
Effects of Massive Use of Disinfectants on the Plankton Communities in Lakes from Wuhan

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
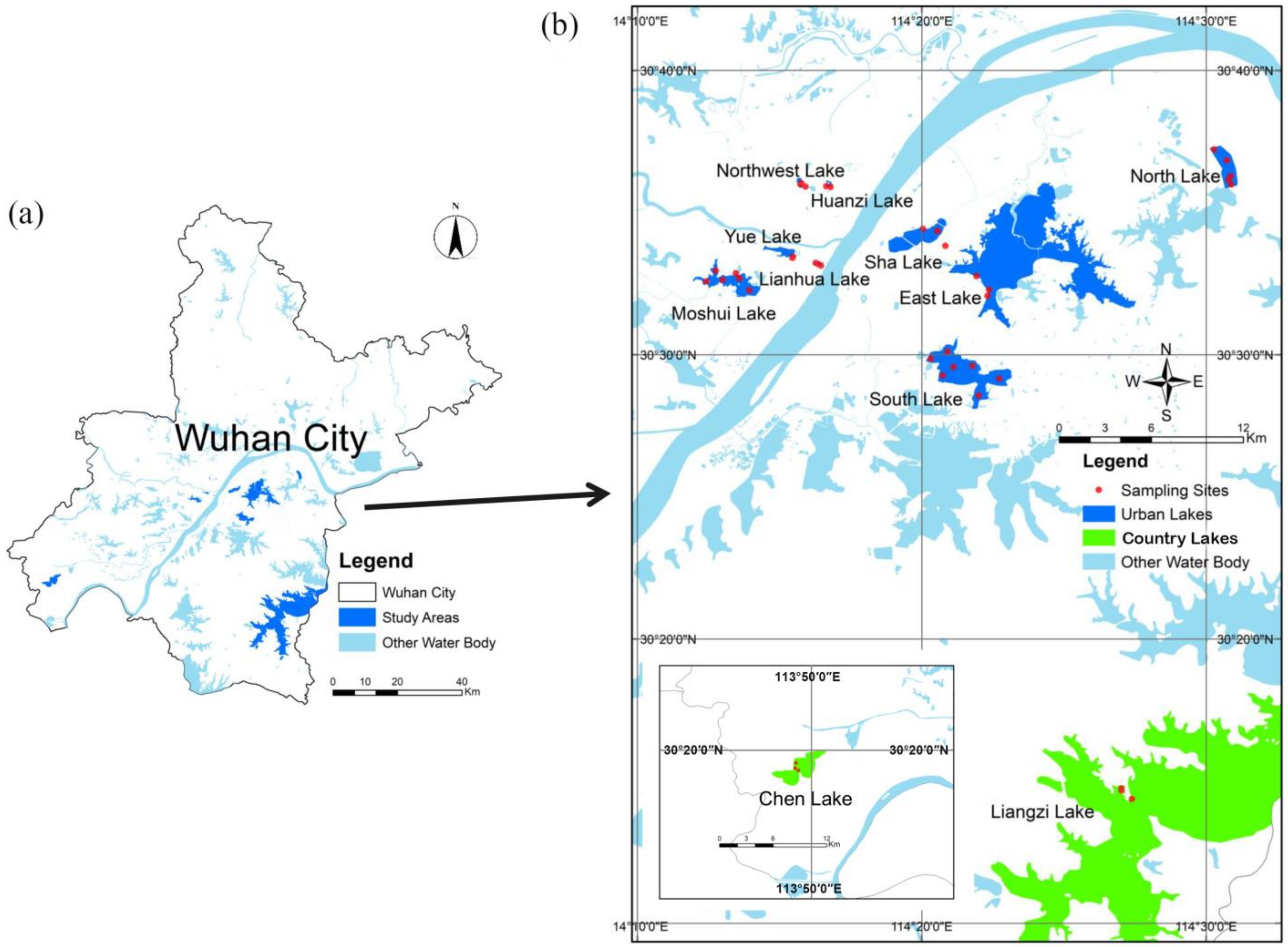
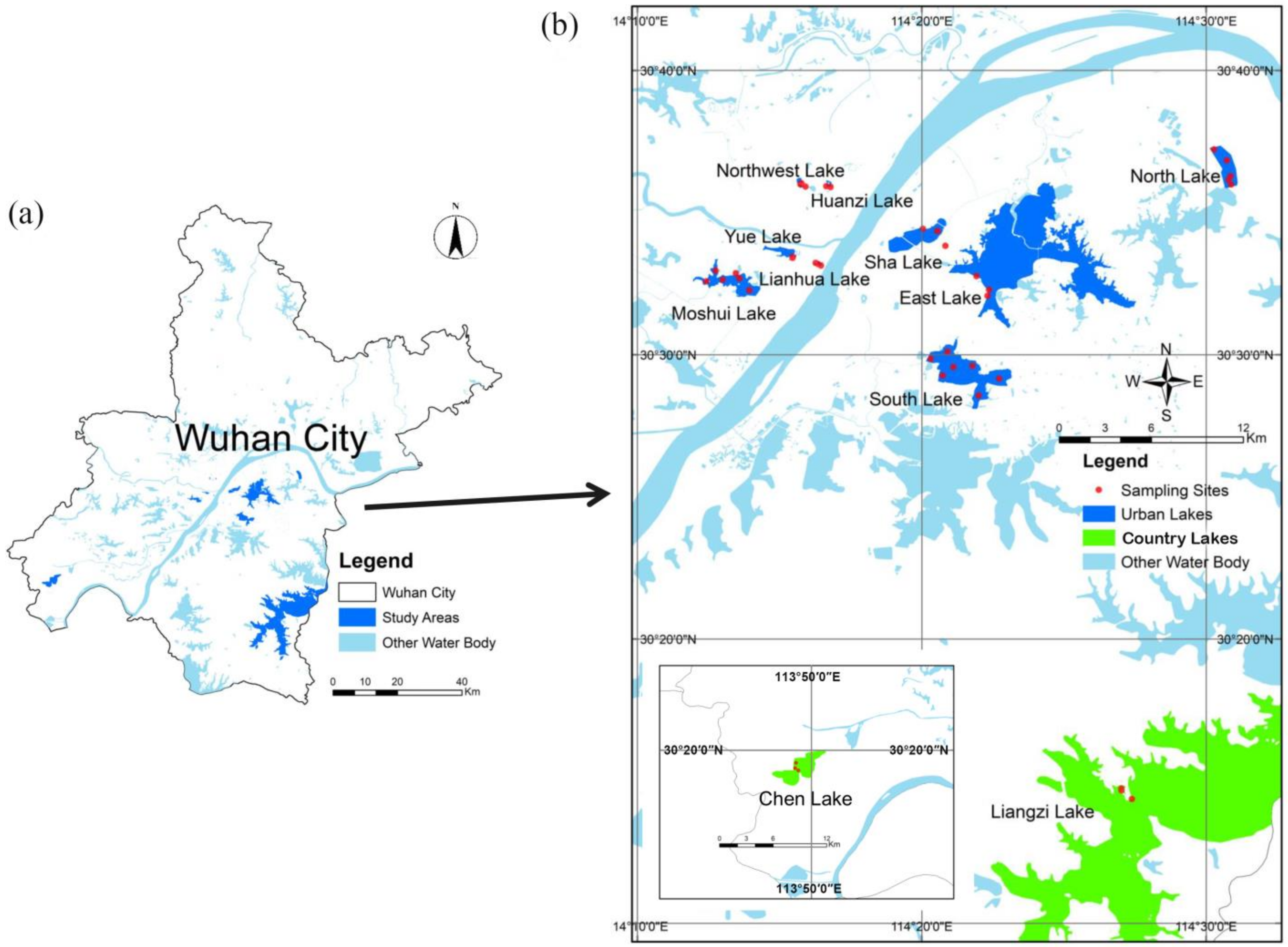
2.1. Sample Collection
2.2. Environmental Variables
2.3. Phytoplankton Analysis
2.4. Zooplankton Analysis
2.5. Statistical Analyses
3. Results
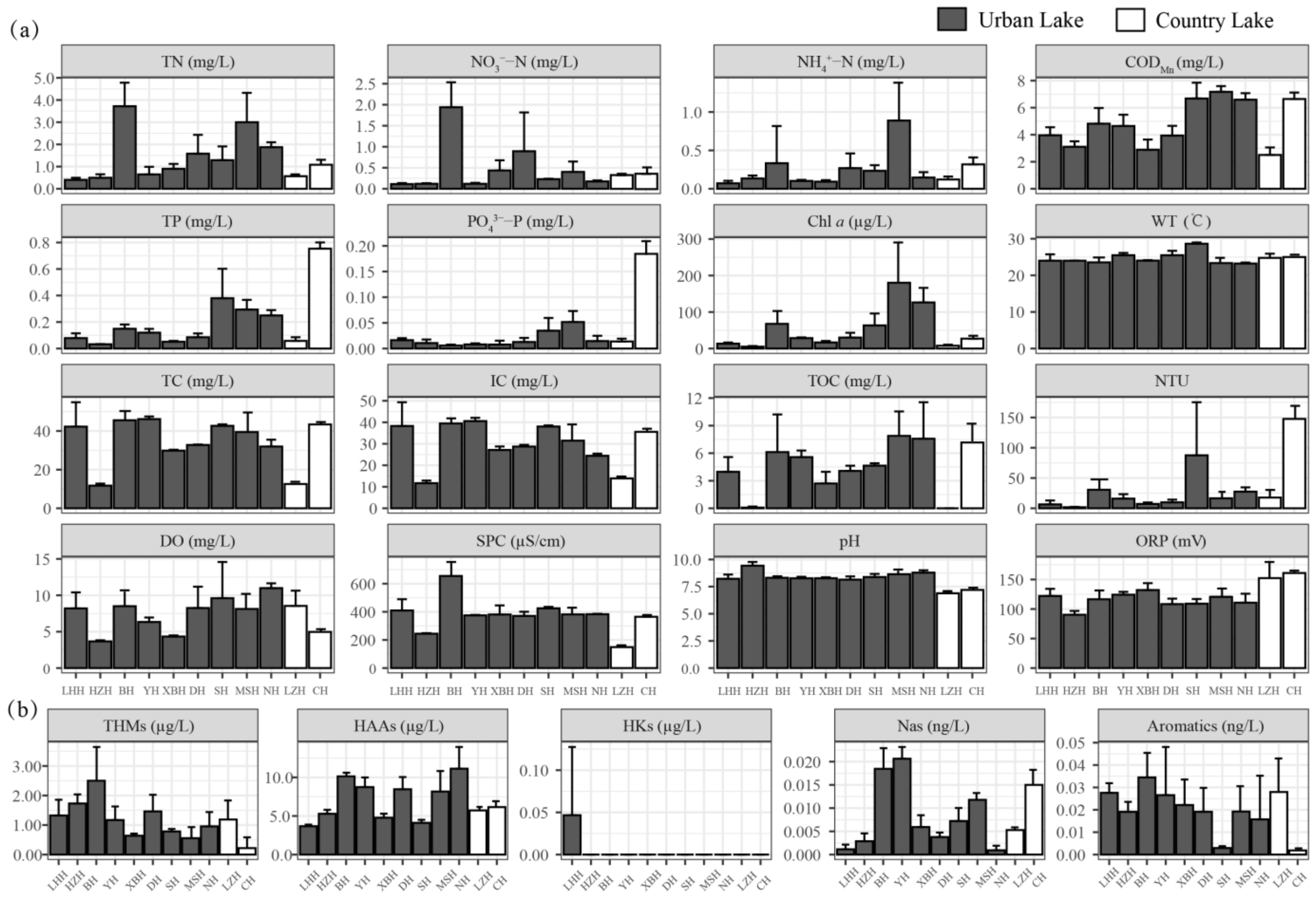
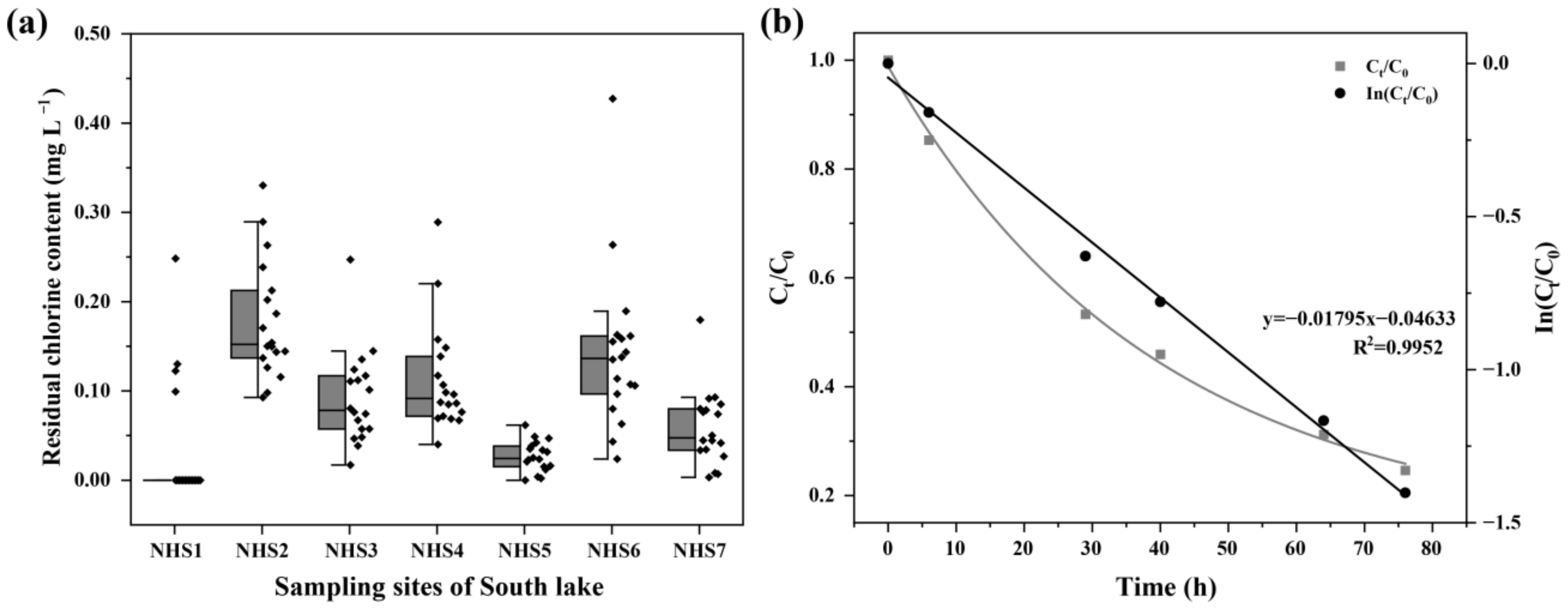
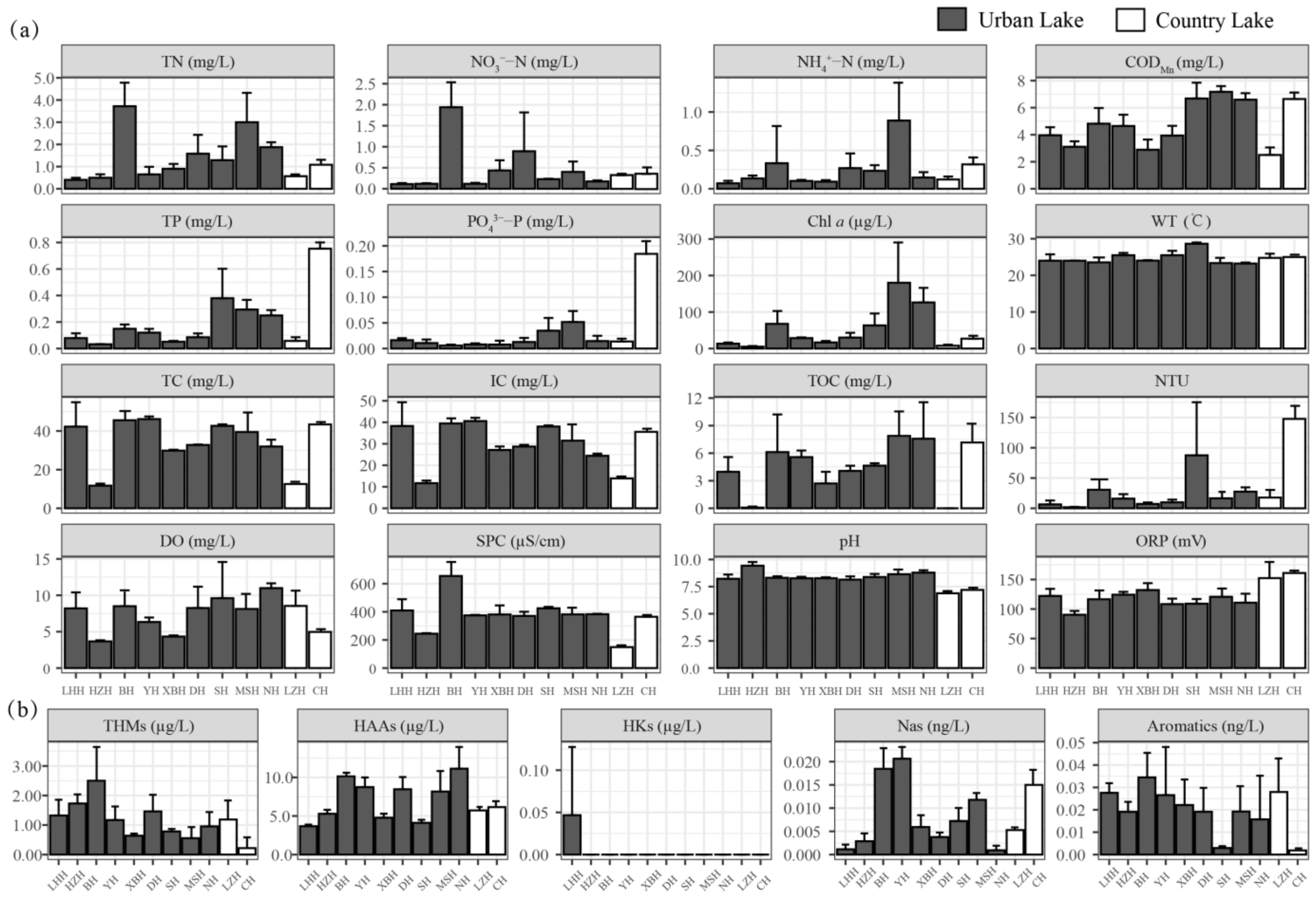
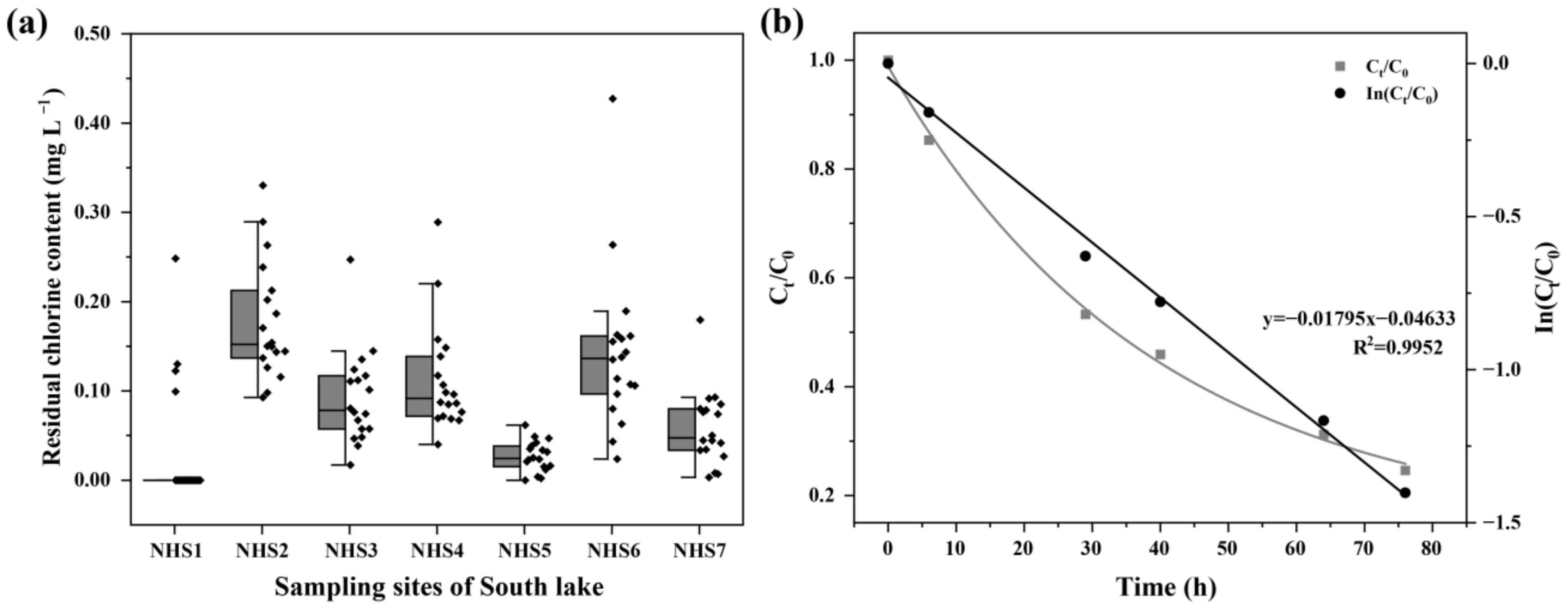
3.1. Variations in Environmental Factors
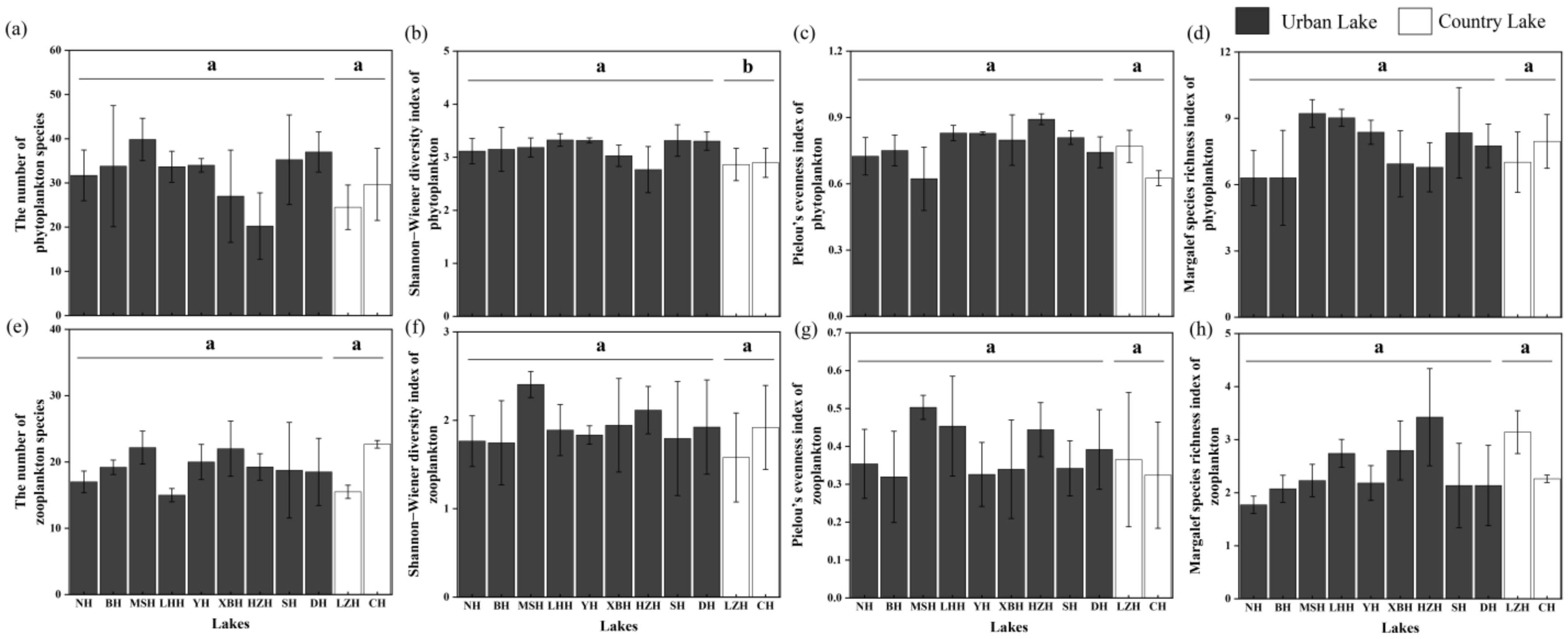
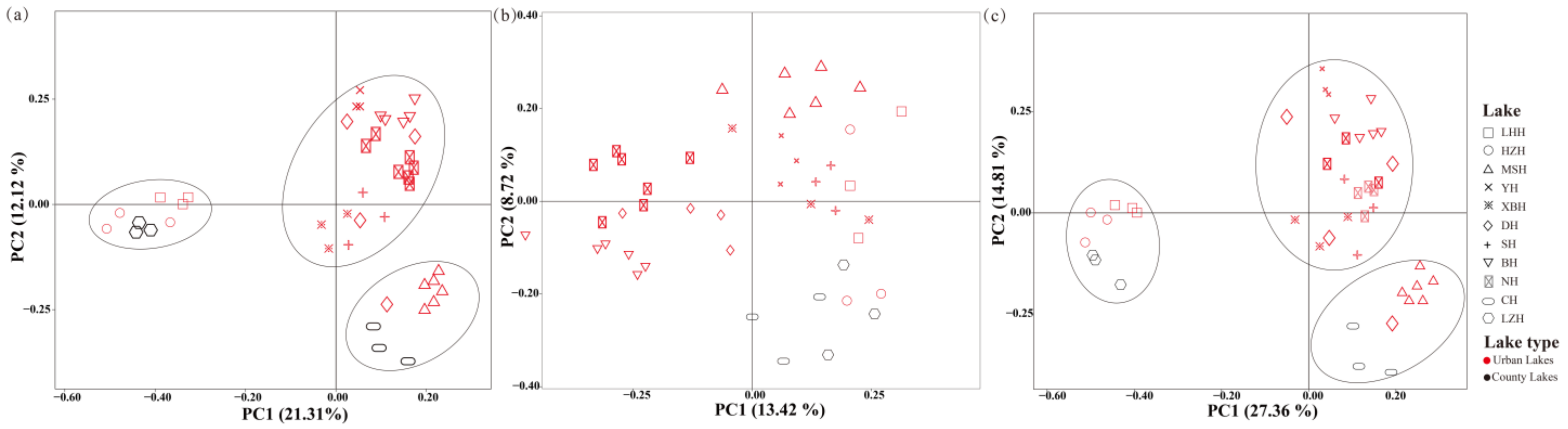
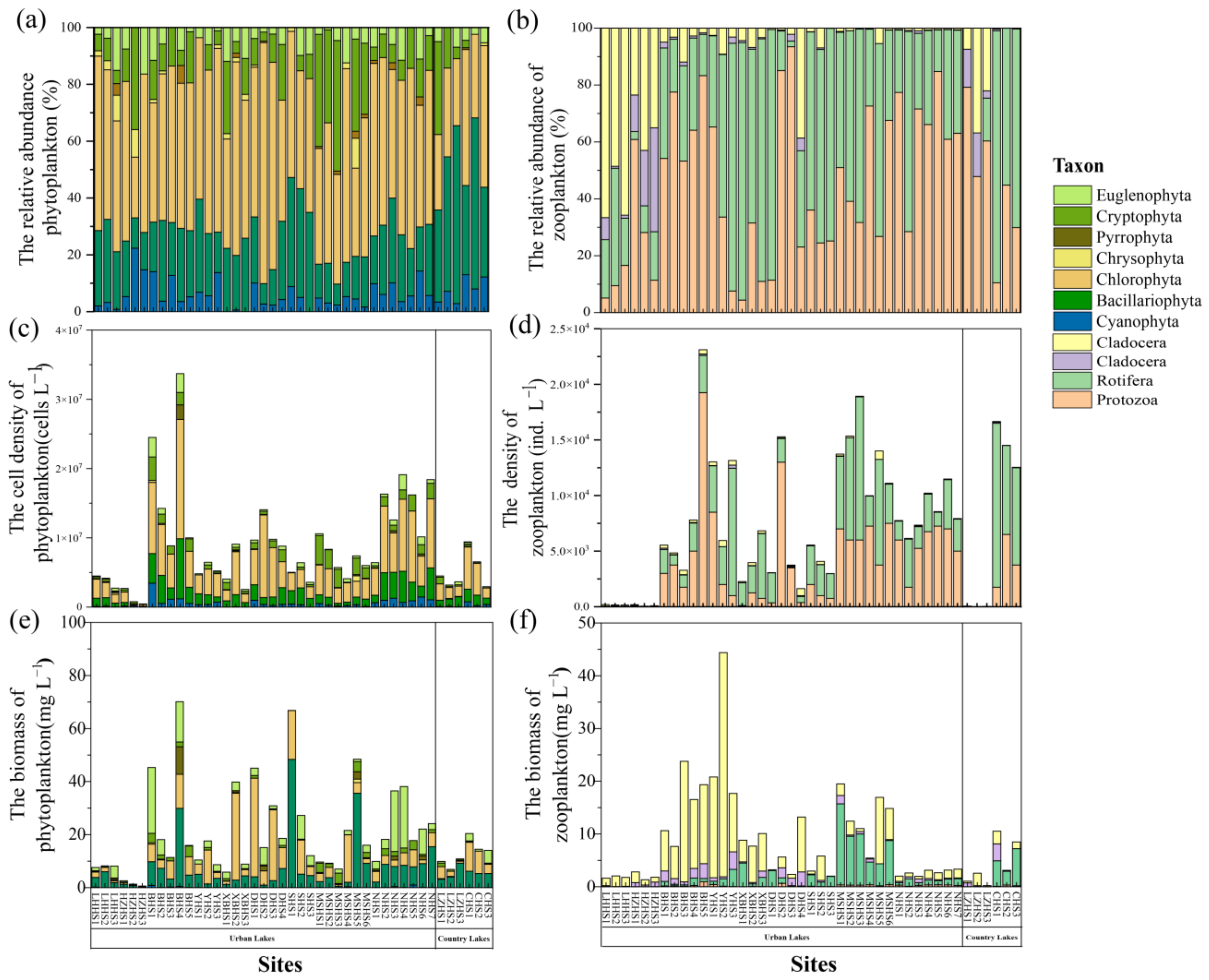
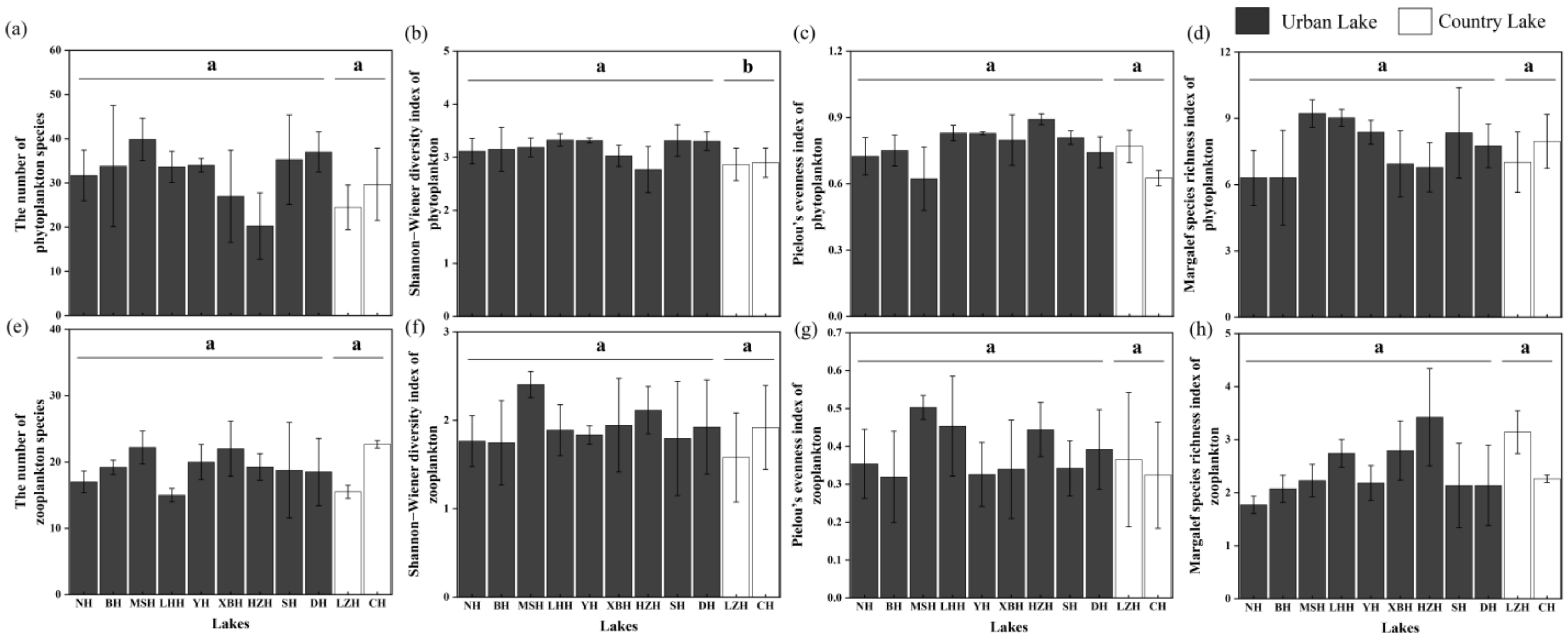
3.2. Spatial Variation of Phytoplankton and Zooplankton Community
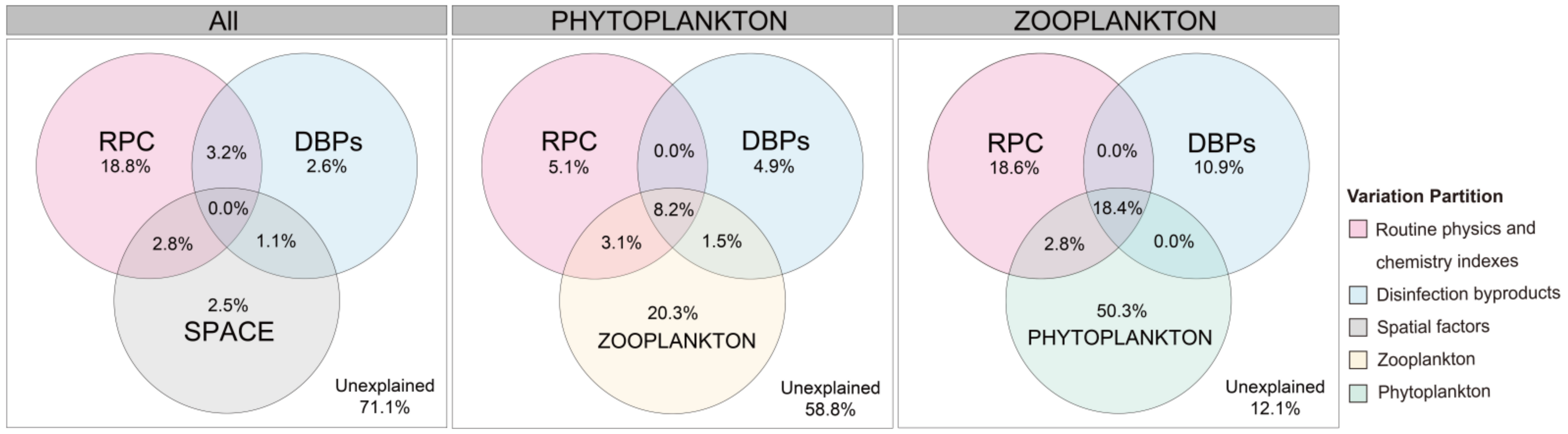
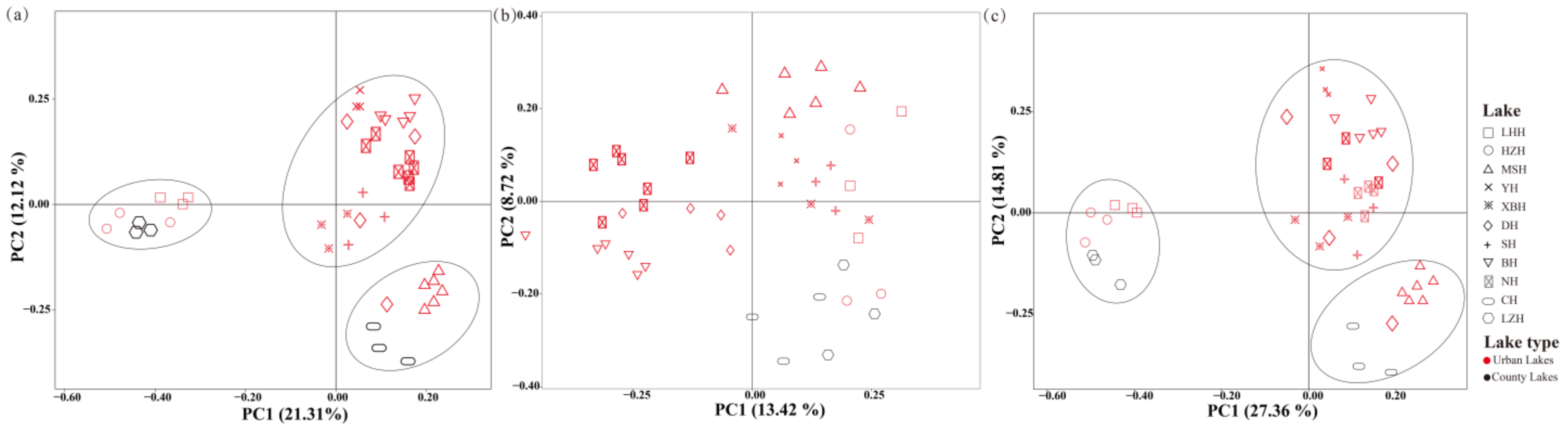
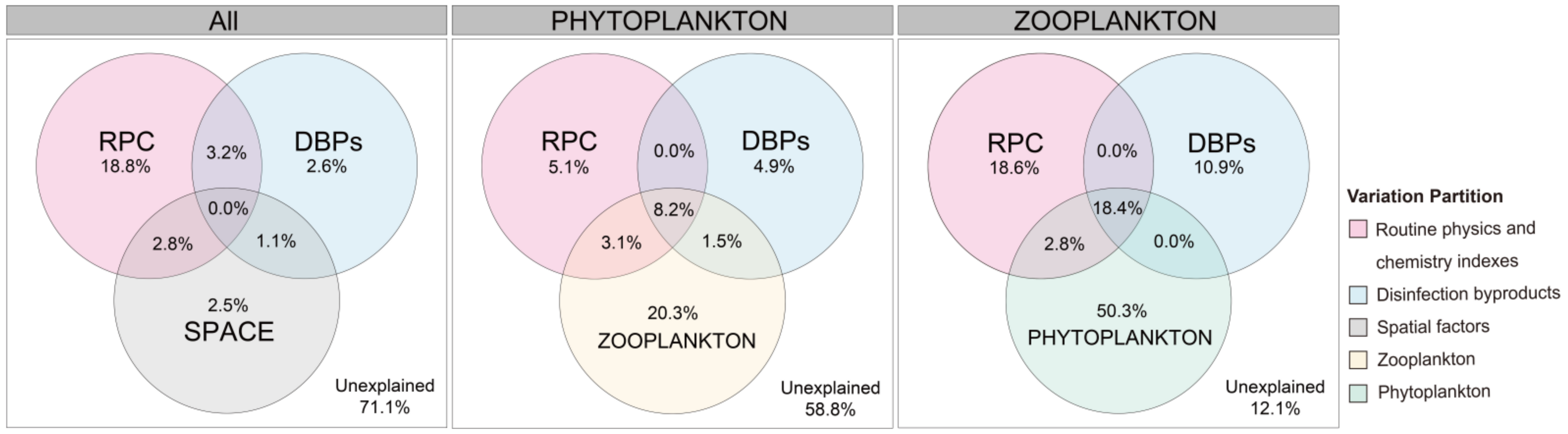
3.3. Community Assembly of Phytoplankton and/or Zooplankton
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fang, D.; Yang, H.; Zhang, H.; Du, H.; Yang, J.; Wei, Q. COVID-19 Lockdowns Promoted Recovery of the Yangtze River’s Aquatic Ecosystem. Water 2022, 14, 3622. [Google Scholar] [CrossRef]
- Sarkodie, S.A.; Owusu, P.A. Global assessment of environment, health and economic impact of the novel coronavirus (COVID-19). Environ. Dev. Sustain. 2021, 23, 5005–5015. [Google Scholar] [CrossRef] [PubMed]
- Tisdell, C.A. Economic, social and political issues raised by the COVID-19 pandemic. Econ. Anal. Policy 2020, 68, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Subpiramaniyam, S. Outdoor disinfectant sprays for the prevention of COVID-19: Are they safe for the environment? Sci. Total Environ. 2021, 759, 144289. [Google Scholar] [CrossRef]
- Dadras, O.; Alinaghi, S.A.S.; Karimi, A.; MohsseniPour, M.; Barzegary, A.; Vahedi, F.; Pashaei, Z.; Mirzapour, P.; Fakhfouri, A.; Zargari, G.; et al. Effects of COVID-19 prevention procedures on other common infections: A systematic review. Eur. J. Med. Res. 2021, 26, 67. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ling, H.; Huang, X.; Li, J.; Li, W.; Yi, C.; Zhang, T.; Jiang, Y.; He, Y.; Deng, S.; et al. Potential spreading risks and disinfection challenges of medical wastewater by the presence of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) viral RNA in septic tanks of Fangcang Hospital. Sci. Total Environ. 2020, 741, 140445. [Google Scholar] [CrossRef]
