
Relationship between Cyanobacterial Abundance and Physicochemical Variables in the Ebro Basin Reservoirs (Spain)
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
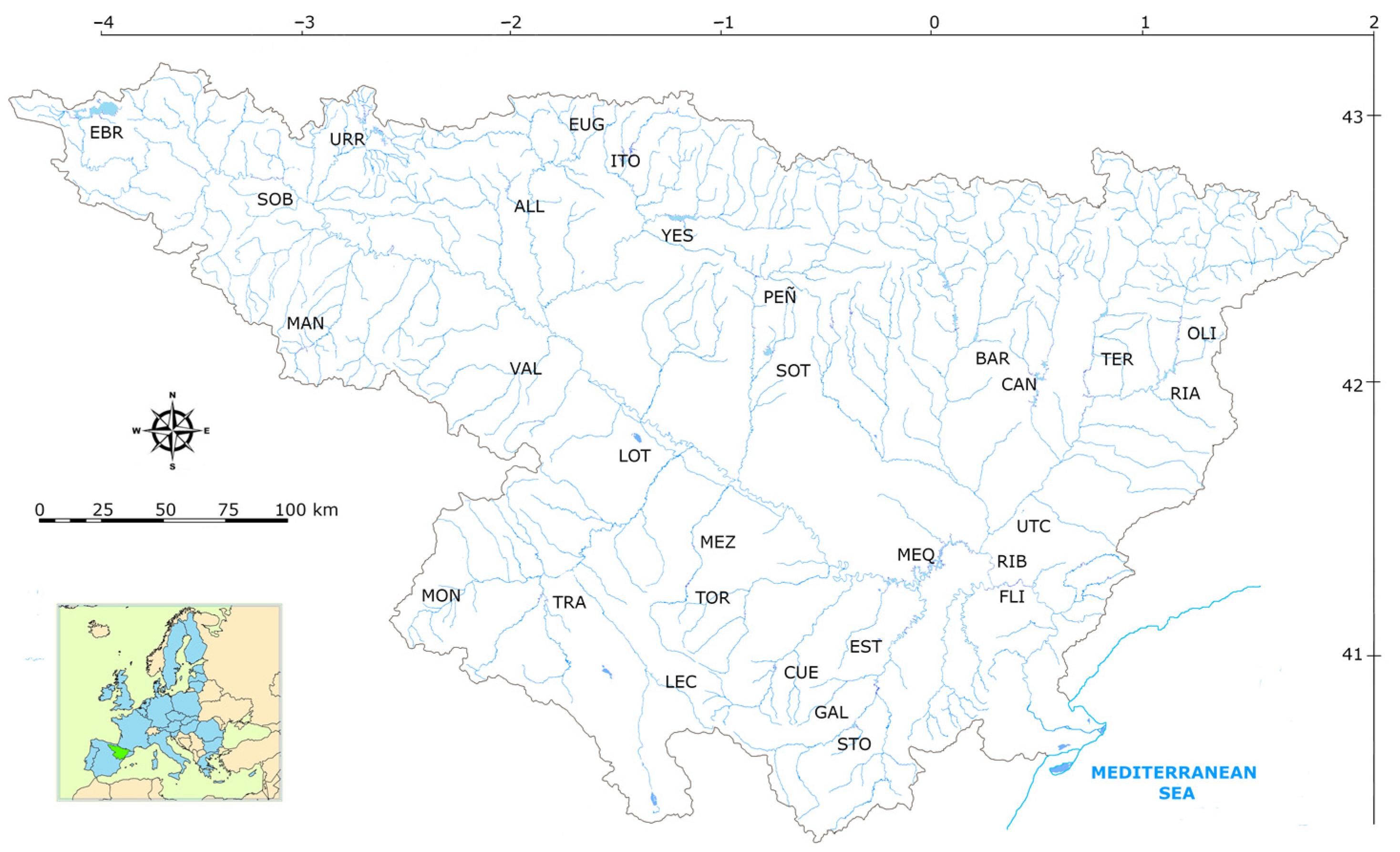
2.1. Study Area
2.2. In Situ Sampling
2.3. Laboratory Analysis
2.3.1. Chemical Variables
2.3.2. Phytoplankton
2.4. Data Processing
2.5. WHO Classification of Water Bodies for Human Health Risk from Cyanobacteria
2.6. Remote Sensing
3. Results
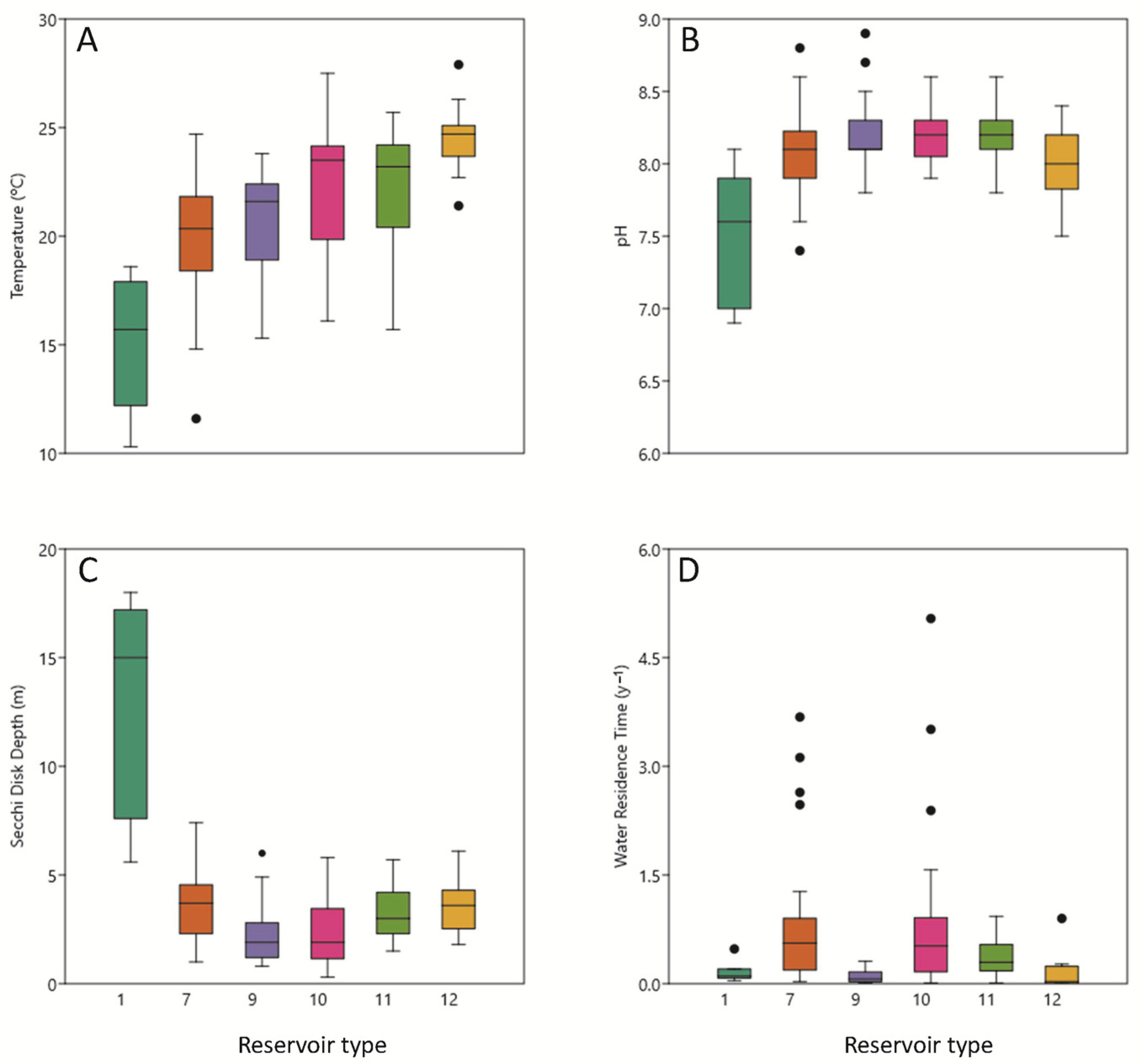
3.1. Physical Conditions
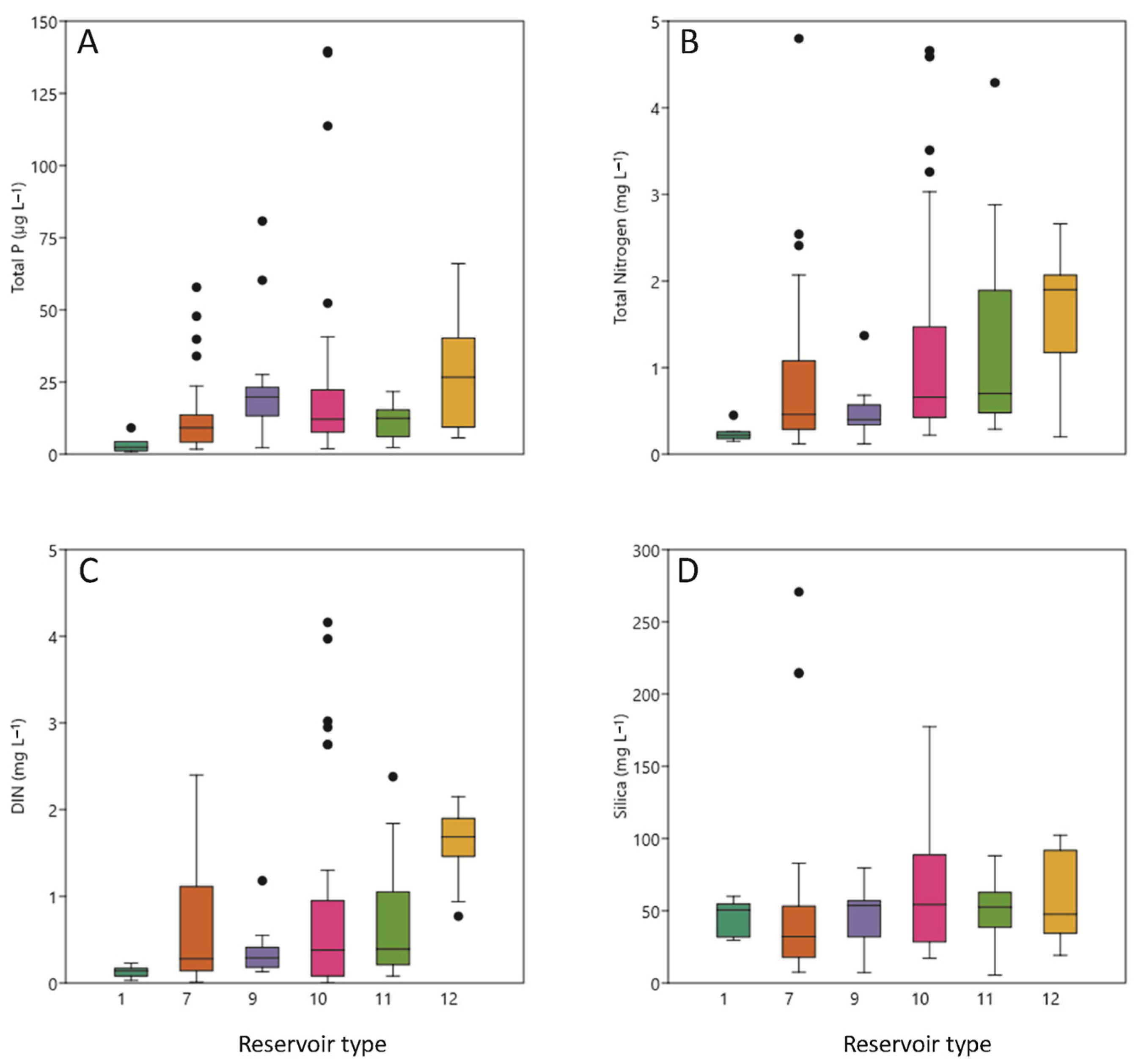
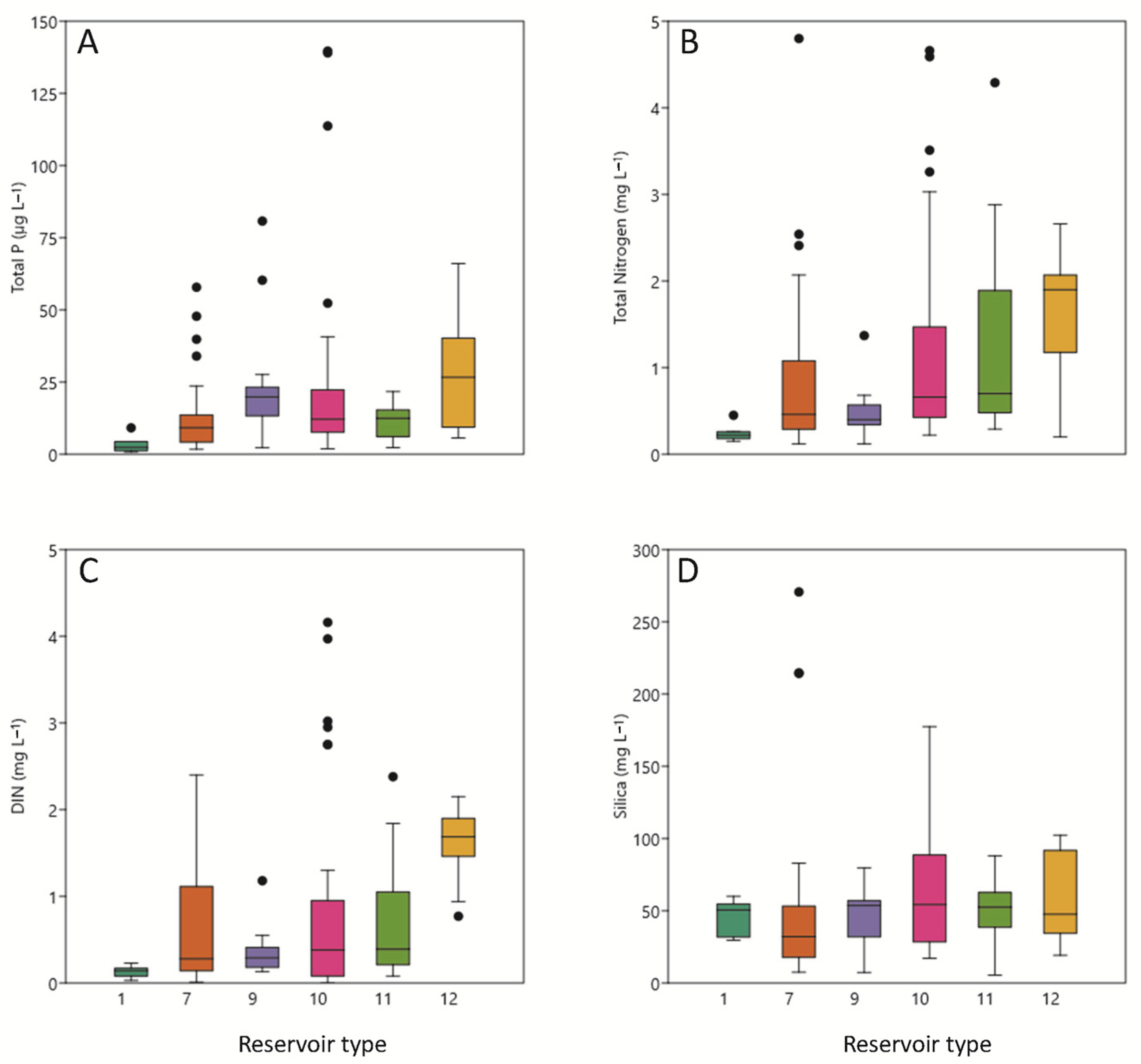
3.2. Chemical Conditions
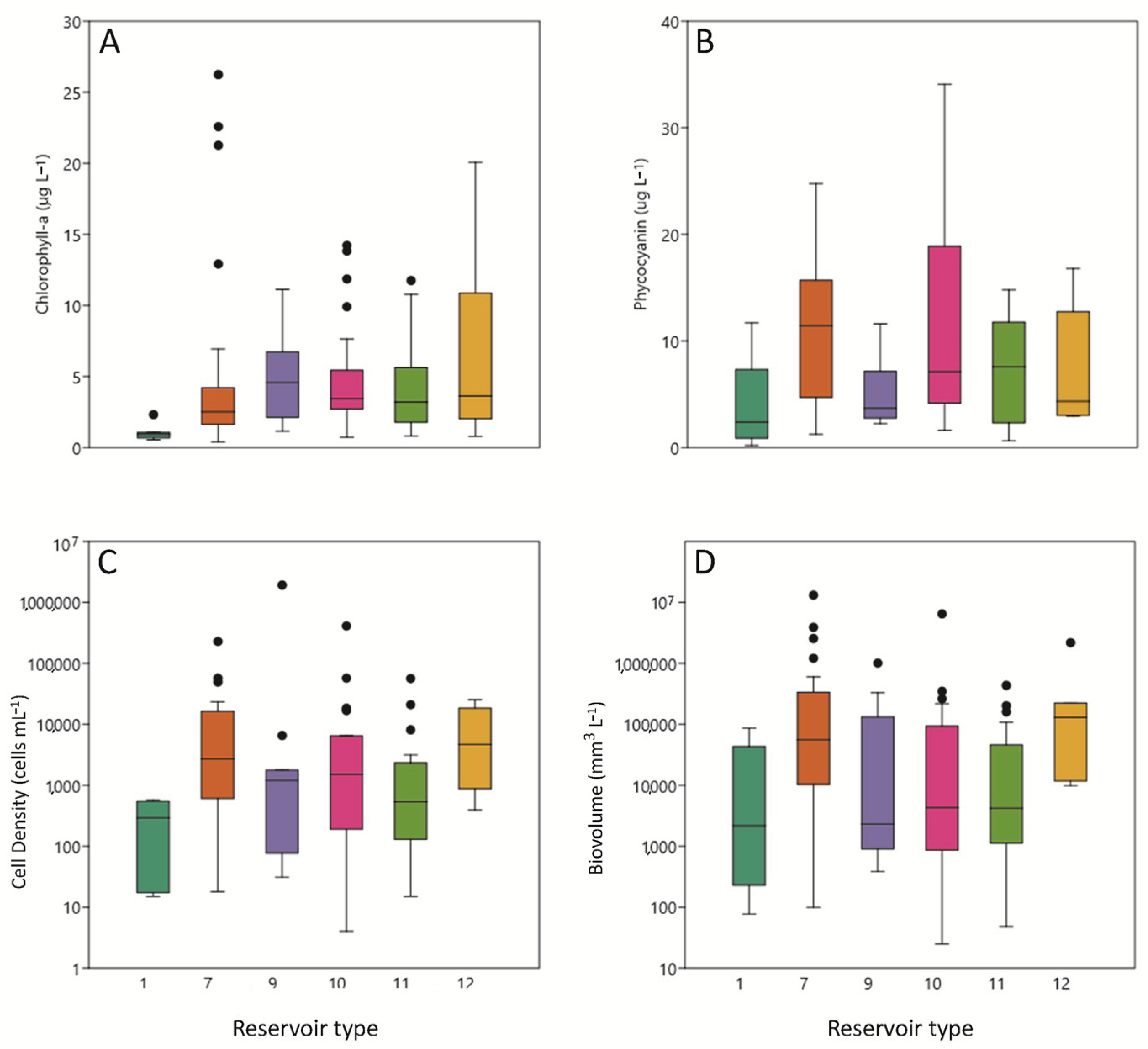
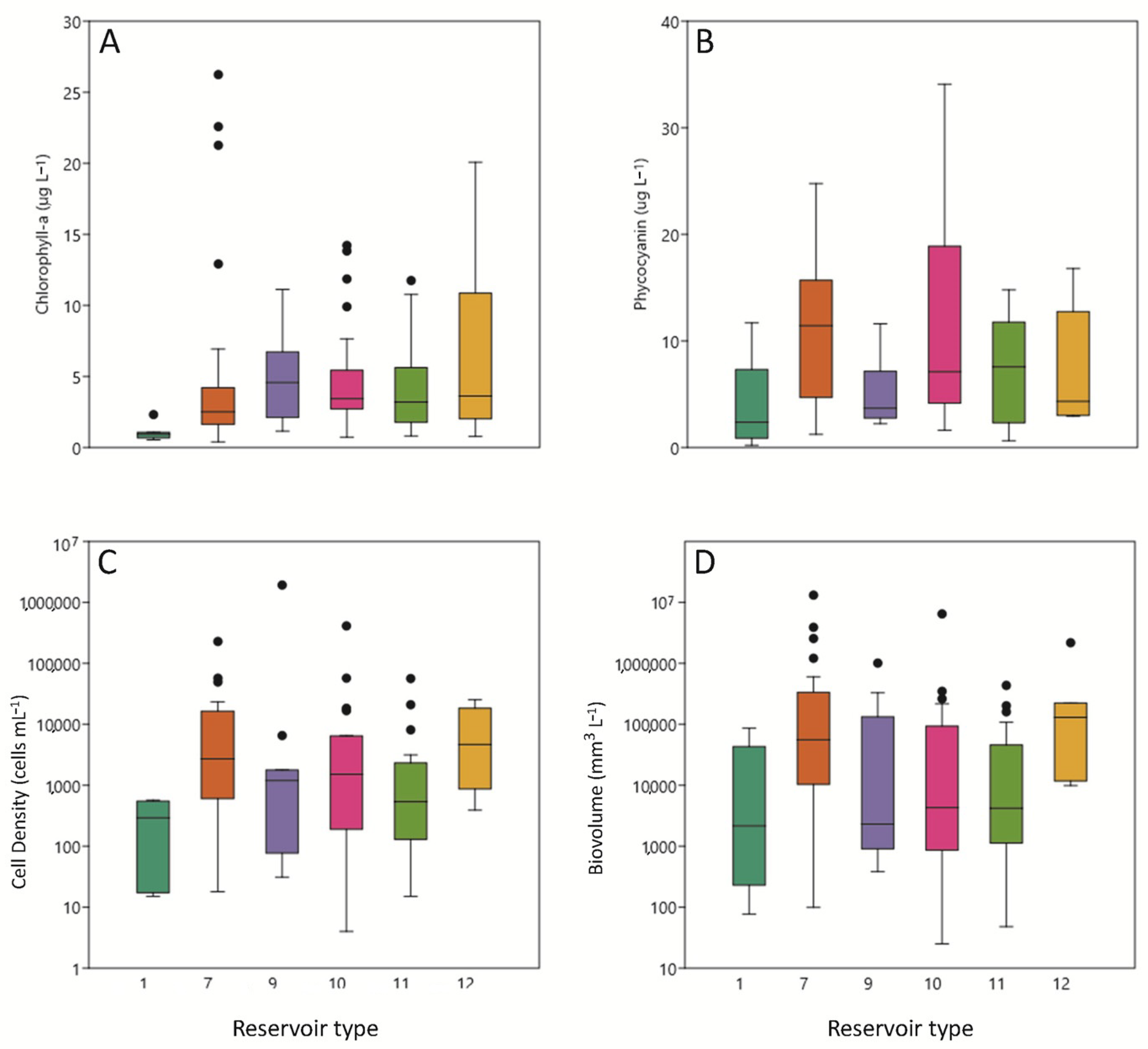
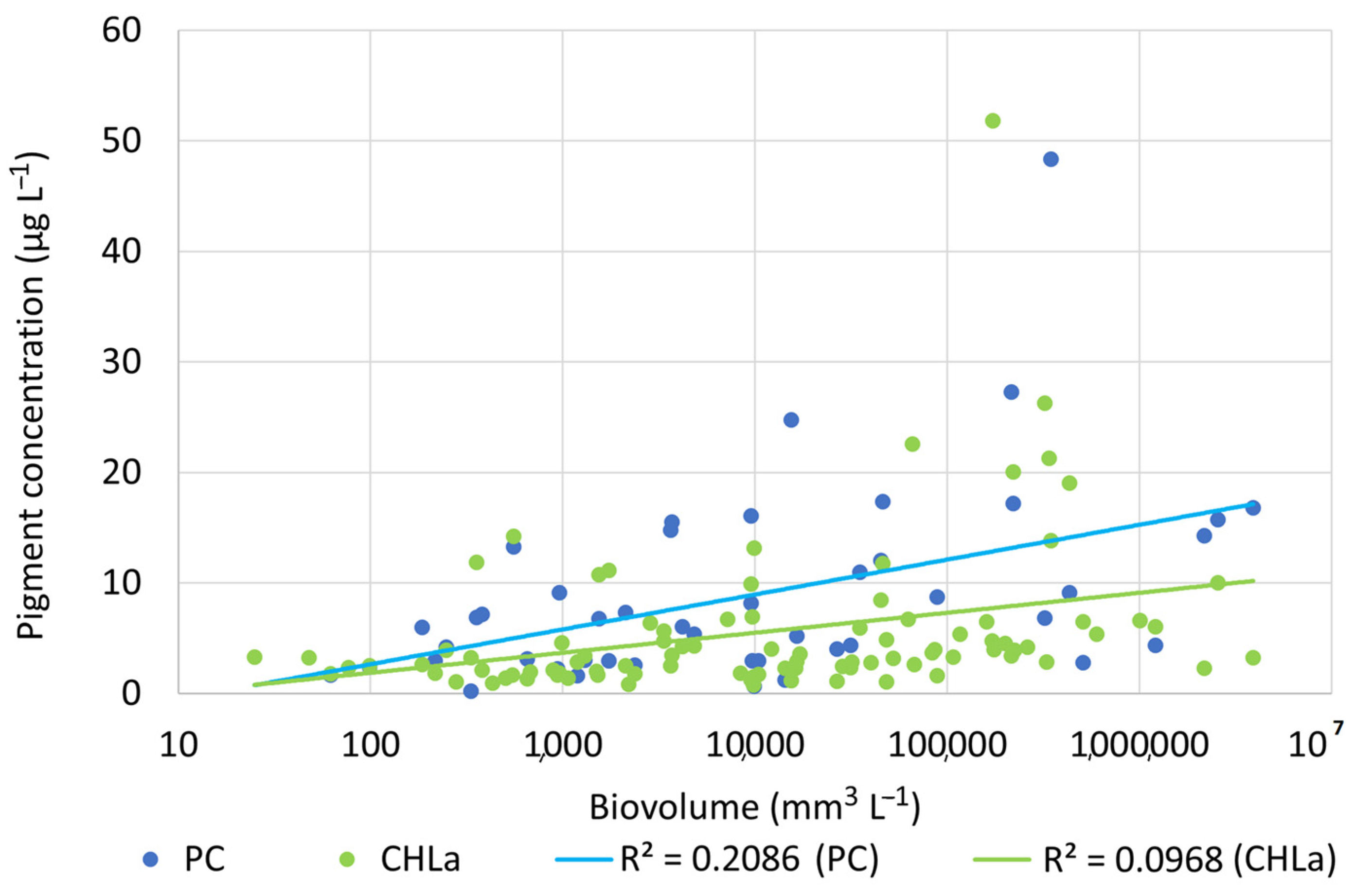
3.3. Phytoplankton Pigments
3.4. Phytoplankton Assemblages
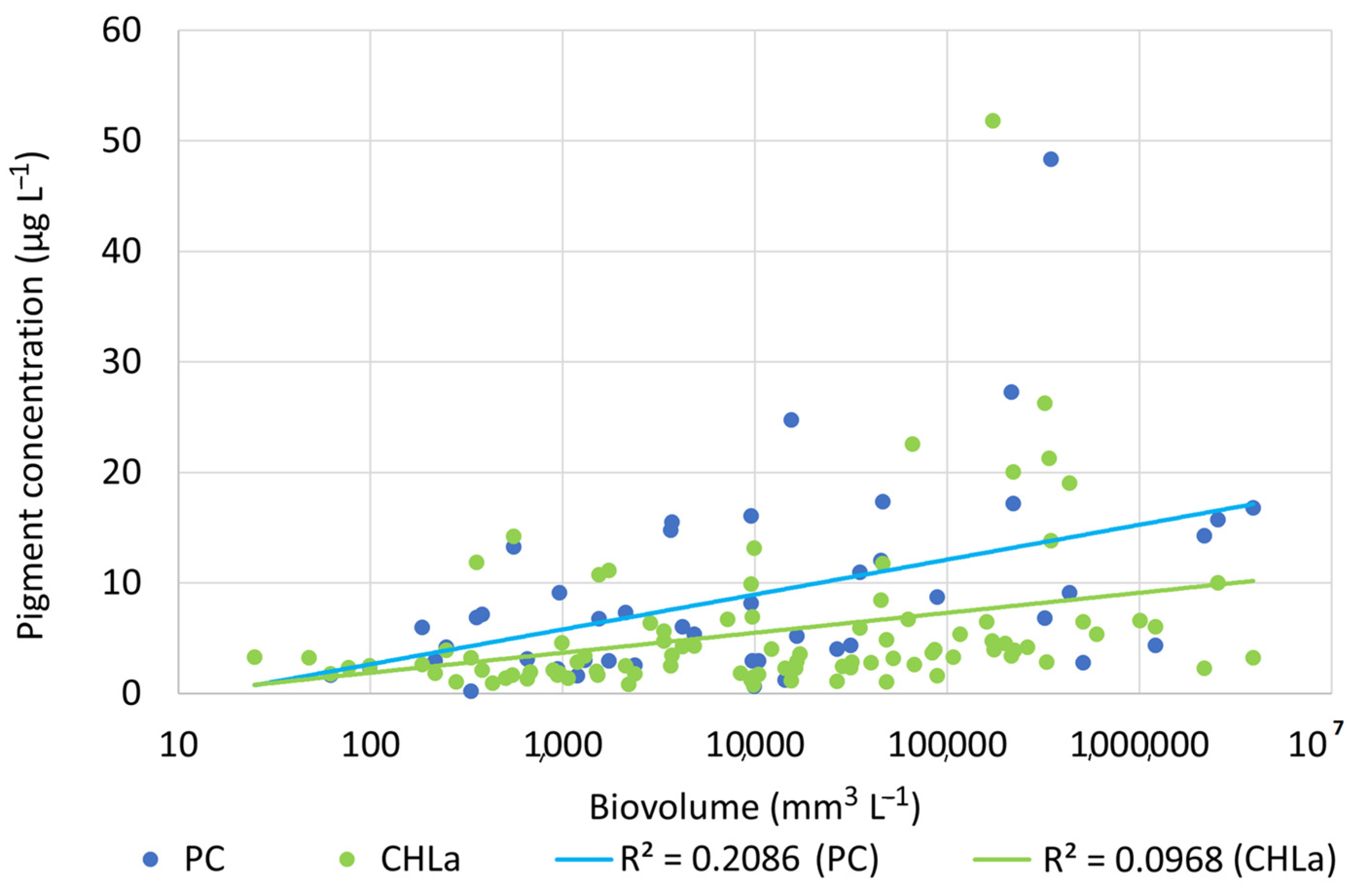
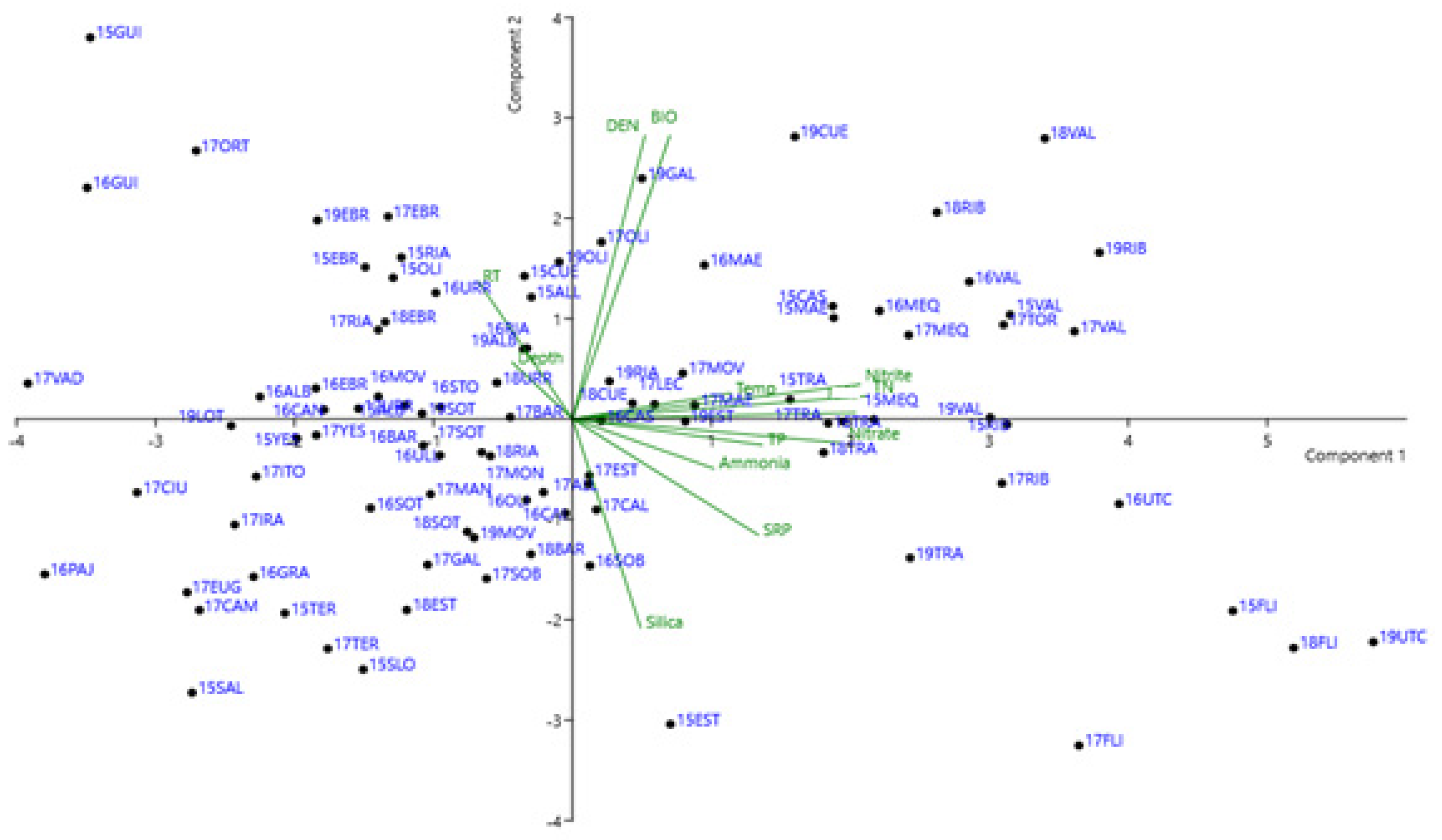
3.5. Pearson’s Correlation and PCA
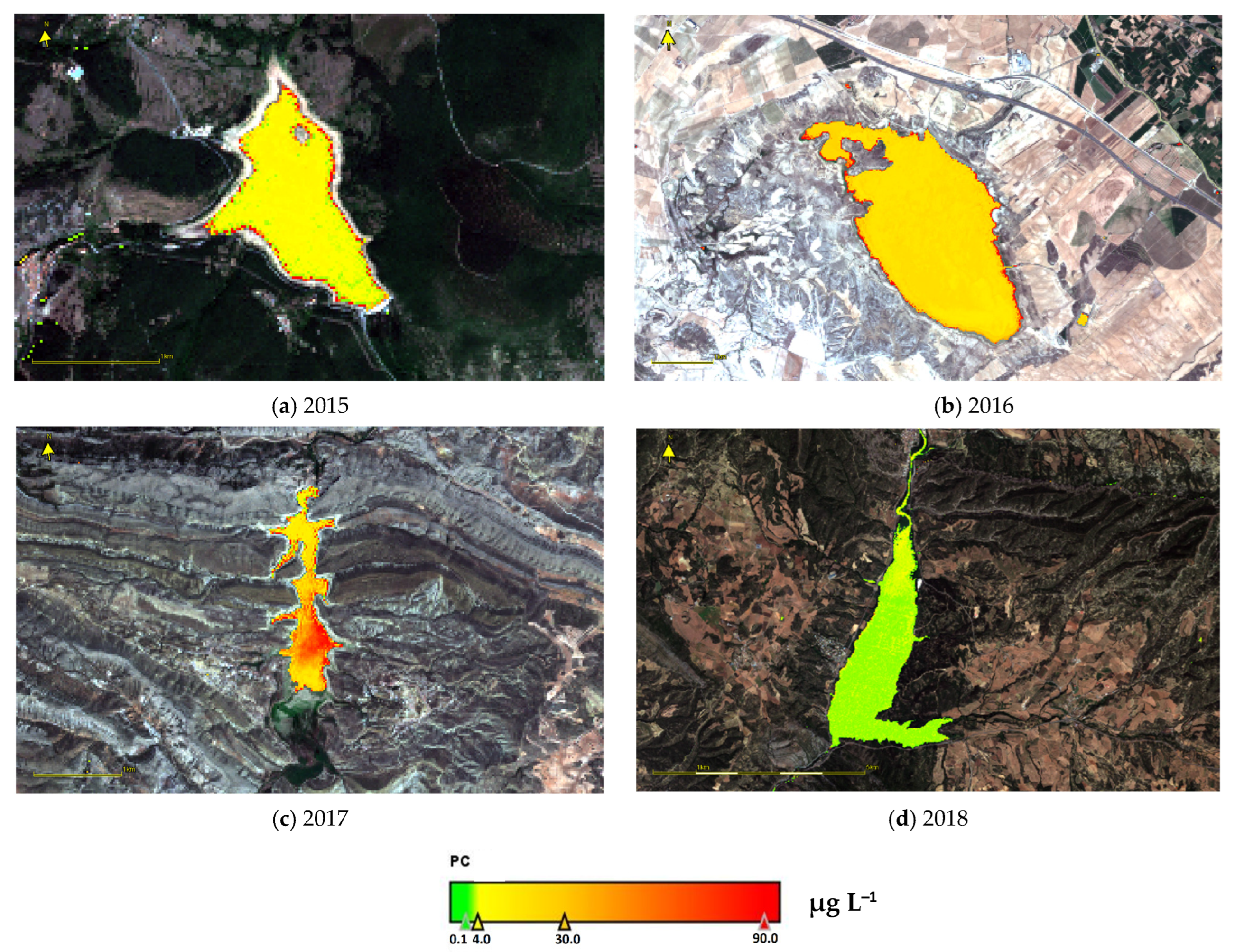
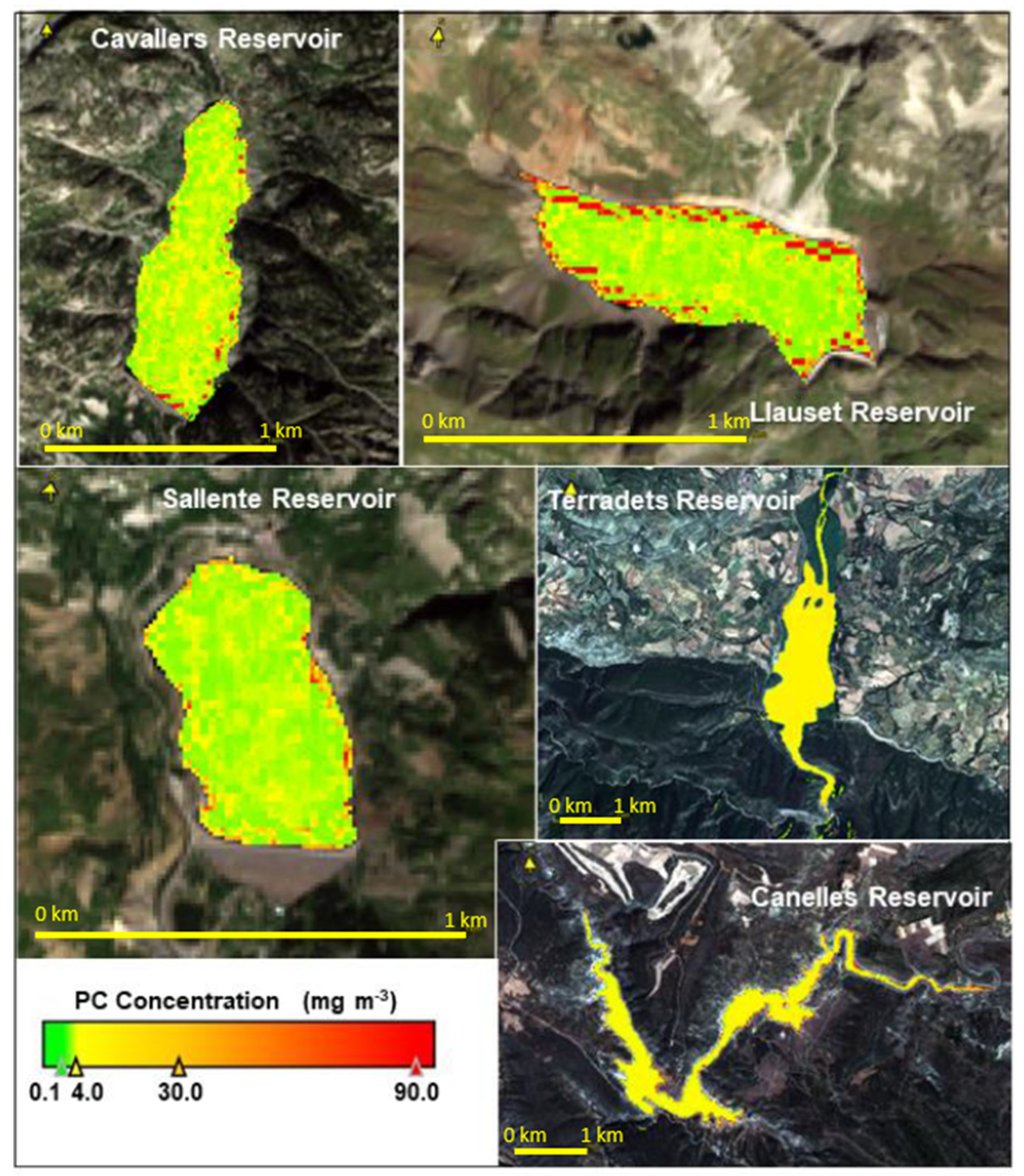
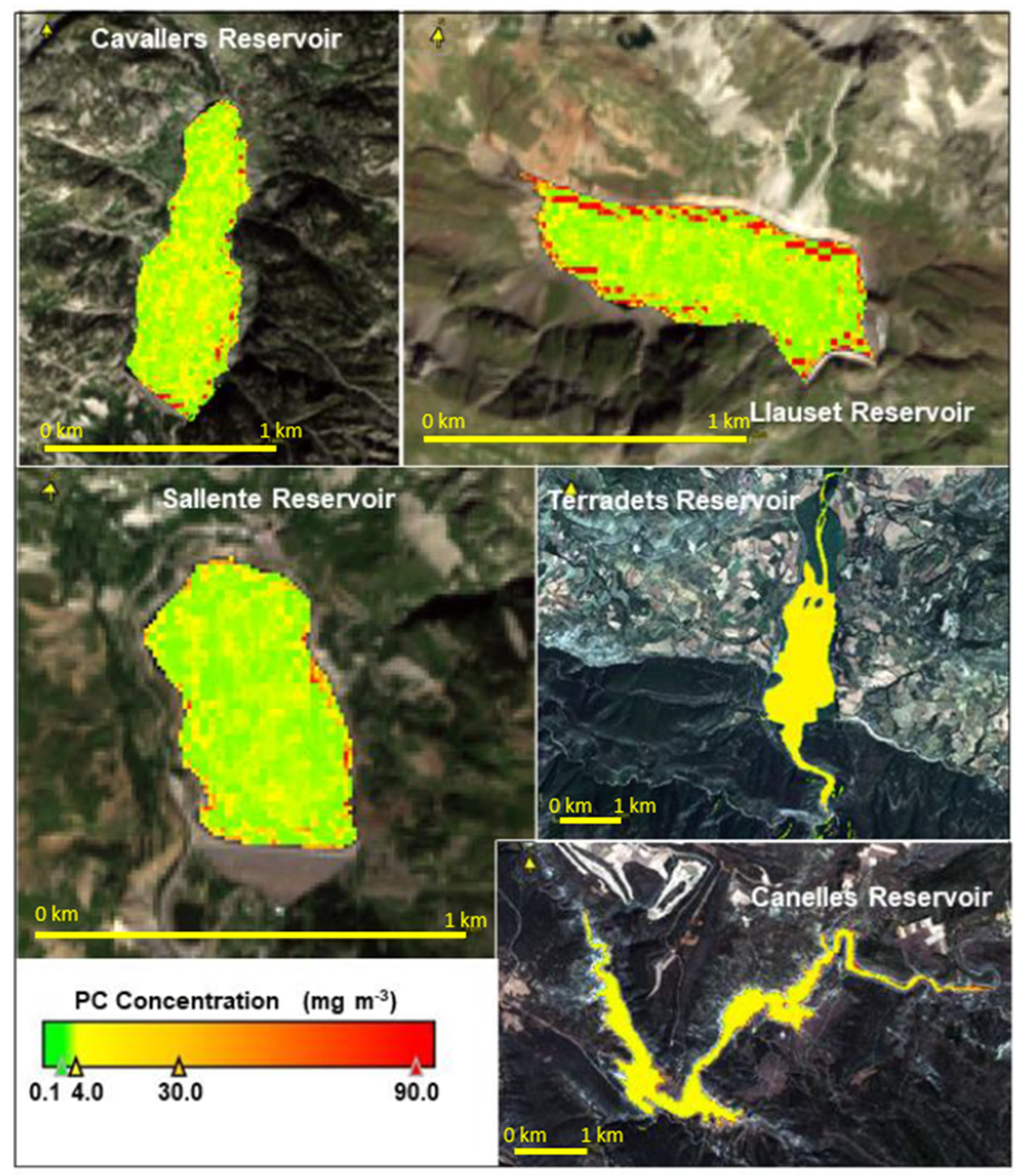
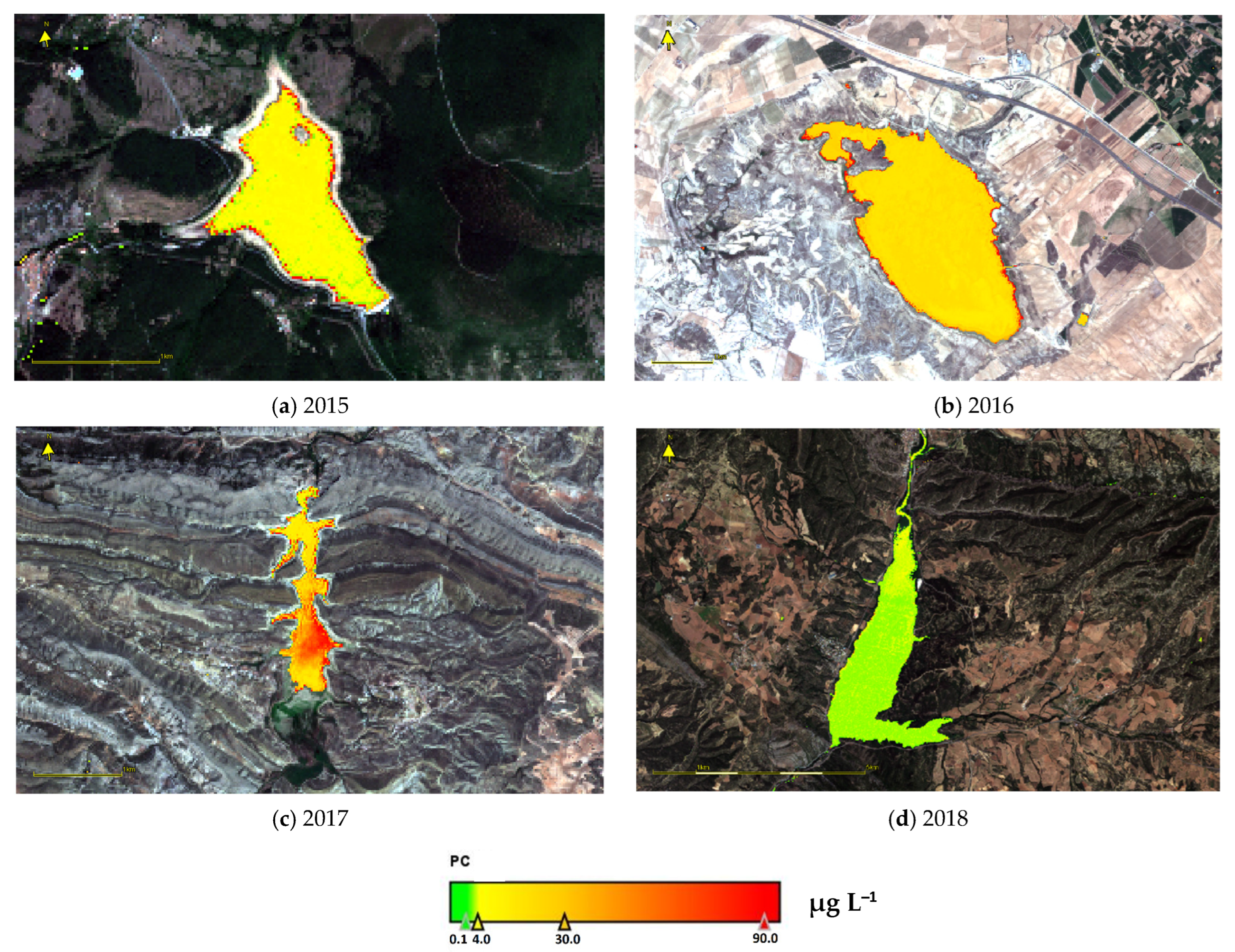
3.6. PC Values Estimated from Remote Sensing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reservoir and Year | Abbreviation | PC (μg L−1) In Situ | PC (μg L−1) Satellite | RMSE μg/L | Reservoir and Year | Abbreviation | PC (μg L−1) In Situ | PC (μg L−1) Satellite | RMSE μg/L |
|---|---|---|---|---|---|---|---|---|---|
