
Ecosystems of Inland Saline Waters in the World of Change




{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Diversity of Saline Inland Waters
3. The Relationship between Biodiversity and Services
4. Saline Lakes, Bioresources, and Business
5. Risks and Challenges of Protecting Inland Saline Lakes and Their Biodiversity
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shadrin, N.; Mirzoeva, N.; Kravchenko, N.; Miroshnichenko, O.; Tereshchenko, N.; Anufriieva, E. Trace elements in the bottom sediments of the Crimean saline lakes. Is it possible to explain their concentration variability? Water 2020, 12, 2364. [Google Scholar] [CrossRef]
- Shadrin, N.; Stetsiuk, A.; Anufriieva, E. Differences in Mercury Concentrations in Water and Hydrobionts of the Crimean Saline Lakes: Does Only Salinity Matter? Water 2022, 14, 2613. [Google Scholar] [CrossRef]
- Mirzoeva, N.; Shadrin, N.; Proskurnin, V.; Arkhipova, S.; Moseychenko, I.; Anufriieva, E. The Sedimentation Rate in the Crimean Hypersaline Lake Aktashskoye Estimated Using the Post-Chernobyl Artificial Radionuclide 90Sr as a Radiotracer. Water 2022, 14, 2506. [Google Scholar] [CrossRef]
- De Necker, L.; Brendonck, L.; Van Vuren, J.; Wepener, V.; Smit, N.J. Aquatic invertebrate community resilience and recovery in response to a supra-seasonal drought in an ecologically important naturally saline lake. Water 2021, 13, 948. [Google Scholar] [CrossRef]
- Zsuga, K.; Inelova, Z.; Boros, E. Zooplankton community structure in Shallow Saline steppe inland waters. Water 2021, 13, 1164. [Google Scholar] [CrossRef]
- Lantushenko, A.; Meger, Y.; Gadzhi, A.; Anufriieva, E.; Shadrin, N. Artemia spp. (Crustacea, Anostraca) in Crimea: New Molecular Genetic Results and New Questions without Answers. Water 2022, 14, 2617. [Google Scholar] [CrossRef]
- Shadrin, N.; Balycheva, D.; Anufriieva, E. Microphytobenthos in the hypersaline water bodies, the case of bay Sivash (Crimea): Is salinity the main determinant of species composition? Water 2021, 13, 1542. [Google Scholar] [CrossRef]
- Saccò, M.; White, N.E.; Campbell, M.; Allard, S.; Humphreys, W.F.; Pringle, P.; Sepanta, F.; Laini, A.; Allentoft, M.E. Metabarcoding under brine: Microbial ecology of five hypersaline lakes at Rottnest Island (WA, Australia). Water 2021, 13, 1899. [Google Scholar] [CrossRef]
- Redón, S.; Gajardo, G.; Vasileva, G.P.; Sánchez, M.I.; Green, A.J. Explaining Variation in Abundance and Species Diversity of Avian Cestodes in Brine Shrimps in the Salar de Atacama and Other Chilean Wetlands. Water 2021, 13, 1742. [Google Scholar] [CrossRef]
- Shadrin, N.; Kolesnikova, E.; Revkova, T.; Latushkin, A.; Chepyzhenko, A.; Dyakov, N.; Anufriieva, E. Macrostructure of benthos along a salinity gradient: The case of Sivash Bay (the Sea of Azov), the largest hypersaline lagoon worldwide. J. Sea Res. 2019, 154, 101811. [Google Scholar] [CrossRef]
- Anufriieva, E.; Kolesnikova, E.; Revkova, T.; Latushkin, A.; Shadrin, N. Human-Induced Sharp Salinity Changes in the World’s Largest Hypersaline Lagoon Bay Sivash (Crimea) and Their Effects on the Ecosystem. Water 2022, 14, 403. [Google Scholar] [CrossRef]
