
Tracking the Evolution of Microbial Communities on Microplastics through a Wastewater Treatment Process: Insight into the “Plastisphere”

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
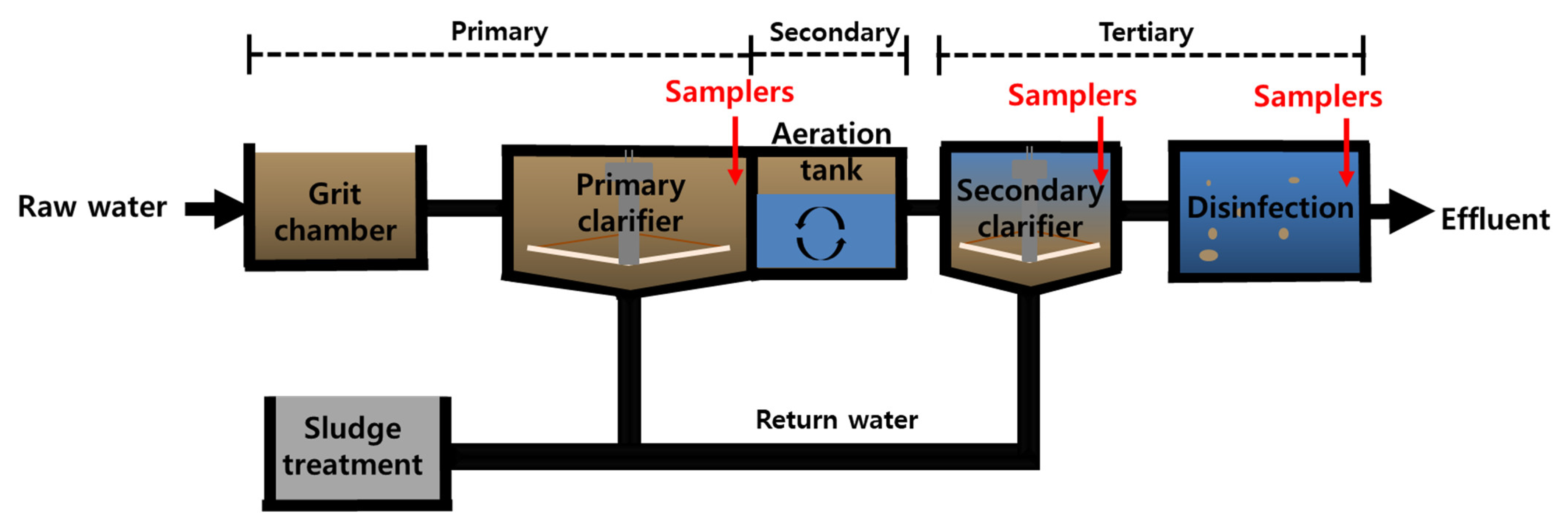
2.1. Description of the WWTP
2.2. Experiment Design: Deployment of MPs in the WWTPs
2.3. Microbial Analyses
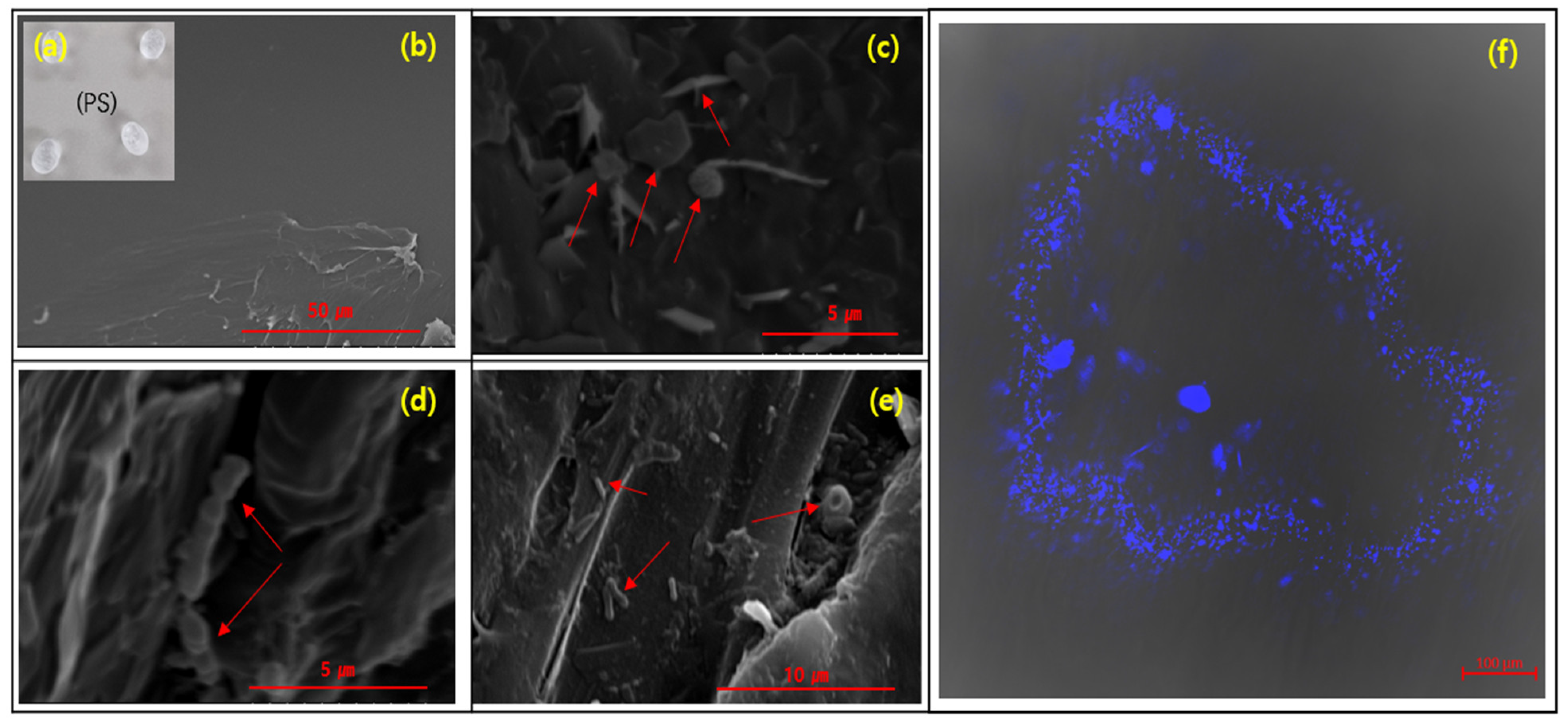
2.3.1. Morphology Analysis
2.3.2. DNA Extraction and High-Throughput Sequencing
2.3.3. Bacterial Community Analysis
3. Results and Discussion
3.1. The Morphology of the Biofilms on the PS
3.2. Bacterial Community Diversity and Clustering
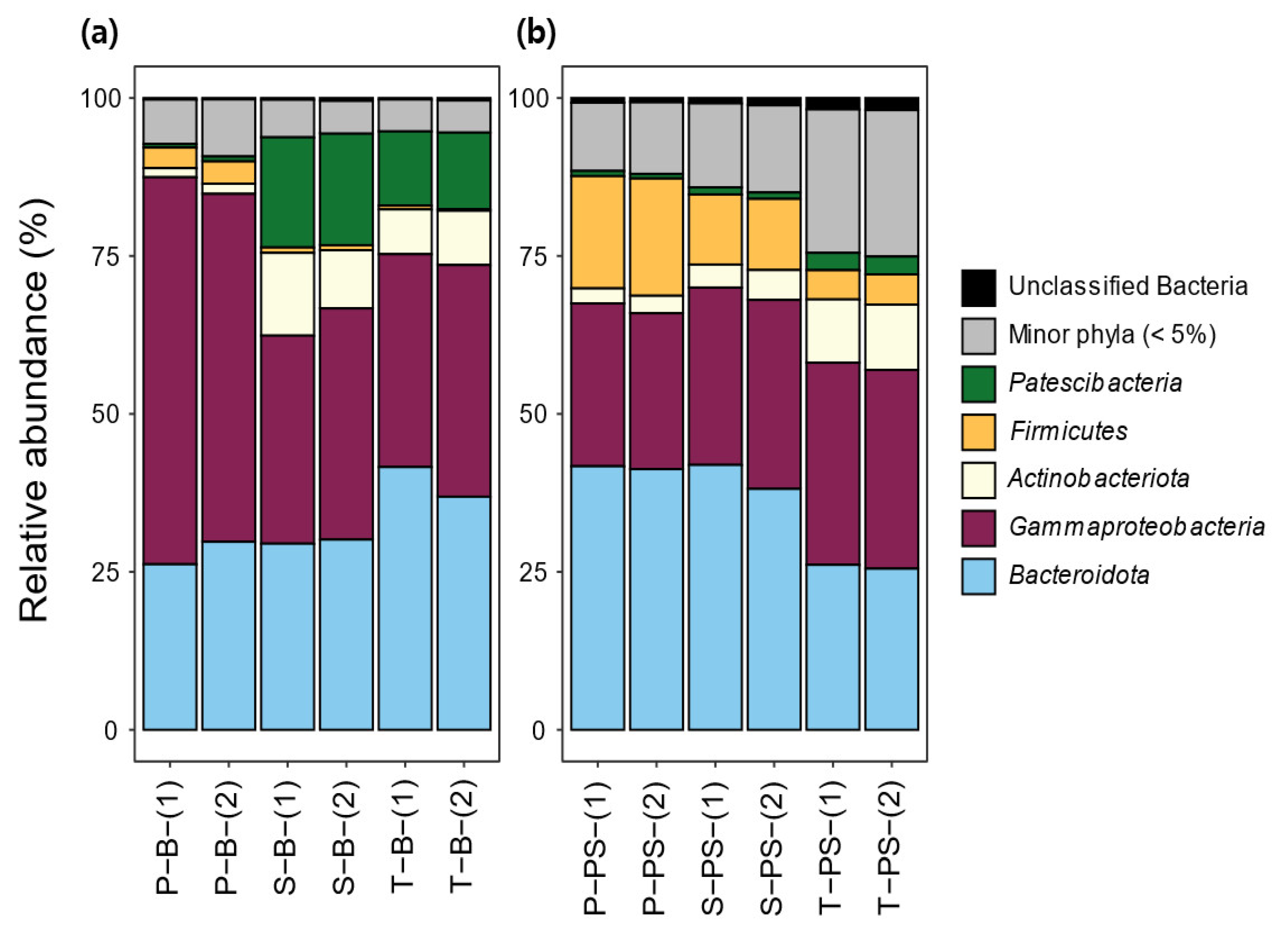
3.3. Bacterial Community Composition and Structure
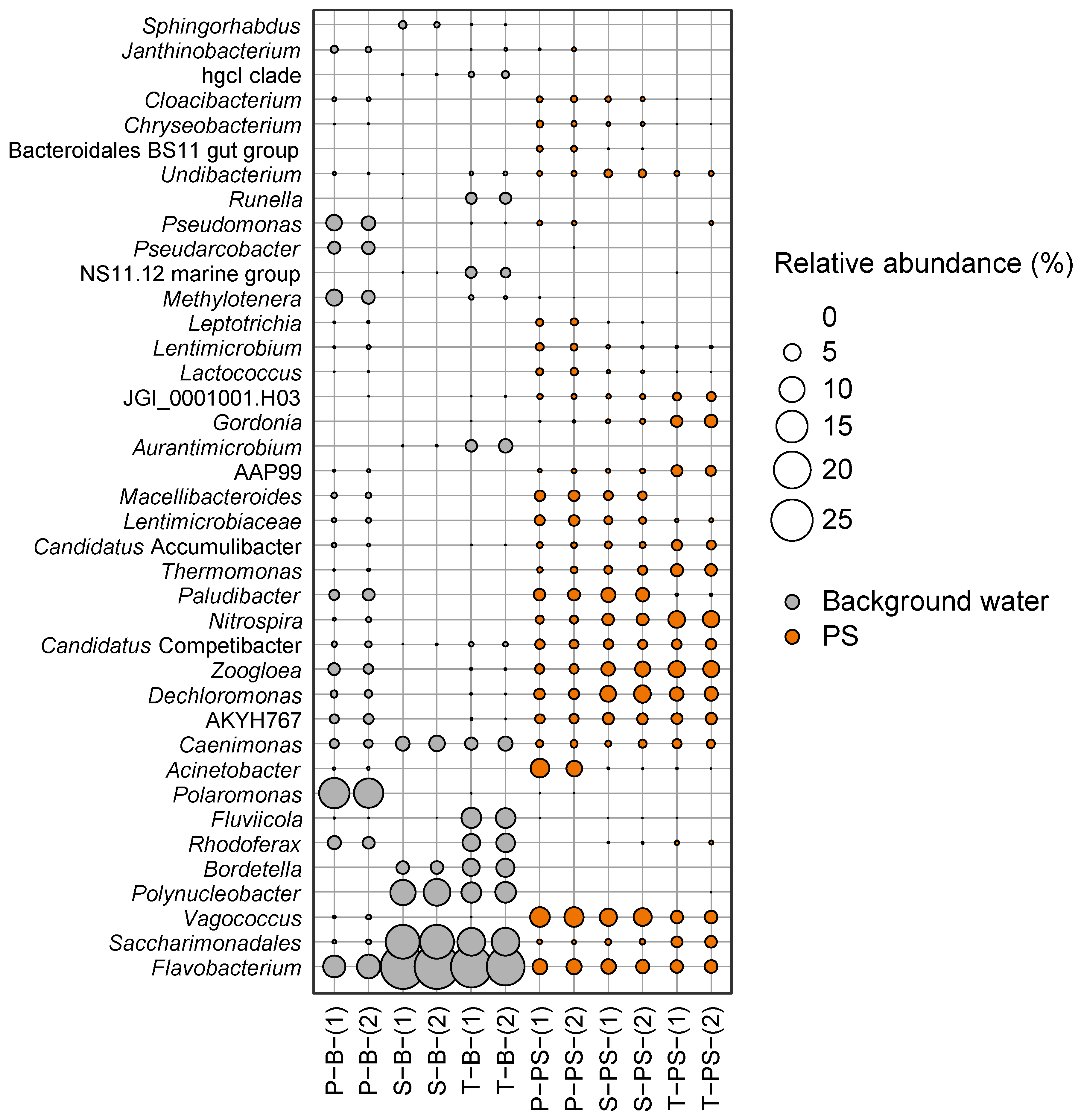
3.4. In-Depth Microbiome at the Genus Level
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sangroniz, A.; Zhu, J.-B.; Tang, X.; Etxeberria, A.; Chen, E.Y.-X.; Sardon, H. Packaging materials with desired mechanical and barrier properties and full chemical recyclability. Nat. Commun. 2019, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Frias, J.P.G.L.; Nash, R. Microplastics: Finding a consensus on the definition. Mar. Pollut. Bull. 2019, 138, 14–147. [Google Scholar] [CrossRef] [PubMed]
- Gago, J.; Galgani, F.; Maes, T.; Thompson, R.C. Microplastics in Seawater: Recommendations from the Marine Strategy Framework Directive Implementation Process. Front. Mar. Sci. 2016, 3, 219. [Google Scholar] [CrossRef]
- Susanti, N.K.Y.; Mardiastuti, A.; Wardiatno, Y. Microplastics and the Impact of Plastic on Wildlife: A Literature Review. IOP Conf. Ser. Earth Environ. Sci. 2020, 528, 012013. [Google Scholar] [CrossRef]
- Xu, S.; Ma, J.; Ji, R.; Pan, K.; Miao, A.-J. Microplastics in aquatic environments: Occurrence, accumulation, and biological effects. Sci. Total Environ. 2020, 703, 134699. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, F.; Li, J.; Chen, Q.; Ma, L.; Zeng, E.; Shi, H. A review of microplastics in table salt, drinking water, and air: Direct human exposure. Environ. Sci. Technol. 2020, 54, 3740–3751. [Google Scholar] [CrossRef]
