
Enhanced Post-Drought Compensatory Growth and Water Utilization in Maize via Rhizosphere Soil Nitrification by Heterotrophic Ammonia-Oxidizing Bacteria
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Experimental Grouping and Treatments
2.1.2. Timeline of the Experiment
2.1.3. Soil Moisture Control Methods
2.2. Measurements and Data Processing
2.2.1. Biomass and Photosynthesis Indicators
2.2.2. Soil Nitrification Rate and Zeatin Riboside Content
2.2.3. Quantitative Real-Time PCR
3. Results
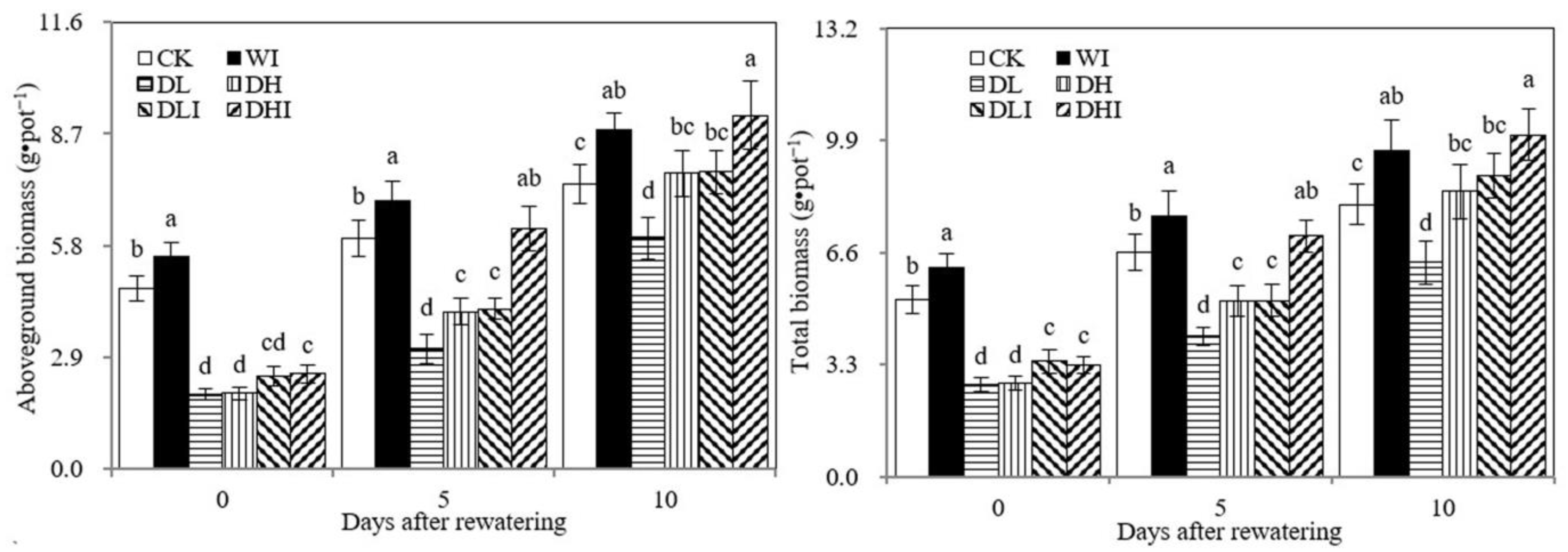
3.1. Changes in Biomass
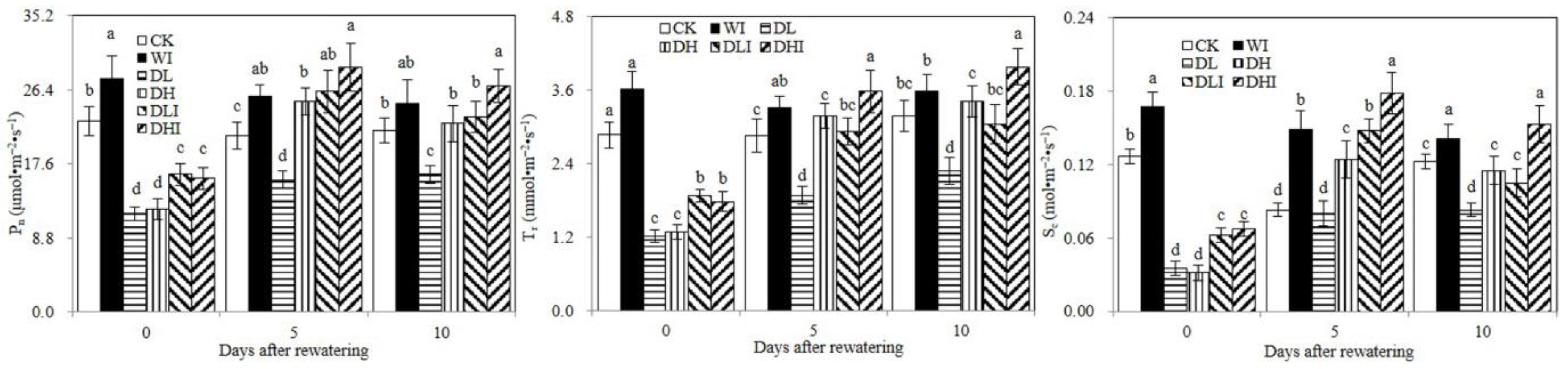
3.2. Photosynthesis Analysis
3.3. Changes in Soil Nitrification Rate and Soil Concentrations of NO3− and NH4+
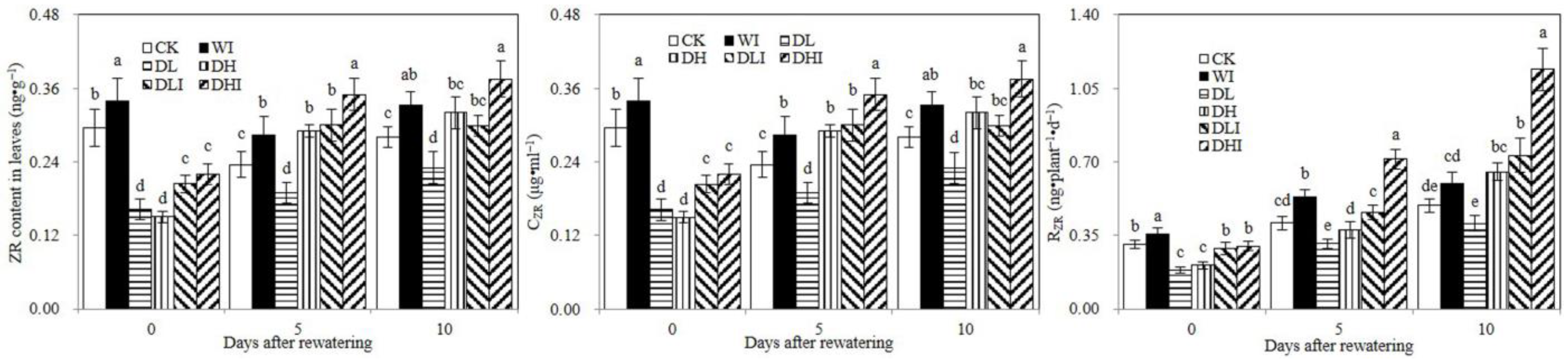
3.4. Changes of Zeatin Riboside
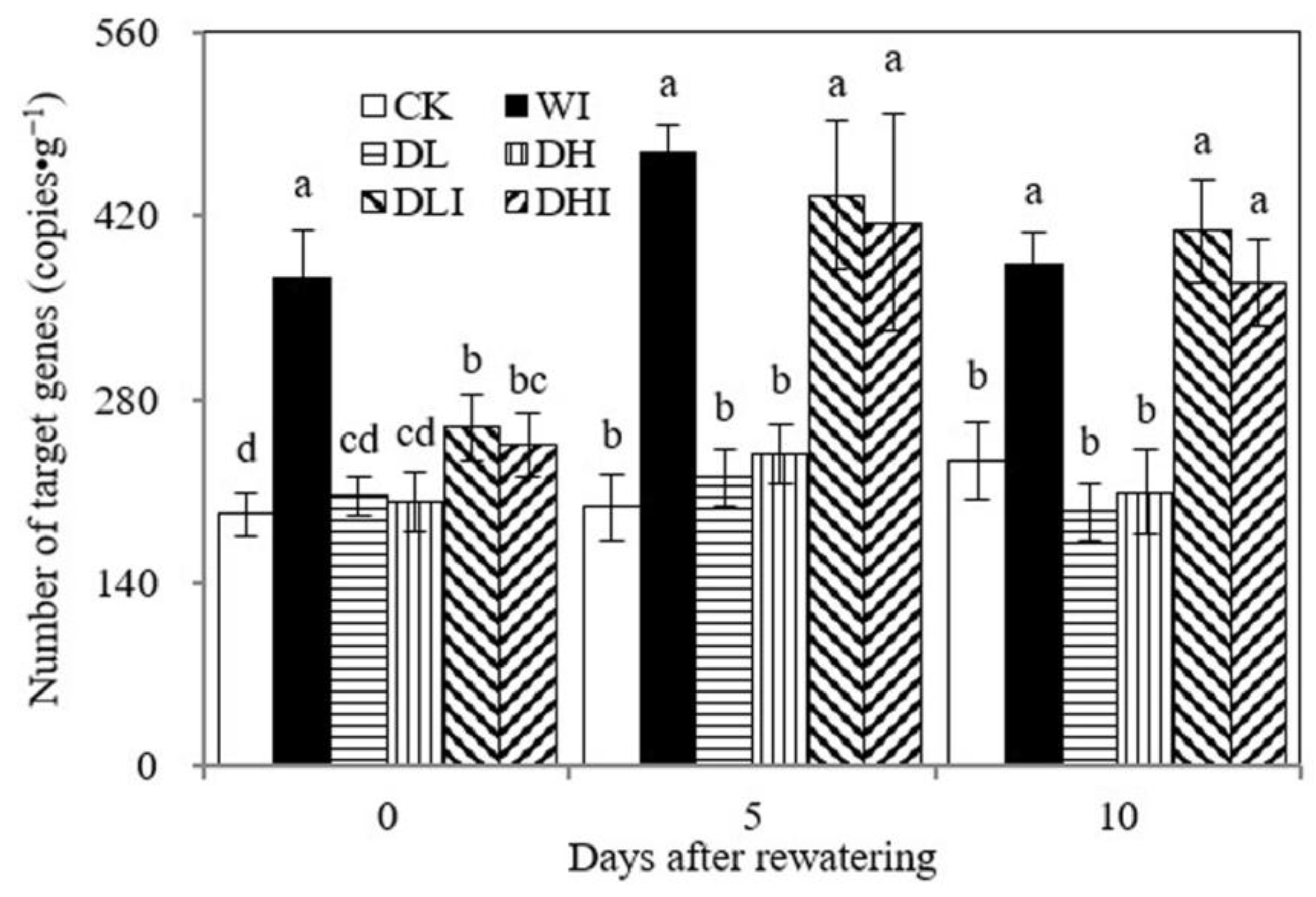
3.5. Copy Number Analysis
4. Discussion
4.1. Limited Rewatering Inhibiting Compensatory Growth
4.2. Limited Rewatering with HAOB Increasing Compensatory Growth
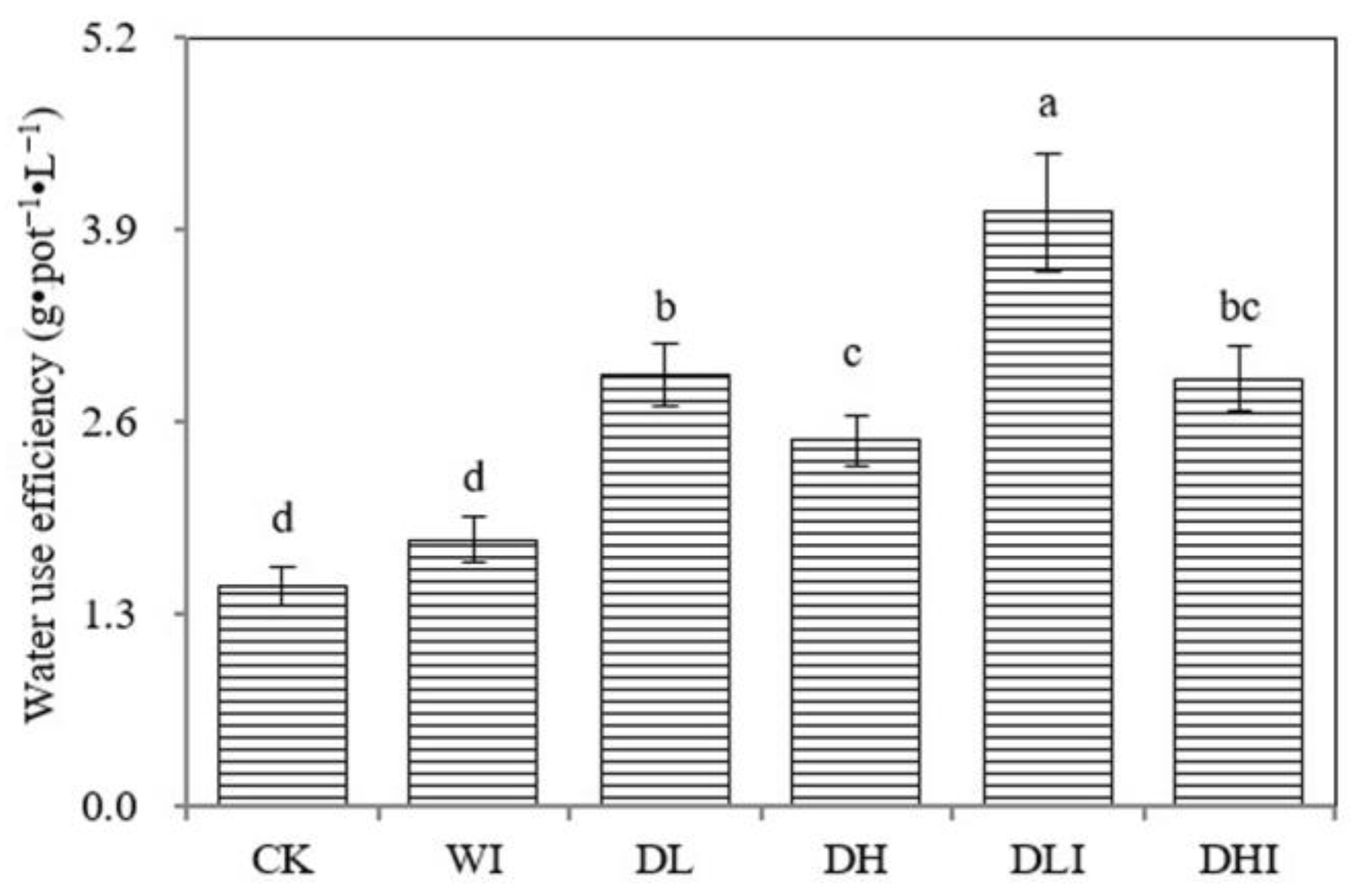
4.3. Achieving Efficient Water Use and Promoting Healthy Crop Growth
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kang, S.Z.; Hao, X.M.; Du, T.S.; Tong, L.; Su, X.L.; Lu, H.N.; Li, X.L.; Huo, Z.L.; Li, S.E.; Ding, R.S. Improving agricultural water productivity to ensure food security in China under changing environment: From research to practice. Agric. Water Manag. 2017, 179, 5–17. [Google Scholar] [CrossRef]
