
Environmental DNA Captures Variations in Fish Assemblages with Distance from Dams in Karst Reservoirs

 ,
,  ,
,
Abstract
1. Introduction
2. Methods
2.1. Sampling Design
2.2. Collection and Processing of eDNA Samples
2.3. Extraction, Amplification, and Sequencing
2.4. Bioinformatics Analysis
2.5. Statistical Analysis
3. Results
3.1. Sequencing and OTU Clustering Results
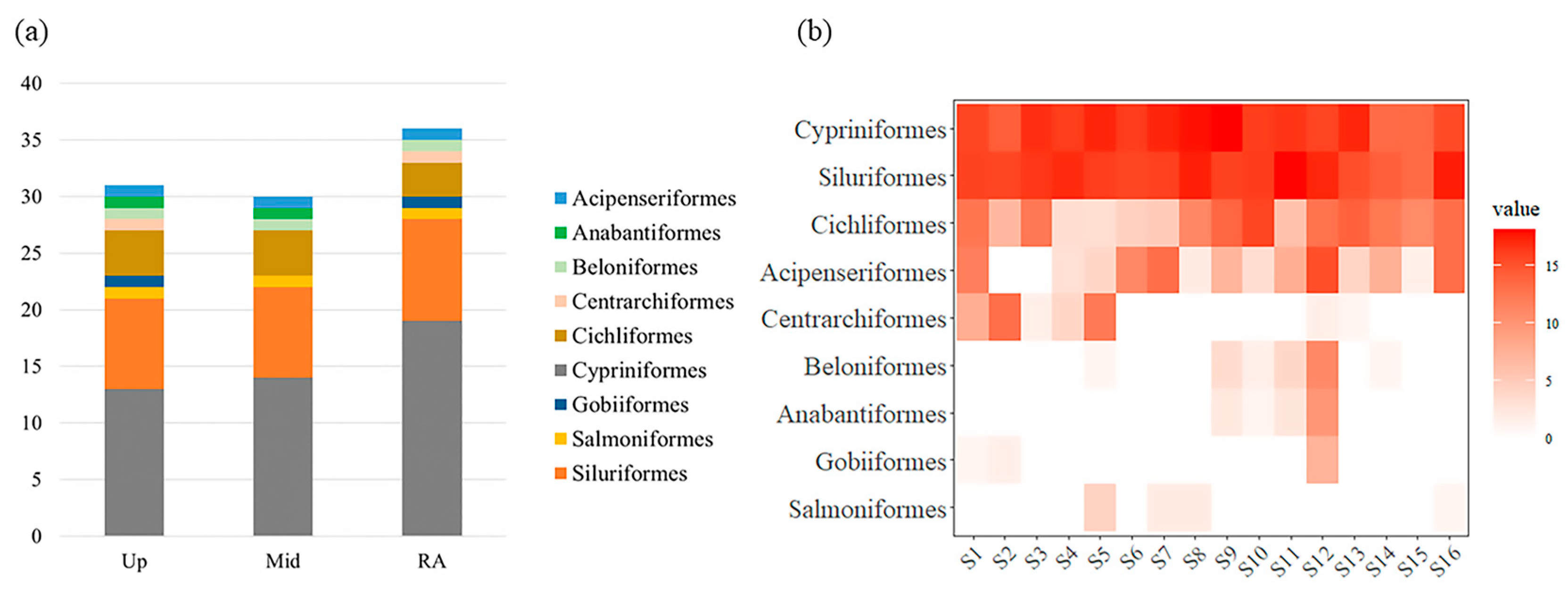
3.2. Fish Species Identified
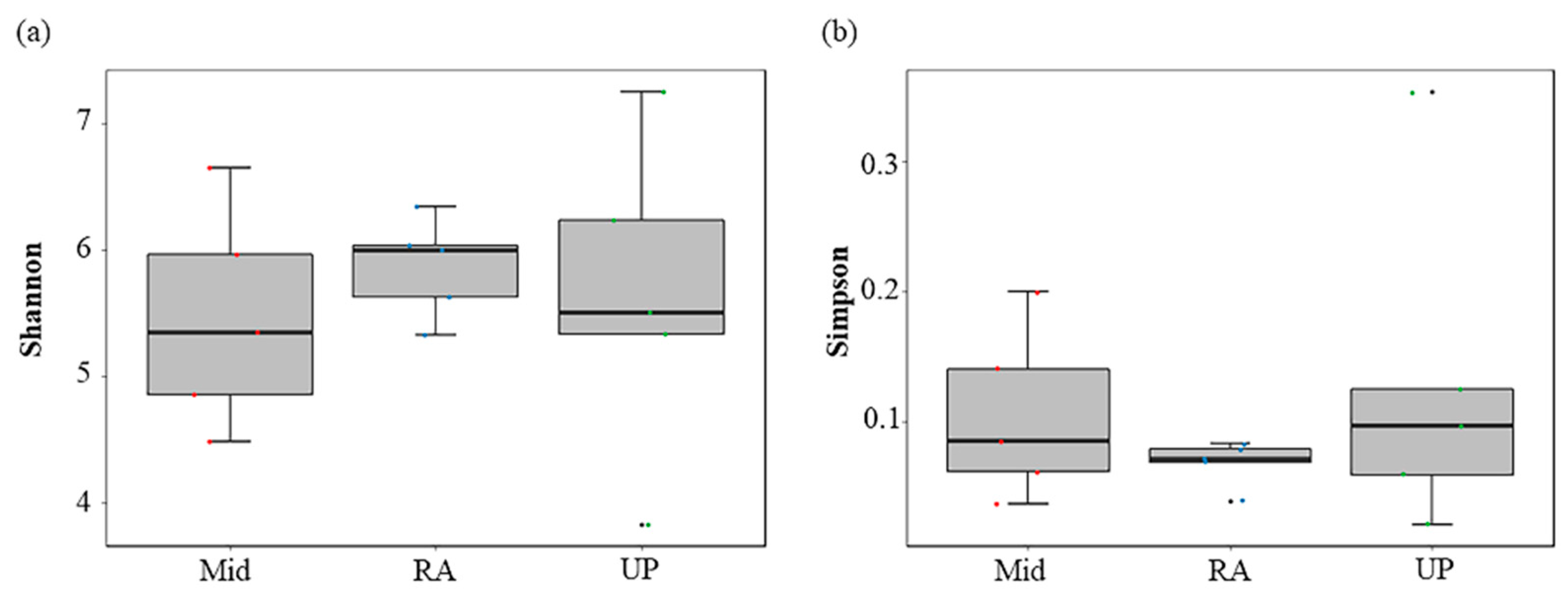
3.3. Spatial Variations of Fish Assemblages
3.4. The Relationship between Fish Diversity and Environmental Factors
4. Discussion
4.1. Status of Fish Assemblages in the Wanfeng Reservoir
4.2. Correlations of Fish Assemblages and Environmental Factors
4.3. Challenges of Karst Plateau Fish Conservation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R960–R967. [Google Scholar] [CrossRef] [PubMed]
- Rolls, R.J.; Hayden, B.; Kahilainen, K.K. Conceptualising the interactive effects of climate change and biological invasions on subarctic freshwater fish. Ecol. Evol. 2017, 7, 4109–4128. [Google Scholar] [CrossRef] [PubMed]
- Azzurro, E.; Sbragaglia, V.; Cerri, J.; Bariche, M.; Bolognini, L.; Souissi, J.B.; Busoni, G.; Coco, S.; Chryssanthi, A.; Fanelli, E.; et al. Climate change, biological invasions, and the shifting distribution of Mediterranean fishes: A large-scale survey based on local ecological knowledge. Glob. Chang. Biol. 2019, 25, 2779–2792. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.J.; Duarte, G.; Segurado, P.; Branco, P. Major threats to European freshwater fish species. Sci. Total Environ. 2021, 797, 149105. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.R.; Lockwood, J.L. Extinction in a field of bullets: A search for causes in the decline of the world’s freshwater fishes. Biol. Conserv. 2001, 102, 97–105. [Google Scholar] [CrossRef]
- Wang, B.; Huang, Y.; Li, J.; Dai, Q.; Wang, Y.; Yang, D. Amphibian species richness patterns in karst regions in Southwest China and its environmental associations. Biodivers. Sci. 2018, 26, 941–950. (In Chinese) [Google Scholar] [CrossRef]
- Li, L.; Li, Q.H.; Chen, J.A.; Wang, J.F.; Jiao, S.L.; Chen, F.F. Temporal and spatial distribution of phytoplankton functional groups and role of environment factors in a deep subtropical reservoir. J. Oceanol. Limnol. 2018, 36, 762–771. [Google Scholar] [CrossRef]
- Dos Santos, J.A.; Silva, C.B.; de Santana, H.S.; Cano-Barbacil, C.; Agostinho, A.A.; Normando, F.T.; Cabeza, J.R.; Roland, F.; García-Berthou, E. Assessing the short-term response of fish assemblages to damming of an Amazonian river. J. Environ. Manag. 2022, 307, 114571. [Google Scholar] [CrossRef]
