
Combining Transcriptomic and Metabolomic Analyses to Investigate the Acute Effects of Microcystin-LR and Nanoplastics of Asian Clams



Abstract
1. Introduction
2. Materials and Methods
2.1. Procurement and Cultivation of Experimental Subjects
2.2. Collection of Samples for NP and MC Exposure
2.3. Sample Processing and Examination
2.4. Bioinformatics Analysis
2.5. Validation of DEGs by qRT-PCR
3. Results
3.1. Transcriptomic Analysis
3.1.1. DEG Analysis
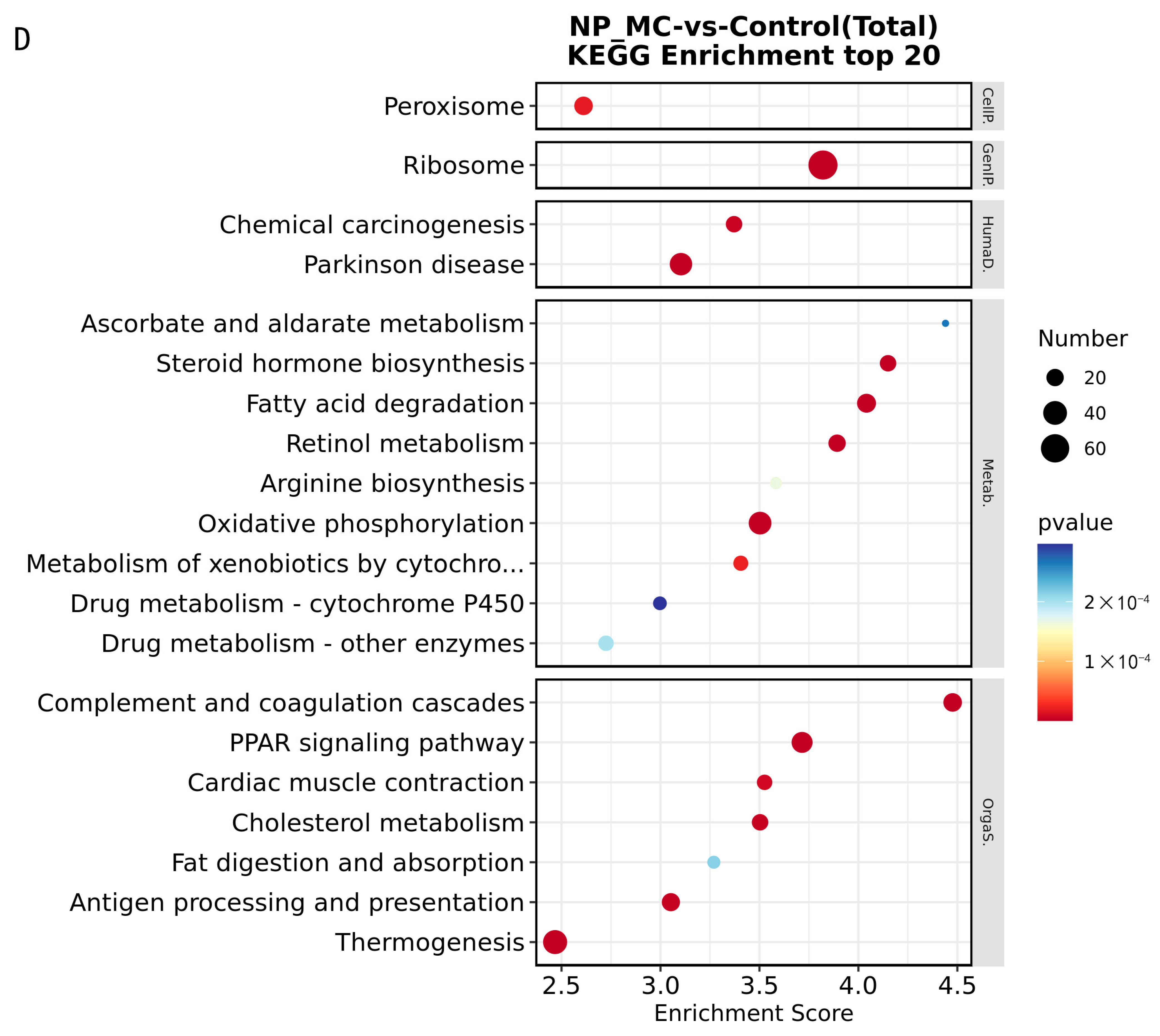
3.1.2. Enrichment Analysis of DEGs
3.2. Metabolomic Analyses
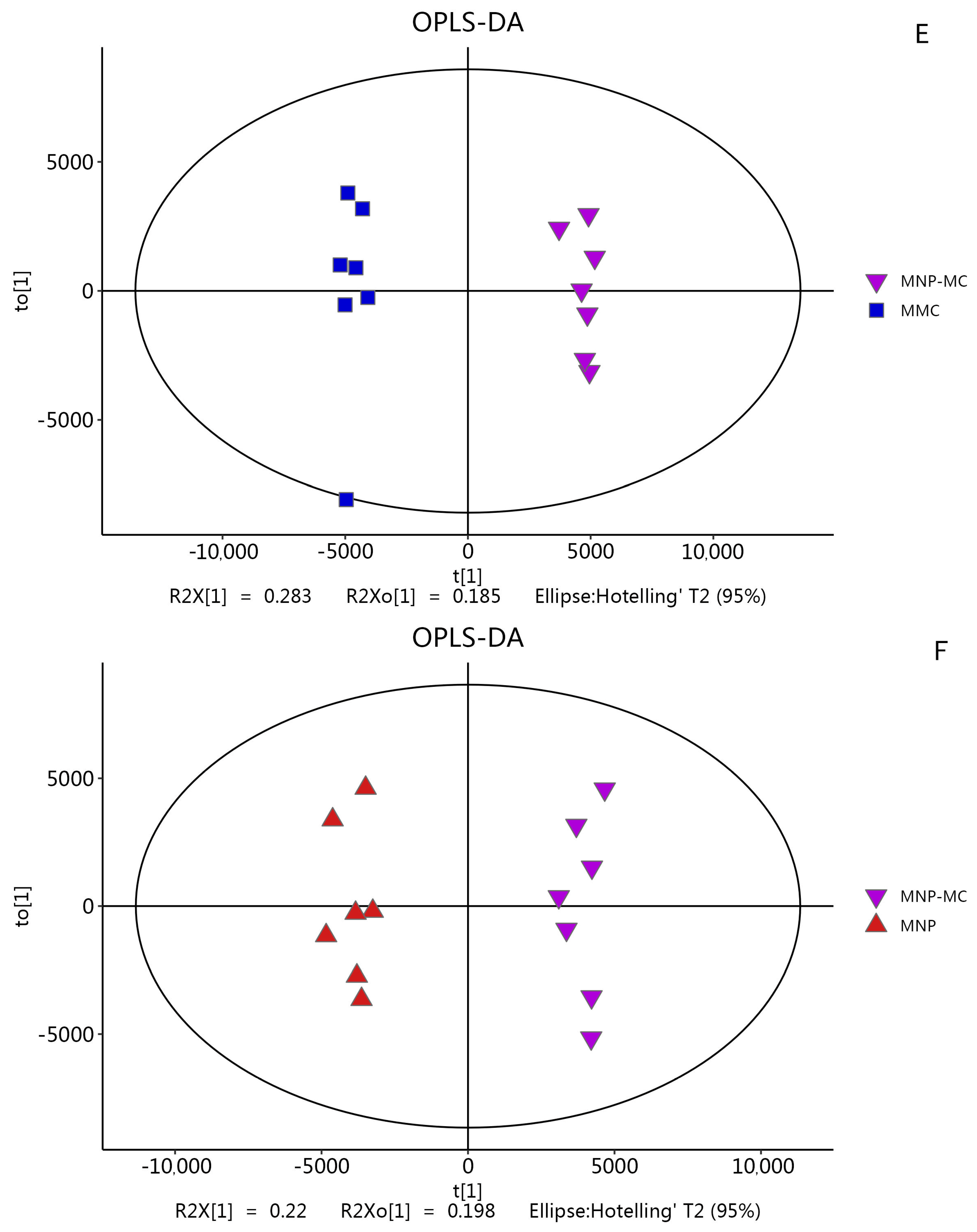
3.2.1. Metabolic Responses to Stress
3.2.2. Analysis of Different Metabolites
3.3. Integrated Transcriptomic and Metabolomic Analysis
4. Discussion
4.1. Analysis of Lysosome Pathway
4.2. Analysis of Citrate Cycle/TCA Cycle Pathway
4.3. Analysis of Histidine Metabolism Pathway
4.4. Analysis of Purine Metabolism Pathway
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Marchant, D.J.; Martínez Rodríguez, A.; Francelle, P.; Jones, J.I.; Kratina, P. Contrasting the Effects of Microplastic Types, Concentrations and Nutrient Enrichment on Freshwater Communities and Ecosystem Functioning. Ecotoxicol. Environ. Saf. 2023, 255, 114834. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Chu, M.L.; Guzman, J.A. Risk Assessment of Harmful Algal Blooms (HAB) Occurrence in the Agroecosystem: A Hydro-Ecologic Modeling Framework and Environmental Risk Matrix. Ecol. Indic. 2022, 145, 109617. [Google Scholar] [CrossRef]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB Science: Directions and Challenges in a Changing Climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef]
- Sun, Y.; Ji, J.; Tao, J.; Yang, Y.; Wu, D.; Han, L.; Li, S.; Wang, J. Current Advances in Interactions between Microplastics and Dissolved Organic Matters in Aquatic and Terrestrial Ecosystems. TrAC Trends Anal. Chem. 2023, 158, 116882. [Google Scholar] [CrossRef]
- Chen, L.; Giesy, J.P.; Adamovsky, O.; Svirčev, Z.; Meriluoto, J.; Codd, G.A.; Mijovic, B.; Shi, T.; Tuo, X.; Li, S.-C.; et al. Challenges of Using Blooms of Microcystis spp. in Animal Feeds: A Comprehensive Review of Nutritional, Toxicological and Microbial Health Evaluation. Sci. Total Environ. 2021, 764, 142319. [Google Scholar] [CrossRef]
- Qu, M.; Nida, A.; Kong, Y.; Du, H.; Xiao, G.; Wang, D. Nanopolystyrene at Predicted Environmental Concentration Enhances Microcystin-LR Toxicity by Inducing Intestinal Damage in Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2019, 183, 109568. [Google Scholar] [CrossRef] [PubMed]
- Straquadine, N.R.W.; Kudela, R.M.; Gobler, C.J. Hepatotoxic Shellfish Poisoning: Accumulation of Microcystins in Eastern Oysters (Crassostrea virginica) and Asian Clams (Corbicula fluminea) Exposed to Wild and Cultured Populations of the Harmful Cyanobacteria, Microcystis. Harmful Algae 2022, 115, 102236. [Google Scholar] [CrossRef]