- Bicer, S.; Erdinc, C.; Comlekcioglu, N. A Comparative Analysis of Co-inoculation of Microbial Biostimulants at Different Irrigation Levels Under Field Conditions on the Cucumber Growth. Gesunde Pflanzen 2023, 75, 1237–1256. [Google Scholar] [CrossRef]
- Khajeeyan, R.; Salehi, A.; Dehnavi, M.M.; Farajee, H.; Kohanmoo, M.A. Physiological and yield responses of Aloe vera plant to biofertilizers under different irrigation regimes. Agric. Water Manag. 2019, 225, 105768. [Google Scholar] [CrossRef]
- Monino, M.J.; Blanco-Cipollone, F.; Vivas, A.; Bodelon, O.G.; Prieto, M.H. Evaluation of different deficit irrigation strategies in the late-maturing Japanese plum cultivar ‘Angeleno’. Agric. Water Manag. 2020, 234, 106111. [Google Scholar] [CrossRef]
- Sanfo, S.; Barbier, B.; Dabire, I.W.P.; Vlek, P.L.G.; Fonta, W.M.; Ibrahim, B.; Barry, B. Rainfall variability adaptation strategies: An ex-ante assessment of supplemental irrigation from farm ponds in southern Burkina Faso. Agric. Syst. 2017, 152, 80–89. [Google Scholar] [CrossRef]
- Peng, M.M.; Han, W.T.; Li, C.Q.; Li, G.; Yao, X.M.; Zhang, M.F. Diurnal and seasonal CO2 exchange and yield of maize cropland under different irrigation treatments in semiarid Inner Mongolia. Agric. Water Manag. 2021, 255, 107041. [Google Scholar] [CrossRef]
- Peng, F.; Lai, C.M.; Li, C.Y.; Ji, C.; Zhang, P.F.; Sun, J.B.; Chen, X.J.; You, Q.G.; Xue, X. Plasticity in over-compensatory growth along an alpine meadow degradation gradient on the Qinghai-Tibetan Plateau. J. Environ. Manag. 2023, 325, 116299. [Google Scholar] [CrossRef]
