
Comparative Biology of Daphniopsis tibetana from Different Habitats under Seawater Acclimation

Key Laboratory of Hydrobiology in Liaoning Province, College of Fisheries and Life Science, Dalian Ocean University, Dalian 116023, China
*
Author to whom correspondence should be addressed.
Water 2023, 15(1), 34; https://doi.org/10.3390/w15010034
Submission received: 15 October 2022
/
Revised: 12 December 2022
/
Accepted: 17 December 2022
/
Published: 22 December 2022
(This article belongs to the Special Issue Advances in Aquaculture Ecology Research)
Abstract
:In this paper, we used experimental ecology methods and third-generation transcriptome sequencing to see the differences in growth, development, and reproduction of three strains of Daphniopsis tibetana Sars, 1903 from different locations in Tibet (Lake Namukacuo, NMKC; Lake Pengcuo, PC; and Lake Zigetangcuo, ZGTC). We also wanted to determine if the genes had changed after seawater-domesticated D. tibetana was reared in a laboratory. The results showed that at 15–16 ppt salinity and 15 ± 0.5 °C, the NMKC strain exhibited the highest survival rate of 26 d, and the ZGTC strain had the lowest survival rate at 53 days of culture. The body length was observed in NMKC (153.6 ± 12.1%), followed by PC (136.4 ± 16.1%), and then ZGTC (86.2 ± 7.6%). Combined, wild-type and seawater-acclimated D. tibetana produced 7252 DEGs, of which 4146 were up-regulated and 3106 were down-regulated. DEGs were subjected to gene ontology enrichment analysis. The DEGs were mainly enriched in single-organism localization, transporter activity, macromolecule localization, and organic substance transport. The Kyoto Encyclopedia of Genes and Genomes enrichment analysis was also performed and revealed that the RNA transport, protein digestion and absorption, and protein processing in the endoplasmic reticulum pathways were highly enriched. The data mined can provide a reference for follow-up research.
1. Intraducation
Daphniopsis tibetana Sars, 1903 belongs to Daphniidae and is a rare saltwater cladoceran [1] that is mainly distributed in high-altitude areas such as Tibet, Qinghai, and Xinjiang in China. D. tibetana preferentially live in low temperatures. Moreover, they are well-suited for living at high altitudes in cold and nutrient-poor saline water bodies. D. tibetana also plays an important role in the study of cladoceran biology [2].
With the rapid development of mariculture, the demand for live farmed animals is increasing. China and other countries have conducted several series of studies on the cultivation and domestication of cladocerans. Among them, a euryhaline species of cold water, D. tibetana is suitable for the water temperature conditions used during the nursery period of northern marine fish and shrimp. Its low temperature tolerance makes D. tibetana preferable to warm-adapted species that need to be raised for cultivation in high-temperature environments. D. tibetana has a longer developmental period and lower fecundity than Moinidae and Daphnia but higher fecundity than marine copepods and marine zooplankton [3]. Additionally, the amino acid composition of D. tibetana can fully meet the essential amino acid needs of most marine and freshwater fish and shrimp, and the contents of certain unsaturated fatty acids in the body are even higher than those of many common species, such as Moina mongolica, Moina spp., Tigiopus japonica, Brachionus plicatilis, and Artemia.
To date, there have been reports on the influence of D. tibetana morphology and structure [4], living habits [5], ecological distribution [3], and classification and evolution [6], and on how environmental factors influence D. tibetana population growth and physiological metabolism [7,8]. However, there has been no research on the biology of D. tibetana from three lakes in Tibet (Lake Namukacuo, NMKC; Lake Pengcuo, PC; and Lake Zigetangcuo, ZGTC) or on the genetic difference between wild-type and seawater-domesticated D. tibetana. This article reports on and compares some biological observations of D. tibetana from these three locations that were domesticated indoors to enrich the biological data on D. tibetana. This information can be used for in-depth study of indoor seawater domestication and large-scale cultivation of D. tibetana.
2. Materials and Methods
2.1. Source and Domestication of Test Animals
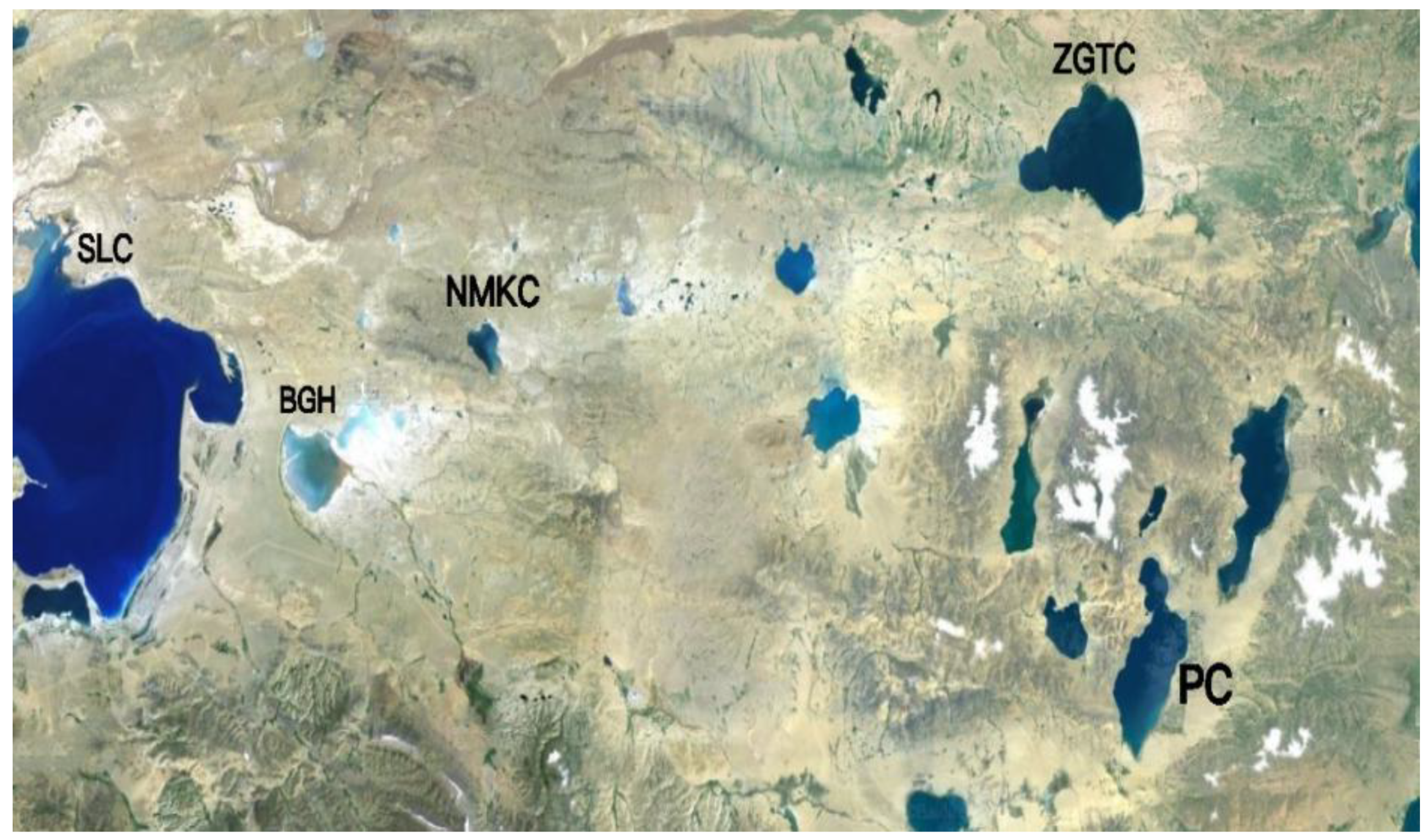
Daphniopsis tibetana is an inland saline cladoceran that is widely distributed in saltwater lakes in Tibet and Qinghai, China, including NMKC (31°83′ N, 89°79′ E), PC (31°89′ N, 90°95′ E), and ZTGC (32°00′ N, 90°90′ E) (Figure 1).
The D. tibetana used in the experiment were collected from NMKC, PC, and ZGTC in Tibet in October 2018. Daphniopsis tibetana were brought back to the laboratory and domesticated in diluted seawater with a salinity of 15–16 ppt at 15 ± 0.5 °C and fed with Chlorella pyrenoidosa. To avoid the negative impact of individual differences on the experiment, observation of life history started with the larval-stage of D. tibetana of the same maternal line. We isolated one gravid mother prior to the experiment and only used its offspring. For the process of domestication, pure water was added to sterilized seawater to adjust the salinity, and the seawater was diluted to 15~16 as culture water. The room temperature was controlled by air conditioning, and the temperature was adjusted to 15 ± 0.5 °C. The Chlorella pyrenoidesa was used as feed with one daily feeding. The culture medium was not replaced during the observation period.
2.2. Experimental Design
Some 30 larval D. tibetana cultured under seawater domestication conditions during the same period were selected for experimentation. One larval D. tibetana each was placed in a 16-mL test tube. During the experiment, its death, molting, time of first birth, interval between births, and number of births were observed and recorded. The experiment was conducted until the D. tibetana died.
2.3. Data Analysis
The population growth parameter was calculated by the following formula:
In the formula, x is the age period (d), lx is the survival rate at stage x (%), mx is the birth rate at stage x, rm is the intrinsic growth rate (d−1), R0 is the net reproductive volume (individual), T is the generation period (d), and λ is the weekly growth rate (d−1).
Microsoft Excel 2010 was used to process test data, and IBM SPSS Statistics 23 (IBM Corporation, Armonk, NY, USA) was used for one-way ANOVA and Duncan’s multiple comparison test to test for significance and variance homogeneity. The arithmetic mean of replicate groups was taken and expressed as mean ± standard deviation; p < 0.05 indicated significant difference; p < 0.01 indicated extremely significant difference.