- Gao, S.; Wu, Q.; Peng, M.; Zeng, J.; Jiang, T.; Ruan, Y.; Xu, L.; Guo, K. Rapid Urbanization Affects Microplastic Communities in Lake Sediments: A Case Study of Lake Aha in Southwest China. J. Environ. Manag. 2023, 338, 117824. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, R.; Zhang, Y.; Mao, L.; Zhu, L.; Zhang, L.; Liu, X.; Jiang, H. Integrate Transcriptomic and Metabolomic Analysis Reveals the Underlying Mechanisms of Behavioral Disorders in Zebrafish (Danio rerio) Induced by Imidacloprid. Sci. Total Environ. 2023, 870, 161541. [Google Scholar] [CrossRef]
- Shi, Q.Q.; Zhang, X.Q.; Zhang, Z.M.; Wang, N.B.; Liu, H.; Zhang, R.R.; Sun, A.L.; Chen, J.; Shi, X.Z. Transcriptome Sequencing and Metabolite Analysis Reveal the Single and Combined Effects of Microplastics and Di-(2-Ethylhexyl) Phthalate on Peneaus vannamei. Sci. Total Environ. 2023, 867, 161549. [Google Scholar] [CrossRef]
- Stephens, A.; Rojo, L.; Araujo-Bernal, S.; Garcia-Carreño, F.; Muhlia-Almazan, A. Cathepsin B from the White Shrimp Litopenaeus vannamei: CDNA Sequence Analysis, Tissues-Specific Expression and Biological Activity. Comp. Biochem. Physiol.—B Biochem. Mol. Biol. 2012, 161, 32–40. [Google Scholar] [CrossRef]
- Lu, Y.-P.; Zheng, P.-H.; Zhang, Z.-L.; Zhang, X.-X.; Li, J.-T.; Wang, D.-M.; Xu, J.-R.; Xian, J.-A.; Wang, A.-L. Hepatopancreas Transcriptome Alterations in Red Claw Crayfish (Cherax quadricarinatus) under Microcystin-LR (MC-LR) Stress. Aquac. Rep. 2023, 29, 101478. [Google Scholar] [CrossRef]
- Yu, J.; Teng, S.; Yue, X.; Wang, H.; Liu, B. The Toll Pathway and Duox-ROS System Are Required for the Clam Antibacterial Immune Response in the Hepatopancreas. Aquaculture 2023, 574, 739637. [Google Scholar] [CrossRef]
- Xu, R.; Zhai, Y.; Yang, J.; Tong, Y.; He, P.; Jia, R. Combined Dynamic Transcriptomics and Metabolomics Analyses Revealed the Effects of Trans-Vp28 Gene Synechocystis sp. PCC6803 on the Hepatopancreas of Litopenaeus vannamei. Fish Shellfish Immunol. 2022, 128, 28–37. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-Mcintyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for Large-Scale Metabolic Profiling of Serum and Plasma Using Gas Chromatography and Liquid Chromatography Coupled to Mass Spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K.; Want, E.J. Global Metabolic Profiling Procedures for Urine Using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar]
- Zhu, B.; Zhang, Z.; Wang, X.; Zhang, W.; Shi, H.; Song, Z.; Ding, S.; Yang, X. Abnormal Histidine Metabolism Promotes Macrophage Lipid Accumulation under Ox-LDL Condition. Biochem. Biophys. Res. Commun. 2022, 588, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.K.; Mukesh, J.; Liu, Z. NGS QC Toolkit: A Toolkit for Quality Control of Next Generation Sequencing Data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Duan, Y.; Xing, Y.; Zhu, X.; Li, H.; Wang, Y.; Nan, Y. Integration of Transcriptomic and Metabolomic Reveals Carbonate Alkalinity Stress Responses in the Hepatopancreas of Litopenaeus vannamei. Aquat. Toxicol. 2023, 260, 106569. [Google Scholar] [CrossRef]