- Park, S.; Kim, I.; Jeon, W.-H.; Moon, H.S. Exploring the vertical transport of Microplastics in subsurface environments: Lab-scale experiments and field evidence. J. Cont. Hydrol. 2023, 257, 104215. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; Zettler, E.R.; Mincer, T.J. Ecology of the plastisphere. Nat. Rev. Microbiol. 2020, 18, 139–151. [Google Scholar] [CrossRef]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L.A. Life in the “Plastisphere”: Microbial Communities on Plastic Marine Debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef]
- Perveen, S.; Pablos, C.; Reynolds, K.; Stanley, S.; Marugán, J. Growth and prevalence of antibiotic-resistant bacteria in microplastic biofilm from wastewater treatment plant effluents. Sci. Total Environ. 2023, 856, 159024. [Google Scholar] [CrossRef] [PubMed]
- Pham, D.N.; Clark, L.; Li, M. Microplastics as hubs enriching antibiotic-resistant bacteria and pathogens in municipal activated sludge. J. Hazard. Mater. Lett. 2021, 2, 100014. [Google Scholar] [CrossRef]
- Iyare, P.U.; Ouki, S.K.; Bond, T. Microplastics removal in wastewater treatment plants: A critical review. Environ. Sci. Water Res. Technol. 2020, 6, 2664–2675. [Google Scholar] [CrossRef]
- Park, H.J.; Oh, M.J.; Kim, P.G.; Kim, G.; Jeong, D.H.; Ju, B.K.; Lee, W.S.; Chung, H.M.; Kang, H.J.; Kwon, J.H. National reconnaissance survey of microplastics in municipal wastewater treatment plants in Korea. Environ. Sci. Technol. 2020, 54, 1503–1512. [Google Scholar] [CrossRef]
- Sol, D.; Laca, A.; Laca, A.; Díaz, M. Approaching the environmental problem of microplastics: Importance of WWTP treatments. Sci. Total Environ. 2020, 740, 140016. [Google Scholar] [CrossRef]
- Tang, K.H.D.; Hadibarata, T. Microplastics removal through water treatment plants: Its feasibility, efficiency, future prospects and enhancement by proper waste management. Environ. Chall. 2021, 5, 100264. [Google Scholar] [CrossRef]