- Ragnvaldsson, D.; Herting, G.; Jönsson, A.; Odnevall, I. Applying Generic Water Quality Criteria to Cu and Zn in a Dynamic Aquatic Environment—The Case of the Brackish Water Formation Strömmen-Saltsjön. Water 2022, 14, 847. [Google Scholar] [CrossRef]
- Shadrin, N.; Anufriieva, E.; Latushkin, A.; Prazukin, A.; Yakovenko, V. Daily Rhythms and Oxygen Balance in the Hypersaline Lake Moynaki (Crimea). Water 2022, 14, 3753. [Google Scholar] [CrossRef]
- Gleick, P.H. Water in Crisis; Oxford University Press: New York, NY, USA, 1993; 504p. [Google Scholar]
- Williams, W.D. Environmental threats to salt lakes and the likely status of inland saline ecosystems in 2025. Environ. Conserv. 2002, 29, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Pinneker, E.V. Brines of the Angara-Lena Artesian Basin; Nauka: Moscow, Russia, 1966; 332p. (In Russian) [Google Scholar]
- Beltyukov, G.V. On the chemical characteristics of underground salt lakes. Peshchery 1969, 7, 44–51. (In Russian) [Google Scholar]
- Maksimovich, G.A.; Beltyukov, G.V.; Golubev, B.M. Salt formations of underground lakes. Peshchery 1966, 6, 25–32. (In Russian) [Google Scholar]
- Zhukov, V.A.; Tolstoy, M.P.; Troyansky, S.V. Artesian Waters of the Carboniferous Deposits of the Paleozoic Basin Near Moscow; GONTI: Moscow, Russia; St. Petersburg, Russia, 1939; 216p. (In Russian) [Google Scholar]
- Lebedeva, N.A. Natural Resources of Underground Waters of the Moscow Artesian Basin.; Nauka: Moscow, Russia, 1972; 148p. [Google Scholar]
- Sidkina, E.S. The brines of the western part of the Tunguska artesian basin. Geokhimiya 2015, 8, 743. (In Russian) [Google Scholar] [CrossRef]
- Ward, J.A.; Slater, G.F.; Moser, D.P.; Lin, L.H.; Lacrampe-Couloume, G.; Bonin, A.S.; Davidson, M.; Hall, J.A.; Mislowack, B.; Bellamy, R.E.; et al. Microbial hydrocarbon gases in the Witwatersrand Basin, South Africa: Implications for the deep biosphere. Geochim. Cosmochim. Acta. 2004, 68, 3239–3250. [Google Scholar] [CrossRef]
- Onstott, T.C.; Colwell, F.S.; Kieft, T.L.; Murdoch, L.; Phelps, T.J. New horizons for deep subsurface microbiology. Microbe 2009, 4, 499–505. [Google Scholar] [CrossRef]
- Colman, D.R.; Poudel, S.; Stamps, B.W.; Boyd, E.S.; Spear, J.R. The deep, hot biosphere: Twenty-five years of retrospection. Proc. Natl. Acad. Sci. USA 2017, 114, 6895–6903. [Google Scholar] [CrossRef] [Green Version]
- Borgonie, G.; García-Moyano, A.; Litthauer, D.; Bert, W.; Bester, A.; van Heerden, E.; Möller, C.; Erasmus, M.; Onstott, T.C. Nematoda from the terrestrial deep subsurface of South Africa. Nature 2011, 474, 79–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ino, K.; Konno, U.; Kouduka, M.; Hirota, A.; Togo, Y.S.; Fukuda, A.; Komatsu, D.; Tsunogai, U.; Tanabe, A.S.; Yamamoto, S.; et al. Deep microbial life in high-quality granitic groundwater from geochemically and geographically distinct underground boreholes. Environ. Microbiol. Rep. 2016, 8, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Saccò, M.; Blyth, A.J.; Venarsky, M.; Humphreys, W.F. Trophic Interactions in Subterranean Environments. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Golubkov, S.M.; Shadrin, N.V.; Golubkov, M.S.; Balushkina, E.V.; Litvinchuk, L.F. Food chains and their dynamics in ecosystems of shallow lakes with different water salinities. Russ. J. Ecol. 2018, 49, 442–448. [Google Scholar] [CrossRef]