| Canelles 2016 | 16CAN | 1.63 | 3.86 | 1.57 | Ebro 2018 | 18EBR | 14.79 | - | - |
| La Sotonera 2016 | 16SOT | 3.00 | 5.47 | 1.74 | Lechago 2018 | 18LEC | 4.31 | 5.22 | 17.43 |
| La Tranquera 2016 | 16TRA | 10.94 | 5 | 4.20 | Monteagudo 2018 | 18MON | 8.70 | 4.39 | 3.05 |
| Mansilla 2016 | 16MAN | 1.54 | 6.65 | 3.61 | Urrunaga 2018 | 18URR | 16.79 | 4.8 | 9.47 |
| Santolea 2016 | 16STO | 4.01 | 4.01 | 0 | El Val 2018 | 18VAL | 11.70 | - | - |
| Ullibari-Gamboa 2016 | 16ULL | 1.24 | - | - | Oliana 2018 | 18OLI | 9.05 | 4.11 | 3.49 |
| Sobrón 2016 | 16SOB | 2.95 | 3.85 | 0.63 | Sobrón 2018 | 18SOB | 15.69 | 4.4 | 8.98 |
| Alloz 2017 | 17ALL | 3.11 | 4.38 | 0.89 | Terradets 2018 | 18TER | 24.77 | 10.41 | 12.15 |
| Ebro 2017 | 17 EBR | 4.34 | - | - | Cueva Foradada 2018 | 18CUE | 13.24 | 12.26 | 0.69 |
| Eugui 2017 | 17EUG | 2.93 | 3.66 | 0.52 | Mezalocha 2018 | 18MEZ | 5.96 | 8.33 | 1.67 |
| Irabia 2017 | 17IRA | 0.21 | - | - | La Sotonera 2018 | 18SOT | 15.52 | 5.65 | 6.98 |
| Itoiz 2017 | 17ITO | 2.37 | 4.61 | 1.58 | Barasona 2018 | 18BAR | 2.57 | 4.13 | 1.11 |
| Maidevera 2017 | 17MAE | 2.94 | - | - | Rialb 2018 | 18RIA | 5.19 | 4.4 | 0.56 |
| El Val 2017 | 17VAL | 6.80 | - | - | La Tranquera 2018 | 18TRA | 17.35 | 4 | 9.44 |
| Oliana 2017 | 17OLI | 15.71 | 5.32 | 9.35 | Flix 2018 | 18FLI | 14.29 | 7.64 | 7.74 |
| La Peña 2017 | 17PEÑ | 4.21 | 5.02 | 0.57 | Ribarroja 2018 | 18RIB | 17.19 | 8.75 | 8.86 |
| Terradets 2017 | 17TER | 7.15 | 14.94 | 8.51 | Ebro 2019 | 19EBR | 4.38 | 2.86 | 1.07 |