- Piria, M.; Simonović, P.; Zanella, D.; Ćaleta, M.; Šprem, N.; Paunović, M.; Tomljanović, T.; Gavrilović, A.; Pecina, M.; Špelić, I.; et al. Long-term analysis of fish assemblage structure in the middle section of the Sava River—The impact of pollution, flood protection and dam construction. Sci. Total Environ. 2019, 651, 143–153. [Google Scholar] [CrossRef]
- Gao, X.; Fujiwara, M.; Winemiller, K.O.; Lin, P.; Li, M.; Liu, H. Regime shift in fish assemblage structure in the Yangtze River following construction of the Three Gorges Dam. Sci. Rep. 2019, 9, 4212. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L.; Tang, W.; Heino, J.; Jiang, X. Effects of dam construction and fish invasion on the species, functional and phylogenetic diversity of fish assemblages in the Yellow River Basin. J. Environ. Manag. 2021, 293, 112863. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, M.T.; Piana, P.A.; Baumgartner, G.; Gomes, L.C. Storage or run-of-river reservoirs: Exploring the ecological effects of dam operation on stability and species interactions of fish assemblages. Environ. Manag. 2020, 65, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.R.; Infante, D.M.; Daniel, W.M.; Wehrly, K.E.; Wang, L.; Brenden, T.O. Assessment of dam effects on streams and fish assemblages of the conterminous USA. Sci. Total Environ. 2017, 586, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Wang, J.; Ye, Y.S.; Li, W.; Correa, S.B.; Zhang, T.; Liu, J. Fish assemblage response to removal of a low-head dam in the lower reach of a tropical island river. Freshw. Biol. 2022, 67, 926–937. [Google Scholar] [CrossRef]
- Wu, Z.; Zhu, S.; Li, Y.; Xia, Y.; Zhang, Y.; Li, X.; Li, J. Spatiotemporal Response of Fish Aggregations to Hydrological Changes in the Lower Pearl River, China, during the Main Spawning Season. Water 2022, 14, 1723. [Google Scholar] [CrossRef]
- Shu, L.; Ludwig, A.; Peng, Z. Standards for Methods Utilizing Environmental DNA for Detection of Fish Species. Genes 2020, 11, 296. [Google Scholar] [CrossRef]
- Pont, G.D.; Ritter, C.D.; Agostinis, A.O.; Stica, P.V.; Horodesky, A.; Cozer, N.; Balsanelli, E.; Netto, O.S.M.; Henn, C.; Ostrensky, A.; et al. Monitoring fish communities through environmental DNA metabarcoding in the fish pass system of the second largest hydropower plant in the world. Sci. Rep. 2021, 11, 23167. [Google Scholar] [CrossRef]
- Ahn, H.; Kume, M.; Terashima, Y.; Ye, F.; Kameyama, S.; Miya, M.; Yamashita, Y.; Kasai, A. Evaluation of fish biodiversity in estuaries using environmental DNA metabarcoding. PLoS ONE 2020, 15, e0231127. [Google Scholar] [CrossRef]