The growth rate of body length was calculated as follows:
In the formula, LGR is the growth rate body length (%), L0 is the initial body length of the D. tibetana used in the experiment, and L1 is the body length at the time of death in the experiment.
2.4. Transcriptome Sequencing
Approximately 400 D. tibetana were randomly selected from each treatment group as a biological replicate. Total RNA was extracted from D. tibetana using TRIzol (Invitrogen, Carlsbad, CA, USA), and DNase I (TaKara, Dalian, China) was used to remove gene DNA. Using a 2100 Bioanalyzer (Agilent, Santa Clara, CA, USA), the concentration and purity of the extracted RNA were detected by ND-2000 microspectrophotometer (Thermo Scientific, Wilmington, DE, USA) to ensure the integrity of all RNA samples (OD260/OD280 = 1.8–2.2, OD260/OD230 ≥ 2.0, RIN ≥ 8.5, 28S/18S ≥ 1.0) and perform transcriptome sequencing. The mRNA sequencing was conducted using the HiSeq platform, and library construction was performed using the Illumina TruSeq™ RNA sample prep kit method as follows: total RNA was extracted (>1 μg), and mRNA was then enriched, fragmented, and inverted into cDNA; then, adapter ligation and illumina sequencing were performed.
2.5. Differentially Expressed Genes (DEGs)
To explore the differential gene expression of wild-type and domesticated D. tibetana, the expression levels of protein-coding genes were calculated by the FPKM method. DEGs were screened, and differential gene expression volcano plots were drawn. Quantitative analysis of gene expression levels was conducted using RSEM (https://deweylab.biostat.wisc.edu/resm/ accessed on 6 June 2019); after obtaining the number of read counts of gene transcripts, DEGseq (http://bioconductor.org/packages/stats/bioc/DESeq/ accessed on 6 June 2019) software was used to analyze gene expression differences between samples. The significance of differential expression was measured by FPKM (fragments per kilobases per million reads) using false discovery rate (FDR) and fold change (FC) as criteria. When a gene exhibited both FDR < 0.05 and |log2FC| > 1, it was considered differentially expressed.
2.6. KEGG Enrichment of DEGs
Functional enrichment analysis of DEGs in different groups was determined using KOBAS (https://kobas.cbi.pku.edu.cn/home.do, accessed on 6 June 2019) leverage of the KEGG database. Genes were classified according to the pathways they participate in or the functions they perform, and the biological processes most relevant to biological phenomena were identified. The Benjamini and Hochberg method was used for multiple test correction, with p ≤ 0.05 indicating that there was significant enrichment in the GO enrichment function or KEGG pathway.
3. Results and Analysis
3.1. Comparison of Survival Rate and Survival Time of D. tibetana from Different Origins
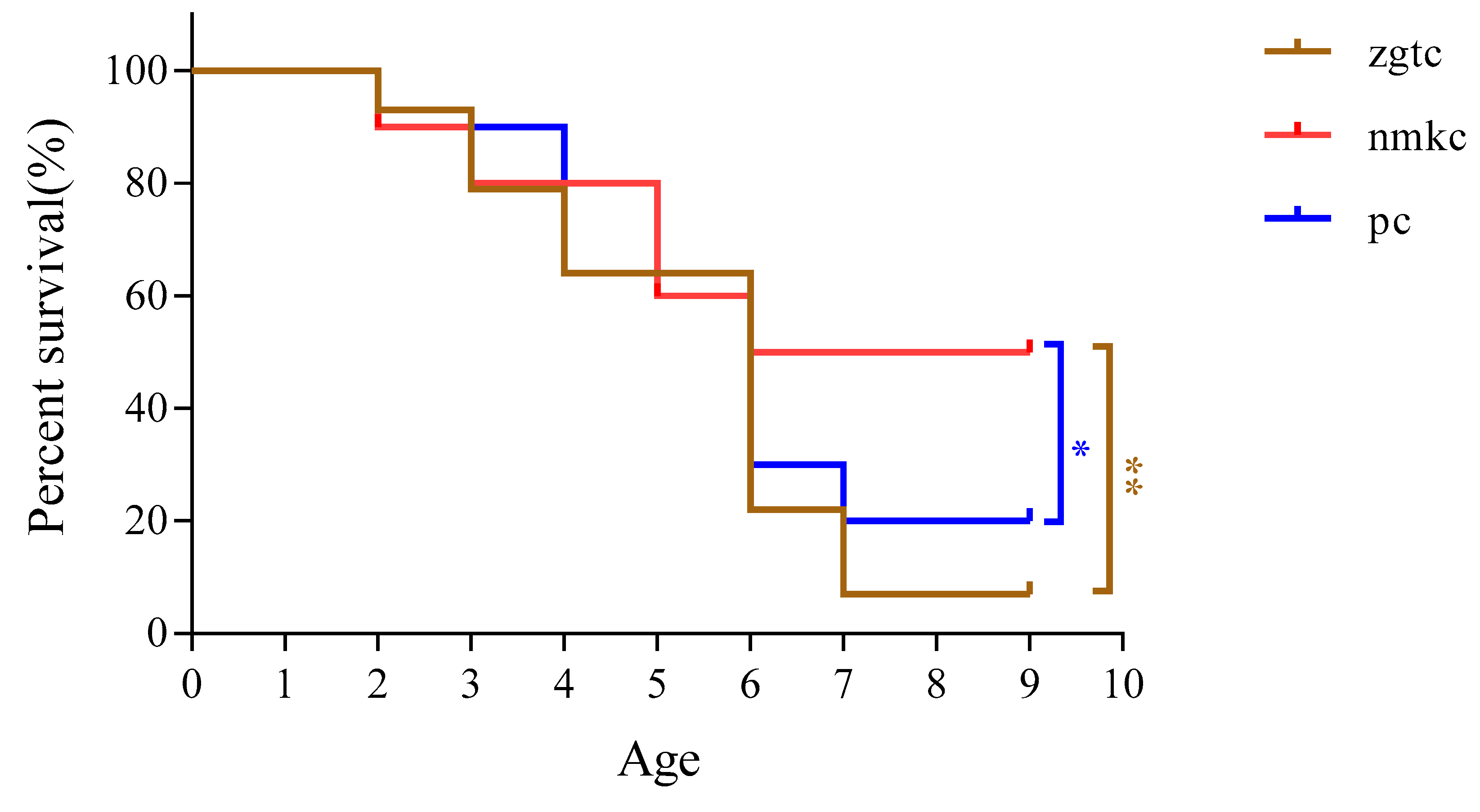
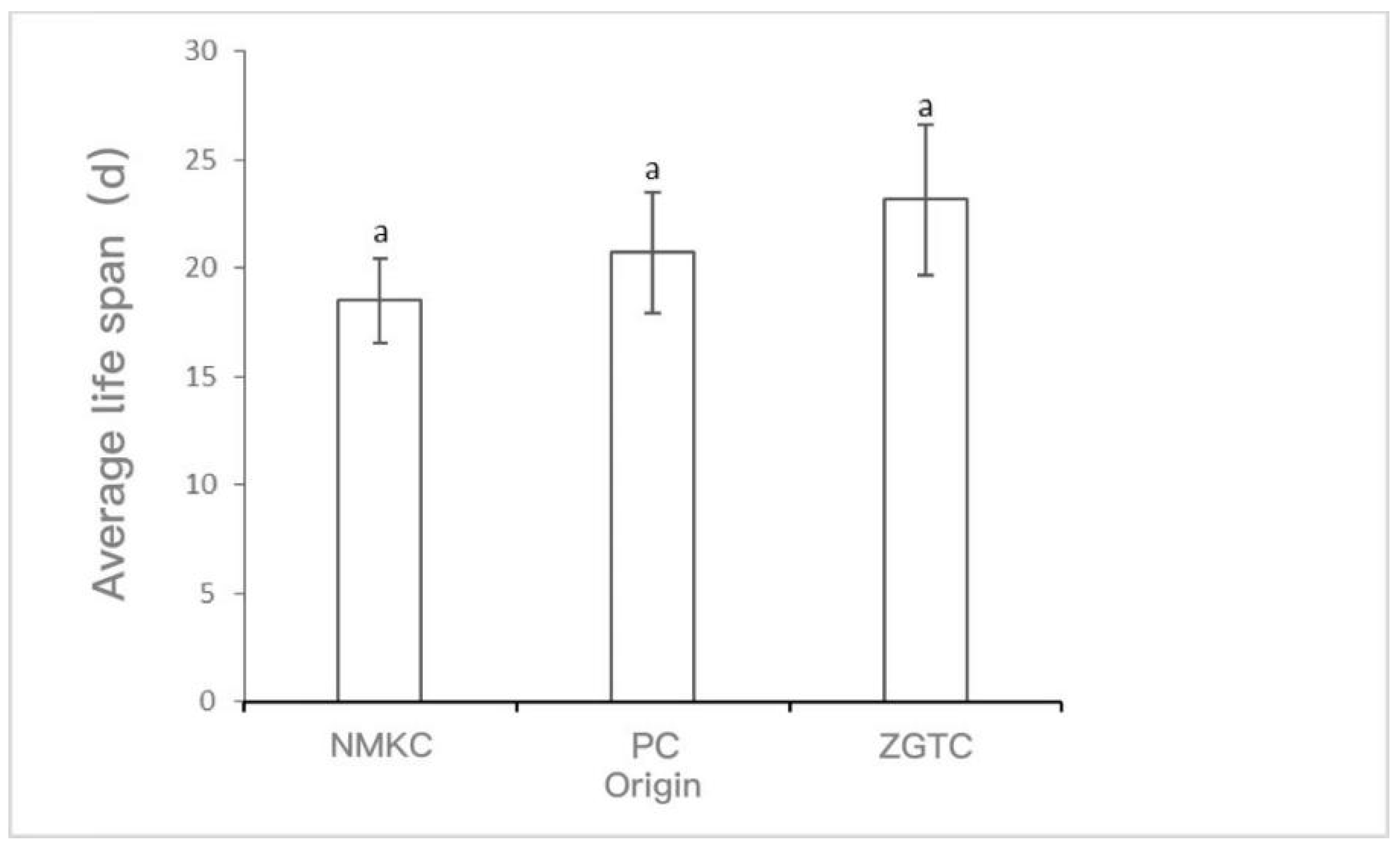
The survival rate and survival time of D. tibetana that originated in each area are shown in Figure 2 and Figure 3. The survival rate of D. tibetana from the three areas dropped rapidly after the fifth instar (Figure 2). In general, the survival rate of the NMKC strain was higher than those from the other two areas. However, the NMKC strain had the shortest survival time and failed to complete the seventh instar. This was followed by the PC strain and finally the ZTGC strain, which had the longest survival time and for which there were still survivors after the ninth instar. The longest survival time of each strain was 26 d for NMKC, 35 d for PC35, and 53 d for ZGTC (Figure 3).
