
Beringian Freshwater Mussel Beringiana beringiana (Unionidae) in Northeast Asia

 , ,
, ,
Abstract
:1. Introduction
- To summarize the available data on the Beringiana distribution in Northeast Asia east of the Verkhoyansk Range and the Okhota River basin;
- To analyze the possible Beringiana dispersion pathways given the high distances between its enclaves and, in particular, the distribution of the species on the two continents.
2. Materials and Methods
3. Results
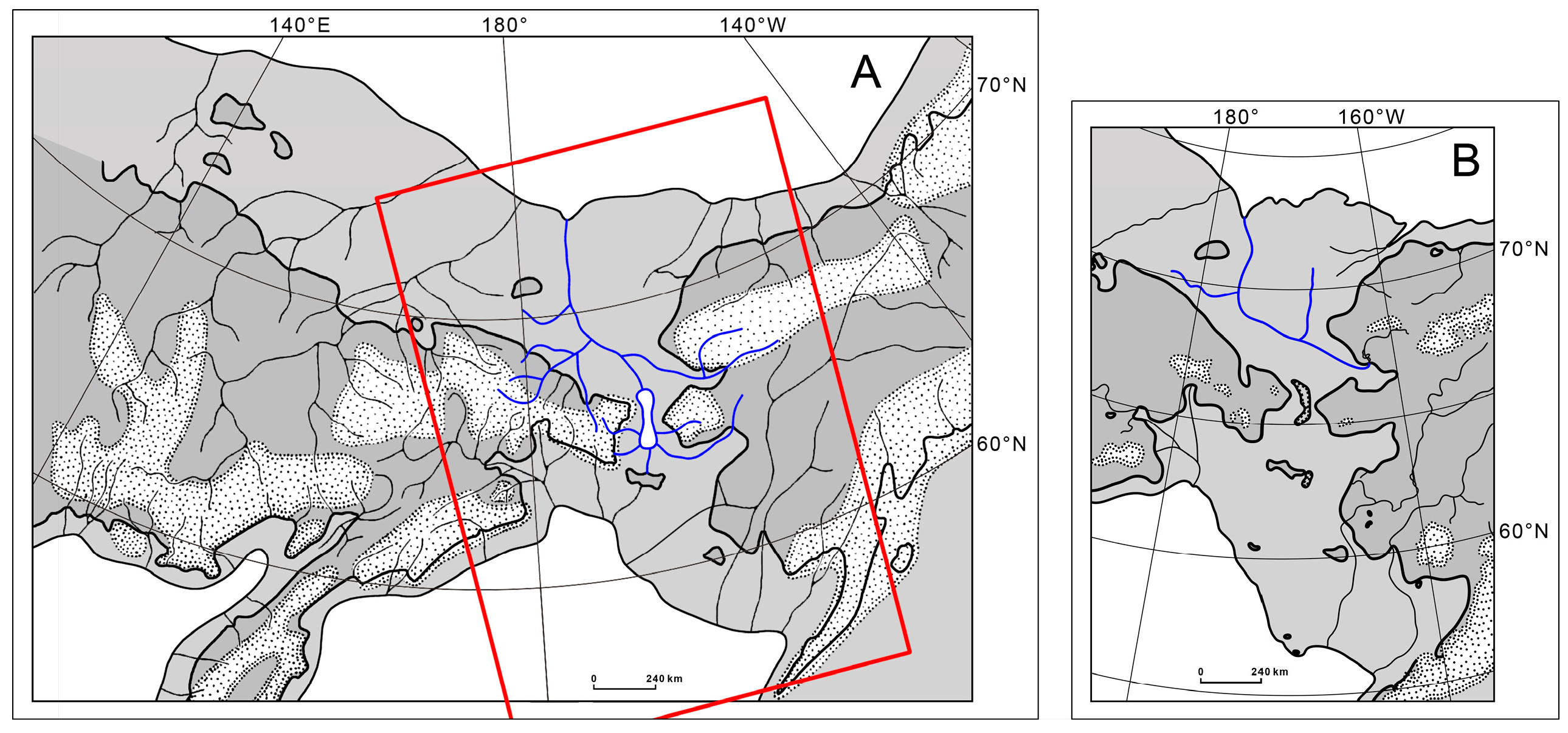
3.1. Distribution
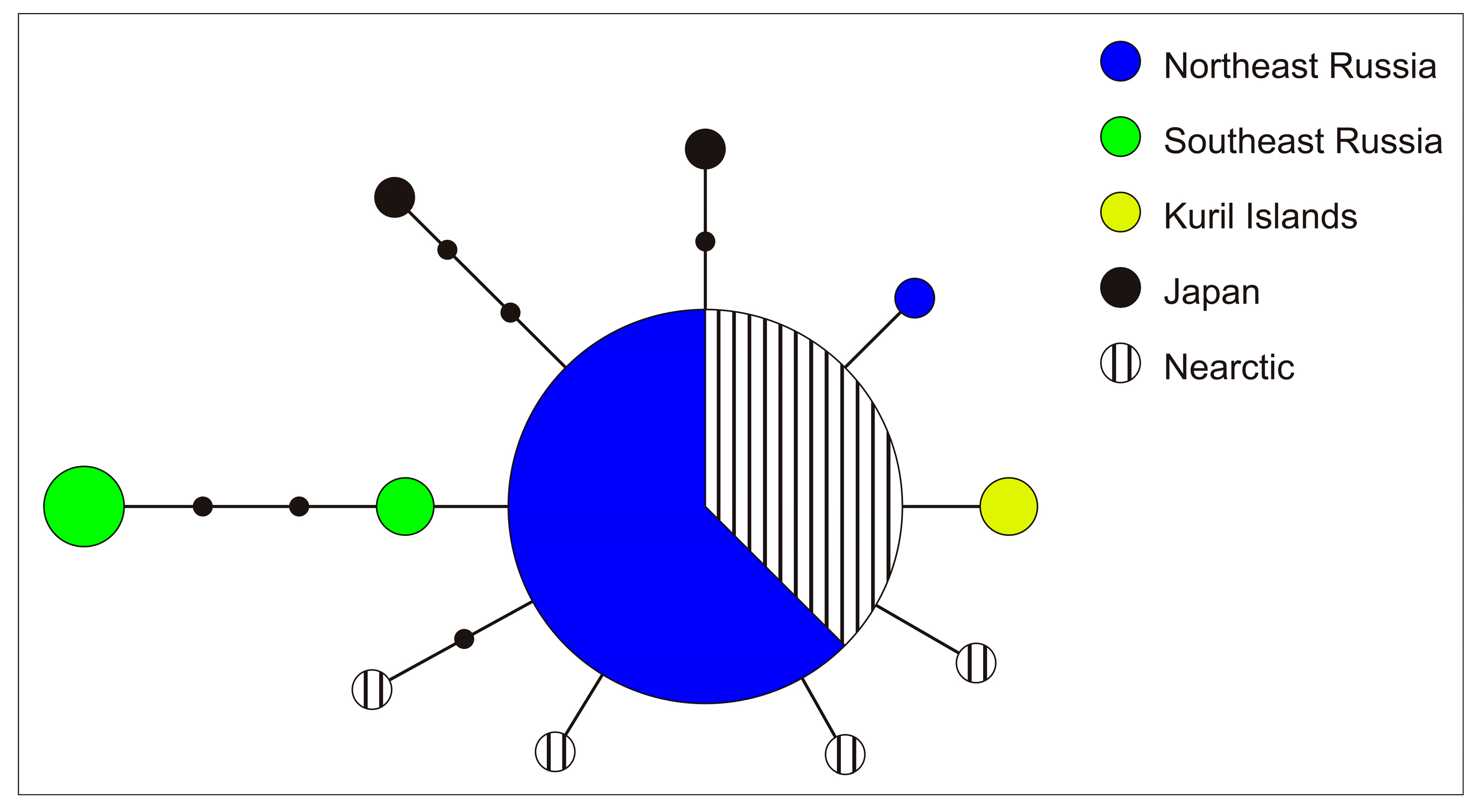
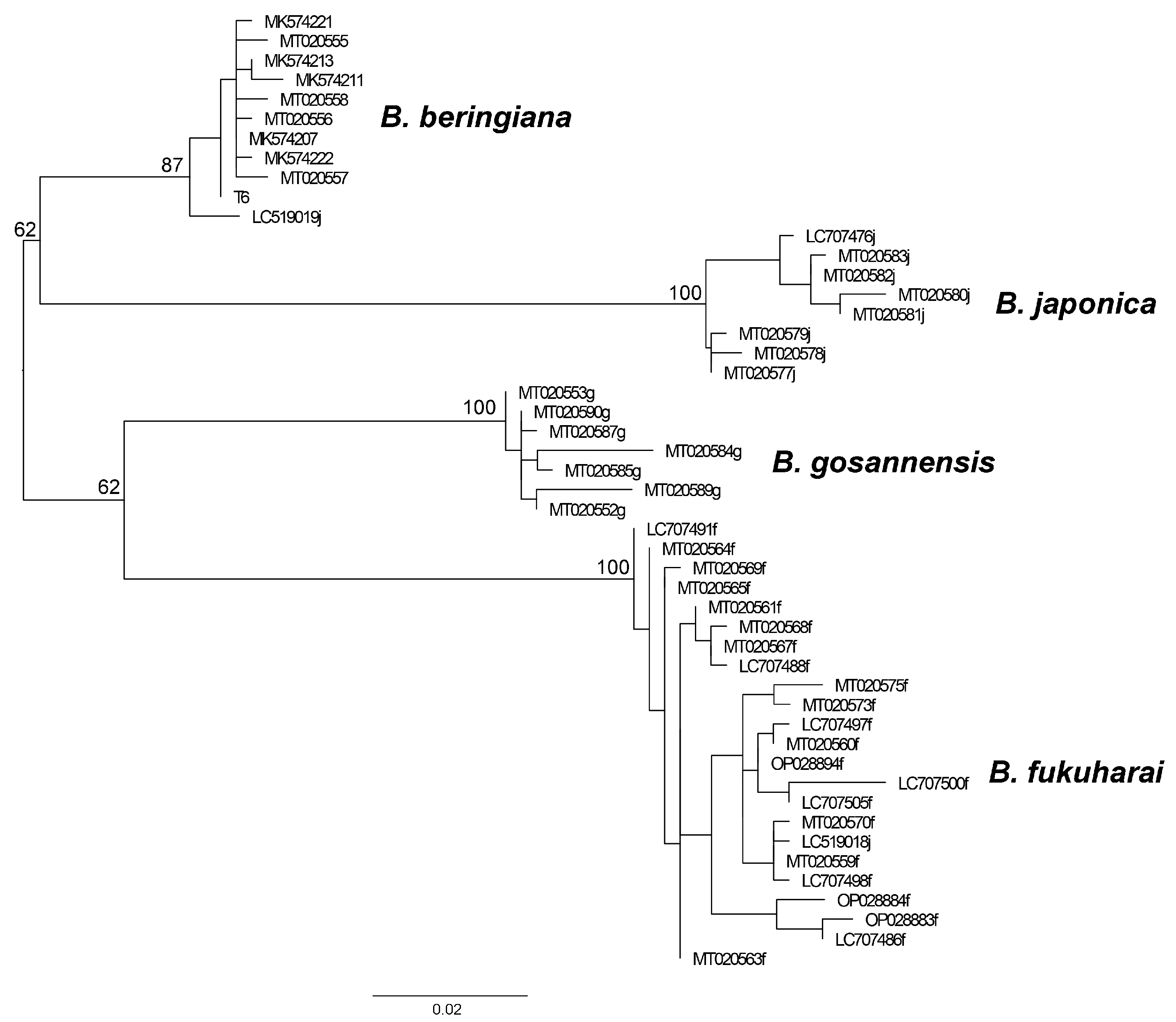
3.2. Genetic Analysis
3.3. Conchological Analysis
- -
- The ratio of shell width (B) to shell height (H) characterizes the degree of prominence (a major trait) of the shell;
- -
- The ratio of the distance between the apex (M) and nymph base (N) to the shell width (B);
- -
- The ratio of the distance between the apex and the anterior margin (Lm) to the shell length (L);
- -
- The ratio of the shell height (H) to the shell length (L) characterizes the general shape of the shell.