- Bohmann, K.; Evans, A.; Gilbert, M.T.P.; Carvalho, G.R.; Creer, S.; Knapp, M.; Yu, D.W.; Bruyn, M.D. Environmental DNA for wildlife biology and biodiversity monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Miaud, C.; Pompanon, F.; Taberlet, P. Species detection using environmental DNA from water samples. Biol. Lett. 2008, 4, 423–425. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, Y.; Zhan, A.; Dong, C.; Zhao, J.; Yao, M. Environmental DNA captures native and non-native fish community variations across the lentic and lotic systems of a megacity. Sci. Adv. 2022, 8, eabk0097. [Google Scholar] [CrossRef] [PubMed]
- Consuegra, S.; O’Rorke, R.; Rodriguez-Barreto, D.; Fernandez, S.; Jones, J.; Leaniz, C.G.D. Impacts of large and small barriers on fish assemblage composition assessed using environmental DNA metabarcoding. Sci. Total Environ. 2021, 790, 148054. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Zhang, H.; Xian, W. Fish Diversity Monitored by Environmental DNA in the Yangtze River Mainstream. Fishes. 2022, 7, 1. [Google Scholar] [CrossRef]
- Li, H.; Yang, F.; Zhang, R.; Liu, S.; Yang, Z.; Lin, L.; Ye, S. Environmental DNA metabarcoding of fish communities in a small hydropower dam reservoir: A comparison between the eDNA approach and established fishing methods. J. Freshw. Ecol. 2022, 37, 337–358. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, Q.; Wang, Y.; Wang, X.; Zhao, J.; Yao, M. Assessment of fish communities using environmental DNA: Effect of spatial sampling design in lentic systems of different sizes. Mol. Ecol. Resour. 2020, 20, 242–255. [Google Scholar] [CrossRef]
- Yamamoto, S.; Minami, K.; Fukaya, K.; Takahashi, K.; Sawada, H.; Murakami, H.; Tsuji, S.; Hashizume, H.; Kubonaga, S.; Horiuchi, T.; et al. Environmental DNA as a ‘Snapshot’ of Fish Distribution: A Case Study of Japanese Jack Mackerel in Maizuru Bay, Sea of Japan. PLoS ONE 2016, 11, e0149786. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, J.D.; Yao, M. A comprehensive and comparative evaluation of primers for metabarcoding eDNA from fish. Methods Ecol. Evol. 2020, 11, 1609–1625. [Google Scholar] [CrossRef]
- Valdez-Moreno, M.; Ivanova, N.V.; Elías-Gutiérrez, M.; Pedersen, S.L.; Bessonov, K.; Bessonov, K. Using eDNA to biomonitor the fish community in a tropical oligotrophic lake. PLoS ONE 2019, 14, e0215505. [Google Scholar] [CrossRef]
