturforsch. C 1998, 53, 995–1003. [Google Scholar] [CrossRef][Green Version]
- Filimon, M.N.; Roman, D.L.; Caraba, I.V.; Isvoran, A. Assessment of the Effect of Application of the Herbicide S-Metolachlor on the Activity of Some Enzymes Found in Soil. Agriculture 2021, 11, 469. [Google Scholar] [CrossRef]
- Lunn, R.D.J.; Tocher, D.A.; Sidebottom, P.J.; Montgomery, M.G.; Keates, A.C.; Carmalt, C.J. Applying the Crystalline Sponge Method to Agrochemicals: Obtaining X-ray Structures of the Fungicide Metalaxyl-M and Herbicide S-Metolachlor. Cryst. Growth Des. 2021, 21, 3024–3036. [Google Scholar] [CrossRef]
- Marín-Benito, J.M.; Herrero-Hernández, E.; Ordax, J.M.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S. The role of two organic amendments to modify the environmental fate of S-metolachlor in agricultural soils. Environ. Res. 2021, 195, 110871. [Google Scholar] [CrossRef]
- HRAC. Global Herbicide Classification Lookup. Available online: Hracglobal.com/tools/classification-lookup (accessed on 12 October 2023).
- Mottes, C.; Lesueur Jannoyer, M.; Le Bail, M.; Guene, M.; Carles, C.; Malezieux, E. Relationships between past and present pesticide applications and pollution at a watershed outlet: The case of a horticultural catchment in Martinique, French West Indies. Chemosphere 2017, 184, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Battaglin, W.A.; Furlong, E.T.; Burkhardt, M.R.; Peter, C.J. Occurrence of sulfonylurea, sulfonamide, imidazolinone, and other herbicides in rivers, reservoirs and ground water in the Midwestern United States, 1998. Sci. Total Environ. 2000, 248, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, B.; Wezel, A.; Guerin, M.; Robin, J. Pesticide contamination of fish ponds in relation to crop area in a mixed farmland-pond landscape (Dombes area, France). Environ. Sci. Pollut. Res. 2022, 29, 66858–66873. [Google Scholar] [CrossRef] [PubMed]
- Brumovsky, M.; Becanova, J.; Kohoutek, J.; Borghini, M.; Nizzetto, L. Contaminants of emerging concern in the open sea waters of the Western Mediterranean. Environ. Pollut. 2017, 229, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, L.; Chen, K.; Yang, H.; Ling, M.; Wu, L.; Zhou, X.; Ma, G.; Bai, L. Combined effects of S-metolachlor and benoxacor on embryo development in zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2022, 238, 113565. [Google Scholar] [CrossRef] [PubMed]
- Ou-Yang, K.; Feng, T.; Han, Y.; Li, G.; Li, J.; Ma, H. Bioaccumulation, metabolism and endocrine-reproductive effects of metolachlor and its S-enantiomer in adult zebrafish (Danio rerio). Sci. Total Environ. 2022, 802, 149826. [Google Scholar] [CrossRef] [PubMed]
- Quintaneiro, C.; Patricio, D.; Novais, S.C.; Soares, A.; Monteiro, M.S. Endocrine and physiological effects of linuron and S-metolachlor in zebrafish developing embryos. Sci. Total Environ. 2017, 586, 390–400. [Google Scholar] [CrossRef]
- Rozmankova, E.; Pipal, M.; Blahova, L.; Njattuvetty Chandran, N.; Morin, B.; Gonzalez, P.; Blaha, L. Environmentally relevant mixture of S-metolachlor and its two metabolites affects thyroid metabolism in zebrafish embryos. Aquat. Toxicol. 2020, 221, 105444. [Google Scholar] [CrossRef]