- Mintenig, S.M.; Int-Veen, I.; Löder, M.G.; Primpke, S.; Gerdts, G. Identification of microplastic in effluents of wastewater treatment plants using focal plane array-based micro-Fourier-transform infrared imaging. Water Res. 2017, 108, 365–372. [Google Scholar] [CrossRef]
- Murphy, F.; Ewins, C.; Carbonnier, F.; Quinn, B. Wastewater Treatment Works (WwTW) as a Source of microplastics in the aquatic environment. Environ. Sci. Technol. 2016, 50, 5800–5808. [Google Scholar] [CrossRef]
- Sun, J.; Dai, X.; Wang, Q.; van Loosdrecht, M.C.; Ni, B.-J. Microplastics in wastewater treatment plants: Detection, occurrence and removal. Water Res. 2019, 152, 21–37. [Google Scholar] [CrossRef]
- Kelly, J.J.; London, M.G.; McCormick, A.R.; Rojas, M.; Scott, J.W.; Hoellein, T.J. Wastewater treatment alters microbial colonization of microplastics. PLoS ONE 2021, 16, e0244443. [Google Scholar] [CrossRef]
- Kruglova, A.; Muñoz-Palazón, B.; Gonzalez-Martinez, A.; Mikola, A.; Vahala, R.; Talvitie, J. The dangerous transporters: A study of microplastic-associated bacteria passing through municipal wastewater treatment. Environ. Pollut. 2022, 314, 120316. [Google Scholar] [CrossRef]
- Boni, W.; Parrish, W.; Patil, S.; Fahrenfeld, N.I. Total coliform and Escherichia coli in microplastic biofilms grown in wastewater and inactivation by peracetic acid. Water Environ. Res. 2021, 93, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.D.; Das, K.R.; Naik, M.M. Co-selection of multi-antibiotic resistance in bacterial pathogens in metal and microplastic contaminated environments: An emerging health threat. Chemosphere 2018, 215, 846–857. [Google Scholar] [CrossRef]
- Shen, M.; Zeng, Z.; Li, L.; Song, B.; Zhou, C.; Zeng, G.; Zhang, Y.; Xiao, R. Microplastics act as an important protective umbrella for bacteria during water/wastewater disinfection. J. Clean. Prod. 2021, 315, 128188. [Google Scholar] [CrossRef]
- Basili, M.; Quero, G.M.; Giovanneilli, D.; Manini, E.; Vignaroli, C.; Avio, C.G.; De Marco, R.; Luna, G.M. Major role of Surrounding environment in Shaping biofilm community composition on Marine Plastic Debris. Front. Mar. Sci. 2020, 7, 262. [Google Scholar] [CrossRef]