- Antognazza, C.M.; Britton, J.R.; Potter, C.; Franklin, E.; Hardouin, E.A.; Roberts, C.G.; Aprahamian, M.; Andreou, D. Environmental DNA as a non-invasive sampling tool to detect the spawning distribution of European anadromous shads (Alosa spp.). Aquat. Conserv.-Mar. Freshw. Ecosyst. 2019, 29, 148–152. [Google Scholar] [CrossRef]
- Santana, C.D.D.; Parenti, L.R.; Dillman, C.B.; Coddington, J.A.; Bastos, D.A.; Baldwin, C.C.; Zuanon, J.; Torrente-Vilara, G.; Covain, R.; Menezes, N.A.; et al. The critical role of natural history museums in advancing eDNA for biodiversity studies: A case study with Amazonian fishes. Sci. Rep. 2021, 11, 18159. [Google Scholar] [CrossRef]
- Lamy, T.; Pitz, K.J.; Chavez, F.P.; Yorke, C.E.; Miller, R.J. Environmental DNA reveals the fine-grained and hierarchical spatial structure of kelp forest fish communities. Sci. Rep. 2021, 11, 14439. [Google Scholar] [CrossRef] [PubMed]
- Elbrecht, V.; Leese, F. Can DNA-Based Ecosystem Assessments Quantify Species Abundance? Testing Primer Bias and Biomass--Sequence Relationships with an Innovative Metabarcoding Protocol. PLoS ONE 2015, 10, e0130324. [Google Scholar] [CrossRef] [PubMed]
- Piñol, J.; Mir, G.; Gomez-Polo, P.; Agustí, N. Universal and blocking primer mismatches limit the use of high-throughput DNA sequencing for the quantitative metabarcoding of arthropods. Mol. Ecol. Resour. 2015, 15, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.P.; Port, J.A.; Yamahara, K.M.; Martone, R.G.; Lowell, N.; Thomsen, P.F.; Mach, M.E.; Bennett, M.; Prahler, E.; Caldwell, M.R.; et al. Environmental monitoring. Harnessing DNA to improve environmental management. Science 2014, 344, 1455–1456. [Google Scholar] [CrossRef]
- Evans, N.T.; Olds, B.P.; Renshaw, M.A.; Turner, C.R.; Li, Y.; Jerde, C.L.; Mahon, A.R.; Pfrender, M.E.; Lamberti, G.A.; Lodge, D.M. Quantification of mesocosm fish and amphibian species diversity via environmental DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 29–41. [Google Scholar] [CrossRef]
- Chen, Y.; Qiu, K.R.; Zhong, Z.Y.; Zhou, T. Influence of environmental factors on the variability of archaeal communities in a karst wetland. Front. Microbiol. 2021, 12, 675665. [Google Scholar] [CrossRef]
- Gorički, Š.; Stanković, D.; Snoj, A.; Kuntner, M.; Jeffery, W.R.; Trontelj, P.; Pavićević, M.; Grizelj, Z.; Năpăruş-Aljančič, M.; Aljančič, G. Environmental DNA in subterranean biology: Range extension and taxonomic implications for Proteus. Sci. Rep. 2017, 7, 45054. [Google Scholar] [CrossRef]