- Beltrame, L.; Calura, E.; Popovici, R.; Draghici, S.; Cavalieri, D.; D’Incalci, M. 666 The Biological Connection Markup Language—A Data Format to Visualize, Annotate and Analyze Biological Pathways. Eur. J. Cancer 2012, 48, S158. [Google Scholar] [CrossRef]
- Cannataro, M.; Guzzi, P.H.; Agapito, G.; Zucco, C.; Milano, M. (Eds.) Chapter 14—Biological Pathway Analysis; Elsevier: Amsterdam, The Netherlands, 2022; pp. 151–165. ISBN 978-0-12-822952-1. [Google Scholar]
- Yao, C.L.; Wu, C.G.; Xiang, J.H.; Li, F.; Wang, Z.Y.; Han, X. The Lysosome and Lysozyme Response in Chinese Shrimp Fenneropenaeus chinensis to Vibrio anguillarum and Laminarin Stimulation. J. Exp. Mar. Biol. Ecol. 2008, 363, 124–129. [Google Scholar] [CrossRef]
- Xu, Y.; Pan, H.; Lu, M.; Liu, Q.; Shafique, L.; Peng, J.; Ahmed, T.; Wang, R.; Zhang, H.; Wang, Q.; et al. Analysis of Transcripts and Splice Isoforms in Red Claw Crayfish (Cherax quadricarinatus) Using Single-Molecule Long-Read Sequencing. Aquaculture 2021, 541, 736828. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, S.H.; Di, J.; Du, H.; Wei, Q.W. Inactivated Mycobacterium Vaccine Induces Innate Immunity against M. Ulcerans Ecovar Liflandii (MuLiflandii ASM001) Infection in Dabry’s Sturgeon (Acipenser dabryanus). Aquaculture 2019, 512, 734312. [Google Scholar] [CrossRef]
- Longarini, E.J.; Dauben, H.; Locatelli, C.; Wondisford, A.R.; Smith, R.; Muench, C.; Kolvenbach, A.; Lynskey, M.L.; Pope, A.; Bonfiglio, J.J.; et al. Modular Antibodies Reveal DNA Damage-Induced Mono-ADP-Ribosylation as a Second Wave of PARP1 Signaling. Mol. Cell 2023, 83, 1743–1760. [Google Scholar] [CrossRef]
- Alu, A.; Han, X.; Ma, X.; Wu, M.; Wei, Y.; Wei, X. The Role of Lysosome in Regulated Necrosis. Acta Pharm. Sin. B 2020, 10, 1880–1903. [Google Scholar] [CrossRef]
- Cacabelos, R.; Carril, J.C.; Sanmartín, A.; Cacabelos, P. Chapter 6—Pharmacoepigenetic Processors: Epigenetic Drugs, Drug Resistance, Toxicoepigenetics, and Nutriepigenetics. In Translational Epigenetics; Cacabelos, R.B.T.-P., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 10, pp. 191–424. ISBN 9780128139394. [Google Scholar]
- Li, M.; Zhang, K.; Long, R.; Sun, Y.; Kang, J.; Zhang, T.; Cao, S. ITRAQ-Based Comparative Proteomic Analysis Reveals Tissue-Specific and Novel Early-Stage Molecular Mechanisms of Salt Stress Response in Carex Rigescens. Environ. Exp. Bot. 2017, 143, 99–114. [Google Scholar] [CrossRef]
- Cui, Q.; Qiu, L.; Yang, X.; Shang, S.; Yang, B.; Chen, M.; Liu, X.; Chen, B.; Fu, X.; Wang, W.; et al. Transcriptome Profiling of the Low-Salinity Stress Responses in the Gills of the Juvenile Pseudopleuronectes yokohamae. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 32, 100612. [Google Scholar] [CrossRef]