- Kang, H.; Park, S.; Lee, B.; Yoon, S.; Oh, H.; Ahn, J.; Kim, S. Characteristics of Microplastics content and estimation of Micorplastics outflow in G city sewage treatment plant. Jour. Wat. Treat. 2020, 28, 43–49. [Google Scholar] [CrossRef]
- Boakyo, Y.D.; Osafo, N.; Amaning Danquah, C.; Adu, F.; Agyare, C. Antimicrobial agents: Antibacterial agents, Anti-biofilm agents, antibacterial natural compounds, and antibacterial chemicals. In Antimicrobial, Antibiotic Resistance, Antibiofilm Strategies and Activity Methods; Kirmusaoğlu, S., Ed.; IntechOpen: London, UK, 2019. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. PNAS 2021, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Kang, B.R.; Kim, J.J.; Hong, J.K.; Schlosser, D.; Lee, T.K. Continuous operation of fungal wheel reactor based on solid-state fermentation for the removal of pharmaceutical and personal care products. J. Environ. Manag. 2023, 331, 117316. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Boylen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. App. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; John Wiley and Sons: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. Vegan: Community Ecology Package. R, Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 November 2017).
- Nguyen, H.T.; Choi, W.; Kim, E.-J.; Cho, K. Microbial community niches on microplastics and prioritized environmental factors under various urban riverine conditions. Sci. Total Environ. 2022, 849, 157781. [Google Scholar] [CrossRef]
- Mughini-Gras, L.; van der Plaats, R.Q.J.; van der Wielen, P.W.J.J.; Bauerlein, P.S.; de Roda Husman, A.M. Riverine microplastic and microbial community compositions: A field study in the Netherlands. Water Res. 2021, 192, 116852. [Google Scholar] [CrossRef]