- Song, G.; Zhu, Y.; Mi, W.; Ajiayan, A.; Bi, Y. Characteristics of zooplankton community and their influencing factors in lakes of Wuhan. J. Henna Norm. Univ. Nat. Sci. Ed. 2022, 50, 1135–1142. (In Chinese) [Google Scholar]
- Zhang, H.; Tang, W.; Chen, Y.; Yin, W. Disinfection threatens aquatic ecosystems. Science 2020, 368, 146–147. [Google Scholar] [CrossRef]
- Hora, P.I.; Pati, S.G.; McNamara, P.J.; Arnold, W.A. Increased Use of Quaternary Ammonium Compounds during the SARS-CoV-2 Pandemic and Beyond: Consideration of Environmental Implications. Environ. Sci. Technol. Lett. 2020, 7, 622–631. [Google Scholar] [CrossRef]
- Wang, J.; Shen, J.; Ye, D.; Yan, X.; Zhang, Y.; Yang, W.; Li, X.; Wang, J.; Zhang, L.; Pan, L. Disinfection technology of hospital wastes and wastewater: Suggestions for disinfection strategy during coronavirus Disease 2019 (COVID-19) pandemic in China. Environ. Pollut. 2020, 262, 114665. [Google Scholar] [CrossRef]
- Nardell, E.A.; Nathavitharana, R.R. Airborne Spread of SARS-CoV-2 and a Potential Role for Air Disinfection. JAMA 2020, 324, 141–142. [Google Scholar] [CrossRef] [PubMed]
- Nabi, G.; Wang, Y.; Hao, Y.; Khan, S.; Wu, Y.; Li, D. Massive use of disinfectants against COVID-19 poses potential risks to urban wildlife. Environ. Res. 2020, 188, 109916. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Fang, C.; Deng, Y.; Xu, Z. Intensified Disinfection Amid COVID-19 Pandemic Poses Potential Risks to Water Quality and Safety. Environ. Sci. Technol 2021, 55, 4084–4086. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, D.L.; von Gunten, U. The Chlorine Dilemma. Science 2011, 331, 42–43. [Google Scholar] [CrossRef]
- He, J.; Shi, M.; Wang, F.; Duan, Y.; Zhao, T.; Shu, S.; Chu, W. Removal of CX3R-type disinfection by-product precursors from rainwater with conventional drinking water treatment processes. Water Res. 2020, 185, 116099. [Google Scholar] [CrossRef]