- West, K.M.; Richards, Z.T.; Harvey, E.S.; Susac, R.; Grealy, A.; Bunce, M. Under the karst: Detecting hidden subterranean assemblages using eDNA metabarcoding in the caves of Christmas Island, Australia. Sci. Rep. 2020, 10, 21479. [Google Scholar] [CrossRef]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Environmental DNA: For Biodiversity Research and Monitoring; Oxford University Press: New York, NY, USA, 2018. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. 2022. Available online: www.fishbase.org (accessed on 23 August 2022).
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastlynimproves diversity estimates from illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R. Package Version 2.5-7. 2020. Available online: http://CRAN.R-project.org/package=vegan (accessed on 23 August 2022).
- Hadley, W.; Winston, C.; Lionel, H.; Lin, P.T.; Kohske, T.; Claus, W.; Kara, W.; Hiroaki, Y.; Dewey, D. ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. R. Package Version 3.3.4. 2021. Available online: https://ggplot2.tidyverse.org (accessed on 23 August 2022).
- Wang, W.Y.; Yang, J.X.; Chen, X.Y. Current Status and Conservation Strategics of Fish Germplasm Resources of Nanpanjiang River System in Yunnan. J. Hydroecol. 2011, 32, 19–29. (In Chinese) [Google Scholar] [CrossRef]
- Shuai, F.M.; Li, X.H.; He, A.Y.; Liu, Q.F.; Zhang, Y.Q.; Wu, Z.; Zhu, S.L. Fish Diversity and Distribution pattern of The Pearl River system in Guangxi. Acta Hydrobiol. Sin. 2020, 44, 819–828. [Google Scholar] [CrossRef]
- Akyol, O.; Özgül, A.; Düzbastılar, F.O.; Şen, H.; Urbina, J.M.O.D.; Ceyhan, T. Influence of some physico-chemical variables on wild fish richness beneath sea-cage fish farms in the Aegean Sea, Turkey. J. Mar. Biol. Assoc. UK 2020, 100, 1171–1179. [Google Scholar] [CrossRef]
- Lemos, C.H.D.P.; Ribeiro, C.V.D.M.; Oliveira, C.P.B.D.; Couto, R.D.; Copatti, C.E. Effects of interaction between pH and stocking density on the growth, haematological and biochemical responses of Nile tilapia juveniles. Aquaculture 2018, 495, 62–67. [Google Scholar] [CrossRef]
- Yanagitsuru, Y.R.; Hewitt, C.R.; Gam, L.T.H.; Phuong-Linh, P.; Rodgers, E.M.; Wang, T.; Bayley, M.; Huong, D.T.T.; Phuong, N.T.; Brauner, C.J. Effect of water pH and calcium on ion balance in five fish species of the Mekong Delta. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 232, 34–39. [Google Scholar] [CrossRef]
- Xu, L.F.; Yao, D.D.; Yang, Y.W.; Guo, X.C.; Li, J.Y.; Jiang, H.B.; An, M.; Dong, X.H.; Shao, J. Fish diversity in karst plateau artificial lakes based on environmental DNA metabarcoding technology. J. South. Agric. 2022. Available online: https://kns.cnki.net/kcms/detail/45.1381.S.20220718.1346.002.html (accessed on 19 July 2022). (In Chinese).