- Bydalek, F.; Webster, G.; Barden, R.; Weightman, A.J.; Kasprzyk-Hordern, B.; Wenk, J. Microplastic biofilm, associated pathogen and antimicrobial resistance dynamics through a wastewater treatment process incorporating a constructed wetland. Water Res. 2023, 235, 119936. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, L.; Chen, H.; Yu, Z.; Yang, J.R.; Xue, Y.; Huang, B.; Yang, J. Community dynamics of free-living and particle-attached bacteria following a reservoir Microcystis bloom. Sci. Total Environ. 2019, 660, 501–511. [Google Scholar] [CrossRef]
- Savlo, D.; Sinclair, L.; Ijaz, U.Z.; Parajka, J.; Reischer, G.H.; Stadler, P.; Blaschke, A.P.; Blöschl, G.; Mach, R.L.; Kirschner, A.K.T.; et al. Bacterial diversity along a 2600 km river continuum. Environ. Microbiol. 2015, 17, 4994–5007. [Google Scholar]
- Moyal, J.; Dave, P.H.; Wu, M.; Karimpour, S.; Brar, S.K.; Zhong, H.; Kwong, R.W.H. Impacts of biofilm formation on the physicochemical properties and toxicity of microplastics: A concise review. Rev. Environ. Contam. Toxicol. 2023, 261, 8. [Google Scholar] [CrossRef]
- Datta, M.S.; Sliwerska, E.; Gore, J.; Polz, M.F.; Cordero, O.X. Microbial interactions lead to rapid micro-scale successions on model marine particles. Nat. Commun. 2016, 7, 11965. [Google Scholar] [CrossRef]
- Jackson, C.R.; Churchill, P.F.; Roden, E.E. Successional changes in bacterial assemblage structure during epilithic biofilm development. Ecology 2001, 82, 555–566. [Google Scholar] [CrossRef]
- Kumar, C.G.; Anand, S.K. Significance of microbial biofilms in food industry: A review. Int. J. Food Microbiol. 1998, 42, 9–27. [Google Scholar] [CrossRef]
- Toyofuku, M.; Inaba, T.; Kiyokawa, T.; Obana, N.; Yawata, Y.; Nomura, N. Environmental factors that shape biofilm formation. Biosci. Biotechnol. Biochem. 2016, 80, 7–12. [Google Scholar] [CrossRef]
- Zhao, K.; Tseng, B.S.; Beckerman, B.; Jin, F.; Gibiansky, M.L.; Harrison, J.J.; Luijten, E.; Parsek, M.R.; Wong, G.C.L. Psl trails guide exploration and microcolony formation in Pseudomonas aeruginosa biofilms. Nature 2013, 497, 388–391. [Google Scholar] [CrossRef]
- Yawata, Y.; Nomura, N.; Uchiyama, H. Development of a Novel Biofilm Continuous Culture Method for Simultaneous Assessment of Architecture and Gaseous Metabolite Production. Appl. Environ. Microbiol. 2008, 74, 5429–5435. [Google Scholar] [CrossRef]