3.2. Comparison of Growth and Reproduction of D. tibetana from Different Origins
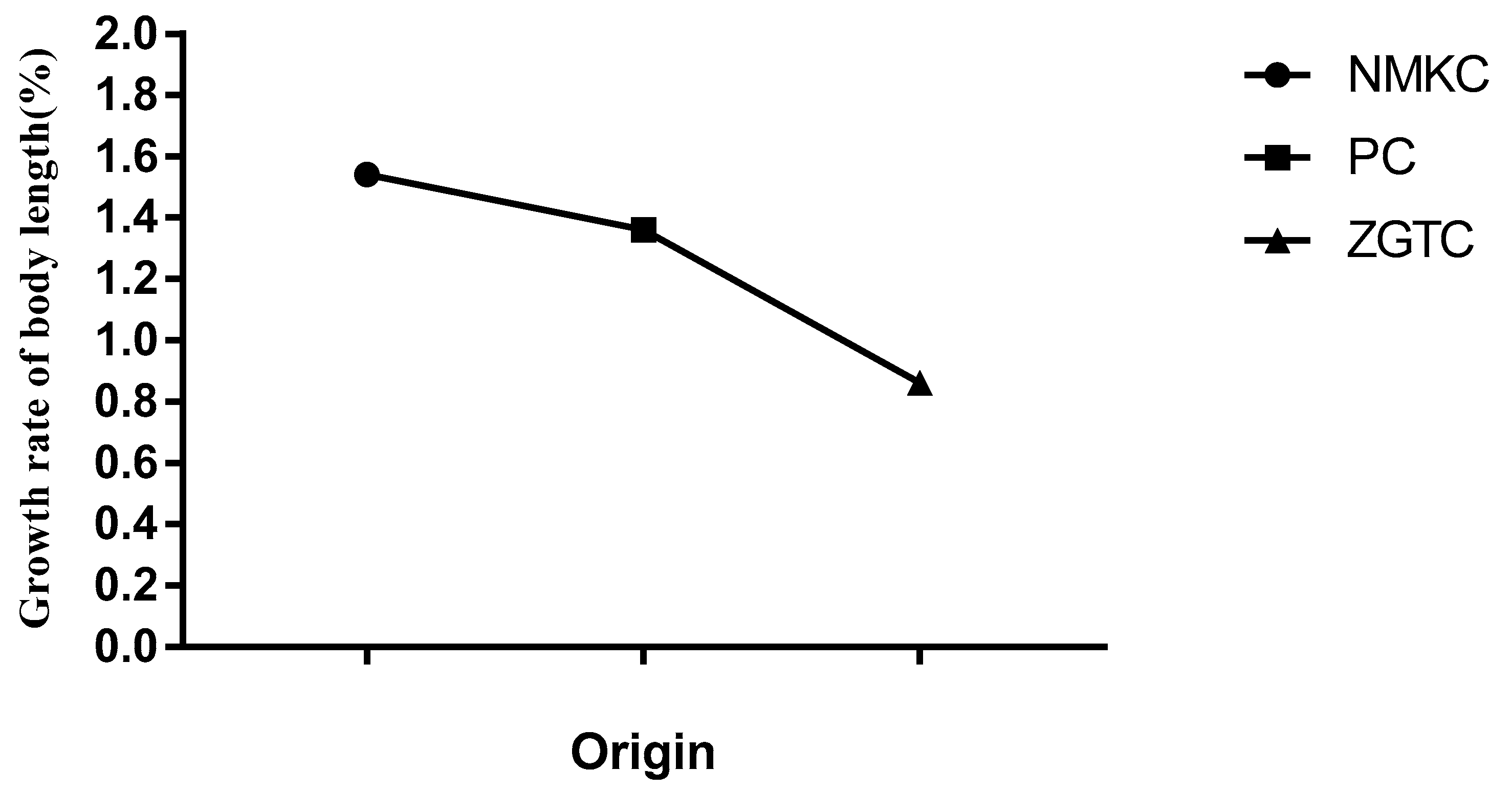
Growth rates of the NMKC and PC strains were significantly higher than that of the ZGTC strain (p < 0.05; Figure 4). The body lengths of the NMKC, PC, and ZGTC strains increased by 1.21 ± 0.91 mm (growth rate, 153.6 ± 12.1%), 1.17 ± 0.13 mm (growth rate, 136.4 ± 16.1%), and 0.70 ± 0.07 mm (growth rate, 86.2 ± 7.6%).
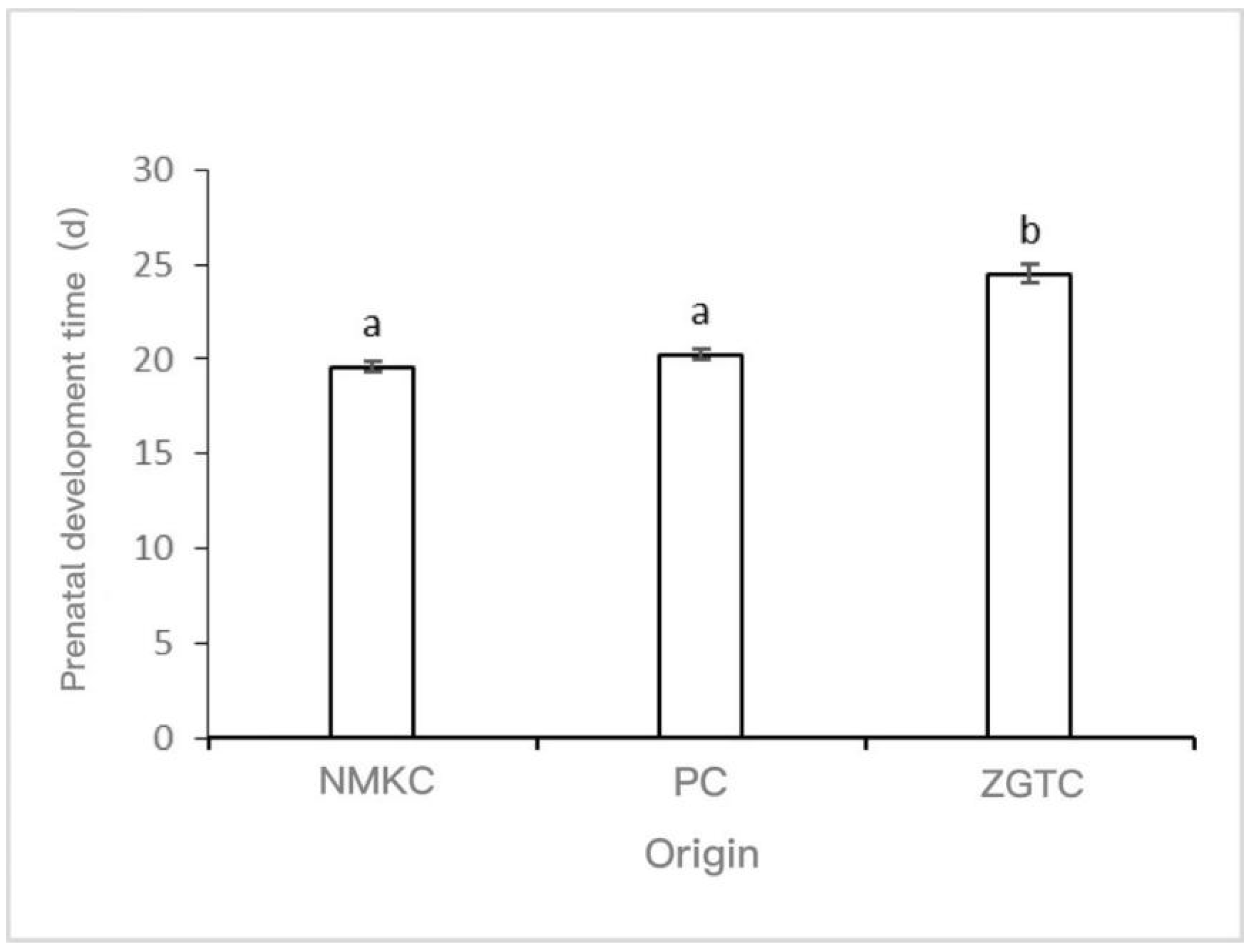
Under the conditions of laboratory seawater acclimation (15 ± 1 °C), the NMKC strain had the shortest prenatal development period (19.6 ± 0.25 d), followed by the PC strain (20.25 ± 0.25 d), and the ZGTC strain had the longest prenatal development period (24.5 ± 0.5 d) (Figure 5). The prenatal development period of the ZGTC strain was significantly different from those of the other two strains (p < 0.05).