- Huang, H.; Wu, Q.; Hu, H.; Mitch, W.A. Dichloroacetonitrile and Dichloroacetamide Can Form Independently during Chlorination and Chloramination of Drinking Waters, Model Organic Matters, and Wastewater Effluents. Environ. Sci. Technol. 2012, 19, 10624–10631. [Google Scholar] [CrossRef]
- Krasner, S.W.; Westerhoff, P.; Chen, B.; RIttmann, B.E.; Amy, G. Occurrence of Disinfection Byproducts in United States Wastewater Treatment Plant Effluents. Environ. Sci. Technol. 2009, 21, 8320–8325. [Google Scholar] [CrossRef]
- Li, X.; Mitch, W.A. Drinking Water Disinfection Byproducts (DBPs) and Human Health Effects: Multidisciplinary Challenges and Opportunities. Environ. Sci. Technol. 2018, 52, 1681–1689. [Google Scholar] [CrossRef]
- Reungoat, J.; Escher, B.I.; Macova, M.; Argaud, F.X.; Gernjak, W.; Keller, J. Ozonation and biological activated carbon filtration of wastewater treatment plant effluents. Water Res. 2012, 46, 863–872. [Google Scholar] [CrossRef]
- Wang, W.; Ye, B.; Yang, L.; Li, Y.; Wang, Y. Risk assessment on disinfection by-products of drinking water of different water sources and disinfection processes. Environ. Int. 2007, 33, 219–225. [Google Scholar] [CrossRef]
- Gong, T.; Tao, Y.; Xian, Q. Selection and applicability of quenching agents for the analysis of polar iodinated disinfection byproducts. Chemosphere 2016, 163, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.J.; Chen, B.Y.; Jiang, Y.L.; Tao, Y.; Zhu, X.S.; Cai, Z.H. Toxicity of 17 Disinfection By-products to Different Trophic Levels of Aquatic Organisms: Ecological Risks and Mechanisms. Environ. Sci. Technol. 2021, 55, 10534–10541. [Google Scholar] [CrossRef]
- Hanson, M.L.; Solomon, K.R. Haloacetic acids in the aquatic environment. Part I: Macrophyte toxicity. Environ. Pollut. 2004, 130, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Zhu, X.; Zhu, Y.; Huang, Y.; Chen, B. Ecotoxicological effects of DBPs on freshwater phytoplankton communities in co-culture systems. J. Hazard. Mater. 2022, 421, 126679. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Song, G.; Bi, Y.; Gao, W.; He, A.; Lu, Y.; Wang, Y.; Jiang, G. Occurrence and Distribution of Disinfection Byproducts in Domestic Wastewater Effluent, Tap Water, and Surface Water during the SARS-CoV-2 Pandemic in China. Environ. Sci. Technol. 2021, 55, 4103–4114. [Google Scholar] [CrossRef]