4. Discussion
4.1. Beringiana beringiana Distribution in Northeast Asia
4.2. Temperature of B. beringiana Habitats in Northeast Asia
4.3. Genetic Studies
4.4. Morphological Variation
4.5. Freshwater Dispersal Pathways of B. beringiana
4.6. On the Possibility of B. beringiana Larva Dispersal with Fish across Seas
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, M.A.; Lopéz, J.A. Mitochondrial phylogeography of a Beringian relict: The endemic freshwater genus of blackfish Dallia (Esociformes). J. Fish Biol. 2014, 84, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Neretina, A.N.; Gololobova, M.A.; Neplyukhina, A.A.; Zharov, A.A.; Rogers, C.D.; Horne, D.J.; Protopopov, A.V.; Kotov, A.A. Crustacean remains from the Yuka mammoth raise questions about non-analogue freshwater communities in the Beringian region during the Pleistocene. Sci. Rep. 2020, 10, 859. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Aksenova, O.V.; Bespalaya, Y.V.; Kondakov, A.V.; Tomilova, A.A.; Khrebtova, I.S.; Gofarov, M.Y.; Bolotov, I.N. One Beringian genus less: A reassessment of Pacifimyxas Kruglov & Starobogatov, 1985 (Mollusca: Gastropoda: Lymnaeidae) questions the current estimates of Beringian biodiversity. J. Zool. Syst. Evol. Res. 2021, 59, 44–59. [Google Scholar]
- Novichkova, A.A.; Chertoprud, E.S. The species structure, biogeographical status, and the relation to the Beringian fauna of microcrustaceans (Cladocera, Copepoda) of the Magadan Area (Far East, Russia). Arthropoda Sel. 2022, 31, 283–292. [Google Scholar] [CrossRef]
- Zuykova, E.I.; Sleptzova, L.P.; Bochkarev, N.A.; Kuchko, Y.A.; Sheveleva, N.G.; Zakharov, E.S.; Pestryakova, L.A.; Kotov, A.A. Mitochondrial Lineage Diversity and Phylogeography of Daphnia (Daphnia) (Crustacea: Cladocera) in North-East Russia. Water 2022, 14, 1946. [Google Scholar] [CrossRef]
- Markevich, N.G.; Solovyev, M.M.; Vlasenko, P.G.; Izotova, G.V.; Kashinskaya, E.N.; Bochkarev, N.A.; Politov, D.V.; Melnik, N.O.; Esin, E.V. Phylogeny, Distribution, and Biology of Pygmy Whitefish (Prosopium coulterii) in the Beringia Region (Chukotka). Diversity 2023, 15, 547. [Google Scholar] [CrossRef]
- Prozorova, L.A. Freshwater and terrestrial molluscs of the coast of the Tauyskaya Bay. In Biodiversity of the Tauy Bay of the Sea of Okhotsk; Chereshnev, I.A., Ed.; Dalnauka: Vladivostok, Russia, 2005; pp. 252–261. (In Russian) [Google Scholar]
- Prozorova, L.A. Beringiana beringiana. Beringiana youkonensis. Beringiana kamchatica. In Red Data Book of the Magadan Region; Andreev, A.V., Ed.; Okhotnik: Magadan, Russia, 2019; pp. 24–26. (In Russian) [Google Scholar]
- Bogatov, V.V.; Starobogatov, J.I. Anodontinae (Bivalvia) of the genus Beringiana. Zool. Zhurnal 2001, 80, 26–31. (In Russian) [Google Scholar]
- Starobogatov, Y.I.; Prozorova, L.A.; Bogatov, V.V.; Sayenko, E.M. Mollusks. In Key to Freshwater Invertebrates of Russia and Adjacent Lands; Tsalolikhin, S.Y., Ed.; Nauka: Saint-Petersburg, Russia, 2004; pp. 11–252. (In Russian) [Google Scholar]
- Chernyshev, A.V. Systematics of genera Naiads (Bivalvia, Unionida) of the Russian Far East. Bull. Russ. Far East Malacol. Soc. 2004, 8, 5–16. (In Russian) [Google Scholar]
- Vinarski, M.V.; Kantor, Y.I. Analytical Catalogue of Fresh and Brackish Water Molluscs of Russia and Adjacent Countries; A.N. Severtsov Institute of Ecology and Evolution: Moscow, Russia, 2016; pp. 1–545. [Google Scholar]