- Dell’Antone, P. Proton Pump-Linked Mg2+-ATPase Activity in Isolated Rat Liver Lysosomes. Arch. Biochem. Biophys. 1988, 262, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.; Nakanishi-Matsui, M. Proton Pumping V-ATPase Inhibitor Bafilomycin A1 Affects Rab7 Lysosomal Localization and Abolishes Anterograde Trafficking of Osteoclast Secretory Lysosomes. Biochem. Biophys. Res. Commun. 2019, 510, 421–426. [Google Scholar] [CrossRef]
- Han, X.; Tang, Y.; Zhang, Y.; Zhang, J.; Hu, Z.; Xu, W.; Xu, S.; Niu, Q. Impaired V-ATPase Leads to Increased Lysosomal PH, Results in Disrupted Lysosomal Degradation and Autophagic Flux Blockage, Contributes to Fluoride-Induced Developmental Neurotoxicity. Ecotoxicol. Environ. Saf. 2022, 236, 113500. [Google Scholar] [CrossRef]
- Collins, M.P.; Forgac, M. Regulation and Function of V-ATPases in Physiology and Disease. Biochim. Biophys. Acta—Biomembr. 2020, 1862, 183341. [Google Scholar] [CrossRef]
- Ruprecht, J.J.; King, M.S.; Zögg, T.; Aleksandrova, A.A.; Pardon, E.; Crichton, P.G.; Steyaert, J.; Kunji, E.R.S. The Molecular Mechanism of Transport by the Mitochondrial ADP/ATP Carrier. Cell 2019, 176, 435–447.e15. [Google Scholar] [CrossRef] [PubMed]
- Chara, O.; Pafundo, D.E.; Schwarzbaum, P.J. Negative Feedback of Extracellular ADP on ATP Release in Goldfish Hepatocytes: A Theoretical Study. J. Theor. Biol. 2010, 264, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Springett, R.; King, M.S.; Crichton, P.G.; Kunji, E.R.S. Modelling the Free Energy Profile of the Mitochondrial ADP/ATP Carrier. Biochim. Biophys. Acta—Bioenerg. 2017, 1858, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Baars, T.L.; Petri, S.; Peters, C.; Mayer, A. Role of the V-ATPase in Regulation of the Vacuolar Fission–Fusion Equilibrium. Mol. Biol. Cell 2007, 18, 3873. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.S.; Sun, Y.; Sun, Y.X.; Zhu, B.J.; Liu, C.L. Characterization and Function of a Cathepsin B in Red Crayfish (Procambarus clarkii) Following Lipopolysaccharide Challenge. Fish Shellfish Immunol. 2016, 56, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Yang, Y.; Guo, Z.; Feng, S.; Wan, Y. A Cathepsin B/GSH Dual-Responsive Fluorinated Peptide for Effective SiRNA Delivery to Cancer Cells. Bioorg. Chem. 2023, 135, 106485. [Google Scholar] [CrossRef]
- Shen, X.B.; Chen, X.; Zhang, Z.Y.; Wu, F.F.; Liu, X.H. Cathepsin C Inhibitors as Anti-Inflammatory Drug Discovery: Challenges and Opportunities. Eur. J. Med. Chem. 2021, 225, 113818. [Google Scholar] [CrossRef]
- Lacoue-Labarthe, T.; Le Bihan, E.; Borg, D.; Koueta, N.; Bustamante, P. Paco Acid Phosphatase and Cathepsin Activity in Cuttlefish (Sepia Officinalis) Eggs: The Effects of Ag, Cd, and Cu Exposure. ICES J. Mar. Sci. 2010, 67, 1517–1523. [Google Scholar] [CrossRef][Green Version]