- Yang, L.; Ivantsova, E.; Souders, C.L., 2nd; Martyniuk, C.J. The agrochemical S-metolachlor disrupts molecular mediators and morphology of the swim bladder: Implications for locomotor activity in zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2021, 208, 111641. [Google Scholar] [CrossRef]
- Dobšíková, R.; Blahová, J.; Modrá, H.; Škorič, M.; Svobodová, Z. The effect of acute exposure to herbicide Gardoprim Plus Gold 500 SC on haematological and biochemical indicators and histopathological changes in common carp (Cyprinus carpio L.). Acta Vet. Brno 2011, 80, 359–363. [Google Scholar] [CrossRef]
- OECD. Test No. 203: Fish, Acute Toxicity Test; OECD: Paris, France, 2019. [Google Scholar]
- OECD. Test No. 215: Fish, Juvenile Growth Test; OECD: Paris, France, 2000. [Google Scholar]
- Svobodová, Z.; Lloyd, R.; Máchová, J.; Vykusová, B. Water Quality and Fish Health; Food and Agriculture Organization of the United Nations: Rome, Italy, 1993; p. 59. [Google Scholar]
- Chen, C.-Y.; Wooster, G.A.; Getchell, R.G.; Bowser, P.R.; Timmons, M.B. Blood chemistry of healthy, nephrocalcinosis-affected and ozone-treated tilapia in a recirculation system, with application of discriminant analysis. Aquaculture 2003, 218, 89–102. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Fenech, M.; Chang, W.P.; Kirsch-Volders, M.; Holland, N.; Bonassi, S.; Zeiger, E. HUMN project: Detailed description of the scoring criteria for the cytokinesis-block micronucleus assay using isolated human lymphocyte cultures. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2003, 534, 65–75. [Google Scholar] [CrossRef] [PubMed]
- RAC. Proposing Harmonised Classification and Labelling at EU Level of S-Metolachlor; RAC: Walsall, UK, 2022; p. 34. [Google Scholar]
- Arnot, J.A.; Gobas, F.A.P.C. A review of bioconcentration factor (BCF) and bioaccumulation factor (BAF) assessments for organic chemicals in aquatic organisms. Environ. Rev. 2006, 14, 257–297. [Google Scholar] [CrossRef]
- Gutiérrez, I.B.; Mesquita, A.F.; Gonçalves, F.J.M.; Marques, J.C.; Gonçalves, A.M.M. Biomarkers’ responses of the benthic clam Scrobicularia plana to the main active ingredients (S-metolachlor and Terbuthylazine) of a common herbicide. Ecol. Indic. 2019, 96, 611–619. [Google Scholar] [CrossRef]
- Quintaneiro, C.; Soares, A.; Monteiro, M.S. Effects of the herbicides linuron and S-metolachlor on Perez’s frog embryos. Chemosphere 2018, 194, 595–601. [Google Scholar] [CrossRef]
- Anderson, J.C.; Marteinson, S.C.; Prosser, R.S. Prioritization of Pesticides for Assessment of Risk to Aquatic Ecosystems in Canada and Identification of Knowledge Gaps. In Reviews of Environmental Contamination and Toxicology; de Voogt, P., Ed.; Springer International Publishing: Cham, Switzerland, 2021; Volume 259, pp. 171–231. [Google Scholar]
- van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Banaee, M.; Mirvagefei, A.R.; Rafei, G.R.; Majazi Amiri, B. Effect of sub-lethal diazinon concentrations on blood plasma biochemistry. Int. J. Environ. Res. 2008, 2, 189–198. [Google Scholar]
- Neskovic, N.K.; Elezovic, I.; Karan, V.; Poleksic, V.; Budimir, M. Acute and subacute toxicity of atrazine to carp (Cyprinus carpio L.). Ecotoxicol. Environ. Saf. 1993, 25, 173–182. [Google Scholar] [CrossRef]