- Dos Santos, D.A.; Hoeinghaus, D.J.; Gomes, L.C. Spatial scales and the invasion paradox: A test using fish assemblages in a Neotropical floodplain. Hydrobiologia 2018, 817, 121–131. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Site | Longitude | Latitude | Tem | Tra | TDS | Sal | pH | DO | NH4+ |
|---|---|---|---|---|---|---|---|---|---|---|
| °C | m | mg/L | ppt | - | mg/L | mg/L | ||||
| S1 | BaXia | 105.11 | 24.95 | 22.00 | 3.00 | 261.95 | 0.19 | 6.93 | 5.90 | 0.14 |
| S2 | DaBa | 105.09 | 24.94 | 25.30 | 2.50 | 261.30 | 0.19 | 7.03 | 8.30 | 0.04 |
| S3 | HongChong | 105.06 | 24.92 | 25.60 | 1.95 | 250.90 | 0.18 | 6.99 | 8.10 | 0.03 |
| S4 | MeiJiaWan | 105.07 | 24.90 | 25.30 | 2.20 | 256.10 | 0.19 | 7.03 | 6.90 | 0.04 |
| S5 | PoNaShu | 105.04 | 24.88 | 25.70 | 2.25 | 257.40 | 0.19 | 7.00 | 5.90 | 0.05 |
| S6 | BaJie | 105.03 | 24.86 | 26.00 | 2.10 | 244.40 | 0.18 | 7.03 | 6.60 | 0.03 |
| S7 | YeYaTan | 105.04 | 24.83 | 26.30 | 1.65 | 219.70 | 0.16 | 7.07 | 7.30 | 0.02 |
| S8 | GeBu | 105.03 | 24.80 | 25.80 | 2.35 | 245.05 | 0.18 | 7.05 | 7.80 | 0.03 |
| S9 | BaAi | 104.95 | 24.77 | 25.90 | 2.15 | 254.80 | 0.19 | 6.97 | 7.90 | 0.04 |
| S10 | BaDaZhang | 104.87 | 24.74 | 27.10 | 2.10 | 227.50 | 0.17 | 6.99 | 8.50 | 0.02 |
| S11 | JinZhongShan | 104.84 | 24.68 | 26.50 | 2.05 | 234.65 | 0.17 | 7.02 | 7.80 | 0.02 |
| S12 | LaoWangZhai | 104.76 | 24.66 | 26.50 | 1.90 | 226.85 | 0.17 | 6.99 | 8.20 | 0.02 |
| S13 | HeShe | 104.73 | 24.62 | 26.40 | 1.70 | 226.85 | 0.17 | 7.06 | 8.20 | 0.02 |
| S14 | BaWan | 104.69 | 24.65 | 26.90 | 1.65 | 223.60 | 0.16 | 6.80 | 11.50 | 0.03 |
| S15 | NanGuShan | 104.61 | 24.68 | 26.50 | 2.25 | 247.00 | 0.18 | 6.96 | 8.30 | 0.03 |
| S16 | SanJIiangKou | 104.54 | 24.73 | 26.30 | 1.75 | 219.05 | 0.16 | 7.09 | 7.09 | 0.02 |
| Taxonomy | S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | S11 | S12 | S13 | S14 | S15 | S16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acipenseriformes | ||||||||||||||||
| Acipenseridae | ||||||||||||||||
| Acipenser sp. | + | + | + | + | + | + | + | + | + | + | + | + | + | + | ||
| Anabantiformes | ||||||||||||||||
| Channidae | ||||||||||||||||
| Channa sp. | + | + | + | + | ||||||||||||
| Beloniformes | ||||||||||||||||
| Adrianichthyidae | ||||||||||||||||
| Oryzias sinensis * | + | + | + | + | + | + | ||||||||||
| Centrarchiformes | ||||||||||||||||
| Sinipercidae | ||||||||||||||||
| Siniperca scherzeri * | + | + | + | + | + | + | ||||||||||
| Cichliformes | ||||||||||||||||
| Cichlidae | ||||||||||||||||
| Coptodon zillii | + | + | + | + | + | + | ||||||||||
| Oreochromis niloticus * | + | + | + | + | + | + | + | + | + | + | + | + | + | + | ||
| Oreochromis tanganicae | + | + | + | + | + | + | + | + | + | |||||||
| Oreochromis sp. | + | + | + | + | + | + | + | + | + | + | + | |||||
| Cypriniformes | ||||||||||||||||
| Acheilognathidae | ||||||||||||||||
| Acheilognathus macropterus * | + | + | + | + | ||||||||||||
| Cyprinidae | ||||||||||||||||
| Ancherythroculter wangi | + | + | + | + | + | + | + | + | + | + | + | + | + | |||
| Barbonymus schwanefeldii | + | + | ||||||||||||||
| Carassius gibelio | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Chanodichthys erythropterus * | + | + | + | |||||||||||||