- Aranishi, F.; Mano, N. Response of Skin Cathepsins to Infection of Edwardsiella tarda in Japanese Flounder. Fish. Sci. 2000, 66, 169–170. [Google Scholar] [CrossRef]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory Metabolism: Glycolysis, the TCA Cycle and Mitochondrial Electron Transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef]
- Nazaret, C.; Heiske, M.; Thurley, K.; Mazat, J.-P. Mitochondrial Energetic Metabolism: A Simplified Model of TCA Cycle with ATP Production. J. Theor. Biol. 2009, 258, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.H.D.; Donnelly, C.; Komlódi, T.; Gnaiger, E. Characterizing Electron Transfer through the Mitochondrial Q-Junction from Fatty Acid Oxidation and TCA Cycle on HEK 293T Cells: Correlation with Respiration. Biochim. Biophys. Acta—Bioenerg. 2022, 1863, 148866. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Beard, K.F.M.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not Just a Circle: Flux Modes in the Plant TCA Cycle. Trends Plant Sci. 2010, 15, 462–470. [Google Scholar] [CrossRef]
- Mishra, R.; Shukla, S.P. Endosulfan Effects on Muscle Malate Dehydrogenase of the Freshwater Catfish Clarias Batrachus. Ecotoxicol. Environ. Saf. 2003, 56, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Ye, J.; Song, M.; Peng, X.; Li, H. Poly I:C Promotes Malate to Enhance Innate Immune Response against Bacterial Infection. Fish Shellfish Immunol. 2022, 131, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Smits, M.; Artigaud, S.; Bernay, B.; Pichereau, V.; Bargelloni, L.; Paillard, C. A Proteomic Study of Resistance to Brown Ring Disease in the Manila Clam, Ruditapes philippinarum. Fish Shellfish Immunol. 2020, 99, 641–653. [Google Scholar] [CrossRef]
- Meng, J.; Zhang, G.; Wang, W.-X. Functional Heterogeneity of Immune Defenses in Molluscan Oysters Crassostrea Hongkongensis Revealed by High-Throughput Single-Cell Transcriptome. Fish Shellfish Immunol. 2022, 120, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B.; Tabor, H. N-Formimino-l-Glutamate Iminohydrolase from Histidine-Adapted Pseudomonas: Purification and properties. J. Biol. Chem. 1972, 247, 1605–1609. [Google Scholar] [CrossRef] [PubMed]
- Kaminskas, E.; Kimhi, Y.; Magasanik, B. Urocanase and N-Formimino-l-Glutamate Formiminohydrolase of Bacillus Subtilis, Two Enzymes of the Histidine Degradation Pathway. J. Biol. Chem. 1970, 245, 3536–3544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, D.; Yu, J.; Su, X.; Li, L. Metabolic Perturbations in Human Hepatocytes Induced by Bis (2-Ethylhexyl)-2,3,4,5-Tetrabromophthalate Exposure: Insights from High-Coverage Quantitative Metabolomics. Anal. Biochem. 2022, 657, 114887. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.; Oberrauch, S.; Allobawi, R.; Cornthwaite-Duncan, L.; Lu, J.; Sharma, R.; Baker, M.; Li, J.; Rao, G.G.; Velkov, T. Untargeted Metabolomics to Evaluate Polymyxin B Toxicodynamics Following Direct Intracerebroventricular Administration into the Rat Brain. Comput. Struct. Biotechnol. J. 2022, 20, 6067–6077. [Google Scholar] [CrossRef] [PubMed]