| Yesa 2017 | 17YES | 2.23 | 3.87 | 1.16 | Oliana 2019 | 19OLI | 2.76 | 2.86 | 0.07 |
| Cueva Foradada 2017 | 17CUE | 3.70 | 12.05 | 9.52 | Sobron 2019 | 19SOB | 11.61 | 3.85 | 8.48 |
| Gallipuén 2017 | 17GAL | 4.21 | - | - | Estanca de Alcañiz 2019 | 19EST | 8.18 | - | - |
| Mezalocha 2017 | 17MEZ | 34.08 | 19.77 | 10.13 | Gallipuen 2019 | 19GAL | 27.28 | 17.17 | 7.15 |
| Moneva 2017 | 17MOV | 48.34 | - | - | La Loteta 2019 | 19LOT | 9.11 | 9.98 | 0.62 |
| Las Torcas 2017 | 17TOR | 2.98 | 4.4 | 1.01 | Moneva 2019 | 19MOV | 6.89 | 28.1 | 14.99 |
| Camarasa 2017 | 17CAM | 1.66 | - | - | La Sotonera 2019 | 19SOT | 7.33 | 5.65 | 1.19 |
| Rialb 2017 | 17RIA | - | - | - | Utchesa-Seca 2019 | 19UTC | 16.09 | 2.83 | 9.37 |
| Type | Reservoirs | Mixing Regime | Geology | Humidity Index (HI) | Basin Area | Annual Temperature |
|---|---|---|---|---|---|---|
| 1 | Lanuza, Pajares. | Monomictics | Siliceous (alkalinity < 1 meq/L) | HI > 0.74 Medium Humidity | Header and upper reaches (basin area < 1000 km2) | <15 °C |
| 7 | Albiña, Alloz, Búbal, Ebro, El Val, Escales, Escarra, Eugui, Irabia, Itoiz, Lechago, Maidevera, Mansilla, Monteagudo, Ortigosa, Sopeira, Ullívarri-Gamboa, Urdalur, Urrúnaga, Vadiello. | Calcareous Alkalinity > 1 meq/L) | HI > 0.74 Medium Humidity | Header and upper reaches (basin area < 1000 km2) | <15 °C | |
| 9 | La Peña, Mediano, Oliana, Sobrón, Terradets, Yesa. | (basin area > 1000 km2) | ||||