- Bolotov, I.N.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V.; Aksenova, O.V.; Aksenov, A.S.; Bespalaya, Y.V.; Borovskoy, A.V.; Danilov, P.P.; Dvoryankin, G.A.; et al. Integrative taxonomy, biogeography and conservation of freshwater mussels (Unionidae). Sci. Rep. 2020, 10, 3072. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Hattori, A.; Kondo, T.; Lee, J.H.; Kim, S.K.; Shirai, A.; Hayashi, H.; Usui, T.; Sakuma, K.; Toriya, T.; et al. Freshwater mussels (Bivalvia: Unionidae) from the Rising Sun (Far East Asia): Phylogeny, systematics and distribution. Mol. Phylogen. Evol. 2020, 146, 106755. [Google Scholar] [CrossRef]
- Sano, I.; Saito, T.; Ito, S.; Ye, B.; Uechi, T.; Seo, T.; Do, V.T.; Kimura, K.; Hirano, T.; Yamazaki, D.; et al. Resolving species-level diversity of Beringiana and Sinanodonta mussels (Bivalvia: Unionidae) in the Japanese archipelago using genome-wide data. Mol. Phyl. Evol. 2022, 175, 107563. [Google Scholar] [CrossRef] [PubMed]
- Bogatov, V.V. Large Bivalve Molluscs of Russia’s Fresh Waters; Dalnauka: Vladivostok, Russia, 2022; pp. 1–288. (In Russian) [Google Scholar]
- Graf, D.L. Palearctic freshwater mussel (Mollusca: Bivalvia: Unionoidae) diversity and the Comparatory Method as a species concept. Proc. Acad. Nat. Sci. Phila. 2007, 156, 71–88. [Google Scholar] [CrossRef]
- Bogatov, V.V.; Neretina, T.V.; Anisimova, A.S.; Abdrakhmanov, A. Evaluation of the Applicability of the Comparatory Method for Species Diagnosis of Unionidae (Bivalvia) by Genetic Analysis. Dokl. Biol. Sci. 2018, 482, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Zatravkin, M.N.; Bogatov, V.V. Large Bivalve Mollusks of the Fresh and Brackish Waters of the Far East of the USSR; Publishing House of the FEB AS USSR: Vladivostok, Russia, 1987; pp. 1–152. (In Russian) [Google Scholar]
- Sayenko, E.M.; Shedko, M.B.; Kholin, S.K. Morphology and some peculiarities of the biology of glochidia of the mollusk genus Beringiana (Bivalvia, Unionidae) of Kamchatka and the Northern Kurils. Vestn. Zool. 2001, 35, 59–68. (In Russian) [Google Scholar]
- Sayenko, E.M. Morphological peculiarities of the glochidia (Bivalvia, Anodontinae) in the Russian Far East island populations. In Proceedings of the Vladimir Ya. Levanidov’s Biennial Memorial Meetings, Vladivostok, Russia, 19–21 March 2003; Volume 2, pp. 166–171. (In Russian). [Google Scholar]
- Prozorova, L.A. Mollusks. In Red Data Book of the Chukotka Autonomous District. Volume 1. Animals; Kondratiev, A.V., Litovka, D.I., Eds.; Teksotel LLC: N. Novgorod, Russia, 2022; pp. 12–21. (In Russian) [Google Scholar]
- Klishko, O. Shell Shape Variability in Mollusks (Bivalvia, Margaritiferidae, Unionoidae) and Their Real Species Diversity. Int. Indep. Sci. J. 2020, 2020, 3–15. [Google Scholar]
- Prozorova, L.A. Beringiana chereshnevi. In Red Book of the Chukchi Autonomous District, V. 1; Chereshnev, I.A., Ed.; Wild North: Magadan, Russia, 2008; pp. 21–31. (In Russian) [Google Scholar]
- Zotin, A.A.; Popov, I.Y. Individual growth of Anodonta beringiana (Unionidae, Bivalvia) in postlarval ontogenesis. Russ. J. Dev. Biol. 2019, 50, 189–193. [Google Scholar] [CrossRef]
- Sayenko, E.M. New data on glochidia of anodontins Beringiana and Kunashiria (Unionidae, Bivalvia). In Proceedings of the Vladimir Ya. Levanidov’s Biennial Memorial Meetings, Vladivostok, Russia, 22–24 March 2021; Volume 9, pp. 151–158. (In Russian). [Google Scholar]
- Shekhovtsov, S.V.; Derzhinsky, Y.A.; Poluboyarova, T.V.; Golovanova, E.V.; Peltek, S.E. Phylogeography and genetic lineages of Aporrectodea rosea (Lumbricidae, Annelida). Eur. J. Soil. Biol. 2020, 99, 103191. [Google Scholar] [CrossRef]