- Fujii, N.; Kuroda, K.; Narihiro, T.; Aoi, Y.; Ozaki, N.; Ohashi, A.; Kindaichi, T. Metabolic Potential of the Superphylum Patescibacteria Reconstructed from Activated Sludge Samples from a Municipal Wastewater Treatment Plant. Microbes Environ. 2022, 37, ME22012. [Google Scholar] [CrossRef] [PubMed]
- Kindaichi, T.; Yamaoka, S.; Uehara, R.; Ozaki, N.; Ohashi, A.; Albertsen, M.; Nielsen, P.H.; Nielsen, J.L. Phylogenetic diversity and ecophysiology of Candidate phylum Saccharibacteria in activated sludge. FEMS Microbiol. Ecol. 2016, 92, fiw078. [Google Scholar] [CrossRef]
- Singleton, C.M.; Petriglieri, F.; Kristensen, J.M.; Kirkegaard, R.H.; Michaelsen, T.Y.; Andersen, M.H.; Kondrotaite, Z.; Karst, S.M.; Dueholm, M.S.; Nielsen, P.H.; et al. Connecting structure to function with the recovery of over 1000 high-quality metagenome-assembled genomes from activated sludge using long-read sequencing. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Keswani, A.; Oliver, D.M.; Gutierrez, T.; Quilliam, R.S. Microbial hitchhikers on marine plastic debris: Human exposure risks at bathing waters and beach environments. Mar. Environ. Res. 2016, 118, 10–19. [Google Scholar] [CrossRef]
- Lee, J.-W.; Nam, J.-H.; Kim, Y.-H.; Lee, K.-H.; Lee, D.-H. Bacterial communities in the initial stage of marine biofilm formation on artificial surfaces. J. Microbiol. 2008, 46, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Oberbeckmann, S.; Löder, M.G.J.; Labrenz, M. Marine microplastic-associated biofilms—A review. Environ. Chem. 2015, 12, 551–562. [Google Scholar] [CrossRef]
- Verduzco Garibay, M.; Fernández del Castillo, A.; Díaz Torres, O.; de Anda, J.; Yebra-Montes, C.; Senés-Guerrero, C.; Gradilla-Hernández, M.S. Characterization of the Spatial Variation of Microbial Communities in a Decentralized Subtropical Wastewater Treatment Plant Using Passive Methods. Water 2021, 13, 1157. [Google Scholar] [CrossRef]