| 10 | Ciurana, Cueva Foradada, Gallipuen, Guiamets, La Estanca, La Loteta, La Sotonera, Las Torcas, Margalef, Mezalocha, Moneva, Pena, Utchesa-Seca | HI < 0.74 Low Humidity | Header and upper reaches (basin area < 1000 km2) | |||
| 11 | Balaguer, Barasona, Calanda, Camarasa, Canelles, Grado, La Tranquera, Rialb, San Lorenzo, Santolea, Talarn. | (basin area > 1000 and < 25,000 km2) | ||||
| 12 | Caspe, Flix, Mequinenza, Ribarroja. | Lower sections of the main axes (basin area > 25,000 km2) | ||||
| 13 | Cavallers, Llauset. | Dimictics | HI > 2 High Humidity |
Appendix B
References
- Burkholder, J.M.; Glibert, P.M. Eutrophication and oligotrophication. In Encyclopedia of Biodiversity, 3rd ed.; Levin, S., Ed.; Academic Press: New York, NY, USA, 2022. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to their Public Health Consequences, Monitoring and Management; E&FN Spon on behalf of the World Health, Organization; Chorus, I., Welker, M., Eds.; CRC Press: London, UK, 1999; pp. 1–416. [Google Scholar]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water—A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; World Health Organization (WHO): Geneva, Switzerland; CRC Press: Boca Raton, FL, USA, 2021.
- Cobo, F. Métodos de control de las floraciones de cianobacterias en aguas continentales. Limnetica 2015, 34, 247–268. [Google Scholar]