- Chen, G.D.; Kong, X.F.; Gan, Y.T.; Zhang, R.Z.; Feng, F.X.; Yu, A.H.; Zhao, C.; Wan, S.M.; Chai, Q. Enhancing the systems productivity and water use efficiency through coordinated soil water sharing and compensation in strip-intercropping. Sci. Rep. 2018, 8, 10494. [Google Scholar] [CrossRef]
- Tian, K.; Wang, Y.; Chen, D.; Cao, M.; Luo, J. Influence of Drought Stress and Post-Drought Rewatering on Phytoremediation Effect of Arabidopsis thaliana. Bull. Environ. Contam. Toxicol. 2022, 108, 594–599. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chang, Y.S.; Lin, L.H. Response of shoot growth, photosynthetic capacity, flowering, and fruiting of potted ‘Nagami’ kumquat to different regulated deficit irrigation. Hortic. Environ. Biota. 2015, 56, 444–454. [Google Scholar] [CrossRef]
- Puertolas, J.; Albacete, A.; Dodd, I.C. Irrigation frequency transiently alters whole plant gas exchange, water and hormone status, but irrigation volume determines cumulative growth in two herbaceous crops. Environ. Exp. Bot. 2020, 176, 104101. [Google Scholar] [CrossRef]
- Maswada, H.F.; Abd El-Razek, U.A.; El-Sheshtawy, A.N.A.; Mazrou, Y.S.A. Effect of Azolla filiculoides on Growth, Physiological and Yield Attributes of Maize Grown under Water and Nitrogen Deficiencies. J. Plant Growth Regul. 2021, 40, 558–573. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Wang, P.Y.; Xiong, X.B.; Zhou, R.; Zhu, Y.; Wang, Y.B.; Wang, N.; Wesly, K.; Xue, W.; Cao, J.; et al. Can shallow-incorporated organic mulching replace plastic film mulching for irrigated maize production systems in arid environments? Field Crop Res. 2023, 297, 108931. [Google Scholar] [CrossRef]