- Folmer, O.; Hoeh, W.R.; Black, M.B.; Vrijenhoek, R.C. Conserved primers for PCR amplification of mitochondrial DNA from different invertebrate phyla. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 117693430500100003. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Rosen, O.V. Terrestrial and freshwater mollusks collected by the Kamchatka expedition of F. P. Ryabushinsky in 1908–1909. Yearly Zool. Mus. Acad. Sci. USSR 1927, 27, 261–274. (In Russian) [Google Scholar]
- Vvedenskaya, T.L.; Ulatov, A.V.; Bonk, T.V. Ecological state of Lake Kultuchnoye (east coast, Kamchatka). In Preservation of the Biodiversity of Kamchatka and Adjacent Seas; Bugaev, V.F., Ed.; Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2013; pp. 72–91. (In Russian) [Google Scholar]
- Gorovaya, O.Y.; Butorina, T.E. Parasitofauna of lake-river Dolly (Salvelinus malma) from Lake Dalnee in Kamchatka. Sci. Work. Dalrybvtuz 2007, 19, 174–181. (In Russian) [Google Scholar]
- Vvedenskaya, T.; Ulatov, A.; Koval, O. Some hydrobiological information about Khalaktyrskoye Lake (Eastern Kamchatka). In Preservation of the Biodiversity of Kamchatka and Adjacent Seas; Bugaev, V.F., Ed.; Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2017; pp. 54–58. (In Russian) [Google Scholar]
- Middendorff, A.T. Voyages in the Extreme North and East of Siberia during the Years 1843 and 1844. V. 2; Imperial Academy of Sciences: Saint-Petersburg, Russia, 1851; pp. 163–465. [Google Scholar]
- Andreev, A.V. Wetlands of Russia. Volume 4. Wetlands of the North-East of Russia; Wetland International: Moscow, Russia, 2001; pp. 1–296. (In Russian) [Google Scholar]
- Khamenkova, E.V. Trophical relations of fishes of Kisi lake of Ola river basin (Magadan region). In Proceedings of the Vladimir Ya. Levanidov’s Biennial Memorial Meetings, Vladivostok, Russia, 21–23 March 2011; Volume 5, pp. 563–570. (In Russian). [Google Scholar]
- Ioganson, V.E.; Kuznetsov, A.S.; Deev, G.N.; Boytsov, Y.A.; Tereshchenko, K.I.; Zhukova, V.N.; Chernyshova, M.R. Rivers: Power sources and runoff regime. In North of the Far East; Shilo, N.A., Ed.; Nauka: Moscow, Russia, 1970; pp. 186–203. (In Russian) [Google Scholar]
- Magritsky, D.V. Peculiarities of changes in the heat runoff of the rivers of the North-east of the Asian part of Russia and the assessment of its value. In Dynamics and Interaction of the Earth’s Geospheres. Part 2; Vershinin, D.A., Ed.; Publishing House TSNTI: Tomsk, Russia, 2021; pp. 57–60. (In Russian) [Google Scholar]
- Derzhavin, A.N. Winter trip to the Kuril lake. In Kamchatka Expedition of Fyodor Pavlovich Ryabushinsky, Equipped with the Assistance of the Russian Geographical Society. Issue 1; Schmidt, P.Y., Ed.; Ryabushinsky: Moscow, Russia, 1916; pp. 246–278. [Google Scholar]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000; pp. 1–464. [Google Scholar]
- Sergeeva, I.S.; Bolotov, I.N.; Bespalaya, Y.V.; Makhrov, A.A.; Bukhanova, A.L.; Artamonova, V.S. Freshwater pearl mussels of the genus Margaritifera (Mollusca: Bivalvia) described as M. elongata (Lamarck, 1819) and M. borealis (Westerlund, 1871) should be classified with M. margaritifera (Linnaeus, 1758). Biol. Bull. 2008, 35, 102–105. [Google Scholar] [CrossRef]
- Klishko, O.K. Pearl mussels of the genus Dahurinaia (Bivalvia, Margaritiferidae): Differently sized groups of Margaritifera dahurica Middendorff, 1850. Biol. Bull. 2014, 41, 434–443. [Google Scholar] [CrossRef]
