
Effect of the Pulsed Addition of Phosphorous on the Growth and Toxin Production of the Bloom-Forming Cyanobacterium Chrysosporum ovalisporum (Forti) Zapomelová et al.
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Experimental Design
2.3. Growth Measurement
2.4. Photosynthetic Activity Parameters
2.5. Intracellular Phosphate
2.6. Exopolysaccharides
2.7. Alkaline Phosphatase
2.8. Cylindrospermopsin (CYN)
2.9. Statistical Analysis
3. Results
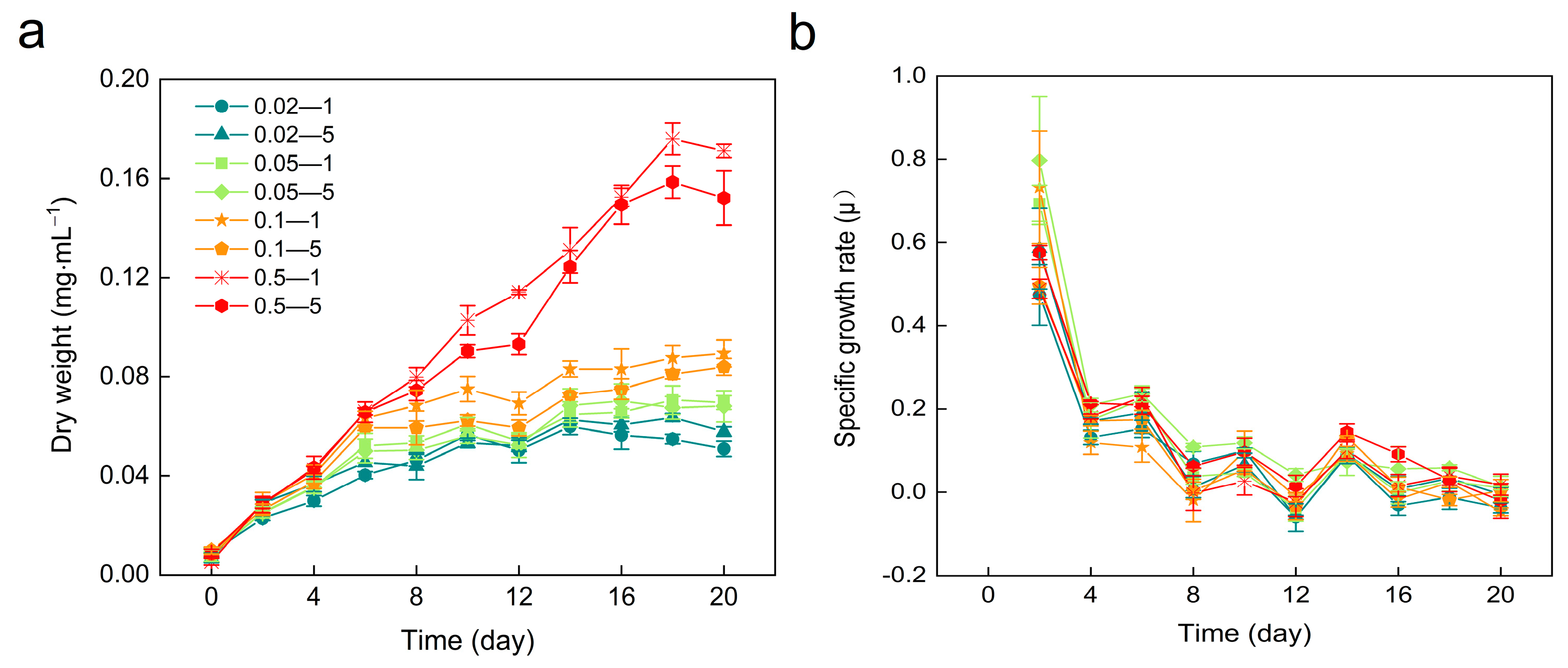
3.1. Biomass and Specific Growth Rate
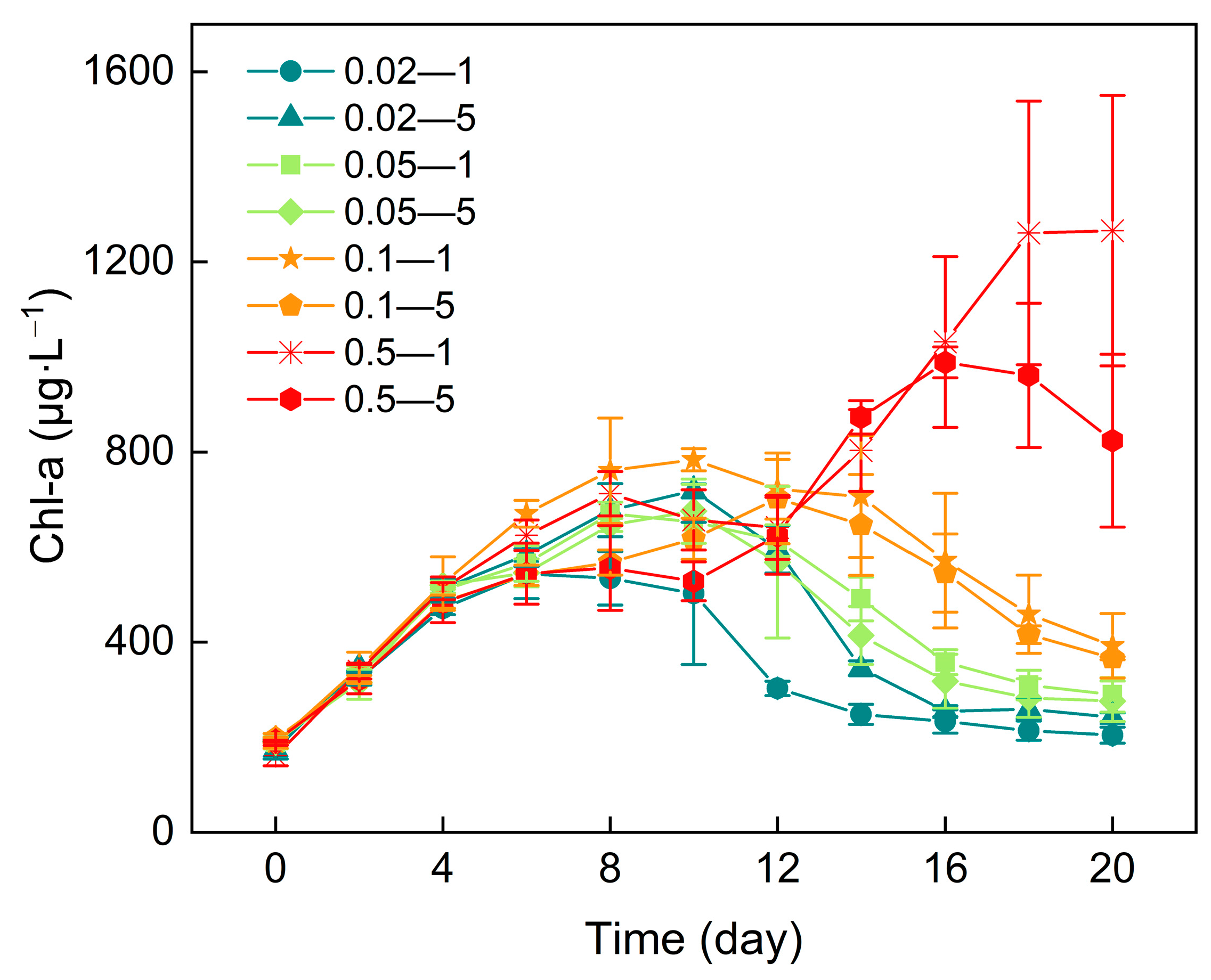
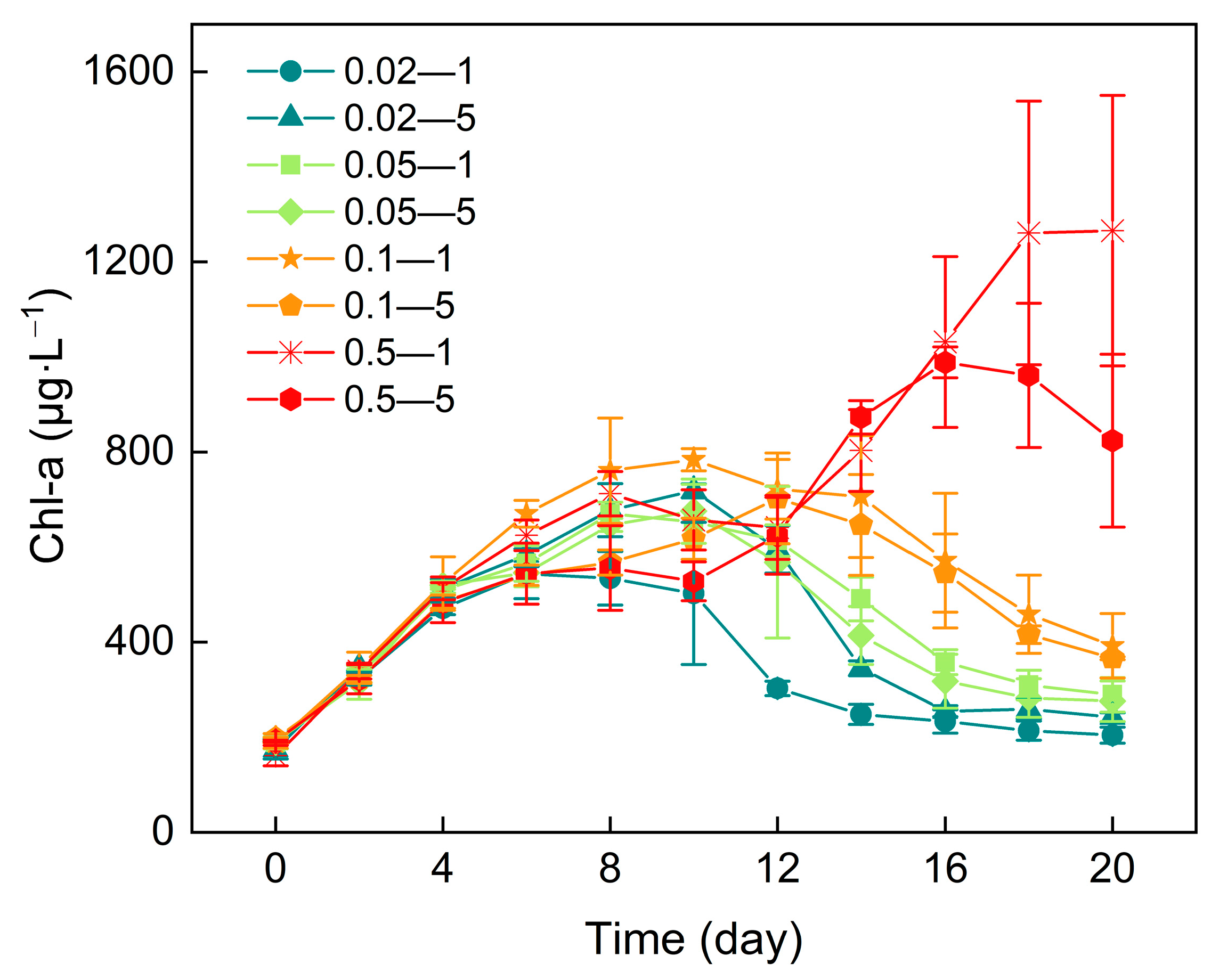
3.2. Chlorophyll-a
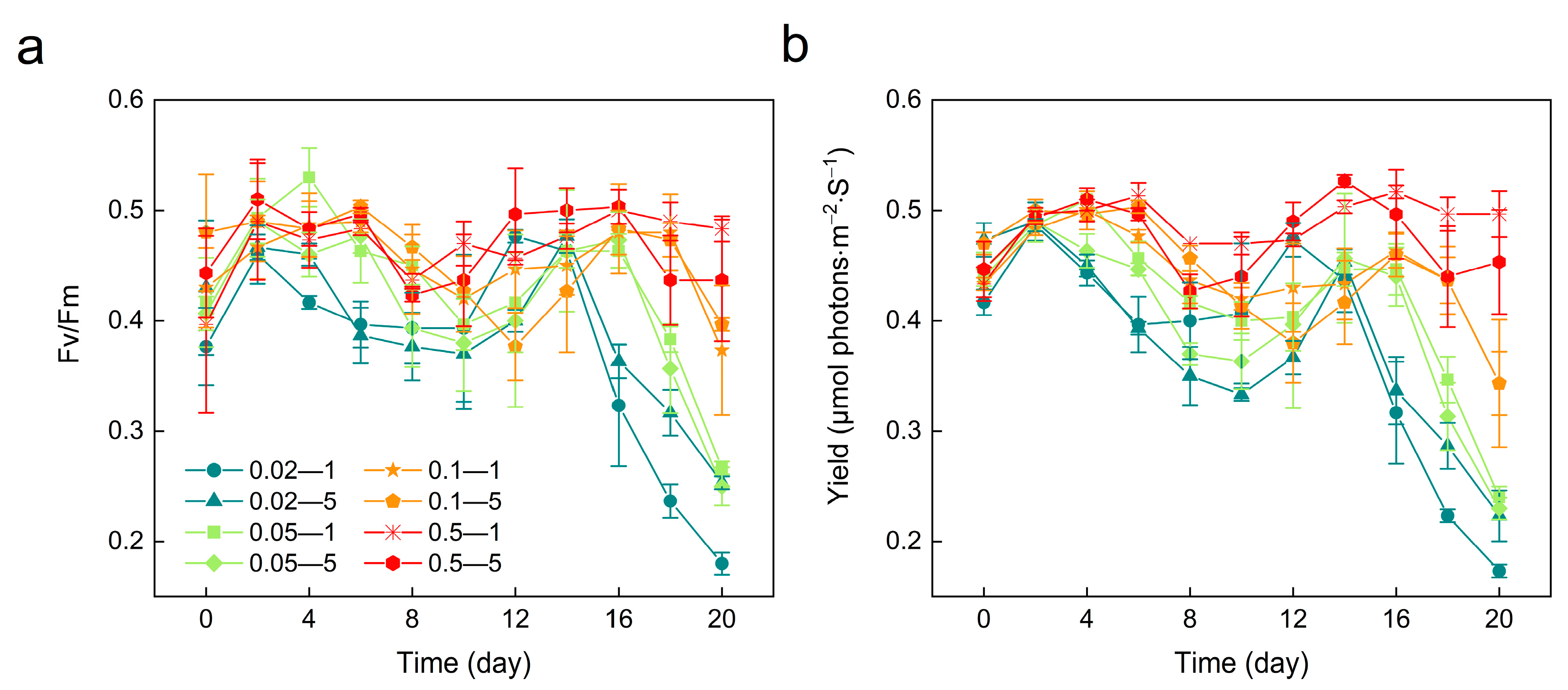
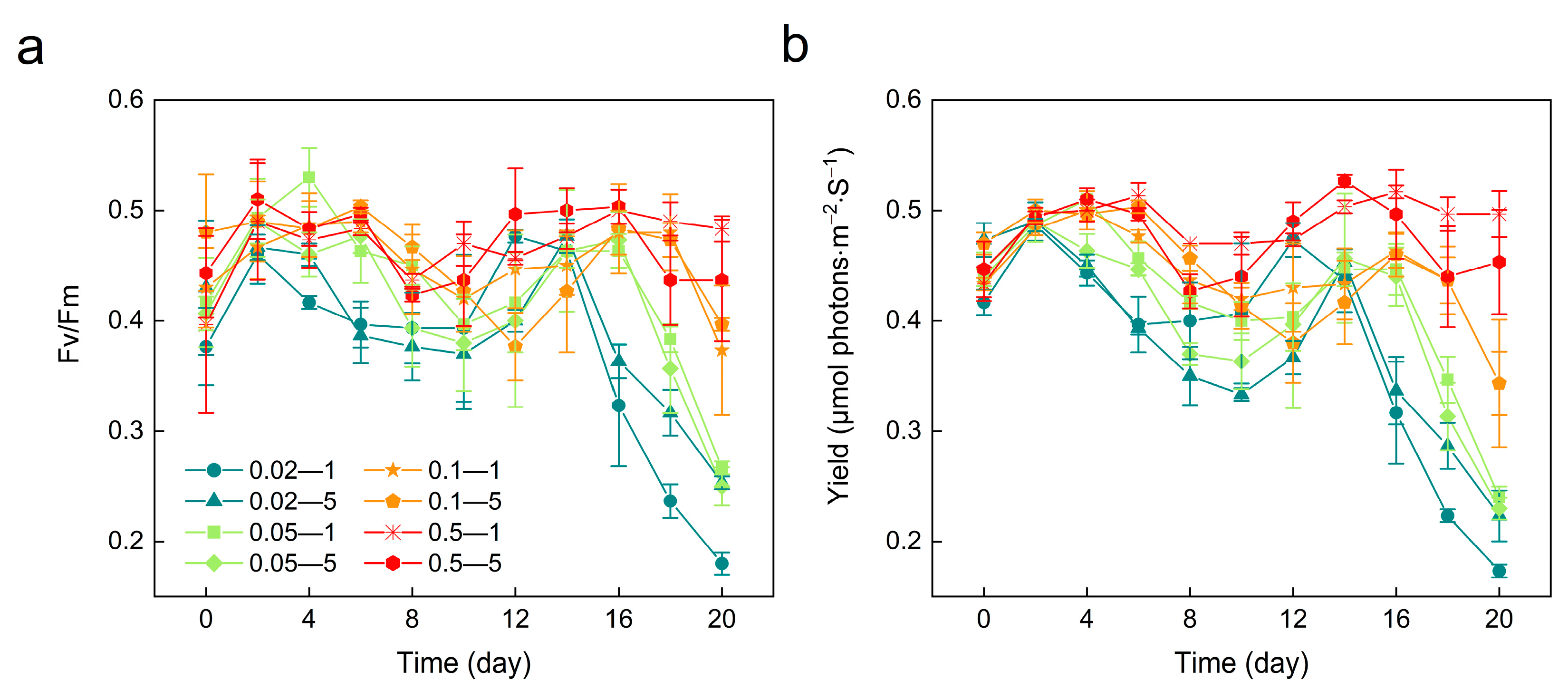
3.3. Photosynthetic Activity Variations
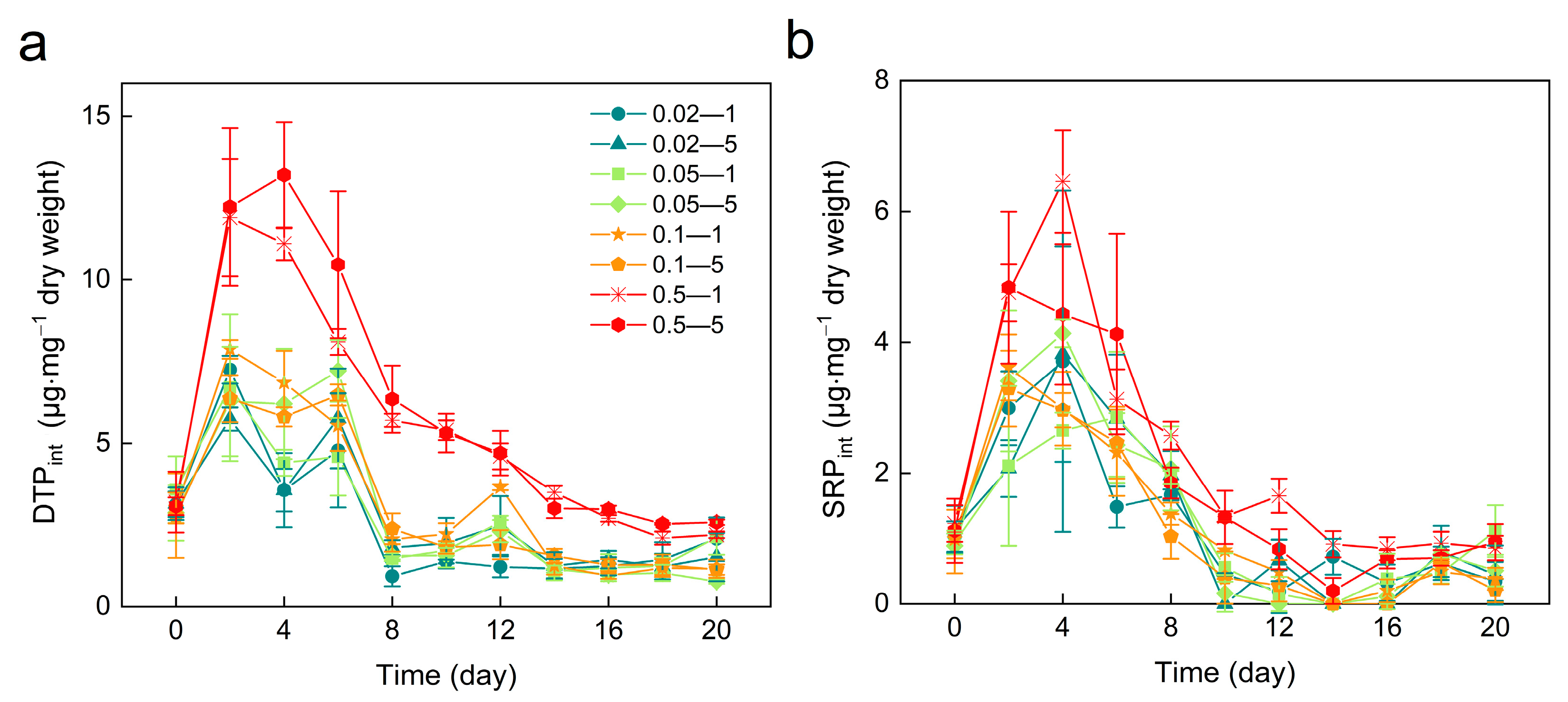
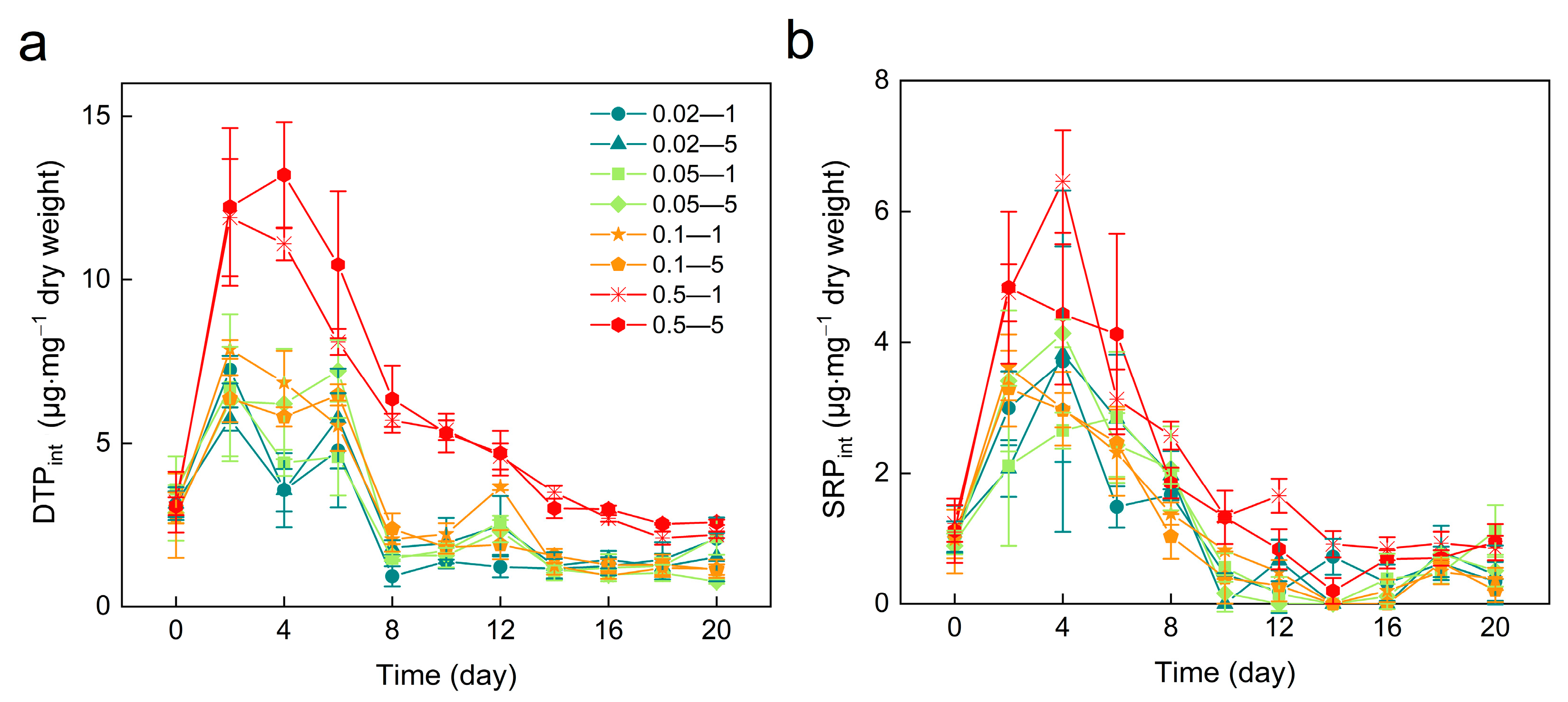
3.4. Intracellular Dissolved Total Phosphorus (DTPint) and Soluble Reactive Phosphorus (SRPint)
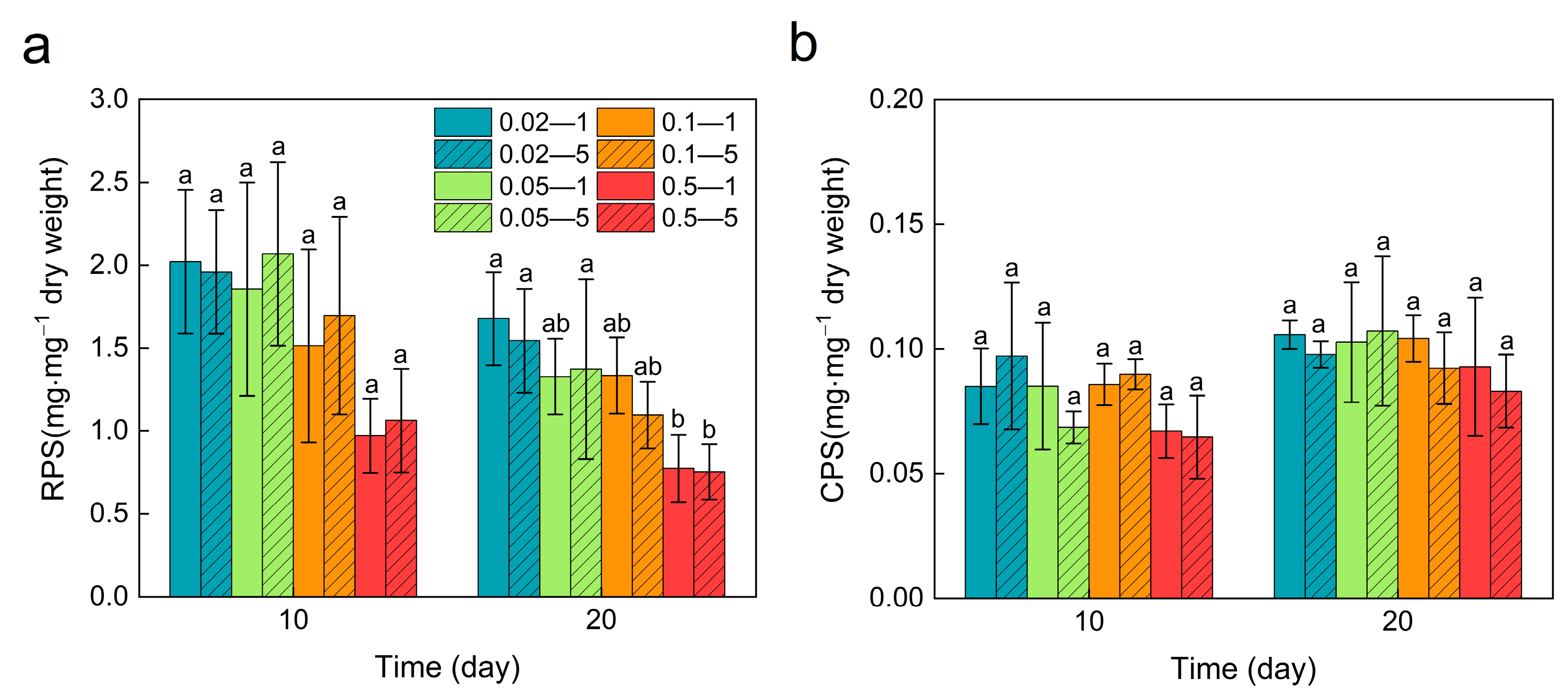
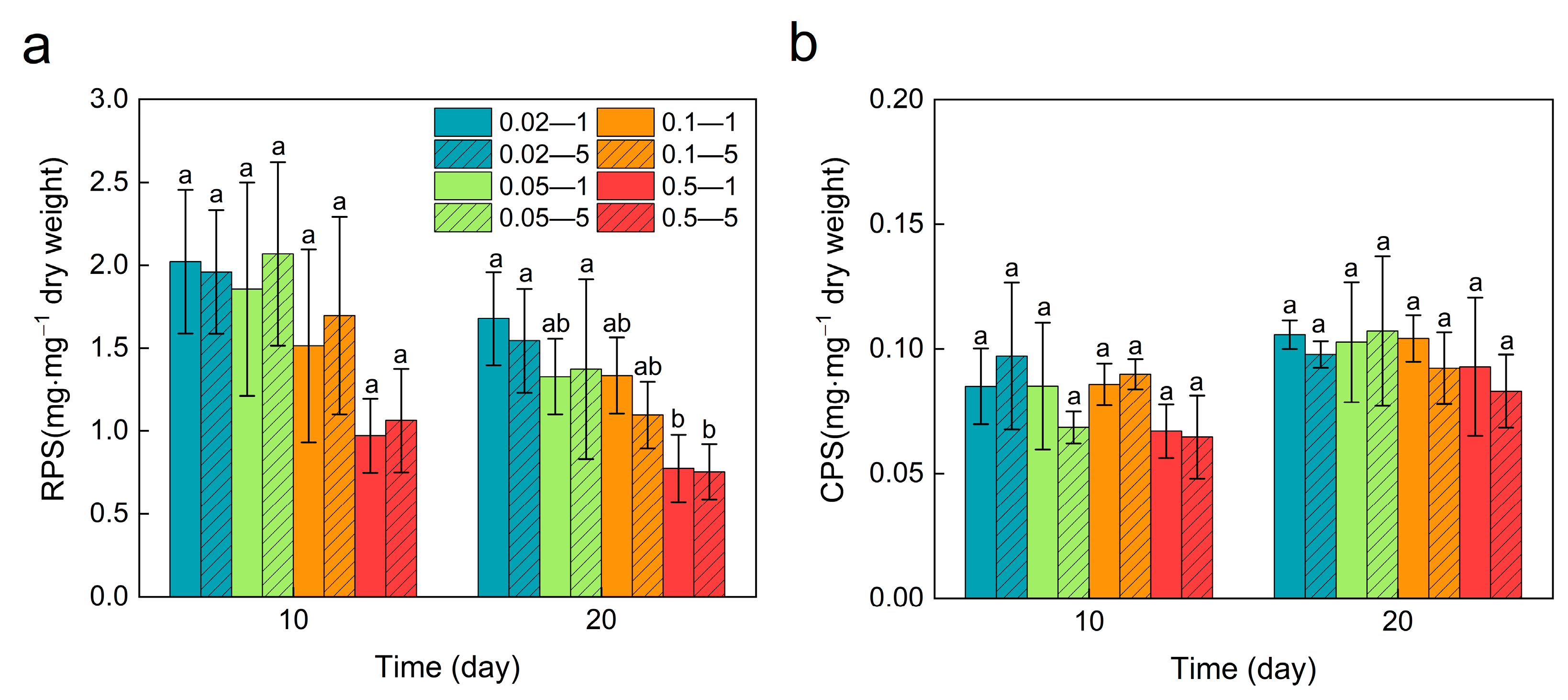
3.5. Exopolysaccharides
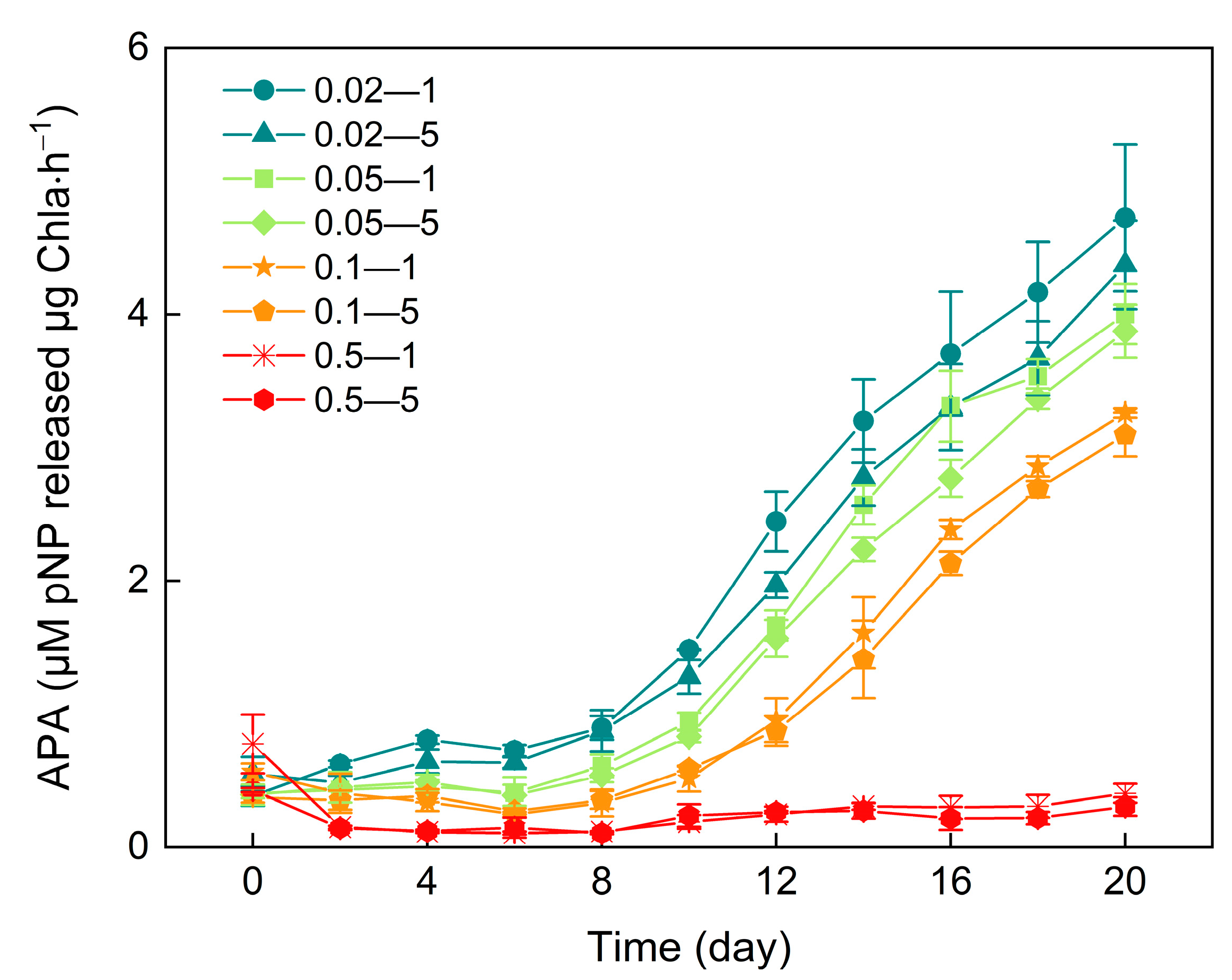
3.6. Alkaline Phosphatase Activity
3.7. Cylindrospermopsin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 21–27. [Google Scholar] [CrossRef]