| Ctenopharyngodon idella * | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Culter alburnus * | + | + | + | + | + | + | + | + | + | + | + | + | + | + | ||
| Cyprinus carpio * | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Hemiculter leucisculus * | + | + | + | + | + | + | + | + | + | + | + | |||||
| Hemiculter tchangi | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Hemiculterella macrolepis | + | + | + | + | + | + | + | + | + | |||||||
| Hypophthalmichthys molitrix * | + | + | + | + | ||||||||||||
| Hypophthalmichthys nobilis * | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Osteochilus salsburyi * | + | + | + | + | + | + | + | + | + | |||||||
| Parabramis pekinensis * | + | + | + | |||||||||||||
| Procypris rabaudi | + | + | + | |||||||||||||
| Squalidus argentatus * | + | + | + | |||||||||||||
| Squaliobarbus curriculus * | + | + | + | + | + | + | + | + | + | |||||||
| Spinibarbus sinensis | + | + | + | + | + | + | ||||||||||
| Xenocypris davidi * | + | + | + | |||||||||||||
| Gobiiformes | ||||||||||||||||
| Gobiidae | ||||||||||||||||
| Rhinogobius similis | + | + | + | |||||||||||||
| Salmoniformes | ||||||||||||||||
| Salmonidae | ||||||||||||||||
| Prosopium coulterii | + | + | + | + | ||||||||||||
| Siluriformes | ||||||||||||||||
| Auchenipteridae | ||||||||||||||||
| Tatia intermedia | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Bagridae | ||||||||||||||||
| Hemibagrus macropterus * | + | + | + | + | + | + | ||||||||||
| Pseudobagrus crassilabris * | + | + | + | + | + | + | + | + | + | + | + | + | ||||
| Pseudobagrus medianalis | + | + | + | + | + | + | + | + | + | |||||||
| Pseudobagrus ondon | + | + | + | + | + | + | + | + | + | + | + | + | + | |||
| Pseudobagrus pratti | + | + | + | + | + | + | + | + | + | + | ||||||
| Tachysurus fulvidraco * | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Clariidae | ||||||||||||||||
| Clarias gariepinus * | + | + | + | + | + | + | + | + | + | + | + | + | ||||
| Siluridae | ||||||||||||||||
| Silurus soldatovi | + | + | + |
| df | SS | MS | F | R2 | p-Value | |
|---|---|---|---|---|---|---|
| Group | 2 | 0.64 | 0.32 | 1.70 | 0.22 | 0.009 |
| Residuals | 12 | 2.27 | 0.19 | 0.78 | ||
| Total | 14 | 2.90 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Yang, D.; Wang, Y.; Li, J.; Guo, X.; Zeng, C.; Jiang, H.; An, M.; Shao, J. Environmental DNA Captures Variations in Fish Assemblages with Distance from Dams in Karst Reservoirs. Water 2023, 15, 73. https://doi.org/10.3390/w15010073
Xu L, Yang D, Wang Y, Li J, Guo X, Zeng C, Jiang H, An M, Shao J. Environmental DNA Captures Variations in Fish Assemblages with Distance from Dams in Karst Reservoirs. Water. 2023; 15(1):73. https://doi.org/10.3390/w15010073
Chicago/Turabian StyleXu, Longfei, Dandan Yang, Yizhou Wang, Junyi Li, Xingchen Guo, Cong Zeng, Haibo Jiang, Miao An, and Jian Shao. 2023. "Environmental DNA Captures Variations in Fish Assemblages with Distance from Dams in Karst Reservoirs" Water 15, no. 1: 73. https://doi.org/10.3390/w15010073
APA StyleXu, L., Yang, D., Wang, Y., Li, J., Guo, X., Zeng, C., Jiang, H., An, M., & Shao, J. (2023). Environmental DNA Captures Variations in Fish Assemblages with Distance from Dams in Karst Reservoirs. Water, 15(1), 73. https://doi.org/10.3390/w15010073