- Moscatello, S.; Belmonte, G. Egg banks in hypersaline lakes of the South-East Europe. Saline Syst. 2009, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M. Saline Lakes and Salt Basin Deposits in China; Science Press: Beijing, China, 2014; 321p. [Google Scholar]
- Timms, B.V. Saline lakes of the Paroo, inland New South Wales, Australia. Hydrobiologia 1993, 267, 269–289. [Google Scholar] [CrossRef]
- Ashton, P.J.; Schoeman, F.R. Limnological studies on the Pretoria Salt Pan, a hypersaline maar lake. Hydrobiologia 1983, 99, 61–73. [Google Scholar] [CrossRef]
- Schagerl, M.; Renaut, R.W. Dipping into the soda lakes of East Africa. In Soda Lakes of East Africa; Schagerl, M., Ed.; Springer: Cham, Switherland, 2016; pp. 3–24. [Google Scholar]
- Hammer, U.T. Saline Lake Ecosystems of the World; Springer Science & Business Media: Berlin, Germany, 1986; 616p. [Google Scholar]
- Laybourn-Parry, J.; Quayle, W.; Henshaw, T. The biology and evolution of Antarctic saline lakes in relation to salinity and trophy. Polar Biol. 2002, 25, 542–552. [Google Scholar] [CrossRef]
- Thomas, D.N.; Dieckmann, G.S. Antarctic sea ice--a habitat for extremophiles. Science 2002, 295, 641–644. [Google Scholar] [CrossRef] [Green Version]
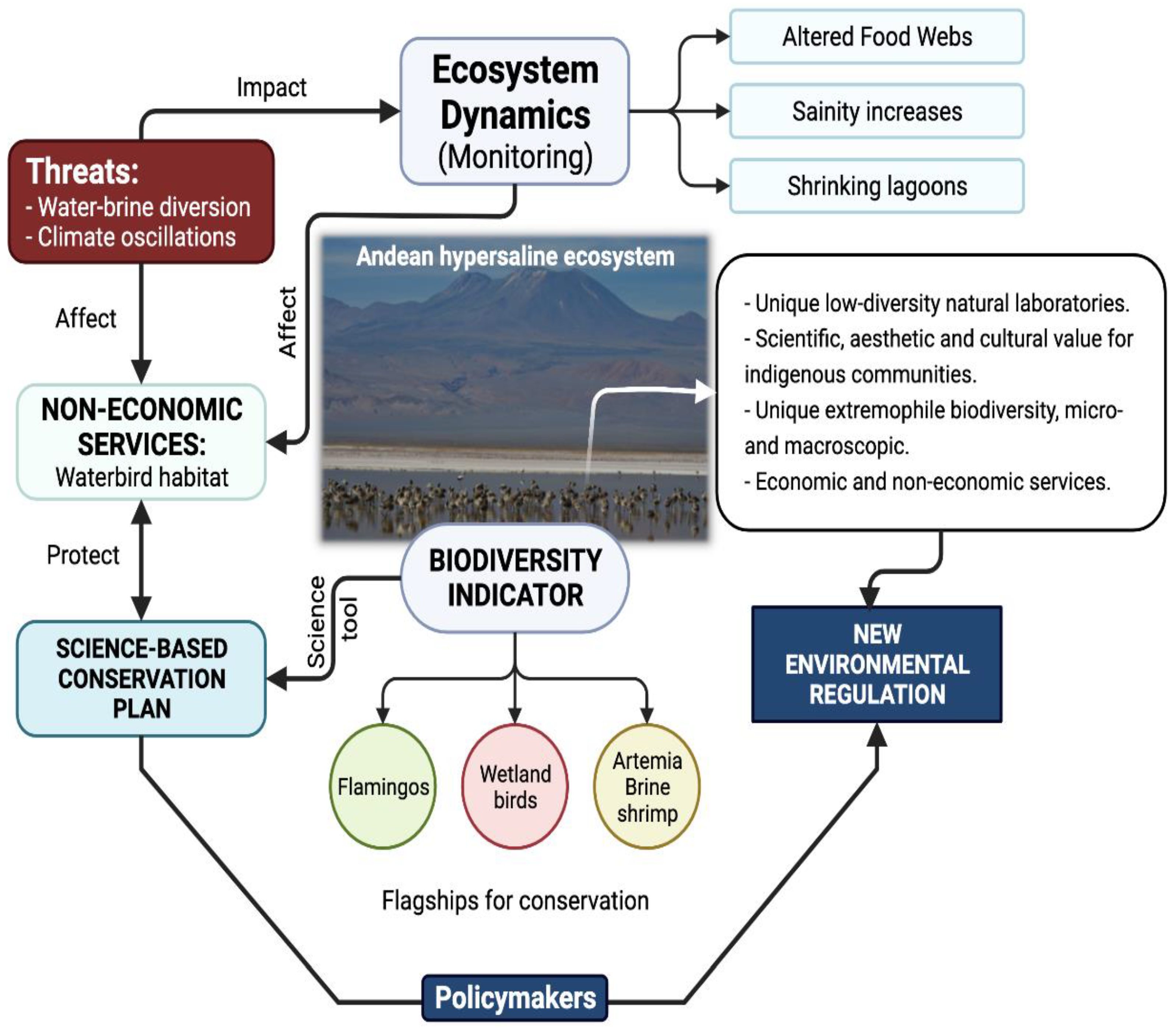
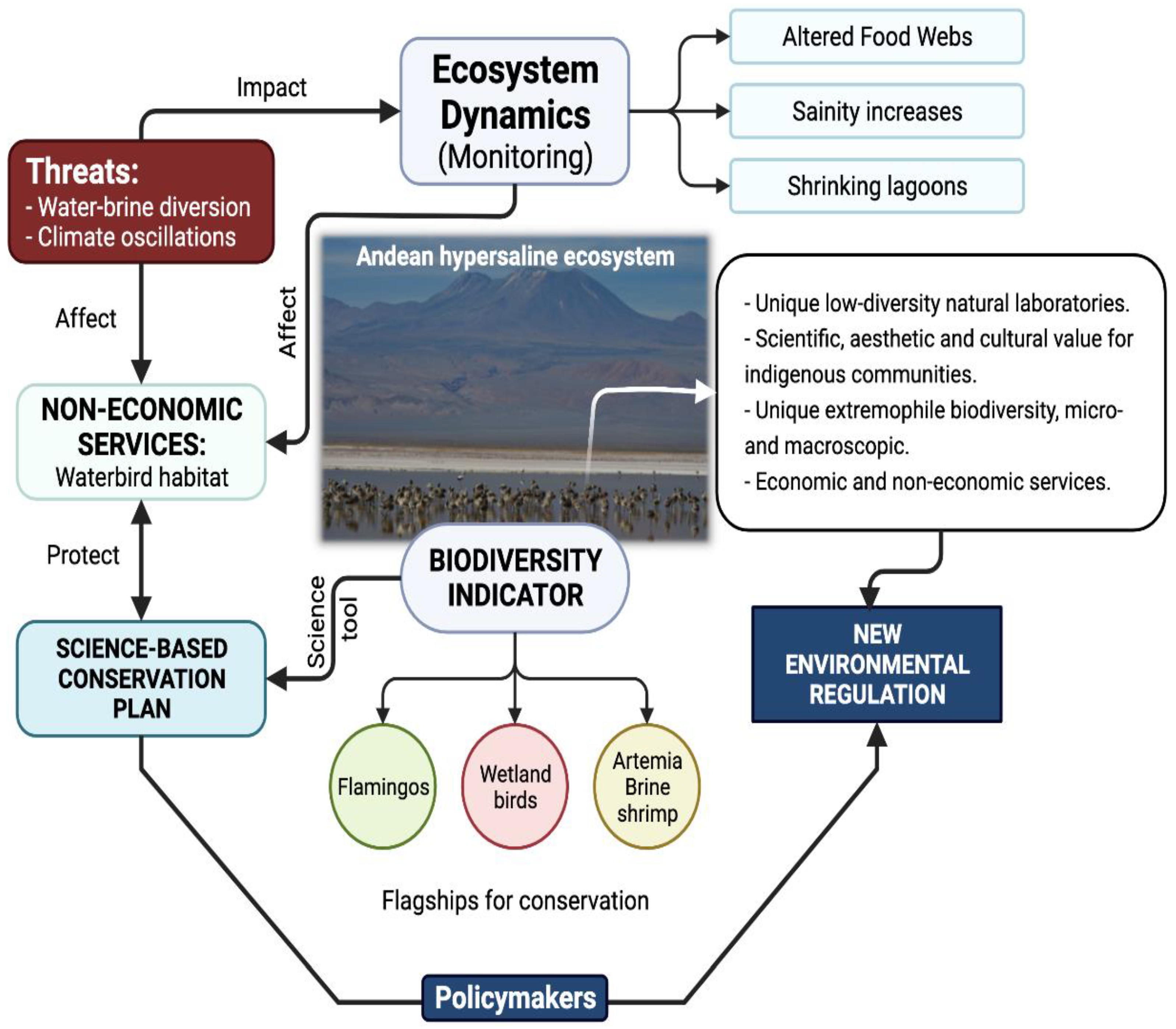
- Gajardo, G.; Redón, S. Andean hypersaline lakes in the Atacama Desert, northern Chile: Between lithium exploitation and unique biodiversity conservation. Conserv. Sci. Pract. 2019, 1, 1–8. [Google Scholar]
- Berdugo, M.; Delgado-Baquerizo, M.; Soliveres, S.; Hernández-Clemente, R.; Zhao, Y.; Gaitán, J.J.; Gross, N.; Saiz, H.; Maire, V.; Lehmann, A.; et al. Global ecosystem thresholds driven by aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Yao, N.; Li, L.; Feng, P.; Feng, H.; Li Liu, D.; Liu, Y.; Jiang, K.; Hu, X.; Li, Y. Projections of drought characteristics in China based on a standardized precipitation and evapotranspiration index and multiple GCMs. Sci. Total Environ. 2020, 704, 135245. [Google Scholar] [CrossRef] [PubMed]
- Kjerfve, B.; Schettini, C.A.; Knoppers, B.; Lessa, G.; Ferreira, H.O. Hydrology and salt balance in a large, hypersaline coastal lagoon: Lagoa de Araruama, Brazil. Estuar. Coast. Shelf Sci. 1996, 42, 701–725. [Google Scholar] [CrossRef] [Green Version]
- Shadrin, N. Hypersaline lakes as the polyextreme habitats for life. In Introduction to Salt Lake Sciences; Zheng, M., Deng, T., Oren, A., Eds.; Science Press: Beijing, China, 2018; pp. 180–187. [Google Scholar]
- Efimov, V.V.; Timofeev, N.A. Heat Balance Studies of the Black and Azov Seas; VNIIGMI MCD: Obninsk, Russia, 1990; 236p. (In Russian) [Google Scholar]