- Poleksić, V.; Karan, V. Effects of trifluralin on carp: Biochemical and histological evaluation. Ecotoxicol. Environ. Saf. 1999, 43, 213–221. [Google Scholar] [CrossRef]
- Velisek, J.; Stara, A.; Zuskova, E.; Svobodova, Z. Use of biometric, hematologic, and plasma biochemical variables, and histopathology to assess the chronic effects of the herbicide prometryn on Common Carp. Vet. Clin. Pathol. 2013, 42, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Velisek, J.; Sudova, E.; Machova, J.; Svobodova, Z. Effects of sub-chronic exposure to terbutryn in common carp (Cyprinus carpio L.). Ecotoxicol. Environ. Saf. 2010, 73, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Banaee, M.; Sureda, A.; Mirvaghefi, A.R.; Ahmadi, K. Effects of diazinon on biochemical parameters of blood in rainbow trout (Oncorhynchus mykiss). Pestic. Biochem. Physiol. 2011, 99, 1–6. [Google Scholar] [CrossRef]
- Stara, A.; Kubec, J.; Zuskova, E.; Buric, M.; Faggio, C.; Kouba, A.; Velisek, J. Effects of S-metolachlor and its degradation product metolachlor OA on marbled crayfish (Procambarus virginalis). Chemosphere 2019, 224, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Nwani, C.D.; Ifo, C.T.; Nwamba, H.O.; Ejere, V.C.; Onyishi, G.C.; Oluah, S.N.; Ikwuagwu, O.E.; Odo, G.E. Oxidative stress and biochemical responses in the tissues of African catfish Clarias gariepinus juvenile following exposure to primextra herbicide. Drug Chem. Toxicol. 2015, 38, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Mikula, P.; Modra, H.; Nemethova, D.; Groch, L.; Svobodova, Z. Effects of subchronic exposure to LASSO MTX® (Alachlor 42% W/V) on hematological indices and histology of the common carp, Cyprinus carpio L. Bull. Environ. Contam. Toxicol. 2008, 81, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Velisek, J.; Stara, A.; Koutnik, D.; Machova, J. Effects of prometryne on early life stages of common carp (Cyprinus carpio L.). Pestic. Biochem. Physiol. 2015, 118, 58–63. [Google Scholar] [CrossRef]
- Velisek, J.; Stara, A.; Machova, J.; Dvorak, P.; Zuskova, E.; Prokes, M.; Svobodova, Z. Effect of terbutryn at environmental concentrations on early life stages of common carp (Cyprinus carpio L.). Pestic. Biochem. Physiol. 2012, 102, 102–108. [Google Scholar] [CrossRef]
- Velisek, J.; Stara, A.; Machova, J.; Svobodova, Z. Effects of long-term exposure to simazine in real concentrations on common carp (Cyprinus carpio L.). Ecotoxicol. Environ. Saf. 2012, 76, 79–86. [Google Scholar] [CrossRef]
- Nešković, N.K.; Poleksić, V.; Elezović, I.; Karan, V.; Budimir, M. Biochemical and histopathological effects of glyphosate on carp, Cyprinus carpio L. Bull. Environ. Contam. Toxicol. 1996, 56, 295–302. [Google Scholar] [CrossRef]
- Qureshi, I.Z.; Bibi, A.; Shahid, S.; Ghazanfar, M. Exposure to sub-acute doses of fipronil and buprofezin in combination or alone induces biochemical, hematological, histopathological and genotoxic damage in common carp (Cyprinus carpio L.). Aquat. Toxicol. 2016, 179, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Kokushi, E.; Koyama, J.; Uno, S.; Ghosh, A.R. Histopathological alterations in gill, liver and kidney of common carp exposed to chlorpyrifos. J. Environ. Sci. Health B 2012, 47, 180–195. [Google Scholar] [CrossRef]