- Burford, M.A.; Beardall, J.; Willis, A.; Orr, P.T.; Magalhaes, V.F.; Rangel, L.M.; Azevedo, S.M.F.O.E.; Neilan, B.A. Understanding the winning strategies used by the bloom-forming cyanobacterium Cylindrospermopsis raciborskii. Harmful Algae 2016, 54, 44–53. [Google Scholar] [CrossRef]
- Yang, Y.M.; Chen, Y.X.; Cai, F.F.; Liu, X.; Wang, Y.L.; Li, R.H. Toxicity-associated changes in the invasive cyanobacterium Cylindrospermopsis raciborskii in response to nitrogen fluctuations. Environ. Pollut. 2018, 237, 1041–1049. [Google Scholar] [CrossRef]
- Lu, Z.; Ye, J.M.; Chen, Z.J.; Xiao, L.J.; Lei, L.M.; Han, B.P.; Paerl, H.W. Cyanophycin accumulated under nitrogen-fluctuating and high-nitrogen conditions facilitates the persistent dominance and blooms of Raphidiopsis raciborskii in tropical waters. Water Res. 2022, 214, 118215. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.Q.; He, S.H.; Zhao, L.; Ji, L.L.; Yang, S.Q.; Wu, Z.X. Physiological and molecular responses of invasive cyanobacterium Raphidiopsis raciborskii to ambient phosphorus deficiency. J. Oceanol. Limnol. 2022, 40, 1792–1803. [Google Scholar] [CrossRef]