- Glibert, P.M. Harmful algae at the complex nexus of eutrophication and climate change. Harmful Algae 2020, 91, 101583. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnology, 3rd ed.; Academic Press: New York, NY, USA, 2001. [Google Scholar]
- Graham, L.E.; Graham, J.M.; Wilcox, L.W.; Cook, M.E. Algae, 3rd ed.; LJLM Press, LLC: Madison, WI, USA, 2016. [Google Scholar]
- Chaffin, J.D.; Bridgeman, T.B. Organic and inorganic nitrogen utilization by nitrogen-stressed cyanobacteria during bloom conditions. J. Appl. Phycol. 2014, 26, 299–309. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency (U.S. EPA). Preventing Eutrophication: Scientific Support for Dual Nutrient Criteria; Fact Sheet #EPA-820-S-15-001; Office of Water, U.S. EPA: Washington, DC, USA, 2015.
- Dolman, A.M.; Rucker, J.; Pick, F.R.; Fastner, J.; Rohlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and cyanotoxins: The influence of nitrogen versus phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Guidelines for Drinking-Water Quality, 4th ed.; World Health Organization: Geneva, Switzerland, 2011; p. 636.
- Ebro Basin Authority (Confederación Hidrográfica del Ebro). Available online: http://www.chebro.es/ (accessed on 8 June 2023).
- Water Framework Directive (2000/60/EC) Common Implementation Strategy; Guidance Document No. 2. Identification of Water Bodies; Office for Official Publications of the European Communities: Luxembourg, 2003.
- Wetzel, R.G.; Likens, G.E. Limnological Analyses, 3rd ed.; Springer: New York, NY, USA, 2000. [Google Scholar]