- Salhotra, A.M.; Adams, E.E.; Harleman, D.R. Effect of salinity and ionic composition on evaporation: Analysis of Dead Sea evaporation pans. Water Resour. Res. 1985, 21, 1336–1344. [Google Scholar] [CrossRef]
- Khromov, S.P.; Petrosyan, M.A. Meteorology and Fundamentals of Climatology. Publishing House of Moscow University: Moscow, Russia, 2001; 527p. [Google Scholar]
- Jellison, R.O. Conservation of saline lakes in the 21st century. In Proceedings of the Environmental Future of Aquatic Ecosystems Conference, Zurich, Switzerland, 23–27 March 2003; pp. 23–27. [Google Scholar]
- Shadrin, N.; Zheng, M.; Oren, A. Past, present and future of saline lakes: Research for global sustainable development. Chin. J. Oceanol. Limnol. 2015, 33, 1349–1353. [Google Scholar] [CrossRef]
- Jia, Q.; Anufriieva, E.; Liu, X.; Kong, F.; Shadrin, N. Intentional introduction of Artemia sinica (Anostraca) in the high-altitude Tibetan lake Dangxiong Co: The new population and consequences for the environment and for humans. Chin. J. Oceanol. Limnol. 2015, 33, 1451–1460. [Google Scholar] [CrossRef]
- Abdrakhmanov, A.R.; Budkova, G.A.; Abrakhmanov, A.A. Salt Lakes of the Sol-Iletsk Resort; Soyuz: Orenburg, Russia, 2008; 196p. (In Russian) [Google Scholar]
- Shadrin, N.; Anufriieva, E.; Galagovets, E. Distribution and historical biogeography of Artemia Leach, 1819 (Crustacea: Anostraca) in Ukraine. Int. J. Artemia Biol. 2012, 2, 30–42. [Google Scholar]
- Williams, W.D. Salinisation: A major threat to water resources in the arid and semi-arid regions of the world. Lakes Reserv. Res. Manag. 1999, 4, 85–91. [Google Scholar] [CrossRef]
- Van Meter, R.J.; Swan, C.M. Road salts as environmental constraints in urban pond food webs. PLoS ONE 2014, 9, e90168. [Google Scholar] [CrossRef]
- Mabidi, A.; Bird, M.S.; Perissinotto, R. Increasing salinity drastically reduces hatching success of crustaceans from depression wetlands of the semi-arid Eastern Cape Karoo region, South Africa. Sci. Rep. 2018, 8, 5983. [Google Scholar] [CrossRef] [Green Version]
- Anufriieva, E.V.; Goher, M.E.; Hussian, A.M.; El-Sayed, S.M.; Hegab, M.H.; Tahoun, U.M.; Shadrin, N.V. Ecosystems of artificial saline lakes. A case of Lake Magic in Wadi El-Rayan depression (Egypt). Knowl. Manag. Aquat. Ecosyst. 2020, 421, 31. [Google Scholar] [CrossRef]
- Kurlansky, M. Salt: A World History; Penguin Books: New York, NY, USA, 2002; 484p. [Google Scholar]
- Shadrin, N.V.; Gerasimenko, L.M.; Mikhodyuk, O.S.; Marian, M.P. Bottom cyanobacteria of hypersaline reservoirs in India. Naukovi Zapisky Ternopil Gosudarstvennogo Pedodologicheskogo Universiteta imeni. V. Gnatiuk. Ser. Biol. 2005, 4, 27–31. (In Russian) [Google Scholar]
- Mara, S.; Deákb, Ş.; Deákb, G.; Stefanescu, L.; Vlad, S.N. Salt Mining Lake Pits in Romania, a Sustainable Heritage. In Mine Water and the Environment, Proceedings of the 10th International Mine Water Association Congress, Karlovy Vary, Czech Republic, 2–5 June 2008; VŠB-Technical University of Ostrava, Faculty of Mining and Geology: Ostrava, Czech Republic, 2008; pp. 595–598. [Google Scholar]