- Makhrov, A.A.; Artamonova, V.S. Instability Stabilized: Mechanisms of Evolutionary Stasis and Genetic Diversity Accumulation in Fishes and Lampreys from Environments with Unstable Abiotic Factors. Contemp. Probl. Ecol. 2020, 13, 370–381. [Google Scholar] [CrossRef]
- Zieritz, A.; Hoffman, J.I.; Amos, W.; Aldridge, D.C. Phenotypic plasticity and genetic isolation-by-distance in the freshwater mussel Unio pictorum (Mollusca: Unionoida). Evol. Ecol. 2010, 24, 923–938. [Google Scholar] [CrossRef]
- Inoue, K.; Hayes, D.M.; Harris, J.L.; Christian, A.D. Phylogenetic and morphometric analyses reveal ecophenotypic plasticity in freshwater mussels Obovaria jacksoniana and Villosa arkansasensis (Bivalvia: Unionidae). Ecol. Evol. 2013, 3, 2670–2683. [Google Scholar] [CrossRef]
- Zając, K.; Zając, T.; Ćmiel, A. What can we infer from the shell dimensions of the thick-shelled river mussel Unio crassus? Hydrobiologia 2018, 810, 415–431. [Google Scholar] [CrossRef]
- Wu, R.; Liu, X.; Guo, L.; Zhou, C.; Ouyang, S.; Wu, X. DNA barcoding, multilocus phylogeny, and morphometry reveal phenotypic plasticity in the Chinese freshwater mussel Lamprotula caveata (Bivalvia: Unionidae). Ecol. Evol. 2022, 12, e9035. [Google Scholar] [CrossRef]
- Chereshnev, I.A. Biological Diversity of Freshwater Ichthyofauna of the North-East of Russia; Dalnauka: Vladivostok, Russia, 1996; pp. 1–198. (In Russian) [Google Scholar]
- Ager, T.A. Late Quaternary vegetation and climate history of the central Bering land bridge from St. Michael Island, western Alaska. Quarter. Res. 2003, 60, 19–32. [Google Scholar] [CrossRef]
- Baranova, Y.P.; Biske, S.F. North-East of the USSR; Nauka: Moscow, Russia, 1964; pp. 1–304. (In Russian) [Google Scholar]
- Hopkins, D.M. Quaternary marine transgression in Alaska. In The Bering Land Bridge; Hopkins, D.M., Ed.; Stanford University Press: Stanford, CA, USA, 1967; pp. 47–90. [Google Scholar]
- Pugachev, O.N. Parasites of Freshwater Fishes of North-East Asia; Publishing House of the Zoological Institute: Leningrad, Russia, 1984; pp. 1–154. (In Russian) [Google Scholar]
- Nakhodkin, N.A.; Nakhodkina, F.N. A river that flows into two oceans. Nauka Tekhnika Yakutii 2020, 1, 82–87. (In Russian) [Google Scholar]
- Chereshnev, I.A. Biogeography of Freshwater Fish Fauna in the Russian Far East; Dalnauka: Vladivostok, Russia, 1998; pp. 1–131. (In Russian) [Google Scholar]
- Koreysha, M.M. Modern Glaciation of the Suntar-Khayat Ridge; USSR Academy of Sciences Press: Moscow, Russia, 1963; pp. 1–170. (In Russian) [Google Scholar]
- Butorina, T.E. Ecological analysis of the parasitic fauna of the charrs (Salvelinus) of the Kamchatka River. In Population Biology and Taxonomy of Salmonids; Konovalov, S.M., Ed.; Institute of Marine Biology: Vladivostok, Russia, 1980; pp. 65–81. (In Russian) [Google Scholar]
- Treasurer, G.W.; Turnbull, T. The pathology and seawater performance of farmed Atlantic salmon infected with glochidia of Margaritifera margaritifera. J. Fish Biol. 2000, 57, 858–866. [Google Scholar] [CrossRef]
- Vikhrev, I.V.; Ieshko, E.P.; Kondakov, A.V.; Mugue, N.S.; Bovykina, G.V.; Efremov, D.A.; Bulakhov, A.G.; Tomilova, A.A.; Yunitsyna, O.A.; Bolotov, I.N. Postglacial expansion routes and mitochondrial genetic diversification of the freshwater pearl mussel in Europe and North America. Diversity 2022, 14, 477. [Google Scholar] [CrossRef]
- De Cicco, A.L. Long-distance movements Anadromous Dolly Varden between Alaska and the USSR. Arctic 1992, 45, 120–123. [Google Scholar]