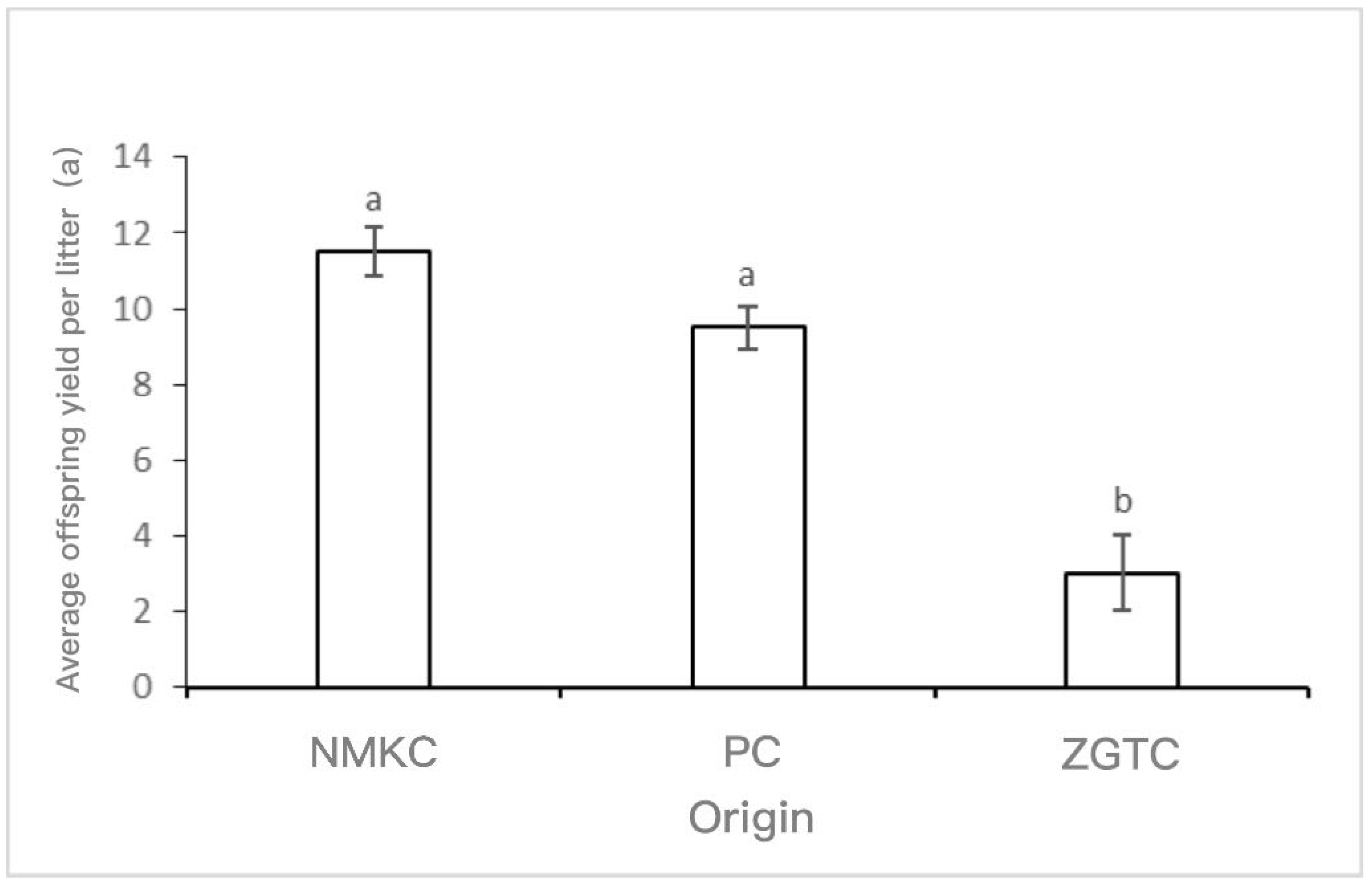
The average number of offspring per litter for the NMKC and PC strains was higher(p < 0.05) than that of the ZGTC strain (p < 0.05) (Figure 6). Among them, the NMKC strain had the most offspring per litter (11.5 ± 0.65), followed by the PC strain (9.5 ± 0.57), and then the ZGTC strain (3.0 ± 1.0).
3.3. Comparison of Population Growth Parameters of D. tibetana from Different Origins
The intrinsic growth rate and net reproductive capacity of the NMKC and PC strains were significantly higher than those of the ZGTC strain (p < 0.05), and the generation cycle of ZGTC was significantly longer than those of the other two strains (p < 0.05) (Table 1). However, the weekly growth rates of the three strains were not significantly different (p > 0.05).
3.4. Embryo Development
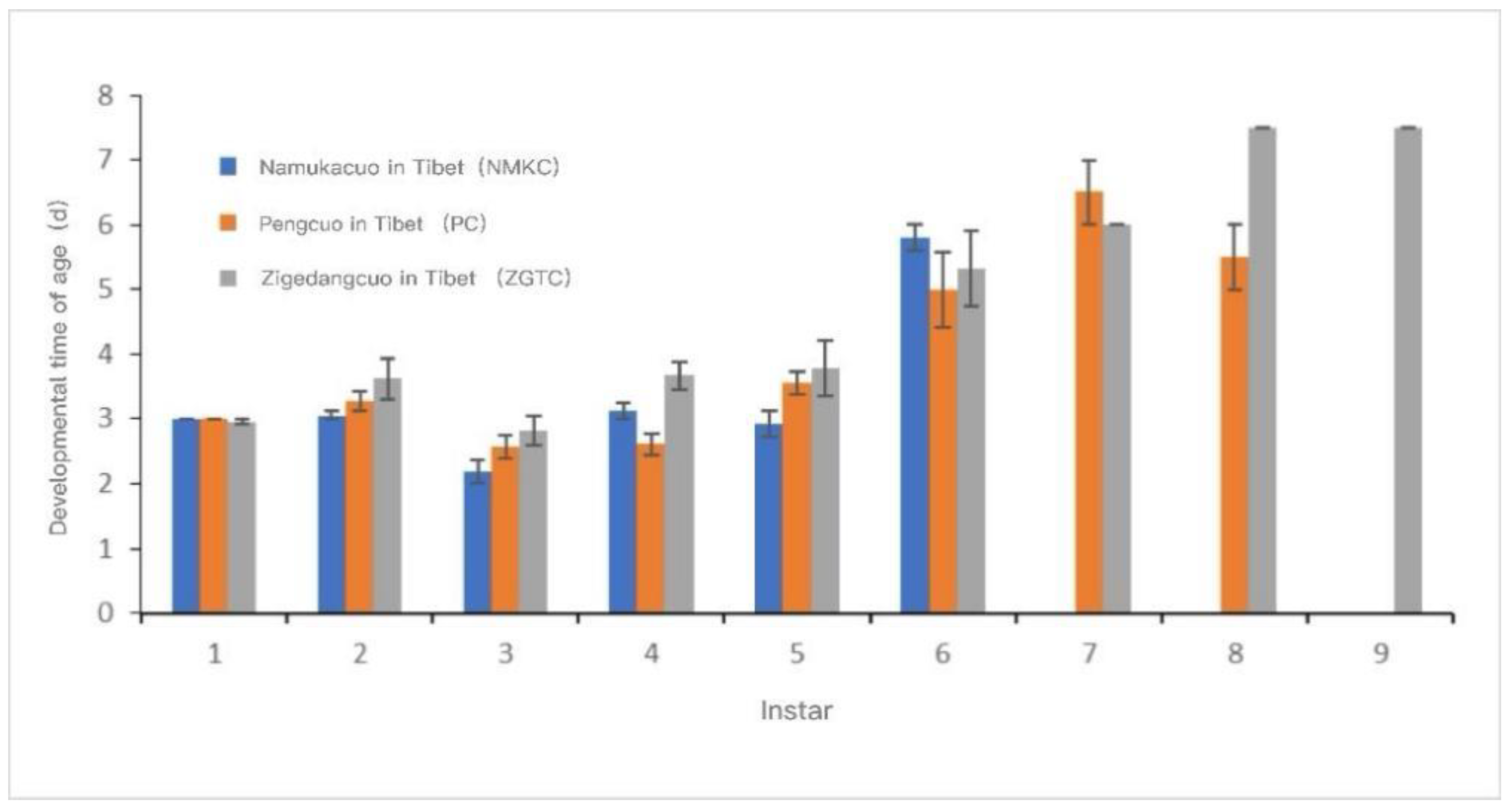
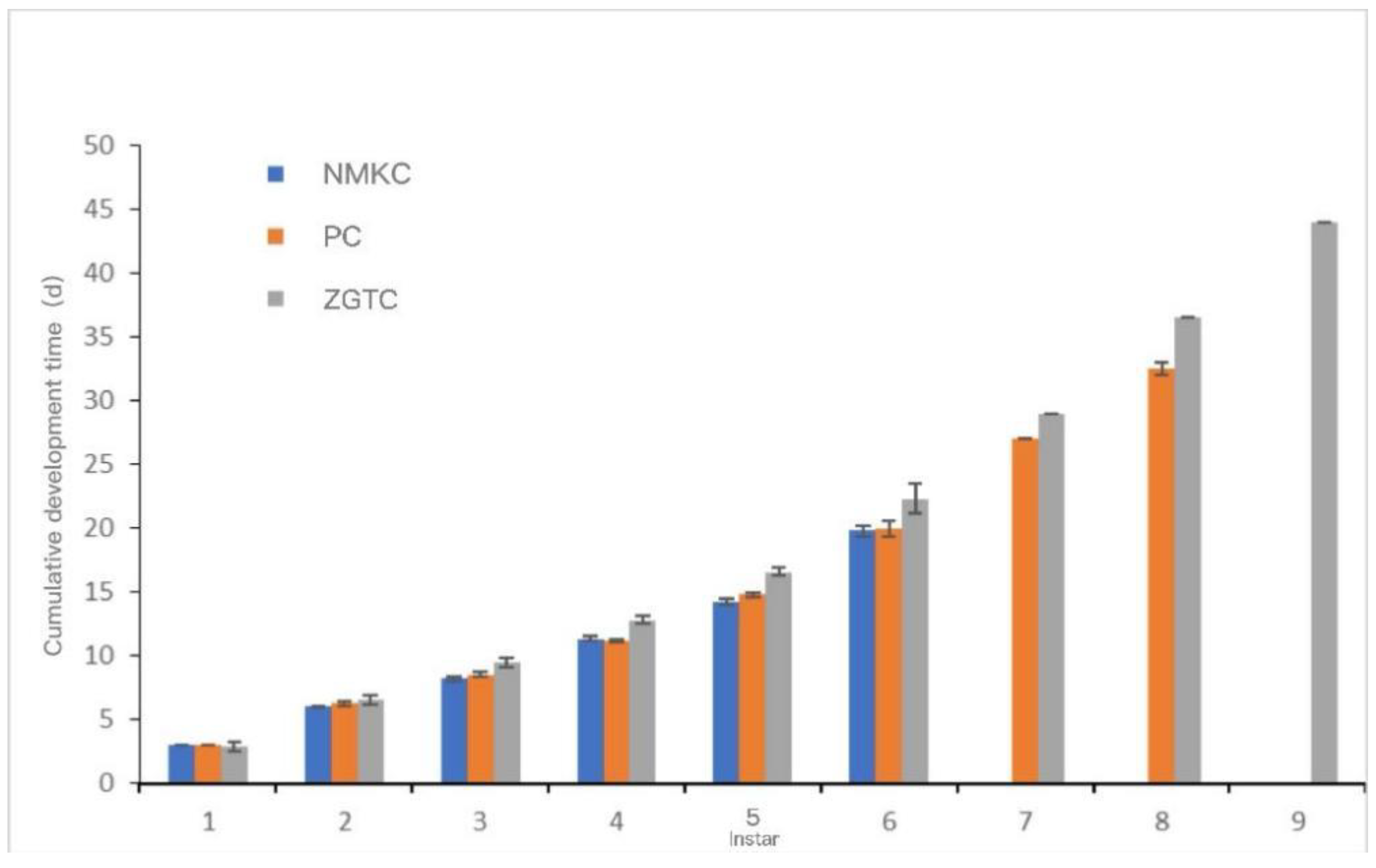
The development time and cumulative development time of each instar of each D. tibetana strain are shown in Figure 7 and Figure 8. In general, the three D. tibetana strains developed to the sixth instar. Among them, the NMKC strain had the shortest survival time; this strain survived after the sixth