- Liu, X.Y.; Chen, L.; Yang, M.T.; Tan, C.Q.; Chu, W.H. The occurrence, characteristics, transformation and control of aromatic disinfection by-products: A review. Water Res. 2020, 184, 116076. [Google Scholar] [CrossRef]
- Howeth, J.G.; Leibold, M.A. Species dispersal rates alter diversity and ecosystem stability in pond metacommunities. Ecology 2010, 91, 2727–2741. [Google Scholar] [CrossRef]
- Zwart, J.A.; Solomon, C.T.; Jones, S.E. Phytoplankton traits predict ecosystem function in a global set of lakes. Ecology 2015, 96, 2257–2264. [Google Scholar] [CrossRef]
- Zhao, K.; Wang, L.; You, Q.; Pan, Y.; Liu, T.; Zhou, Y.; Zhang, J.; Pang, W.; Wang, Q. Influence of cyanobacterial blooms and environmental variation on zooplankton and eukaryotic phytoplankton in a large, shallow, eutrophic lake in China. Sci. Total Environ. 2021, 773, 145421. [Google Scholar] [CrossRef]
- Zhao, K.; Wang, L.; Riseng, C.; Wehrly, K.; Pan, Y.; Song, K.; Da, L.; Pang, W.; You, Q.; Tian, H.; et al. Factors determining zooplankton assemblage difference among a man-made lake, connecting canals, and the water-origin river. Ecol. Indic. 2018, 84, 488–496. [Google Scholar] [CrossRef]
- Roberts, J.F.; van Egmond, R.; Price, O.R. Toxicity of haloacetic acids to freshwater algae. Ecotoxicol. Environ. Saf. 2010, 73, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yang, J.; Lv, H.; Yu, X.; Wilkinson, D.M.; Yang, J. Phytoplankton Communities Exhibit a Stronger Response to Environmental Changes than Bacterioplankton in Three Subtropical Reservoirs. Environ. Sci. Technol. 2015, 49, 10850–10858. [Google Scholar] [CrossRef]
- Yi, J.P.; Huang, X.; Hou, J.; Xiong, J.W.; Qian, Z.; Liu, S.; Zhang, J.Q.; Yin, D.C.; Li, J.Y.; Su, Q.K.; et al. Occurrence and distribution of PPCPs in water from two largest urban lakes of China: First perspective from DGT in-situ measurement. Sci. Total Environ. 2023, 904, 166656. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Peng, X.X.; Ming, X.Y.; Huang, X.N.; Gu, K.D. Survey of Microcystins-LR Levels in Lakes in Wuhan, China. J. Environ. Health 2011, 28, 142–144. [Google Scholar]