- Chowdhury, S.; Thangarajan, R.; Bolan, N.; O’Reilly-Wapstra, J.; Kunhikrishnan, A.; Naidu, R. Nitrification potential in the rhizosphere of Australian native vegetation. Soil Res. 2017, 55, 58–69. [Google Scholar] [CrossRef]
- Beeckman, F.; Motte, H.; Beeckman, T. Nitrification in agricultural soils: Impact, actors and mitigation. Curr. Opin. Biotech. 2018, 50, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Wang, J.J.; Sun, R.H.; Hou, X.G.; Zhao, W.; Shi, J.; Zhang, Y.F.; Qi, L.; Li, X.L.; Dong, P.H.; et al. Correlation of the maize compensatory growth mechanism after post-drought rewatering with cytokinin induced by root nitrate absorption. Agric. Water Manag. 2016, 166, 77–85. [Google Scholar] [CrossRef]
- Wang, X.L.; Ma, K.; Qi, L.; Liu, Y.H.; Shi, J.; Li, X.L.; Zhang, L.X.; Liu, W.; Song, P. Effect of ammonia-oxidizing bacterial strain that survives drought stress on maize compensatory growth upon post-drought rewatering. Front. Plant Sci. 2022, 13, 947476. [Google Scholar] [CrossRef]
- Wu, D.; Wang, X.L.; Zhu, X.H.; Wang, H.H.; Liu, W.; Qi, L.; Song, P.; Zhang, M.M.; Zhao, W. Effect of Ammonia-oxidizing bacterial strains that coexist in rhizosphere soil on Italian ryegrass regrowth. Microorganisms 2022, 10, 2122. [Google Scholar] [CrossRef]