- Zhou, Y.; Nguyen, B.T.; Zhou, C.; Straka, L.; Lai, Y.J.S.A.; Xia, S.Q.; Rittmann, B.E. The distribution of phosphorus and its transformations during batch growth of Synechocystis. Water Res. 2017, 122, 355–362. [Google Scholar] [CrossRef]
- Bar-yosef, Y.; Murik, O.; Sukenik, A.; Hadas, O.; Kaplan, A. Multiannual variations in phytoplankton populations: What distinguished the blooms of Aphanizomenon ovalisporum in Lake Kinneret in 2010 from 2009? Environ. Microbiol. Rep. 2012, 4, 498–503. [Google Scholar] [CrossRef]
- Aubriot, L.; Bonilla, S. Rapid regulation of phosphate uptake in freshwater cyanobacterial blooms. Aquat. Microb. Ecol. 2012, 67, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Amaral, V.; Bonilla, S.; Aubriot, L. Growth optimization of the invasive cyanobacterium Cylindrospermopsis raciborskii in response to phosphate fluctuations. Eur. J. Phycol. 2014, 49, 134–141. [Google Scholar] [CrossRef]
- Akcaalan, R.; Koker, L.; Oguz, A.; Spoof, L.; Meriluoto, J.; Albay, M. First Report of Cylindrospermopsin Production by Two Cyanobacteria (Dolichospermum mendotae and Chrysosporum ovalisporum) in Lake Iznik, Turkey. Toxins 2014, 6, 3173–3186. [Google Scholar] [CrossRef]