- Zhao, Z.W.; He, W.; Wu, R.L.; Xu, F.L. Distribution and Relationships of Polycyclic Aromatic Hydrocarbons (PAHs) in Soils and Plants near Major Lakes in Eastern China. Toxics 2022, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Tong, W. Research of Land Use Change and Scenario Simulation in Wuhan Based on Time Series Landsat Images. Master’s Thesis, China University of Minning and Technology, Xuzhou, China, 2021. [Google Scholar]
- Wang, C.; Li, Q.; Ge, F.; Hu, Z.; He, P.; Chen, D.; Xu, D.; Wang, P.; Zhang, Y.; Zhang, L.; et al. Responses of aquatic organisms downstream from WWTPs to disinfectants and their by-products during the COVID-19 pandemic, Wuhan. Sci. Total Environ. 2022, 818, 151711. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- Zhang, Z.S.; Huang, X.F. Research Methods of Freshwater Plankton; Beijing Press: Beijing, China, 1991; pp. 333–362. [Google Scholar]
- McBurney, J.G.R. Measurement of invertebrate biomass by volume. N. Z. Entomol. 1978, 6, 428–429. [Google Scholar] [CrossRef]
- Gao, S.B.; Chi, S.Y.; Li, S.X.; Zheng, J.X.; Dong, F.Y. Fish resource investigation and species diversity analysis of Nanxi River in Zhejiang Province. J. Hydroecol. 2017, 38, 72–81. [Google Scholar]
- Zhou, J.; Ning, D. Stochastic community assembly: Does it matter in microbial ecology? Microbiol. Mol. Biol. Rev. 2017, 81, e00002–e00017. [Google Scholar] [CrossRef]
- Luo, Y.; Liu, N.; Sun, S.W.; Hou, S.; Guo, C.S.; Xu, J. Occurrence and ecological risk of typical DBPs in Chinese surface water. China Environ. Sci. 2021, 41, 1806–1814. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Drinking Water Standards and Health Advisories; U.S. Environmental Protection Agency: Washington, DC, USA, 2018. [Google Scholar]
- Jin, W.; Zhou, J.; Chen, B.; Zhu, X.; Cui, C. Modeling volatilization and adsorption of disinfection byproducts in natural watersheds. J. Environ. Monitor. 2012, 14, 2990–2999. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hu, L.; Deng, W.; Ying, G.; Hong, H.; Eric, P.K.T.; Damià, B. Pilot Study of Pollution Characteristics and Ecological Risk of Disinfection Byproducts in Natural Waters in Hong Kong. Environ. Toxicol. Chem. 2022, 41, 2613–2621. [Google Scholar] [CrossRef] [PubMed]