- Zou, J.; Snider, J.L.; Zhu, H.H.; He, J.Q.; Li, Y.X.; Zhou, Z.G.; Wang, Y.H.; Meng, Y.L.; Chen, B.L.; Zhao, W.Q.; et al. Soil drought duration and severity affect cotton boll biomass by altering recovery times and carbon dynamics of subtending leaf. J. Agron. Crop Sci. 2022, 209, 94–115. [Google Scholar] [CrossRef]
- Wang, X.L.; Qin, R.R.; Sun, R.H.; Wang, J.J.; Hou, X.G.; Qi, L.; Qin, D.H. No post-drought compensatory growth of corns with root cutting based on cytokinin induced by roots. Agric. Water Manag. 2018, 205, 9–20. [Google Scholar] [CrossRef]
- Wang, X.L.; Duan, P.L.; Sun, R.H.; Qi, L.; Shi, J.; Li, X.L.; Zhang, L.X. Effects of Soil nitrification on compensatory growth upon post-drought rewatering of maizes based on cytokinin. Int. J. Agric. Biol. 2020, 23, 882–888. [Google Scholar]
- Qin, R.R.; Wang, X.L. Effects of crown height on the compensatory growth of Italian ryegrass based on combined effects of stored organic matter and cytokinin. Grassl. Sci. 2020, 66, 29–39. [Google Scholar] [CrossRef]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmulling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, eaba0196. [Google Scholar] [CrossRef]
- Zaicovski, C.B.; Zimmerman, T.; Nora, L.; Nora, F.R.; Silva, J.A.; Rombaldi, C.V. Water stress increases cytokinin biosynthesis and delays postharvest yellowing of broccoli florets. Postharvest Biol. Technol. 2008, 49, 436–439. [Google Scholar] [CrossRef]
- Kobayashi, H.; Inoue, S.; Gyokusen, K. Photosynthesis-nitrogen relationship in a Hinoki cypress (Chamaecyparis obtusa) canopy: A comparison with Japanese cedar (Cryptomeria japonica). Photosynthetica 2012, 50, 317–320. [Google Scholar] [CrossRef]