- Rueda, F.; Moreno-Ostos, E.; Armengol, J. The residence time of river water in reservoirs. Ecol. Model. 2006, 191, 260–274. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater, 18th ed.; APHA 4500-NO3 E/4599-NO2 B APHA 4500-N C APHA 4500-SIO2 C; APHA (American Public Health Association): Washington, DC, USA, 2005. [Google Scholar]
- Shoaf, W.T.; Lium, B.W. Improve extraction of chlorophyll a and b from algae using dimethyl sulfoxide. Limnol. Oceanogr. 1976, 21, 926–928. [Google Scholar] [CrossRef]
- Jeffrey, S.T.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Sobiechowska-Sasim, M.; Ston-Egiert, J.; Kosakowska, A. Quantitative analysis of extracted phycobilin pigments in cyanobacteria—An assessment of spectrophotometric and spectrofluorometric methods. J. Appl. Phycol. 2014, 26, 2065–2074. [Google Scholar] [CrossRef] [Green Version]
- Vollenweider, R.A. (Ed.) A Manual on Methods for Measuring Primary Productivity in Aquatic Environments, 2nd ed.; IBP Handbook No. 12; Blackwell Scientific: Oxford, UK, 1974. [Google Scholar]
- DIN EN 15204:2006-12; Water Quality—Guidance Standard on the Enumeration of Phytoplankton Using Inverted Microscopy (Utermöhl Technique). European Committee for Standardization: Brussels, Belgium, 2006.