instar, but soon died and failed to complete the seventh instar. The PC strain developed to the eighth instar. ZGTC had the longest development time but failed to complete the 10th instar. The cumulative development time of the ZGTC strain was longer than those of the other two D. tibetana strains.
3.5. Transcriptomic Analysis of D. tibetana
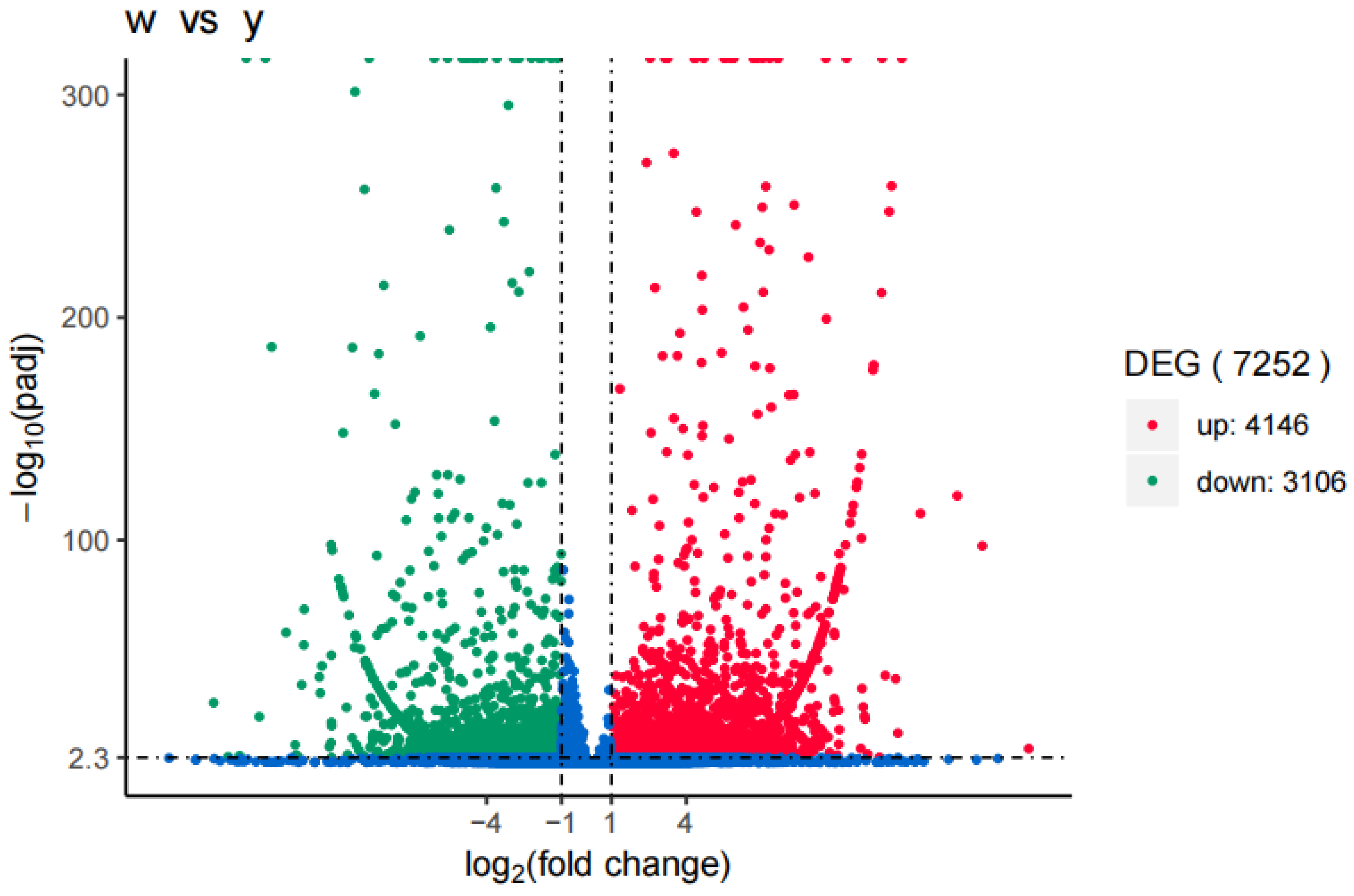
Compared with Y, 7252 differentially expressed genes were generated after acclimation with seawater, among which 4146 were up-regulated and 3106 were down-regulated. The results indicated that D. tibetana had significant gene expression difference after acclimation in seawater(Figure 9).
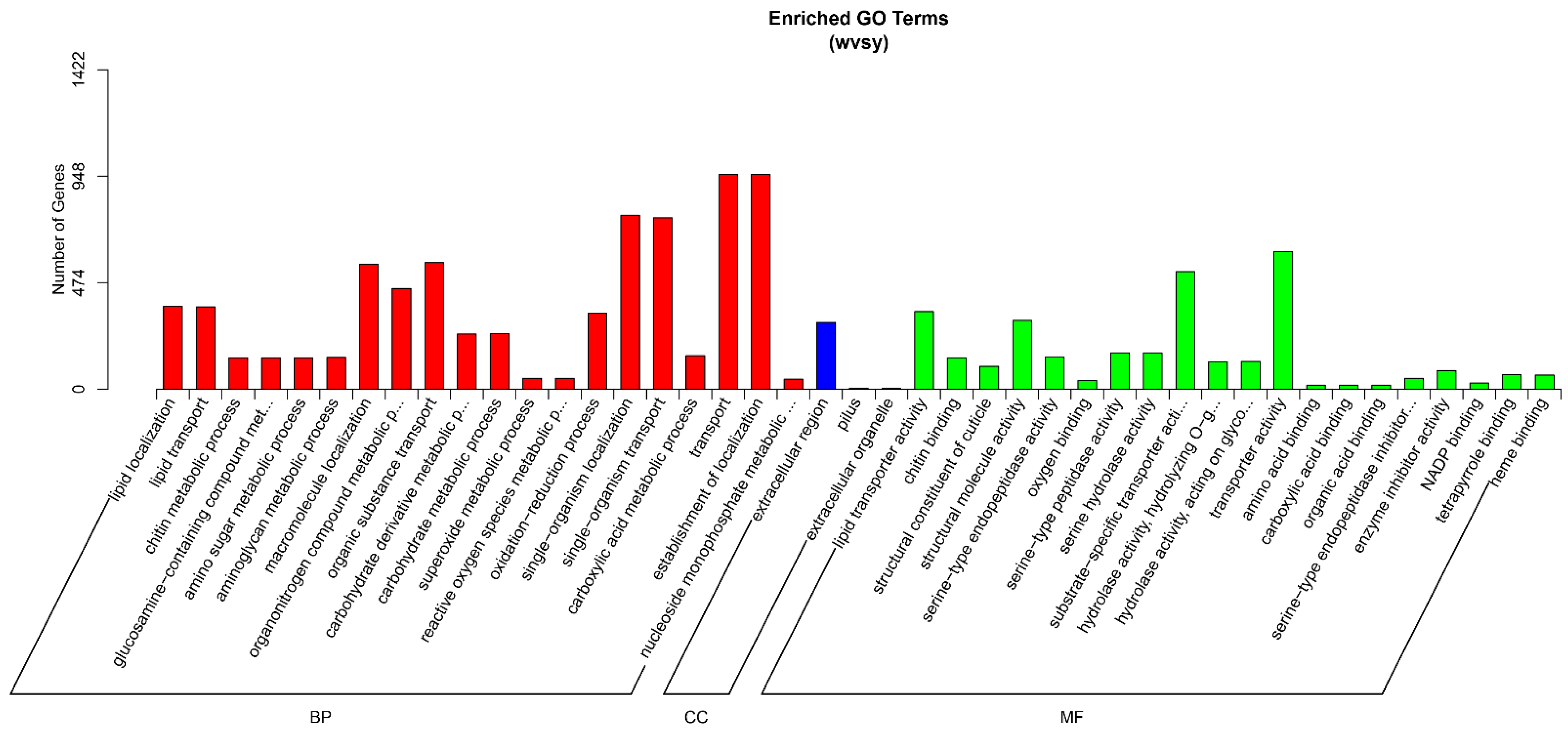
Results of GO enrichment analysis performed on the DEGs of wild-type and domesticated D. tibetana are shown in Figure 10. The DEGs between wild-type and domesticated D. tibetana were mainly enriched in four biological process terms (establishment of localization, transport, single-organism operation, and single-organism localization), one cellular component term (extracellular region), and two molecular function terms (substrate-specific transporter activity and transporter activity).