- Crawford, A.; Holiday, J.; Merrick., C.; Brayan, J.; Van-Asten, M.; Bowling, L. Use of three monitoring approaches to manage a major Chrysosporum ovalisporum bloom in the Murray River, Australia. Environ. Monit. Assess. 2017, 189, 202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shang, G.X.; Zhang, J.Y.; Yang, L.; Tan, H.X.; Wang, L.Q. First report of bloom forming cyanobacterial species Chrysosporum ovalisporum in China. Plant Sci. J. 2018, 36, 185–190. (In Chinese) [Google Scholar]
- Hadas, O.; Pinkas, R.; Malinsky-Rushansky, N.; Shalev-Alon, G.; Delphine, E.; Berner, T.; Sukenik, A.; Kaplan, A. Physiological variables determined under laboratory conditions may explain the bloom of Aphanizomenon ovalisporum in Lake Kinneret. Eur. J. Phycol. 2002, 37, 259–267. [Google Scholar] [CrossRef]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavement in the water body by toxic Aphanizomenon ovalisporum inducing alkaline phosphatase in phytoplanktons. Curr. Biol. 2010, 20, 1557–1561. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.F.; Yin, S.C.; Chou, Q.C.; Zhou, D.; Jeppesen, E.; Wang, L.Q.; Zhang, W. Community-level and function response of photoautotrophic periphyton exposed to oxytetracycline hydrochloride. Environ. Pollut. 2021, 294, 118593. [Google Scholar] [CrossRef]