- Xing, H.; Li, S.; Wang, Z.; Gao, X.; Xu, S.; Wang, X. Oxidative stress response and histopathological changes due to atrazine and chlorpyrifos exposure in common carp. Pestic. Biochem. Physiol. 2012, 103, 74–80. [Google Scholar] [CrossRef]
- Yancheva, V.; Georgieva, E.; Velcheva, I.; Iliev, I.; Stoyanova, S.; Vasileva, T.; Bivolarski, V.; Todorova-Bambaldokova, D.; Zulkipli, N.; Antal, L.; et al. Assessment of the exposure of two pesticides on common carp (Cyprinus carpio Linnaeus, 1758): Are the prolonged biomarker responses adaptive or destructive? Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 261, 109446. [Google Scholar] [CrossRef] [PubMed]
- Au, D.W.T. The application of histo-cytopathological biomarkers in marine pollution monitoring: A review. Mar. Pollut. Bull. 2004, 48, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Bernet, D.; Schmidt-Posthaus, H.; Wahli, T.; Burkhardt-Holm, P. Evaluation of two monitoring approaches to assess effects of waste water disposal on histological alterations in fish. Hydrobiologia 2004, 524, 53–66. [Google Scholar] [CrossRef]
- Nikolić, D.; Skorić, S.; Rašković, B.; Lenhardt, M.; Krpo-Ćetković, J. Impact of reservoir properties on elemental accumulation and histopathology of European perch (Perca fluviatilis). Chemosphere 2020, 244, 125503. [Google Scholar] [CrossRef]
- Rašković, B.; Jarić, I.; Koko, V.; Spasić, M.; Dulić, Z.; Marković, Z.; Poleksić, V. Histopathological indicators: A useful fish health monitoring tool in common carp (Cyprinus carpio Linnaeus, 1758) culture. Cent. Eur. J. Biol. 2013, 8, 975–985. [Google Scholar] [CrossRef]
- Harper, C.; Wolf, J.C. Morphologic effects of the stress response in fish. ILAR J. 2009, 50, 387–396. [Google Scholar] [CrossRef]
- Monteiro, S.M.; Rocha, E.; Mancera, J.M.; Fontaínhas-Fernandes, A.; Sousa, M. A stereological study of copper toxicity in gills of Oreochromis niloticus. Ecotoxicol. Environ. Saf. 2009, 72, 213–223. [Google Scholar] [CrossRef]
- Mueller, M.E.; Sanchez, D.A.; Bergman, H.L.; McDonald, D.G.; Rhem, R.G.; Wood, C.M. Nature and time course of acclimation to aluminum in juvenile brook trout (Salvelinus fontinalis). II. Gill histology. Can. J. Fish. Aquat. Sci. 1991, 48, 2016–2027. [Google Scholar] [CrossRef]
- Destro, A.L.F.; Silva, S.B.; Gregorio, K.P.; de Oliveira, J.M.; Lozi, A.A.; Zuanon, J.A.S.; Salaro, A.L.; da Matta, S.L.P.; Goncalves, R.V.; Freitas, M.B. Effects of subchronic exposure to environmentally relevant concentrations of the herbicide atrazine in the Neotropical fish Astyanax altiparanae. Ecotoxicol. Environ. Saf. 2021, 208, 111601. [Google Scholar] [CrossRef] [PubMed]
- Ghelichpour, M.; Taheri Mirghaed, A.; Mirzargar, S.S.; Joshaghani, H.; Ebrahimzadeh Mousavi, H. Plasma proteins, hepatic enzymes, thyroid hormones and liver histopathology of Cyprinus carpio (Linnaeus, 1758) exposed to an oxadiazin pesticide, indoxacarb. Aquac. Res. 2017, 48, 5666–5676. [Google Scholar] [CrossRef]
- Jiraungkoorskula, W.; Upathama, E.S.; Kruatrachue, M.; Sahaphong, S.; Vichasri-Gramsa, S.; Pokethitiyooka, P. Histopathological effects of Roundup, a glyphosate herbicide, on Nile tilapia (Oreochromis niloticus). Sci. Asia 2002, 28, 121–127. [Google Scholar] [CrossRef]