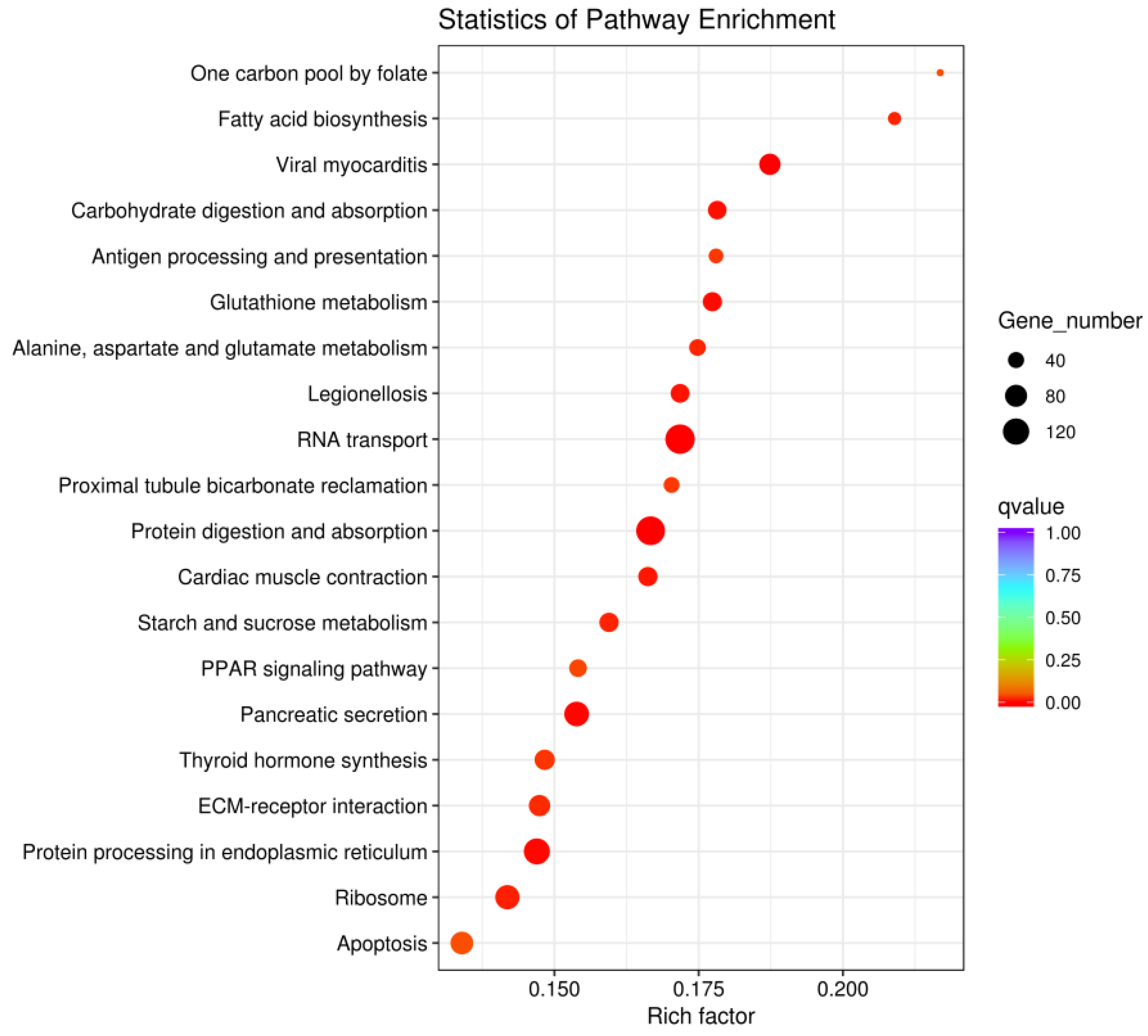
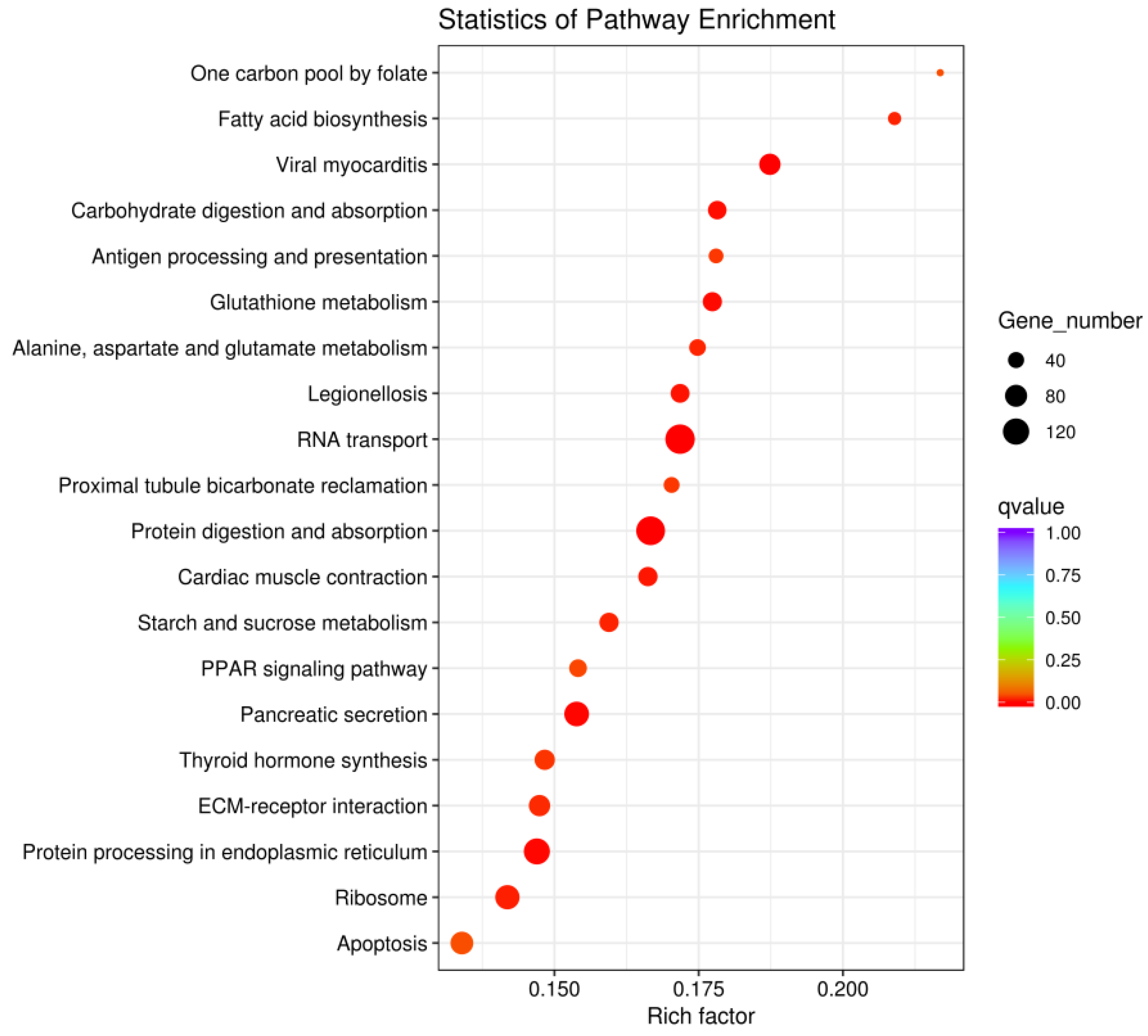
Compared with wild-type D. tibetana, domesticated D. tibetana had higher enrichment of RNA transport, protein digestion and absorption, and protein processing in endoplasmic reticulum pathways (Figure 11).
4. Discussion
ZGTC salinity is greater and the water body larger than that of PC, although the ecological environments are similar, and D. tibetana is less dense than in NMKC. The salinity of NMKC is between 15–25 ppt, and the composition of salt ions is different from that of other lakes because of its unique geographical location and water environment characteristics; this could be why the D. tibetana biomass is greater in NMKC.
A previous study on the salt lakes in northern Tibet revealed that the fish biomass in these salt lakes is low, and D. tibetana has become the main food for some water birds in Tibet [9]. Because most of the salt lakes are not connected, the inhabiting activities of birds may be the main reason that D. tibetana can be distributed in each salt lake even though the water ecosystem of each salt lake is different and there is a certain amount of geographical isolation. This may be the main reason why D. tibetana formed different strains.