- Sommer, U.; Adrian, R.; De, L.; Domis, S.; Elser, J.J.; Gaedke, U.; Winder, M. Beyond the Plankton Ecology Group (PEG) Model: Mechanisms Driving Plankton Succession. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- Nabout, J.C.; Siqueira, T.; Bini, L.M.; Nogueira, I.D.S. No evidence for environmental and spatial processes in structuring phytoplankton communities. Acta Oecol. 2009, 35, 720–726. [Google Scholar] [CrossRef]
- Abrantes, N.; Antunes, S.C.; Pereira, M.J.; Gonçalves, F. Seasonal succession of cladocerans and phytoplankton and their interactions in a shallow eutrophic lake (Lake Vela, Portugal). Acta Oecol. 2006, 29, 54–64. [Google Scholar] [CrossRef]
- Yang, W.; Zheng, Z.; Zheng, C.; Lu, K.; Ding, D.; Zhu, J. Temporal variations in a phytoplankton community in a subtropical reservoir: An interplay of extrinsic and intrinsic community effects. Sci. Total Environ. 2018, 612, 720–727. [Google Scholar] [CrossRef]
- Picapedra, P.H.S.; Fernandes, C.; Baumgartner, G.; Sanches, P.V. Zooplankton communities and their relationship with water quality in eight reservoirs from the midwestern and southeastern regions of Brazil. Braz. J. Biol. 2021, 81, 701–713. [Google Scholar] [CrossRef]
- Lazzaro, X.; Bouvy, M.; Ribeiro-Filho, R.A.; Oliviera, V.S.; Sales, L.T.; Vasconcelos, A.R.M.; Mata, M.R. Do fish regulate phytoplankton in shallow eutrophic Northeast Brazilian reservoirs? Freshw. Boil. 2003, 48, 649–668. [Google Scholar] [CrossRef]
- Dai, W.; Zhang, J.; Tu, Q.; Deng, Y.; Qiu, Q.; Xiong, J. Bacterioplankton assembly and interspecies interaction indicating increasing coastal eutrophication. Chemosphere 2017, 177, 317–325. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Gsell, A.S.; Mooij, W.M.; Van Donk, E.; Van Den Wyngaert, S.; De Senerpont Domis, L.N. Chytrid infections and diatom spring blooms: Paradoxical effects of climate warming on fungal epidemics in lakes. Freshw. Biol. 2011, 56, 754–766. [Google Scholar] [CrossRef]