- Speare, D.J.; Ferguson, H.W. Fixation artifacts in rainbow trout (Salmo gairdneri) gills: A morphometric evaluation. Can. J. Fish. Aquat. Sci. 1989, 46, 780–785. [Google Scholar] [CrossRef]
- Wolf, J.C.; Baumgartner, W.A.; Blazer, V.S.; Camus, A.C.; Engelhardt, J.A.; Fournie, J.W.; Frasca, S.; Groman, D.B.; Kent, M.L.; Khoo, L.H.; et al. Nonlesions, misdiagnoses, missed diagnoses, and other interpretive challenges in fish histopathology studies: A guide for investigators, authors, reviewers, and readers. Toxicol. Pathol. 2014, 43, 297–325. [Google Scholar] [CrossRef]
- Velisek, J.; Svobodova, Z.; Piackova, V.; Groch, L.; Nepejchalova, L. Effects of clove oil anaesthesia on common carp (Cyprinus carpio L.). Vet. Med. 2012, 50, 269–275. [Google Scholar] [CrossRef]
- Wolf, J.C. Comparing apples and oranges and pears and kumquats: The misuse of index systems for processing histopathology data in fish toxicological bioassays. Environ. Toxicol. Chem. 2018, 37, 1688–1695. [Google Scholar] [CrossRef]
- Bolognesi, C.; Perrone, E.; Roggieri, P.; Pampanin, D.M.; Sciutto, A. Assessment of micronuclei induction in peripheral erythrocytes of fish exposed to xenobiotics under controlled conditions. Aquat. Toxicol. 2006, 78, S93–S98. [Google Scholar] [CrossRef]
- Buschini, A.; Martino, A.; Gustavino, B.; Monfrinotti, M.; Poli, P.; Rossi, C.; Santoro, M.; Dörr, A.J.M.; Rizzoni, M. Comet assay and micronucleus test in circulating erythrocytes of Cyprinus carpio specimens exposed in situ to lake waters treated with disinfectants for potabilization. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2004, 557, 119–129. [Google Scholar] [CrossRef]
- Grisolia, C.K.; Starling, F.L.R.M. Micronuclei monitoring of fishes from Lake Paranoá, under influence of sewage treatment plant discharges. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2001, 491, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Gustavino, B.; Scornajenghi, K.A.; Minissi, S.; Ciccotti, E. Micronuclei induced in erythrocytes of Cyprinus carpio (teleostei, pisces) by X-rays and colchicine. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2001, 494, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Llorente, M.T.; Martos, A.; Castaño, A. Detection of Cytogenetic Alterations and Blood Cell Changes in Natural Populations of Carp. Ecotoxicology 2002, 11, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Nepomuceno, J.C.; Ferrari, Í.; Spanó, M.A.; Centeno, A.J. Detection of micronuclei in peripheral erythrocytes of Cyprinus carpio exposed to metallic mercury. Environ. Mol. Mutagen. 1997, 30, 293–297. [Google Scholar] [CrossRef]
- Al-Sabti, K. Clastogenic effects of five carcinogenic-mutagenic chemicals on the cells of the common carp, Cyprinus carpio L. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 1986, 85, 5–9. [Google Scholar] [CrossRef]
- EC. Commision Regulation (EU) No 1317/2013 Amending Annexes II, III and V to Regulation (EC) No 396/2005 of the European Parliament and of the Council as Regards Maximum Residue Levels for 2,4-D, Beflubutamid, Cyclanilide, Diniconazole, Florasulam, Metolachlor and S-Metolachlor, and Milbemectin in or on Certain Products. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32013R1317 (accessed on 30 November 2023).