- Cohen, M.J.; Morrison, J.I.; Glenn, E.P. Haven or Hazard: The Ecology and Future of the Salton Sea: A Report; Pacific Institute for Studies in Development, Environment, and Security: Oakland, CA, USA, 1999; 64p. [Google Scholar]
- El-Shabrawy, G.M.; Dumont, H.J. The Fayum depression and its lakes. In The Nile; Dumont, H.J., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 95–124. [Google Scholar]
- Embabi, N.S. The Fayum Depression. In Landscapes and Landforms of Egypt. Landforms and Evolution; Embaby, N.S., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 153–162. [Google Scholar]
- Null, S.E.; Wurtsbaugh, W.A. Water development, consumptive water uses, and Great Salt Lake. In Great Salt Lake Biology; Baxter, B.K., Butler, J.K., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–21. [Google Scholar]
- Zadereev, E.; Lipka, O.; Karimov, B.; Krylenko, M.; Elias, V.; Pinto, I.S.; Alizade, V.; Anker, Y.; Feest, A.; Kuznetsova, D.; et al. Overview of past, current, and future ecosystem and biodiversity trends of inland saline lakes of Europe and Central Asia. Inland Waters 2020, 10, 438–452. [Google Scholar] [CrossRef]
- Foroumandi, E.; Nourani, V.; Kantoush, S.A. Investigating the main reasons for the tragedy of large saline lakes: Drought, climate change, or anthropogenic activities? A call to action. J. Arid Environ. 2022, 196, 104652. [Google Scholar] [CrossRef]
- Kjerfve, B. Coastal lagoons. ElsevierOceanogr. Ser. 1994, 60, 1–8. [Google Scholar]
- Pérez-Ruzafa, Á.; Marcos, C.; Pérez-Ruzafa, I.M. Recent advances in coastal lagoons ecology: Evolving old ideas and assumptions. Transit. Waters Bull. 2012, 5, 50–74. [Google Scholar]
- El-Shabrawy, G.M.; Anufriieva, E.; Shadrin, N. Tintinnina (Ciliophora) and Foraminifera in plankton of hypersaline Lagoon Bardawil (Egypt): Spatial and temporal variability. Turk. J. Zool. 2018, 42, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Tweedley, J.R.; Dittmann, S.R.; Whitfield, A.K.; Withers, K.; Hoeksema, S.D.; Potter, I.C. Hypersalinity: Global distribution, causes and effects on the biota of estuaries and lagoons. In Coasts and Estuaries: The Future; Wolanski, E., Day, J., Elliott, M., Ramesh, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 523–546. [Google Scholar]
- Roberts, D.A.; Johnston, E.L.; Knott, N.A. Impacts of desalination plant discharges on the marine environment: A critical review of published studies. Water Res. 2010, 44, 5117–5128. [Google Scholar] [CrossRef]
- Uddin, S.; Al Ghadban, A.N.; Khabbaz, A. Localized hypersaline waters in Arabian Gulf from desalination activity—An example from South Kuwait. Environ. Monit. Assess. 2011, 181, 587–594. [Google Scholar] [CrossRef]
- Anufriieva, E.; Shadrin, N. The long-term changes in plankton composition: Is Bay Sivash transforming back into one of the world’s largest habitats of Artemia sp. (Crustacea, Anostraca)? Aquac. Res. 2020, 51, 341–350. [Google Scholar] [CrossRef]
- Zinchenko, T.D.; Golovatyuk, L.V.; Abrosimova, E.V.; Popchenko, T.V.; Nikulina, T.D. Changes in macrozoobenthos communities under a mineralization gradient in the rivers of the basin of the hyperhaline lake Elton (2006–2013). Izv. RAS SamSC 2017, 19, 140–156. (In Russian) [Google Scholar]