- Liu, L.; Yang, J.; Lv, H.; Yu, Z. Synchronous dynamics and correlations between bacteria and phytoplankton in a subtropical drinking water reservoir. FEMS Microbiol. Ecol. 2014, 90, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Paver, S.F.; Hayek, K.R.; Gano, K.A.; Fagen, J.R.; Brown, C.T.; Davis-Richardson, A.G.; Crabb, D.B.; Rosario-Passapera, R.; Giongo, A.; Triplett, E.W.; et al. Interactions between specific phytoplankton and bacteria affect lake bacterial community succession. Environ. Microbiol. 2013, 15, 2489–2504. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lv, H.; Yang, J.; Liu, L.; Yu, X.; Chen, H. Decline in water level boosts cyanobacteria dominance in subtropical reservoirs. Sci. Total Environ. 2016, 557–558, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Sondergaard, M.; Jeppesen, E.; Mortensen, E.; Dall, E.; Kristensen, P.; Sortkjaer, O. Phytoplankton biomass reduction after planktivorous fish reduction in a shallow, eutrophic lake—A combined effect of reduced internal P-loading and increased zooplankton grazing. Hydrobiologia 1990, 200, 229–240. [Google Scholar] [CrossRef]
- Wiltshire, K.H.; Malzahn, A.M.; Wirtz, K.; Greve, W.; Janisch, S.; Mangelsdorf, P.; Manly, B.F.J.; Boersma, M. Resilience of North Sea Phytoplankton Spring Bloom Dynamics: An Analysis of Long-Term Data at Helgoland Roads. Limnol. Oceanogr. 2008, 53, 1294–1302. [Google Scholar] [CrossRef]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of Microbial Community Resistance and Resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A. Trait-Based Community Ecology of Phytoplankton. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 615–639. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Urban Lakes | Country Lakes | Mann–Whitney U Test | ||||||
|---|---|---|---|---|---|---|---|---|
| Avg | Max | Min | Avg | Max | Min | Z | p | |
| TN (mg L−1) | 1.82 | 5.48 | 0.30 | 0.83 | 1.22 | 0.47 | −1.761 | 0.081 |
| NO3−-N (mg L−1) | 0.54 | 2.50 | 0.09 | 0.35 | 0.46 | 0.20 | 0.935 | 0.369 |
| NH4+-N (mg L−1) | 0.30 | 1.54 | 0.03 | 0.22 | 0.41 | 0.11 | 0.683 | 0.516 |
| TP (mg L−1) | 0.18 | 0.62 | 0.03 | 0.40 | 0.80 | 0.03 | 0.467 | 0.661 |
| PO43−-P (mg L−1) | 0.02 | 0.09 | 0.00 | 0.10 | 0.20 | 0.00 | 1.061 | 0.297 |
| CODMn (mg L−1) | 5.20 | 8.00 | 2.23 | 4.56 | 7.09 | 2.06 | −0.935 | 0.369 |