- Soria, J.M.; Montagud, D.; Sòria-Perpinyà, X.; Dolores Sendra, M.; Vicente, E. Phytoplankton Reservoir Trophic Index (PRTI): A new tool for ecological quality studies. Inland Waters 2019, 9, 301–308. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; W.H. Freeman and Co.: New York, NY, USA, 2012. [Google Scholar]
- Jackson, D.A. Stopping rules in principal components analysis—A comparison of heuristic and statistical approaches. Ecology 1993, 74, 2204–2214. [Google Scholar] [CrossRef]
- Boyer, G.L. The occurrence of cyanobacterial toxins in New York lakes: Lessons from the MERHAB-Lower Great Lakes Program. Lake Reserv. Manag. 2007, 23, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Brient, L.; Lengronne, M.; Bertrand, E.; Rolland, D.; Sipel, A.; Steinmann, D.; Baudin, I.; Legeas, M.; Le Rouzic, B.; Bormans, M. A Phycocyanin probe as a tool for monitoring cyanobacteria in freshwater bodies. J. Environ. Monit. 2008, 10, 248–255. [Google Scholar] [CrossRef] [PubMed]
- European Space Agency. Sentinel-2 Spectral Response Functions. 2017. Available online: https://sentinel.esa.int/web/sentinel/user-guides/sentinel-2-msi/document-library/-/asset_publisher/Wk0TKajiISaR/content/sentinel-2a-spectralresponses (accessed on 12 April 2023).
- Radin, C.; Sòria-Perpinyà, X.; Delegido, J. Multitemporal water quality study in Sitjar (Castelló, Spain) reservoir using 674 Sentinel-2 images. Rev. Teledetec. 2020, 56, 117–130. [Google Scholar] [CrossRef]
- National Institute of Aerospace Technology (INTA). Available online: https://www.inta.es/INTA/en/ (accessed on 30 January 2023).