- Michelato, M.; Zaminhan, M.; Boscolo, W.R.; Nogaroto, V.; Vicari, M.; Artoni, R.F.; Furuya, V.R.B.; Furuya, W.M. Dietary Histidine Requirement of Nile Tilapia Juveniles Based on Growth Performance, Expression of Muscle-Growth-Related Genes and Haematological Responses. Aquaculture 2017, 467, 63–70. [Google Scholar] [CrossRef]
- Wu, P.; Qu, B.; Feng, L.; Jiang, W.-D.; Kuang, S.-Y.; Jiang, J.; Tang, L.; Zhou, X.-Q.; Liu, Y. Dietary Histidine Deficiency Induced Flesh Quality Loss Associated with Changes in Muscle Nutritive Composition, Antioxidant Capacity, Nrf2 and TOR Signaling Molecules in on-Growing Grass Carp (Ctenopharyngodon idella). Aquaculture 2020, 526, 735399. [Google Scholar] [CrossRef]
- Gao, Y.-J.; Liu, Y.-J.; Chen, X.-Q.; Yang, H.-J.; Li, X.-F.; Tian, L.-X. Effects of Graded Levels of Histidine on Growth Performance, Digested Enzymes Activities, Erythrocyte Osmotic Fragility and Hypoxia-Tolerance of Juvenile Grass Carp Ctenopharyngodon idella. Aquaculture 2016, 452, 388–394. [Google Scholar] [CrossRef]
- Sarih, S.; Djellata, A.; Roo, J.; Hernández-Cruz, C.M.; Fontanillas, R.; Rosenlund, G.; Izquierdo, M.; Fernández-Palacios, H. Effects of Increased Protein, Histidine and Taurine Dietary Levels on Egg Quality of Greater Amberjack (Seriola dumerili, Risso, 1810). Aquaculture 2019, 499, 72–79. [Google Scholar] [CrossRef]
- Furuta, T.; Takahashi, H.; Shibasaki, H.; Kasuya, Y. Reversible Stepwise Mechanism Involving a Carbanion Intermediate in the Elimination of Ammonia from L-Histidine Catalyzed by Histidine Ammonia-Lyase. J. Biol. Chem. 1992, 267, 12600–12605. [Google Scholar] [CrossRef]
- Hubert, L.; Sutton, V.R. Chapter 12—Disorders of Purine and Pyrimidine Metabolism. In Clinical Aspects and Laboratory Determination; Garg, U., Smith, L., Eds.; Elsevier: San Diego, CA, USA, 2017; pp. 283–299. ISBN 978-0-12-802896-4. [Google Scholar]
- Carballeira, J.D.; Fernandez-Lucas, J.; Quezada, M.A.; Hernaiz, M.J.; Alcantara, A.R.; Simeó, Y.; Sinisterra, J.V. Biotransformations, 3rd ed.; Academic Press: Oxford, UK, 2009; pp. 212–251. ISBN 978-0-12-373944-5. [Google Scholar]
- Baldissera, M.D.; Souza, C.F.; Doleski, P.H.; Guarda, N.S.; Moresco, R.N.; Santos, R.C.V.; Baldisserotto, B. Stimulation of Splenic and Lymphocytic Acetylcholinesterase and Adenosine Deaminase Activities in Rhamdia quelen Experimentally Infected with Pseudomonas aeruginosa: Impairment of Immune System. Aquaculture 2017, 473, 417–422. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Boaventura, T.P.; Nakayama, C.L.; Baldisserotto, B.; Luz, R.K. Purinergic Signaling as a Potential Target of Hypoxia Stress-Induced Impairment of the Immune System in Freshwater Catfish Lophiosilurus alexandri. Aquaculture 2018, 496, 197–202. [Google Scholar] [CrossRef]