- Wassenaar, P.N.H.; Verbruggen, E.M.J.; Cieraad, E.; Peijnenburg, W.; Vijver, M.G. Variability in fish bioconcentration factors: Influences of study design and consequences for regulation. Chemosphere 2020, 239, 124731. [Google Scholar] [CrossRef]
- Tsuda, T. Bioconcentration of Pesticides in Fish from Rivers and Lakes. In Pesticides; Stoytcheva, M., Ed.; IntechOpen: Rijeka, Croatia, 2011; pp. 333–350. ISBN 978-953-307-532-7. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue Alteration | Importance Factor |
|---|---|
| Gills | |
| Hyperaemia | 1 |
| Presence of EGC in gills | 1 |
| Hyperplasia of epithelial cells | 2 |
| Hyperplasia of goblet cells | 2 |
| Oedema of primary epithelium | 1 |
| Oedema of secondary epithelium | 1 |
| Atrophy of secondary lamellae | 2 |
| Necrosis | 3 |
| Liver | |
| Leukocyte infiltration | 2 |
| Pyknosis of hepatocytes’ nuclei | 1 |
| Presence of ceroid pigment | 1 |
| Hyperaemia | 1 |
| Congestion of sinusoids | 1 |
| Fibrosis of portal areas | 2 |
| Necrosis | 3 |
| Group | ||||
|---|---|---|---|---|
| Parameter | C0 | C1 | C2 | C3 |
| Weight (g) | 3.99 ± 0.66 | 3.80 ± 0.58 | 4.16 ± 0.54 | 3.81 ± 0.28 |
| Total length (cm) | 8.00 ± 0.47 | 8.20 ± 0.79 | 8.64 ± 0.33 | 8.76 ± 0.16 |
| Condition factor (g·cm−3) | 0.78 ± 0.18 | 0.69 ± 0.18 | 0.63 ± 0.06 | 0.54 ± 0.02 |
| Group | ||||
|---|---|---|---|---|
| Parameter | C0 | C1 | C2 | C3 |
| TP (g·L−1) | 18.97 ± 2.67 | 19.58 ± 2.90 | 20.52 ± 3.82 | 20.80 ± 3.22 |
| ALT (µkat·L−1) | 1.05 ± 0.34 | 0.85 ± 0.32 | 0.97 ± 0.36 | 0.84 ± 0.29 |
| AST (µkat·L−1) | 6.62 ± 3.12 | 5.70 ± 2.42 | 7.73 ± 3.93 | 7.08 ± 3.64 |
| ALP (µkat·L−1) | 0.44 ± 0.32 | 0.63 ± 0.71 | 0.72 ± 0.79 | 0.48 ± 0.43 |
| ACP (µkat·L−1) | 0.06 ± 0.03 | 0.07 ± 0.02 | 0.08 ± 0.03 | 0.08 ± 0.03 |
| LDH (µkat·L−1) | 16.65 ± 9.58 | 11.97 ± 6.68 | 17.90 ± 8.69 | 15.26 ± 7.70 |
| Group | ||||
|---|---|---|---|---|
| Tissue Alteration | C0 | C1 | C2 | C3 |
| Gills | ||||
| Hyperaemia | 2.7 ± 1.6 | 3.0 ± 1.4 | 2.2 ± 1.5 | 1.7 ± 1.7 |
| Presence of EGC in gills | 0.8 ± 1.0 | 0.5 ± 0.8 | 1.7 ± 1.4 | 1.5 ± 0.8 |
| Hyperplasia of epithelial cells | 0.5 ± 0.5 | 0.4 ± 0.4 | 1.3 ± 1.2 | 2.4 ± 2.5 |
| Hyperplasia of goblet cells | 0.8 ± 2.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.2 ± 0.4 |
| Oedema of primary epithelium | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.7 ± 1.6 |
| Oedema of secondary epithelium * | 2.7 ± 2.2 ab | 1.5 ± 1.4 a | 1.0 ± 0.9 a | 3.7 ± 1.7 b |
| Atrophy of secondary lamellae | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.3 ± 0.7 |
| Necrosis | 0.0 ± 0.0 | 0.1 ± 0.3 | 0.7 ± 0.9 | 0.1 ± 0.2 |