- Xu, H.; Paerl, H.W.; Qin, B.; Zhu, G.; Gaoa, G. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China. Limnol. Oceanogr. 2010, 55, 420–432. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.F.; Han, Z.Y.; Wang, M.X.; Ma, Y.C.; Wang, T.; Wang, L.Q.; Zhang, W. Toxicological effects of enrofloxacin and its removal by freshwater microgreen algae Dictyosphaerium sp. Environ. Sci. 2020, 41, 2688–2697. (In Chinese) [Google Scholar]
- Ray, J.M.; Bhaya, D.; Block, M.A.; Grossman, A.R. Isolation, transcription, and inactivation of the gene for an atypical alkaline phosphatase of Synechococcus sp. strain PCC 7942. J. Bacteriol. 1999, 173, 4297–4309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Li, Z.G. Determination of 10 algal toxins in water by UHPLC-MS /MS. Environ. Chem. 2000, 39, 2325–2328. (In Chinese) [Google Scholar]
- Mainston, C.P.; Parr, W. Phosphorus in rivers-ecology and management. Sci. Total Environ. 2002, 282, 25–47. [Google Scholar] [CrossRef]
- Dong, W.G.; Zhao, W.W.; Pang, X.B.; Zhang, M.; Wang, Y.F.; Zeng, D.Q. Chrysosporum ovalisporum—A new record species of cyanobacteria and habitat characteristics of its bloom in an aquaculture pond in northern china. Chin. J. Fish. 2021, 34, 24–30. (In Chinese) [Google Scholar]
- Wu, Z.X.; Zeng, B.; Li, R.H.; Song, L.R. Physiological regulation of Cylindrospermopsis raciborskii (Nostocales, cyanobacteria) in response to inorganic phosphorus limitation. Harmful Algae 2012, 15, 53–58. [Google Scholar] [CrossRef]
- Cruz, A.A.D.L.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.X.; Loftin, K.; Oshea, K.; Zhao, C.; et al. A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Process. Impacts 2013, 15, 1979–2003. [Google Scholar] [CrossRef]
- Bacsi, I.; Vasas, G.; Suranyi, G.; Hamvas, M.; Mathe, C.; Toth, E.; Borbely, G. Alteration of cylindrospermopsin production in sulfate- or phosphate-starved cyanobacterium Aphanizomenon ovalisporum. FEMS Microbiol. Lett. 2006, 269, 303–310. [Google Scholar] [CrossRef]
- Adav, S.S.; Lee, D.J. Extraction of extracellular polymeric substances from aerobic granule with compact interior structure. J. Hazard. Mater. 2008, 154, 1120–1126. [Google Scholar] [CrossRef]
- Konopka, A. The effect of nutrient limitation and its interaction with light upon the products of photosynthesis in Merismopedia tenuissima (Cyanophyceae). J. Phycol. 1983, 19, 403–409. [Google Scholar] [CrossRef]