- Hossain, M.S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M.; Ono, S.; Fujieda, T. Comparison of the Effects of Inosine and Inosine Monophosphate on Growth, Immune Response, Stress Resistance and Gut Morphology of Juvenile Red Sea Bream, Pagrus Major. Aquaculture 2016, 458, 64–74. [Google Scholar] [CrossRef]
- Miller, S.G.; Hafen, P.S.; Brault, J.J. Increased Adenine Nucleotide Degradation in Skeletal Muscle Atrophy. Int. J. Mol. Sci. 2020, 21, 88. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M.; Dawood, M.A.O.; Kader, M.A.; Bulbul, M.; Fujieda, T. Efficacy of Nucleotide Related Products on Growth, Blood Chemistry, Oxidative Stress and Growth Factor Gene Expression of Juvenile Red Sea Bream, Pagrus Major. Aquaculture 2016, 464, 8–16. [Google Scholar] [CrossRef]
- Kambe, J.; Miyata, S.; Li, C.; Yamamoto, Y.; Nagaoka, K. Xanthine-Induced Deficits in Hippocampal Behavior and Abnormal Expression of Hemoglobin Genes. Behav. Brain Res. 2023, 449, 114476. [Google Scholar] [CrossRef]
- Xu, R.; Zheng, R.; Wang, Y.; Ma, R.; Tong, G.; Wei, X.; Feng, D.; Hu, K. Transcriptome Analysis to Elucidate the Toxicity Mechanisms of Fenvalerate, Sulfide Gatifloxacin, and Ridomil on the Hepatopancreas of Procambarus clarkii. Fish Shellfish Immunol. 2021, 116, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Perera, N.C.N.; Godahewa, G.I.; Lee, S.; Kim, M.-J.; Hwang, J.Y.; Kwon, M.G.; Hwang, S.D.; Lee, J. Manganese-Superoxide Dismutase (MnSOD), a Role Player in Seahorse (Hippocampus abdominalis) Antioxidant Defense System and Adaptive Immune System. Fish Shellfish Immunol. 2017, 68, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Ishibe, K.; Osatomi, K.; Hara, K.; Kanai, K.; Yamaguchi, K.; Oda, T. Comparison of the Responses of Peritoneal Macrophages from Japanese Flounder (Paralichthys olivaceus) against High Virulent and Low Virulent Strains of Edwardsiella tarda. Fish Shellfish Immunol. 2008, 24, 243–251. [Google Scholar] [CrossRef]

























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward and Reverse Primer (5′–3′) | TM (°C) | Amplicon Length (bp) | Application Efficiency |
|---|---|---|---|---|
| β-actin | ACCTATCTATGAGGGGTACGCC | 61 | 136 | 100.2% |
| AATTTCTCTTTCGGCTGTTGTG | 58 | |||
| COX1 | ATGTTCTATCAATGGGAGC | 58 | 190 | 102.1% |
| TCTGAGTAGCGTCGTGGT | 56 | |||
| COX3 | GATTCTTCTGAGCGTTCTAT | 52 | 187 | 104.3% |
| TGGTTTCGTTTACCTTCTAT | 53 | |||
| GADD45 | TCGAGTGTGCAAGCTACCTG | 60 | 147 | 99.8% |
| CCTTGACAACGTGTATGTCGTT | 59 | |||
| TLR1 | ATACCGCTAAATGTTCCAAG | 54 | 179 | 99.4% |
| GTACTGAAAGGCGTGATGTT | 56 | |||
| NFKB | ATTTGGAGGATTGAATGTAG | 55 | 240 | 99.7% |
| ATAATGTGAGTTCCGTTGTC | 52 | |||