Different geographical populations of the same Daphnia species must adapt to the specific ecological environment of their habitat; therefore, certain interspecies differences occur. In May and July 2001, Zhao [10] investigated the biological and ecological characteristics of 22 lakes in northern Tibet; the lake salinities ranged between 1 and 390 ppt, and 95 taxa phytoplankton and 42 zooplankton taxa were recorded. Moreover, Na+ and Mg2+ were the main cations in lake water; however, CO32−was the dominant anion under low salinity, whereas Cl− was the dominant anion with increasing salinity. This is consistent with the results of our laboratory’s investigation in a few of Tibet’s salt lakes in September 2018. Therefore, this experiment used the optimum temperature (15 °C) and salinity (15 ppt) for D. tibetana survival and growth to further explore the dynamic changes of D. tibetana seasonal populations in three different areas [9].
Under certain environmental conditions, a change in the intrinsic growth rate of a population can reflect small changes in the environment and is an important indicator of the reproductive ability of a species [11,12].This study found that there was no significant difference in the D. tibetana intrinsic growth rate, weekly growth rate, and generation cycle between the NMKC and PC strains, but the net reproductive capacity of NMKC was significantly less than that of PC. This is because individuals of the NMKC strain gave birth only once during their entire life cycle, whereas individuals of the PC strain gave birth more than once. However, the ANOVA results for the experimental data of these two strains showed that the average prenatal development period, average reproductive volume per litter, and growth rate of body length were not significant (p > 0.05). This finding shows that the D. tibetana of NMKC and PC may be the same geographic population. However, on average, the prenatal development period of the NMKC strain was shorter, and the average reproductive capacity per individual was the largest. This may be because the water used in this experiment was closer to the salinity of NMKC and had less impact on this strain. Compared with the other two groups, ZGTC had obvious differences in intrinsic growth rate and net reproductive capacity; this may result from the geographical isolation and salinity changes having important impacts on D. tibetana biology.
From the perspective of salinity, the three lakes are all inland salt lakes; however, the populations of these cladocerans in different areas have very different adaptability to salinity domestication [13].The salinity of NMKC and PC are both 16 ppt, whereas that of ZGTC is 21 ppt. Under the experimental conditions, the salinity used was closer to that of NMKC and PC; therefore, compared with the ZGTC strain, the NMKC and PC strains had the characteristics of shorter prenatal development period and larger average reproductive volume per individual. However, because of the dry climate in Tibet, slow changes in salinity during the evaporation and concentration of water also play a natural role in domesticating aquatic organisms.
There is little difference between the pH values of NMKC, PC, and ZGTC (9.54, 9.86, and 10.06, respectively). Moreover, the temperatures of NMKC, PC, and ZGT Care 13 °C, 16.5 °C, and 11.5 °C, respectively, and the control temperature in this experiment (15 ± 0.5 °C) was closer to NMKC and PC. Zhao [14] noted that geographical isolation and salinity changes have important impacts on the genetic diversity of D. tibetana from different water bodies. Additionally, Wang [7] found that there were obvious interspecies differences caused by geographical isolation. This study compared the distribution of D. tibetana in Tibet with some biological observations of indoor domesticated strains and further confirmed that there are differences in genetic diversity among different geographic populations of D. tibetana. However, this difference cannot be attributed simply to geographical isolation. It may be that in Tibet, D. tibetana has genetic diversity differences that result from long-term adaptation to different ecological factors. This difference is based mainly on what factors directly or indirectly affect the organisms and can be used to identify differences among different geographic populations.
In addition, 7252 DEGs were identified based on the third-generation transcriptome sequencing data of wild-type and domesticated D. tibetana that were analyzed in the laboratory. After D. tibetana was moved from the wild to the laboratory, numerous DEGs were generated. Significant enrichment of GO terms revealed that the DEGs are mainly involved in molecular functions, such as substrate-specific transporter activity and transporter activity, and they are mainly located in the cellular components of the extracellular region. Moreover, the majority of DEGs were associated with biological processes and were enriched in the establishment of localization, transport, single-organism operation, and single-organism localization. In the KEGG pathway enrichment analysis of DEGs, the RNA transport pathway, protein digestion and absorption pathway, and protein processing in endoplasmic reticulum pathway were highly enriched. Through these annotations, a large amount of wild-type and domesticated D. tibetana transcriptome information, which can more effectively help us understand the genetic characteristics of D. tibetana at the molecular level, was obtained. This is of great significance for further exploration of gene function in the future and provides basic data for exploring the functional genes related to D. tibetana resistance to environmental stress and studying related physiological functions.
5. Conclusions
Under laboratory domestication at a temperature of 15 ± 0.5 °C and a salinity of 15–16 ppt, the ZGTC strain had the longest life span, but the NMKC and PC strains had significantly higher growth rates of body length than the ZGTC strain (p < 0.05).The prenatal development period of the NMKC strain was the shortest (19.6 ± 0.25 d), but the average number of offspring per litter was the largest (11.5 ± 0.65). The intrinsic growth rate and net reproductive capacity of the NMKC and PC strains were significantly higher than those of the ZGTC population (p < 0.05). Three generations of transcriptome sequencing of wild-type D. tibetana after it was moved from the wild to the laboratory were performed in the laboratory, and correlation analysis was performed on the determined DEGs. In total, 7252 DEGs were generated in the comparison between wild-type and domesticated D. tibetana after seawater domestication, of which 4146 were up-regulated and 3106 were down-regulated. After seawater domestication, a series of biological processes and related genes in D. tibetana cells were affected. In GO enrichment analysis, the DEGs were mainly enriched in four biological process terms (establishment of localization, transport, single organism operation, and single organism localization), one cellular component term (extracellular region), and two molecular function terms (substrate-specific transporter activity and transporter activity. In KEGG pathway enrichment analysis, the DEGs were highly enriched in the RNA transport pathway, protein digestion and absorption pathway, and protein processing in the endoplasmic reticulum pathway.
Author Contributions
Conceptualization, W.Z. (Wan Zhang) and J.Z.; methodology, W.Z. (Wen Zhao); software, W.Z. (Wan Zhang); validation, J.W., S.W. and D.Y.; formal analysis, J.Z.; investigation, W.Z. (Wen Zhao); resources, W.Z. (Wen Zhao); data curation, W.Z. (Wen Zhao); writing—original draft preparation, W.Z. (Wan Zhang); writing—review and editing, J.Z.; visualization, W.Z. (Wan Zhang); supervision, W.Z. (Wen Zhao); project administration, J.W.; funding acquisition, W.Z. (Wen Zhao). All authors have read and agreed to the published version of the manuscript.
Funding
The paper was supported by National Key R&D Plan Blue Granary Science and Technology Innovation Project (grant No.2020YFD0900200).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Because we still have a lot of follow-up studies going on, the data is not available.
Conflicts of Interest
The authors declare no conflict of interest.
References
- He, Z.H.; Zhao, W. Biological resource in inland saline waters in North China. J. Dalian Fish. Univ. 2002, 3, 157–166. [Google Scholar] [CrossRef]
- Jiang, X.Z.; Du, N.S. Zoology of China (Arthropod Crustacea Freshwater Cladocera); Science Press: Beijing, China, 1979; pp. 122–124. [Google Scholar]
- Wen, Z.; Qiaohan, W.; Mianping, Z.; Yuanyi, Z.; Hailei, W. A preliminary study on the biology of Daphniopsis tebitana Sars. J. Dalian Fish. Univ. 2002, 17, 209–214. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, Q.H. The morphological redescription of Daphniopsis tebitana Sars. J. Dalian Fish. Univ. 2005, 20, 165–173. [Google Scholar] [CrossRef]
- Manca, M.; Carnmarano, P.; Spagnuolo, T. Notes on Cladocera and Copepoda from high altitude lakes in the Mount Everest Region (Nepal). Hydrobiologia 1994, 287, 225–231. [Google Scholar] [CrossRef]
- Zhao, W.; Li, R. Molecular phylogeny of four strains of Daphniopsis tibetana Sars based on mitochondrial 12S rRNA gene sequences. J. Dalian Ocean. Univ. 2012, 27, 300–305. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, W.; Wei, J.; Wang, S.; Xie, X. Acute effects of UVB radiation on the survival, growth, development, and reproduction of Daphniopsis tibetana Sars (Crustacea: Cladocera). Environ. Sci. Pollut. Res. Int. 2019, 26, 10916–10925. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Huo, Y.; Zhang, T.; Wang, S.; Shi, T. Effects of lithium on the survival, growth, and reproduction of Daphniopsis tibetana Sars (Crustacea: Cladocera). Chin. J. Oceanol. Limnol. 2017, 35, 754–762. [Google Scholar] [CrossRef]
- Zhao, W.; He, Z.H.; Ren, S.R. Biology and Technology of Culture and Utilization in Marine Water for Cladocera in Inland Saline Waters; Science Press: Beijing, China, 2008; p. 163. ISBN 978-7-03-020277-2. [Google Scholar]
- Wen, Z.; Mian-Ping, Z.; Xian-Zhong, X.; Xi-Fang, L.; Gan-Lin, G.; Zhi-Hui, H. Biological and ecological features of saline lakes in northern Tibet, China. Hydrobiologia 2005, 541, 189–203. [Google Scholar] [CrossRef]
- Lin, C.S. Theoretical and laboratory studies of animal population dynamics I. The significance and application of laboratory populations and of mathematical models in the studies of animal population dynamics. Acta Zool. Sin. 1963, 15, 371–381. [Google Scholar]
- Krebs, C.J. Ecology: The Experimental Analysis of Distribution and Abundance, 4th ed.; Harper Collins College Publishers: New York, NY, USA, 1994; pp. 184–186. [Google Scholar]
- He, Z.H.; An, S.S. The adaptability of Moina rectirostris to seawater salinity. Chin. J. Zool. 1986, 2, 25–28. [Google Scholar] [CrossRef]
- Zhao, W.; Huo, Y.; Zhang, T.; Wang, S.; Shi, T. Genetic diversity of four population of cladocera Daphniopsis tebitana. J. Dalian Ocean. Univ. 2011, 26, 108–113. [Google Scholar] [CrossRef]
Figure 1.