- Velasco, J.; Millán, A.; Hernández, J.; Gutiérrez, C.; Abellán, P.; Sánchez, D.; Ruiz, M. Response of biotic communities to salinity changes in a Mediterranean hypersaline stream. Saline Syst. 2006, 2, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccò, M.; White, N.E.; Harrod, C.; Salazar, G.; Aguilar, P.; Cubillos, C.F.; Meredith, K.; Baxter, B.K.; Oren, A.; Anufriieva, E.; et al. Salt to conserve: A review on the ecology and preservation of hypersaline ecosystems. Biol. Rev. 2021, 96, 2828–2850. [Google Scholar] [CrossRef] [PubMed]
- Wurtsbaugh, W.A.; Miller, C.; Null, S.E.; DeRose, R.J.; Wilcock, P.; Hahnenberger, M.; Howe, F.; Moore, J. Decline of the world’s saline lakes. Nat. Geosci. 2017, 10, 816–821. [Google Scholar] [CrossRef]
- Intriago, P.; Jones, D.A. Bacteria as food for Artemia. Aquaculture 1993, 113, 115–127. [Google Scholar] [CrossRef]
- Tkavc, R.; Ausec, L.; Oren, A.; Gunde-Cimerman, N. Bacteria associated with Artemia spp. along the salinity gradient of the solar salterns at Eilat (Israel). FEMS Microbiol. Ecol. 2011, 77, 310–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajardo, G.M.; Beardmore, J.A. The brine shrimp Artemia: Adapted to critical life conditions. Front. Physiol. 2012, 22, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Los Ríos, P.A.; Gajardo, G.O. The brine shrimp Artemia (Crustacea; Anostraca): A model organism to evaluate management policies in aquatic resources. Rev. Chil. Hist. Nat. 2004, 77, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. Halophilic microbial communities and their environments. Curr. Opin. Biotechnol. 2015, 33, 119–124. [Google Scholar] [CrossRef]
- Golubkov, S.; Kemp, R.; Golubkov, M.; Balushkina, E.; Litvinchuk, L.; Gubelit, Y. Biodiversity and the functioning of hypersaline lake ecosystems from Crimea Peninsula (Black Sea). Fundam. Appl. Limnol. 2007, 169, 79–87. [Google Scholar] [CrossRef]
- Green, A.J.; Figuerola, J. Recent advances in the study of long-distance dispersal of aquatic invertebrates via birds. Divers. Distrib. 2005, 11, 149–156. [Google Scholar] [CrossRef]
- Green, A.J.; Sánchez, M.I.; Amat, F.; Figuerola, J.; Hontoria, F.; Ruiz, O.; Hortas, F. Dispersal of invasive and native brine shrimps Artemia (Anostraca) via waterbirds. Limnol. Oceanogr. 2005, 50, 737–742. [Google Scholar] [CrossRef] [Green Version]
- Sorgeloos, P.; Dhert, P.; Candreva, P. Use of the brine shrimp, Artemia spp., in marine fish larviculture. Aquaculture 2001, 200, 147–159. [Google Scholar] [CrossRef]
- Le, T.H.; Van Hoa, N.; Sorgeloos, P.; Van Stappen, G. Artemia feeds: A review of brine shrimp production in the Mekong Delta, Vietnam. Rev. Aquac. 2019, 11, 1169–1175. [Google Scholar] [CrossRef]
- Van Stappen, G.; Sui, L.; Hoa, V.N.; Tamtin, M.; Nyonje, B.; de Medeiros Rocha, R.; Sorgeloos, P.; Gajardo, G. Review on integrated production of the brine shrimp Artemia in solar salt ponds. Rev. Aquac. 2020, 12, 1054–1071. [Google Scholar] [CrossRef]