- De Moura, F.B.; Da, S.; Vieira, M.R.; Simões, A.D.N.; da Silva, F.S.L.; De Medeiros, D.C.; Paes, R.D.A.; de Oliveira, S.A.A.; do Nascimento, C.A.H.; Júnior, E.W.S. Participation of cytokinin on gas exchange and antioxidant enzymes activities. Indian J. Plant Physiol. 2017, 22, 16–29. [Google Scholar] [CrossRef]
- Wu, W.Q.; Du, K.; Kang, X.Y.; Wei, H.R. The diverse roles of cytokinins in regulating leaf development. Hortic. Res. 2021, 8, 118. [Google Scholar] [CrossRef]
- Poudel, M.; Mendes, R.; Costa, L.A.S.; Bueno, C.G.; Meng, Y.; Folimonova, S.Y.; Garrett, K.A.; Martins, S.J. The Role of Plant-Associated Bacteria, Fungi, and Viruses in Drought Stress Mitigation. Front. Microbiol. 2021, 12, 743512. [Google Scholar] [CrossRef]
- Hu, H.W.; Macdonald, C.A.; Trivedi, P.; Holmes, B.; Bodrossy, L.; He, J.Z.; Singh, B.K. Water addition regulates the metabolic activity of ammonia oxidizers responding to environmental perturbations in dry subhumid ecosystems. Environ. Microbiol. 2015, 17, 444–461. [Google Scholar] [CrossRef]
- Keshavarz, H. Study of water deficit conditions and beneficial microbes on the oil quality and agronomic traits of canola (Brassica napus L.). Grasas Aceites 2020, 71, e373. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Definition | Symbol | Definition |
|---|---|---|---|
| HAOB | Heterotrophic ammonia-oxidizing bacteria | Sc | Stomatal conductance |