- Moreno, J.; Vargas, M.A.; Olivares, H.; Rivas, J.; Guerrero, M.G. Exopolysaccharide production by the cyanobacterium Anabaena sp. ATCC 33047 in batch and continuous culture. J. Biotechnol. 1998, 60, 175–182. [Google Scholar] [CrossRef]
- Guerrini, F.; Cangini, M.; Boni, L.; Trost, P.; Pistocchi, R. Metabolic responses of the diatom Achnanthes brevipes (Bacillariophyceae) to nutrient limitation. J. Phycol. 1999, 36, 882–890. [Google Scholar] [CrossRef]
- Song, H.; Lavoie, M.; Fan, X.; Tan, H.; Liu, G.; Xu, P.; Fu, Z.; Paerl, H.W.; Qian, H. Allelopathic interactions of linoleic acid and nitric oxide increase the competitive ability of Microcystis aeruginosa. ISME J. 2017, 11, 1865–1876. [Google Scholar] [CrossRef] [Green Version]
- Urbani, R.; Magaletti, E.; Sist, P.; Cicero, A.M. Extracellular carbohydrates released by the marine diatoms Cylindrotheca Closterium, Thalassiosira pseudonana and Skeletonema costatum: Effect of P-depletion and growth status. Sci. Total Environ. 