- Reddy, M.V.; Mohan, S.V. Effect of substrate load and nutrients concentration on the polyhydroxyalkanoates (PHA) production using mixed consortia through wastewater treatment. Bioresour. Technol. 2012, 114, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Joly-Guillou, M.-L. Clinical impact and pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [Google Scholar] [CrossRef]
- Venâncio, I.; Luís, Â.; Domingues, F.; Oleastro, M.; Pereira, L.; Ferreira, S. The Prevalence of Arcobacteraceae in Aquatic Environments: A Systematic Review and Meta-Analysis. Pathogens 2022, 11, 244. [Google Scholar] [CrossRef]
- Galafassi, S.; Sabatino, R.; Sathicq, M.B.; Eckert, E.M.; Fontaneto, D.; Fontana, G.D.; Mossotti, R.; Corno, G.; Volta, P.; Di Cesarem, A. Contribution of microplastic particles to the spread of resistances and pathogenic bacteria in treated wastewaters. Water Res. 2021, 201, 117368. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Rhymes, J.M.; Wade, M.J.; Kevill, J.L.; Malham, S.K.; Grimsley, J.M.; Rimmer, C.; Weightman, A.J.; Farkas, K. Suitability of aircraft wastewater for pathogen detection and public health surveillance. Sci. Total. Environ. 2022, 856, 159162. [Google Scholar] [CrossRef]
- Rummel, C.D.; Jahnke, A.; Gorokhova, E.; Kühnel, D.; Schmitt-Jansen, M. Impacts of Biofilm Formation on the Fate and Potential Effects of Microplastic in the Aquatic Environment. Environ. Sci. Technol. Lett. 2017, 4, 258–267. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, D.; Jin, B.; Su, Y.; Zhang, Y. Deciphering the role of polyethylene microplastics on antibiotic resistance genes and mobile genetic elements fate in sludge thermophilic anaerobic digestion process. Chem. Eng. J. 2023, 452, 139520. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Influent Flow Rate (m3/d) | Effluent Flow Rate (m3/d) |
|---|---|---|
| 21 February | 190,483 | 180,619 |
| 22 February | 198,156 | 189,031 |
| 23 February | 188,177 | 175,208 |
| 24 February | 173,533 | 164,001 |
| 25 February | 183,788 | 188,776 |
| 26 February | 184,733 | 184,461 |
| 27 February | 176,604 | 171,453 |
| 28 February | 185,600 | 174,135 |
| 1 March | 309,517 | 262,622 |
| 2 March | 183,798 | 179,252 |