| Liver | ||||
| Leukocyte infiltration * | 0.0 ± 0.0 a | 1.0 ± 1.7 ab | 2.3 ± 0.8 b | 1.0 ± 1.1 ab |
| Pyknosis of hepatocytes’ nuclei | 0.0 ± 0.0 | 0.3 ± 0.8 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Presence of ceroid pigment | 0.0 ± 0.0 | 0.7 ± 1.0 | 1.0 ± 1.1 | 0.3 ± 0.8 |
| Hyperaemia | 0.3 ± 0.8 | 0.3 ± 0.8 | 0.7 ± 1.0 | 0.7 ± 1.6 |
| Congestion of sinusoids | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.3 ± 0.8 |
| Fibrosis of portal areas | 0.0 ± 0.0 | 0.3 ± 0.8 | 0.3 ± 0.8 | 0.7 ± 1.0 |
| Necrosis | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.7 ± 1.0 | 0.7 ± 1.0 |
| Indices | ||||
| Gill index (IG) | 8.9 ± 5.0 | 6.3 ± 3.3 | 9.7 ± 4.0 | 13.6 ± 8.4 |
| Liver index (IL) * | 0.3 ± 0.8 a | 4.0 ± 3.8 ab | 9.0 ± 4.3 b | 6.7 ± 5.9 ab |
| Total index (IT) * | 9.2 ± 5.0 a | 10.3 ± 5.2 ab | 18.7 ± 6.4 ab | 20.3 ± 10.5 b |
| Parameter | Group | |||
|---|---|---|---|---|
| C0 | C1 | C2 | C3 | |
| Micronuclei (‰) | 0.13 ± 0.11 | 0.00 ± 0.00 | 0.15 ± 0.15 | 0.00 ± 0.00 |
| Nuclear bud (‰) | 0.05 ± 0.09 | 0.08 ± 0.11 | 0.10 ± 0.09 | 0.23 ± 0.39 |
| Fragmented-apoptotic cell (‰) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.05 ± 0.09 | 0.38 ± 0.39 |
| Bi-nucleated cell (‰) | 0.07 ± 0.12 | 0.16 ± 0.01 | 0.31 ± 0.16 | 0.23 ± 0.26 |
| Total nuclear abnormalities (‰) | 0.30 ± 0.12 | 0.48 ± 0.44 | 0.46 ± 0.31 | 0.83 ± 0.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rašković, B.; Poleksić, V.; Vuković, G.; Špirović Trifunović, B.; Božić, G.; Ćupić Miladinović, D.; Marković, Z.; Brkić, D. Acute and Subchronic Exposure of the Common Carp (Cyprinus carpio) to Herbicide S-Metolachlor. Water 2023, 15, 4182. https://doi.org/10.3390/w15234182
Rašković B, Poleksić V, Vuković G, Špirović Trifunović B, Božić G, Ćupić Miladinović D, Marković Z, Brkić D. Acute and Subchronic Exposure of the Common Carp (Cyprinus carpio) to Herbicide S-Metolachlor. Water. 2023; 15(23):4182. https://doi.org/10.3390/w15234182
Chicago/Turabian StyleRašković, Božidar, Vesna Poleksić, Gorica Vuković, Bojana Špirović Trifunović, Gavrilo Božić, Dejana Ćupić Miladinović, Zoran Marković, and Dragica Brkić. 2023. "Acute and Subchronic Exposure of the Common Carp (Cyprinus carpio) to Herbicide S-Metolachlor" Water 15, no. 23: 4182. https://doi.org/10.3390/w15234182
APA StyleRašković, B., Poleksić, V., Vuković, G., Špirović Trifunović, B., Božić, G., Ćupić Miladinović, D., Marković, Z., & Brkić, D. (2023). Acute and Subchronic Exposure of the Common Carp (Cyprinus carpio) to Herbicide S-Metolachlor. Water, 15(23), 4182. https://doi.org/10.3390/w15234182