2005, 353, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Lei, L.M.; Lu, Y.; Peng, L.; Han, B.P. Phosphorus deficiency stimulates dominance of Cylindrospermopsis through facilitating cylindrospermopsin-induced alkaline phosphatase secretion: Integrating field and laboratory-based evidences. Environ. Pollut. 2021, 290, 117946. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, S.; Huang, R.; Wang, N.; Li, C.; Jeppesen, E.; Wang, L.; Zhang, W. Effect of the Pulsed Addition of Phosphorous on the Growth and Toxin Production of the Bloom-Forming Cyanobacterium Chrysosporum ovalisporum (Forti) Zapomelová et al. Water 2023, 15, 351. https://doi.org/10.3390/w15020351
Yin S, Huang R, Wang N, Li C, Jeppesen E, Wang L, Zhang W. Effect of the Pulsed Addition of Phosphorous on the Growth and Toxin Production of the Bloom-Forming Cyanobacterium Chrysosporum ovalisporum (Forti) Zapomelová et al. Water. 2023; 15(2):351. https://doi.org/10.3390/w15020351
Chicago/Turabian StyleYin, Sicheng, Ruohan Huang, Ni Wang, Changxing Li, Erik Jeppesen, Liqing Wang, and Wei Zhang. 2023. "Effect of the Pulsed Addition of Phosphorous on the Growth and Toxin Production of the Bloom-Forming Cyanobacterium Chrysosporum ovalisporum (Forti) Zapomelová et al." Water 15, no. 2: 351. https://doi.org/10.3390/w15020351