- Shadrin, N.; Yakovenko, V.; Anufriieva, E. Suppression of Artemia spp. (Crustacea, Anostraca) populations by predators in the Crimean hypersaline lakes: A review of the evidence. Int. Rev. Hydrobiol. 2019, 104, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Panwar, R.; Ober, H.; Pinkse, J. The uncomfortable relationship between business and biodiversity: Advancing research on business strategies for biodiversity protection. Bus. Strateg. Environ. 2022; in press. [Google Scholar] [CrossRef]
- Wurtsbaugh, W.A.; Sima, S. Contrasting Management and Fates of Two Sister Lakes: Great Salt Lake (USA) and Lake Urmia (Iran). Water 2022, 14, 3005. [Google Scholar] [CrossRef]
- Wright, J. Artemia, the ‘Magic Powder’ Fueling a Multi-Billion-Dollar Industry. Global Aquaculture Advocate. 2017. Available online: https://www.globalseafood.org/advocate/artemia-the-magic-powder-fueling-a-multi-billion-dollar-industry/ (accessed on 19 December 2022).
- Belovsky, G.E.; Stephens, D.; Perschon, C.; Birdsey, P.; Paul, D.; Naftz, D.; Baskin, R.; Larson, C.; Mellison, C.; Luft, J.; et al. The Great Salt Lake Ecosystem (Utah, USA): Long term data and a structural equation approach. Ecosphere 2011, 2, 1–40. [Google Scholar] [CrossRef]
- Wurtsbaugh, W.A. The Great Salt Lake Ecosystem (Utah, USA): Long term data and a structural equation approach: Comment. Ecosphere 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Conover, M.R.; Caudell, J.N. Energy Budgets for Eared Grebes on the Great Salt Lake and Implications for Harvest of Brine Shrimp. J. Wildl. Manag. 2009, 73, 1134–1139. [Google Scholar] [CrossRef]
- Marconi, P.; Arengo, F.; Clark, A. The arid Andean plateau waterscapes and the lithium triangle: Flamingos as flagships for conservation of high-altitude wetlands under pressure from mining development. Wetl. Ecol. Manag. 2022, 30, 827–852. [Google Scholar] [CrossRef]
- Gutierrez, J.S.; Moore, J.N.; Donnelly, J.P.; Dorador, C.; Navedo, J.G.; Senner, N.R. Climate change and lithium mining influence flamingo abundance in the Lithium Triangle. Proc. R. Soc. B Biol. Sci. 2022, 289, 20212388. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Sepúlveda, R. Environmental degradation through mining for energy resources: The case of the shrinking Laguna Santa Rosa wetland in the Atacama Region of Chile. Energy Geosci. 2022, 3, 182–190. [Google Scholar] [CrossRef]
- Prazukin, A.V.; Anufriieva, E.V.; Shadrin, N.V. Is biomass of filamentous green algae Cladophora spp. (Chlorophyta, Ulvophyceae) an unlimited cheap and valuable resource for medicine and pharmacology? A review. Rev. Aquac. 2020, 12, 2493–2510. [Google Scholar] [CrossRef]
- Anufriieva, E.V. How can saline and hypersaline lakes contribute to aquaculture development? A review. J. Oceanol. Limnol. 2018, 36, 2002–2009. [Google Scholar] [CrossRef]