| TBK1 | AGAACTGCTGAAGGCTGTCAAC | 61 | 167 | 103.8% |
| ATTCCTTCTCCTCAAAGCCAAG | 58 | |||
| NBR1 | TGTGAATCCTGCGAGGACAA | 57 | 184 | 101.6% |
| CGCCTTCTGGATAGATGGGT | 59 | |||
| CASP8 | ACCTCATAGGCTATTCCACG | 55 | 155 | 102.7% |
| GGCTACCTTTCGGTTTACTT | 54 | |||
| CASP7 | CCGACGAAGACAAAGAGGTT | 56 | 189 | 97.4% |
| ACTCCATGTTTCCAGCGAAG | 57 |
| Group | Pathway Name | Up Stream | Down Stream | Kegg Level 2 |
|---|---|---|---|---|
| Glycolysis/Gluconeogenesis | 1 | 17 | Carbohydrate metabolism | |
| MC-control | Citrate cycle (TCA cycle) | 3 | 14 | Carbohydrate metabolism |
| Alanine, aspartate, and glutamate metabolism | 14 | 2 | Amino acid metabolism | |
| Cysteine and methionine metabolism | 4 | 1 | Amino acid metabolism | |
| NP-control | Arginine and proline metabolism | 4 | 1 | Metabolism of other amino acids |
| Glutathione metabolism | 2 | 2 | Amino acid metabolism | |
| Alanine, aspartate, and glutamate metabolism | 16 | 1 | Amino acid metabolism | |
| NP-MC -control | Glycolysis/Gluconeogenesis | 0 | 17 | Carbohydrate metabolism |
| Citrate cycle (TCA cycle) | 1 | 13 | Carbohydrate metabolism | |
| Glycolysis/Gluconeogenesis | 1 | 8 | Carbohydrate metabolism | |
| NP-MC | Glycine, serine, and threonine metabolism | 2 | 6 | Amino acid metabolism |
| Alanine, aspartate, and glutamate metabolism | 7 | 1 | Amino acid metabolism | |
| Alanine, aspartate, and glutamate metabolism | 10 | 4 | Amino acid metabolism | |
| NP-MC -MC | Glycine, serine, and threonine metabolism | 0 | 10 | Amino acid metabolism |
| Arginine and proline metabolism | 5 | 3 | Amino acid metabolism | |
| Alanine, aspartate, and glutamate metabolism | 10 | 5 | Amino acid metabolism | |
| NP-MC -NP | Glycine, serine, and threonine metabolism | 1 | 10 | Amino acid metabolism |
| Glycolysis/Gluconeogenesis | 1 | 10 | Carbohydrate metabolism |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wang, J.; Liu, S.; Zhou, Y.; Liu, X. Combining Transcriptomic and Metabolomic Analyses to Investigate the Acute Effects of Microcystin-LR and Nanoplastics of Asian Clams. Water 2023, 15, 3519. https://doi.org/10.3390/w15193519
Zhang J, Wang J, Liu S, Zhou Y, Liu X. Combining Transcriptomic and Metabolomic Analyses to Investigate the Acute Effects of Microcystin-LR and Nanoplastics of Asian Clams. Water. 2023; 15(19):3519. https://doi.org/10.3390/w15193519
Chicago/Turabian StyleZhang, Jiahua, Jie Wang, Shikun Liu, Yin Zhou, and Xingguo Liu. 2023. "Combining Transcriptomic and Metabolomic Analyses to Investigate the Acute Effects of Microcystin-LR and Nanoplastics of Asian Clams" Water 15, no. 19: 3519. https://doi.org/10.3390/w15193519
APA StyleZhang, J., Wang, J., Liu, S., Zhou, Y., & Liu, X. (2023). Combining Transcriptomic and Metabolomic Analyses to Investigate the Acute Effects of Microcystin-LR and Nanoplastics of Asian Clams. Water, 15(19), 3519. https://doi.org/10.3390/w15193519