| 3 March | 184,328 | 176,680 |
| 4 March | 183,367 | 174,389 |
| 5 March | 175,715 | 167,251 |
| 6 March | 183,289 | 177,079 |
| 7 March | 180,822 | 176,478 |
| 8 March | 188,371 | 181,152 |
| 9 March | 186,326 | 182,325 |
| 10 March | 188,041 | 184,768 |
| 11 March | 171,762 | 170,640 |
| 12 March | 182,149 | 171,640 |
| 13 March | 177,429 | 167,192 |
| 14 March | 187,904 | 174,614 |
| Avg. ± sd. | 189,265 ± 27,519 | 180,636 ± 19,500 |
| Sample ID | Description | Shannon’s Index | Richness |
|---|---|---|---|
| P-B | Background water of primary treatment | 4.19 ± 0.20 | 340 ± 103 |
| S-B | Background water of secondary treatment | 3.10 ± 0.20 | 187 ± 17 |
| T-B | Background water of tertiary treatment | 3.55 ± 0.06 | 275 ± 60 |
| P-PS | PS in primary treatment | 5.06 ± 0.005 | 447 ± 13 |
| S-PS | PS in secondary treatment | 4.97 ± 0.03 | 444 ± 29 |
| T-PS | PS in tertiary treatment | 4.97 ± 0.01 | 420 ± 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, J.-K.; Oh, H.; Lee, T.K.; Kim, S.; Oh, D.; Ahn, J.; Park, S. Tracking the Evolution of Microbial Communities on Microplastics through a Wastewater Treatment Process: Insight into the “Plastisphere”. Water 2023, 15, 3746. https://doi.org/10.3390/w15213746
Hong J-K, Oh H, Lee TK, Kim S, Oh D, Ahn J, Park S. Tracking the Evolution of Microbial Communities on Microplastics through a Wastewater Treatment Process: Insight into the “Plastisphere”. Water. 2023; 15(21):3746. https://doi.org/10.3390/w15213746
Chicago/Turabian StyleHong, Jin-Kyung, Hyecheol Oh, Tae Kwon Lee, Seogku Kim, Daemin Oh, Jaehwan Ahn, and Saerom Park. 2023. "Tracking the Evolution of Microbial Communities on Microplastics through a Wastewater Treatment Process: Insight into the “Plastisphere”" Water 15, no. 21: 3746. https://doi.org/10.3390/w15213746
APA StyleHong, J.-K., Oh, H., Lee, T. K., Kim, S., Oh, D., Ahn, J., & Park, S. (2023). Tracking the Evolution of Microbial Communities on Microplastics through a Wastewater Treatment Process: Insight into the “Plastisphere”. Water, 15(21), 3746. https://doi.org/10.3390/w15213746