- Calderón-Arreola, J.B.; Alcocer, J.; Oseguera, L.A. A Note of a Unique Inland, Saline Water Fishery: Brine Flies (Diptera: Ephydridae) of Lake Cuitzeo, Mexico. Water 2022, 14, 900. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, W.; Huo, Y.; Yin, X.; Wei, J.; Wang, S.; Wang, Y. Influence of Seawater Salinity on the Survival, Growth, Development and Neonate Production of Scapholeberis mucronata (O. F. Müller) (Crustacea: Cladocera). Water 2022, 14, 3706. [Google Scholar] [CrossRef]
- Yakovenko, V.; Shadrin, N.; Anufriieva, E. The Prawn Palaemon adspersus in the Hypersaline Lake Moynaki (Crimea): Ecology, Long-Term Changes, and Prospects for Aquaculture. Water 2022, 14, 2786. [Google Scholar] [CrossRef]
- Shadrin, N.; Yakovenko, V.; Anufriieva, E. Feeding of the Amphipod Gammarus aequicauda in the Presence of the Planktonic Cladoceran Moina salina and the Benthic Chironomid Larvae Baeotendipes noctivagus. Water 2022, 14, 3948. [Google Scholar] [CrossRef]
- Laikre, L.; Hoban, S.; Bruford, M.W.; Segelbacher, G.; Allendorf, F.W.; Gajardo, G.; Rodríguez, A.G.; Hedrick, P.W.; Heuertz, M.; Hohenlohe, P.A.; et al. Post-2020 goals overlook genetic diversity. Science 2020, 367, 1083–1085. [Google Scholar] [CrossRef] [Green Version]
- Hoban, S.; Bruford, M.; Jackson, J.D.U.; Lopes-Fernandes, M.; Heuertz, M.; Hohenlohe, P.A.; Paz-Vinas, I.; Sjögren-Gulve, P.; Segelbacher, G.; Vernesi, C.; et al. Genetic diversity targets and indicators in the CBD post-2020 Global Biodiversity Framework must be improved. Biol. Conserv. 2020, 248, 108654. [Google Scholar] [CrossRef]
- Gajardo, G.M.; Sorgeloos, P.; Beardmore, J.A. Inland hypersaline lakes and the brine shrimp Artemia as simple models for biodiversity analysis at the population level. Saline Syst. 2006, 2, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigolot, C. Transdisciplinarity as a discipline and a way of being: Complementarities and creative tensions. Humanit. Soc. Sci. Commun. 2020, 7, 1–5. [Google Scholar] [CrossRef]
- Garcés, I.; Alvarez, G. Water mining and extractivism of the Salar de Atacama, Chile. WIT Trans. Ecol. Environ. 2020, 245, 189–199. [Google Scholar]
- Lorca, M.; Olivera Andrade, M.; Escosteguy, M.; Köppel, J.; Scoville-Simonds, M.; Hufty, M. Mining indigenous territories: Consensus, tensions and ambivalences in the Salar de Atacama. Extr. Ind. Soc. 2022, 9, 101047. [Google Scholar] [CrossRef]
- Forget, M.; Bos, V. Harvesting lithium and sun in the Andes: Exploring energy justice and the new materialities of energy transitions. Energy Res. Soc. Sci. 2022, 87, 102477. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shadrin, N.; Anufriieva, E.; Gajardo, G. Ecosystems of Inland Saline Waters in the World of Change. Water 2023, 15, 52. https://doi.org/10.3390/w15010052
Shadrin N, Anufriieva E, Gajardo G. Ecosystems of Inland Saline Waters in the World of Change. Water. 2023; 15(1):52. https://doi.org/10.3390/w15010052
Chicago/Turabian StyleShadrin, Nickolai, Elena Anufriieva, and Gonzalo Gajardo. 2023. "Ecosystems of Inland Saline Waters in the World of Change" Water 15, no. 1: 52. https://doi.org/10.3390/w15010052
APA StyleShadrin, N., Anufriieva, E., & Gajardo, G. (2023). Ecosystems of Inland Saline Waters in the World of Change. Water, 15(1), 52. https://doi.org/10.3390/w15010052