- Pérez-González, R.; Sòria-Perpinyà, X.; Soria, J.M.; Delegido, J.; Urrego, P.; Sendra, M.D.; Ruíz-Verdú, A.; Vicente, E.; Moreno, J. Phycocyanin monitoring in some Spanish water bodies with Sentinel-2 imagery. Water 2021, 13, 2866. [Google Scholar] [CrossRef]
- Burkholder, J.M.; Kinder, C.A.; Dickey, D.A.; Reed, R.E.; Arellano, C.; James, J.L.; Mackenzie, L.M.; Allen, E.H.; Lindor, N.L.; Mathis, J.G.; et al. Classical discrete indicators and diel DO vs. trend analysis in assessing eutrophication of potable-water reservoirs. Ecol. Appl. 2022, 32, e2541. [Google Scholar] [CrossRef]
- Jiang, Y.; Ji, B.; Wong, R.N.S.; Wong, M.H. Statistical study on the effects of environmental factors on the growth and microcystins production of bloom-forming cyanobacterium—Microcystis aeruginosa. Harmful Algae 2008, 7, 127–136. [Google Scholar] [CrossRef]
- Paerl, H.W.; Havens, K.E.; Hall, N.S.; Otten, T.G.; Zhu, M.; Xu, H.; Qin, B. Mitigating a global expansion of toxic cy-anobacterial blooms: Confounding effects and challenges posed by climate change. Mar. Freshw. Res. 2019, 715, 579–592. [Google Scholar]
- Yan, X.; Xu, X.; Wang, M.; Wang, G.; Wu, S.; Li, Z.; Yang, Y. Climate warming and cyanobacteria blooms: Looks at their relationships from a new perspective. Water Res. 2017, 125, 449–457. [Google Scholar] [CrossRef]
- Mantzouki, E.; Lürling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Woźniak, E.; Koreivienė, J.; Warming, T.P. Temperature effects explain continental scale distribution of cyanobacterial toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Peng, C.; Wang, J.; Huang, S.; Hu, Y.; Zhang, J.; Li, D. Temperature and silicate are significant driving factors for the seasonal shift of dominant diatoms in a drinking water reservoir. J. Oceanol. Limnol. 2019, 37, 568–579. [Google Scholar] [CrossRef]
- Catherine, Q.; Susanna, W.; Isidora, E.S.; Mark, H.; Aurelie, V.; Jean-François, H. A review of current knowledge on toxic benthic freshwater cyanobacteria–ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef] [PubMed]
- Sòria-Perpinyà, X.; Vicente, E.; Urrego, P.; Pereira-Sandoval, M.; Ruíz-Verdú, A.; Delegido, J.; Moreno, J. Remote sensing of cyanobacterial blooms in a hypertrophic lagoon (Albufera of València, Eastern Iberian Peninsula) using multitemporal Sentinel-2 images. Sci. Total Environ. 2020, 698, 134305. [Google Scholar] [CrossRef] [PubMed]
- Hilborn, E.D.; Beasley, V.R. One health and cyanobacteria in freshwater systems: Animal illnesses and deaths are sentinel events for human health risks. Toxins 2015, 7, 1374–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]



| Name | Position | Depth m (max) | Volume ×106 m3 | Elevation m.a.s.l | Res. Time (Years) | Climate | |
|---|---|---|---|---|---|---|---|
| Lat. | Lon. | ||||||
| Alloz | 42.70 | −1.92 | 60 | 65 | 468 | 0.48 | Cfa |
| Barasona | 42.14 | 0.33 | 66 | 85 | 448 | 0.24 | Cfa |
| Canelles | 42.03 | 0.65 | 150 | 201 | 506 | 0.00 | Cfb |
| C. Foradada | 40.97 | −0.69 | 65 | 22 | 580 | 0.65 | Bsk |
| Ebro | 42.97 | −4.07 | 34 | 540 | 838 | 1.55 | Cfb |
| Est. Alcañiz | 41.06 | −0.18 | 15 | 7 | 342 | 0.14 | BSk |
| Eugui | 42.97 | −1.51 | 43 | 21 | 628 | 0.18 | Cfb |
| Flix | 41.23 | 0.53 | 26 | 11 | 41 | 0.01 | BSk |
| Gallipuén | 40.87 | −0.41 | 36 | 4 | 694 | 0.71 | Cfb |
| Itoiz | 42.48 | −1.21 | 107 | 418 | 573 | 0.57 | Cfb |
| Lechago | 40.96 | −1.30 | 18.5 | 7 | 891 | - | Csa |
| Loteta | 41.82 | −1.32 | 34 | 100 | 288 | 3.51 | BSk |
| Mansilla | 42.16 | −2.91 | 70 | 68 | 930 | 0.09 | Cfa |
| Mezalocha | 41.42 | −1.07 | 45 | 4 | 473 | 1.17 | Cfa |
| Mequinenza | 41.22 | 16.33 | 79 | 1534 | 106 | 0.13 | Csa |
| Moneva | 41.17 | −0.83 | 45 | 8 | 615 | 0.95 | Cfb |
| Oliana | 42.12 | 1.3 | 102 | 84 | 519 | 0.08 | Cfa |
| Peña | 40.82 | 0.13 | 61 | 18 | 561 | - | Bsk |
| Rialb | 41.97 | 1.23 | 99 | 402 | 430 | 0.36 | Cfa |
| Ribarroja | 41.33 | 0.36 | 60 | 207 | 70 | 0.03 | Csb |
| Santolea | 40.77 | −0.31 | 44 | 48 | 596 | 0.60 | Csa |
| Sobrón | 42.76 | −3.15 | 39 | 20 | 511 | 0.06 | Cfb |
| Sotonera | 42.11 | −0.68 | 31 | 189 | 417 | 0.58 | Cfa |
| Terradets | 42.05 | 0.88 | 47 | 33 | 372 | 0.04 | Cfa |
| Torcas | 41.29 | −1.08 | 41 | 7 | 624 | 0.27 | Cfa |
| Tranquera | 41.24 | −1.78 | 81 | 84 | 684 | 0.68 | BSk |
| Urrúnaga | 42.98 | −2.65 | 31 | 72 | 547 | 0.31 | Csb |
| Utchesa | 41.51 | 0.52 | 16.6 | 4 | 147 | - | Cfb |
| El Val | 42.61 | −1.78 | 50 | 24 | 620 | 0.42 | Cfa |
| Yesa | 42.61 | −1.18 | 60.7 | 447 | 488 | 0.23 | Cfb |
| Drinking Water | Bath Water | Density (cel./mL) | Biovolume (mm3/L) | Chlorophyll a (µg/L) | PC (µg/L) |
|---|---|---|---|---|---|
| Surveillance level | 200 | 0.02 | 0.1 | <0.1 | |
| Alert level I | 2000 | 0.2 | 1.0 | 4 | |
| Guidance level I | 20,000 | 2 | 10 | 30 ± 2 | |
| Alert level II | Guidance level II | 100,000 | 10 | 50 | 90 ± 2 |
| Depth | 1 | |||||||||||
| Temperature | −0.268 | 1 | ||||||||||
| pH | 0.001 | 0.280 | 1 | |||||||||
| Nitrate | −0.075 | 0.328 | −0.034 | 1 | ||||||||
| Ammonia | −0.131 | 0.178 | −0.016 | 0.314 | 1 | |||||||
| Total N | −0.169 | 0.451 | 0.078 | 0.887 | 0.396 | 1 | ||||||
| SRP | −0.161 | 0.148 | −0.033 | 0.304 | 0.313 | 0.346 | 1 | |||||
| Total P | −0.378 | 0.284 | 0.168 | 0.255 | 0.424 | 0.435 | 0.515 | 1 | ||||
| Silica | −0.095 | −0.001 | −0.173 | 0.140 | 0.261 | 0.147 | 0.235 | 0.220 | 1 | |||
| RT | 0.035 | 0.072 | −0.012 | −0.208 | 0.045 | 0.001 | −0.149 | −0.119 | −0.068 | 1 | ||
| Biovolume | −0.237 | 0.129 | 0.164 | 0.063 | 0.080 | 0.242 | 0.060 | 0.303 | −0.249 | 0.119 | 1 | |
| Cell number | −0.237 | 0.481 | 0.236 | 0.149 | 0.047 | 0.265 | 0.087 | 0.204 | −0.295 | 0.059 | 0.741 | 1 |
| Depth | Temp. | pH | Nitrate | Ammonia | Total N | SRP | Total P | Silica | RT | Biovolume | Cell number |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-González, R.; Sòria-Perpinyà, X.; Soria, J.; Sendra, M.D.; Vicente, E. Relationship between Cyanobacterial Abundance and Physicochemical Variables in the Ebro Basin Reservoirs (Spain). Water 2023, 15, 2538. https://doi.org/10.3390/w15142538
Pérez-González R, Sòria-Perpinyà X, Soria J, Sendra MD, Vicente E. Relationship between Cyanobacterial Abundance and Physicochemical Variables in the Ebro Basin Reservoirs (Spain). Water. 2023; 15(14):2538. https://doi.org/10.3390/w15142538
Chicago/Turabian StylePérez-González, Rebeca, Xavier Sòria-Perpinyà, Juan Soria, Maria D. Sendra, and Eduardo Vicente. 2023. "Relationship between Cyanobacterial Abundance and Physicochemical Variables in the Ebro Basin Reservoirs (Spain)" Water 15, no. 14: 2538. https://doi.org/10.3390/w15142538