| Turbidity (NTU) | 23.1 | 186.7 | 1.1 | 82.7 | 164.5 | 11.2 | 1.815 | 0.069 |
| Chl a (μg L−1) | 75.85 | 299.02 | 3.64 | 14.32 | 34.88 | 0.94 | −2.552 | 0.009 |
| WT (°C) | 24.4 | 29.0 | 21.8 | 24.6 | 26.0 | 23.8 | 0.954 | 0.350 |
| DO (mg L−1) | 8.0 | 15.1 | 3.5 | 6.7 | 10.4 | 4.7 | −0.935 | 0.350 |
| SPC (S m−1) | 411.51 | 832.00 | 242.00 | 261.98 | 366.60 | 155.90 | −2.804 | 0.003 |
| pH | 8.51 | 9.64 | 7.76 | 7.04 | 7.40 | 6.74 | −3.882 | 0.000 |
| ORP (mV) | 114.8 | 145.6 | 81.5 | 157.7 | 170.6 | 120.8 | 3.379 | 0.000 |
| TC (mg L−1) | 36.18 | 59.28 | 11.04 | 28.56 | 44.82 | 13.47 | −0.683 | 0.516 |
| IC (mg L−1) | 30.81 | 46.72 | 10.99 | 25.46 | 37.09 | 14.65 | −1.186 | 0.250 |
| TOC (mg L−1) | 5.37 | 15.47 | 0.00 | 3.59 | 9.31 | 0.00 | −0.809 | 0.428 |
| Urban Lakes | Country Lakes | Mann–Whitney U Test | ||||||
|---|---|---|---|---|---|---|---|---|
| Avg | Max | Min | Avg | Max | Min | Z | p | |
| THM (μg L−1) | 1.23 | 4.37 | 0.00 | 0.70 | 1.80 | 0.00 | −1.618 | 0.111 |
| HAA (μg L−1) | 7.86 | 14.55 | 3.07 | 5.92 | 7.00 | 5.26 | −1.186 | 0.250 |
| HKs (μg L−1) | 0.00 | 0.14 | 0.00 | 0.00 | 0.00 | 0.00 | −0.408 | 0.930 |
| Nas (ng L−1) | 0.008 | 0.024 | 0.000 | 0.010 | 0.019 | 0.005 | 1.187 | 0.250 |
| Aromatics (ng L−1) | 0.02 | 0.05 | 0.00 | 0.01 | 0.04 | 0.00 | −1.366 | 0.182 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, G.; Xu, P.; Zhu, Y.; Abate, A.R.; Mi, W.; Bi, Y. Effects of Massive Use of Disinfectants on the Plankton Communities in Lakes from Wuhan. Water 2023, 15, 3875. https://doi.org/10.3390/w15223875
Song G, Xu P, Zhu Y, Abate AR, Mi W, Bi Y. Effects of Massive Use of Disinfectants on the Plankton Communities in Lakes from Wuhan. Water. 2023; 15(22):3875. https://doi.org/10.3390/w15223875
Chicago/Turabian StyleSong, Gaofei, Pingping Xu, Yuxuan Zhu, Adilo Rediat Abate, Wujuan Mi, and Yonghong Bi. 2023. "Effects of Massive Use of Disinfectants on the Plankton Communities in Lakes from Wuhan" Water 15, no. 22: 3875. https://doi.org/10.3390/w15223875
APA StyleSong, G., Xu, P., Zhu, Y., Abate, A. R., Mi, W., & Bi, Y. (2023). Effects of Massive Use of Disinfectants on the Plankton Communities in Lakes from Wuhan. Water, 15(22), 3875. https://doi.org/10.3390/w15223875