| CK | Control with sufficient Water | Tr | Transpiration rate |
| WI | Sufficient water + HAOB strain | ZR | Zeatin riboside |
| DL | Limited rewatering | RZR | Delivery rate of ZR from roots to leaves |
| DH | Sufficient rewatering | CZR | ZR concentration in xylem sap |
| DHI | Sufficient rewatering + HAOB strain | Bt | Temporary weight of the whole pot |
| DLI | Limited rewatering + HAOB strain | Bd | Net weight of dried soil of pot |
| NO3− | Soil nitrate | Be | Weight of the empty pot |
| NH4+ | Soil ammonium | FWC | Field water capacity |
| Pn | Photosynthetic rate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-L.; Tian, S.-S.; Yu, H.; Sun, R.-H.; Qi, L.; Song, P.; Yang, S.-J. Enhanced Post-Drought Compensatory Growth and Water Utilization in Maize via Rhizosphere Soil Nitrification by Heterotrophic Ammonia-Oxidizing Bacteria. Water 2023, 15, 3933. https://doi.org/10.3390/w15223933
Wang X-L, Tian S-S, Yu H, Sun R-H, Qi L, Song P, Yang S-J. Enhanced Post-Drought Compensatory Growth and Water Utilization in Maize via Rhizosphere Soil Nitrification by Heterotrophic Ammonia-Oxidizing Bacteria. Water. 2023; 15(22):3933. https://doi.org/10.3390/w15223933
Chicago/Turabian StyleWang, Xiao-Ling, Shi-Shi Tian, Hao Yu, Run-Hong Sun, Lin Qi, Peng Song, and Shen-Jiao Yang. 2023. "Enhanced Post-Drought Compensatory Growth and Water Utilization in Maize via Rhizosphere Soil Nitrification by Heterotrophic Ammonia-Oxidizing Bacteria" Water 15, no. 22: 3933. https://doi.org/10.3390/w15223933