- Lindberg, G.U. Large Fluctuations in Ocean Level during the Quaternary Period; Nauka: Leningrad, Russia, 1972; pp. 1–548. (In Russian) [Google Scholar]
- Esin, E.V.; Bocharova, E.S.; Mugue, N.S.; Markevich, G.N. Occurrence of sympatric charr groups, Salvelinus, Salmonidae, in the lakes of Kamchatka: A legacy of the last glaciations. J. Fish Biol. 2017, 91, 628–644. [Google Scholar] [CrossRef]
- Osinov, A.G.; Volkov, A.A.; Alekseyev, S.S.; Sergeev, A.A.; Oficerov, M.V.; Kirillov, A.F. On the origin and phylogenetic position of Arctic charr (Salvelinus alpinus complex, Salmonidae) from Lake Cherechen’ (middle Kolyma River basin): Controversial genetic data. Polar Biol. 2017, 40, 777–786. [Google Scholar] [CrossRef]
- Esin, E.V.; Markevich, G.N. Evolution of the Charrs, Genus Salvelinus (Salmonidae). 1. Origins and Expansion of the Species. J. Ichthyol. 2018, 58, 187–203. [Google Scholar] [CrossRef]
- Oleinik, A.G.; Kukhlevsky, A.D.; Skurikhina, L.A. The Relationships of the Charrs Salvelinus sp. 4 (Salmoniformes: Salmonidae) from Lake Nachikinskoe, Kamchatka, as Inferred from Complete Mitochondrial Genome Analysis. Russ. J. Mar. Biol. 2022, 48, 55–59. [Google Scholar] [CrossRef]
- Oleinik, A.G.; Skurikhina, L.A.; Kukhlevsky, A.D.; Bondar, E.I. On the origin of endemic stone charr in the Kamchatka River basin. Hydrobiologia 2019, 840, 21–33. [Google Scholar] [CrossRef]
- Osinov, A.G.; Senchukova, A.L.; Mugue, N.S.; Pavlov, S.D.; Chereshnev, I.A. Speciation and genetic divergence of three species of charr from ancient Lake El’gygytgyn (Chukotka) and their phylogenetic relationships with other representatives of the genus Salvelinus. Biol. J. Linn. Soc. 2015, 116, 63–85. [Google Scholar] [CrossRef]
- Esin, E.V.; Markevich, G.N.; Zlenko, D.V.; Shkil, F.N. Thyroid-Mediated Metabolic Differences Underlie Ecological Specialization of Extremophile Salmonids in the Arctic Lake El’gygytgyn. Front. Ecol. Evol. 2021, 9, 715110. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Trait | |||
|---|---|---|---|---|
| (M − N)/B | Lm/L | B/H | H/L | |
| Kukhtuy, n = 12 | 0.83 ± 0.03 13.6 | 0.18 ± 0.01 10.1 | 0.71 ± 0.02 9.4 | 0.54 ± 0.01 4.7 |
| Chukcha, n = 31 | 0.82 ± 0.02 14.5 | 0.22 ± 0.01 13.9 | 0.63 ± 0.01 13.2 | 0.60 ± 0.01 5.5 |
| Ola, n = 18 | 0.96 ± 0.05 22.8 | 0.19 ± 0.01 15.1 | 0.57 ± 0.02 14.6 | 0.61 ± 0.01 4.5 |
| Anadyr, n = 10 | 0.96 ± 0.04 12.8 | 0.20 ± 0.01 15.0 | 0.61 ± 0.01 6.7 | 0.57 ± 0.01 6.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulakhova, N.A.; Makhrov, A.A.; Lazutkin, A.N.; Shekhovtsov, S.V.; Poluboyarova, T.V.; Berman, D.I. Beringian Freshwater Mussel Beringiana beringiana (Unionidae) in Northeast Asia. Water 2023, 15, 3538. https://doi.org/10.3390/w15203538
Bulakhova NA, Makhrov AA, Lazutkin AN, Shekhovtsov SV, Poluboyarova TV, Berman DI. Beringian Freshwater Mussel Beringiana beringiana (Unionidae) in Northeast Asia. Water. 2023; 15(20):3538. https://doi.org/10.3390/w15203538
Chicago/Turabian StyleBulakhova, Nina A., Alexander A. Makhrov, Anatoly N. Lazutkin, Sergey V. Shekhovtsov, Tatiana V. Poluboyarova, and Daniil I. Berman. 2023. "Beringian Freshwater Mussel Beringiana beringiana (Unionidae) in Northeast Asia" Water 15, no. 20: 3538. https://doi.org/10.3390/w15203538
APA StyleBulakhova, N. A., Makhrov, A. A., Lazutkin, A. N., Shekhovtsov, S. V., Poluboyarova, T. V., & Berman, D. I. (2023). Beringian Freshwater Mussel Beringiana beringiana (Unionidae) in Northeast Asia. Water, 15(20), 3538. https://doi.org/10.3390/w15203538