Sampling points of each lake. SLC, Selincuo Lake; BGH, Bangehu Lake.
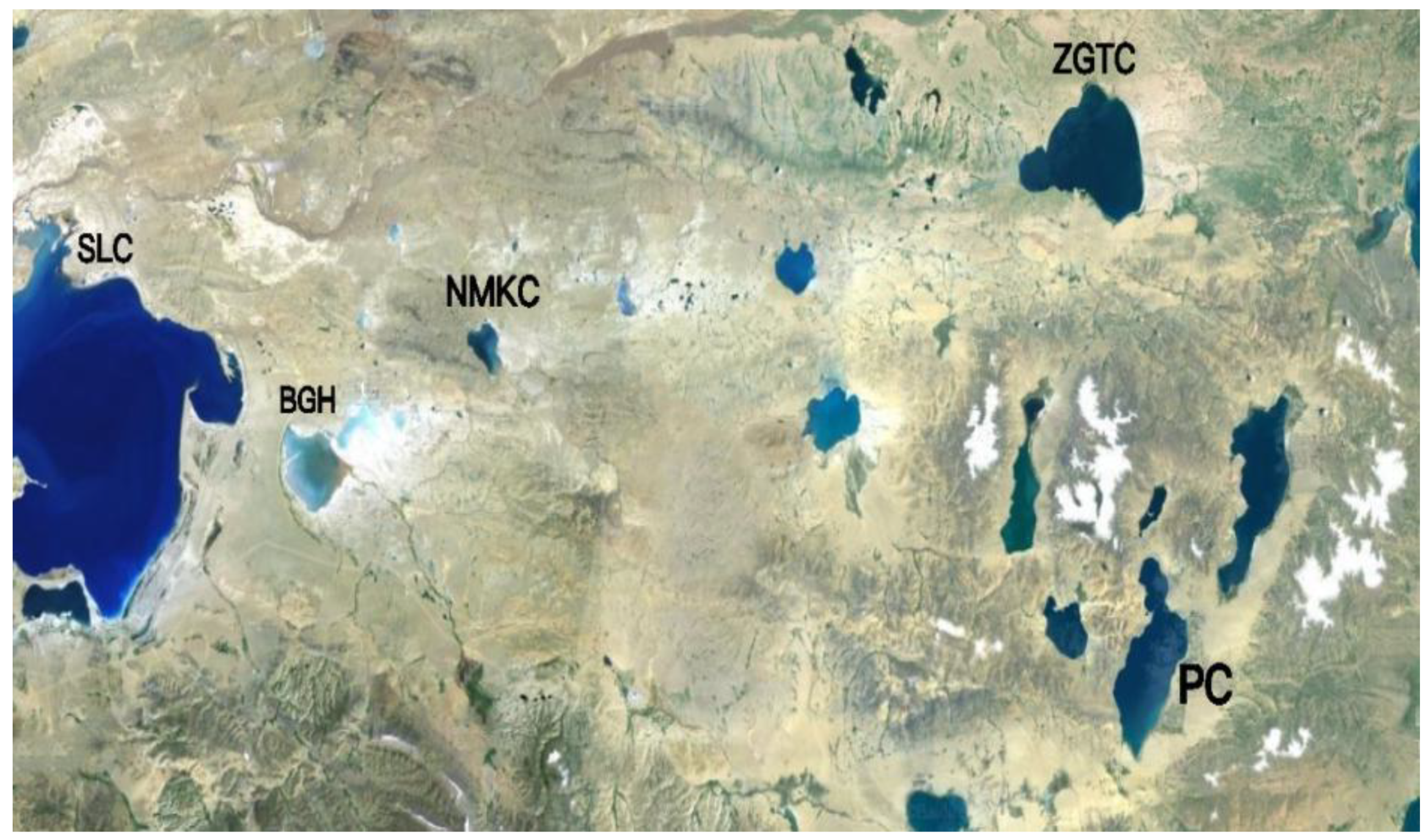
Figure 2.
Survival rates of Daphniopsis tibetana from different origins.*: Have difference; **: Have a significant difference.
Figure 2.
Survival rates of Daphniopsis tibetana from different origins.*: Have difference; **: Have a significant difference.

Figure 3.
Life span of Daphniopsis tibetana from different origins. Means with different lowercase letters are significantly different (p < 0.05). a: no significant difference.
Figure 3.
Life span of Daphniopsis tibetana from different origins. Means with different lowercase letters are significantly different (p < 0.05). a: no significant difference.
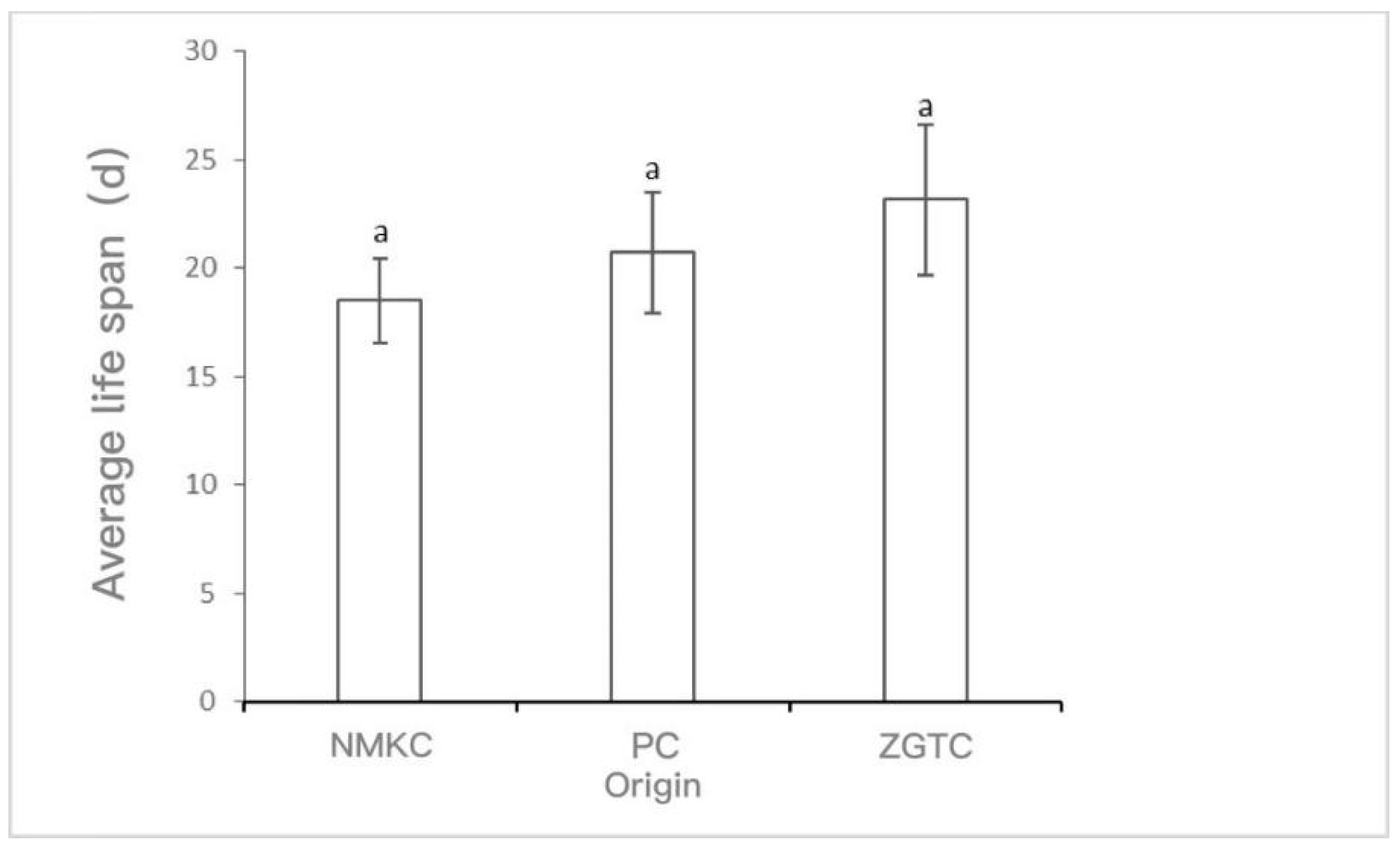
Figure 4.
Growth rate of body length of Daphniopsis tibetana from different origins. Means with different lowercase letters are significantly different (p < 0.05).
Figure 4.
Growth rate of body length of Daphniopsis tibetana from different origins. Means with different lowercase letters are significantly different (p < 0.05).

Figure 5.
Prenatal development time of Daphniopsis tibetana from different origins. Means with different lowercase letters are significantly different (p < 0.05). a: no significant difference; b: significant difference.
Figure 5.
Prenatal development time of Daphniopsis tibetana from different origins. Means with different lowercase letters are significantly different (p < 0.05). a: no significant difference; b: significant difference.

Figure 6.
Average number of offspring per litter of Daphniopsis tibetana from different origins. Means with different lowercase letters are significantly different (p < 0.05). a: no significant difference; b: significant difference.
Figure 6.
Average number of offspring per litter of Daphniopsis tibetana from different origins. Means with different lowercase letters are significantly different (p < 0.05). a: no significant difference; b: significant difference.

Figure 7.
Instar development time of Daphniopsis tibetana from different origins.
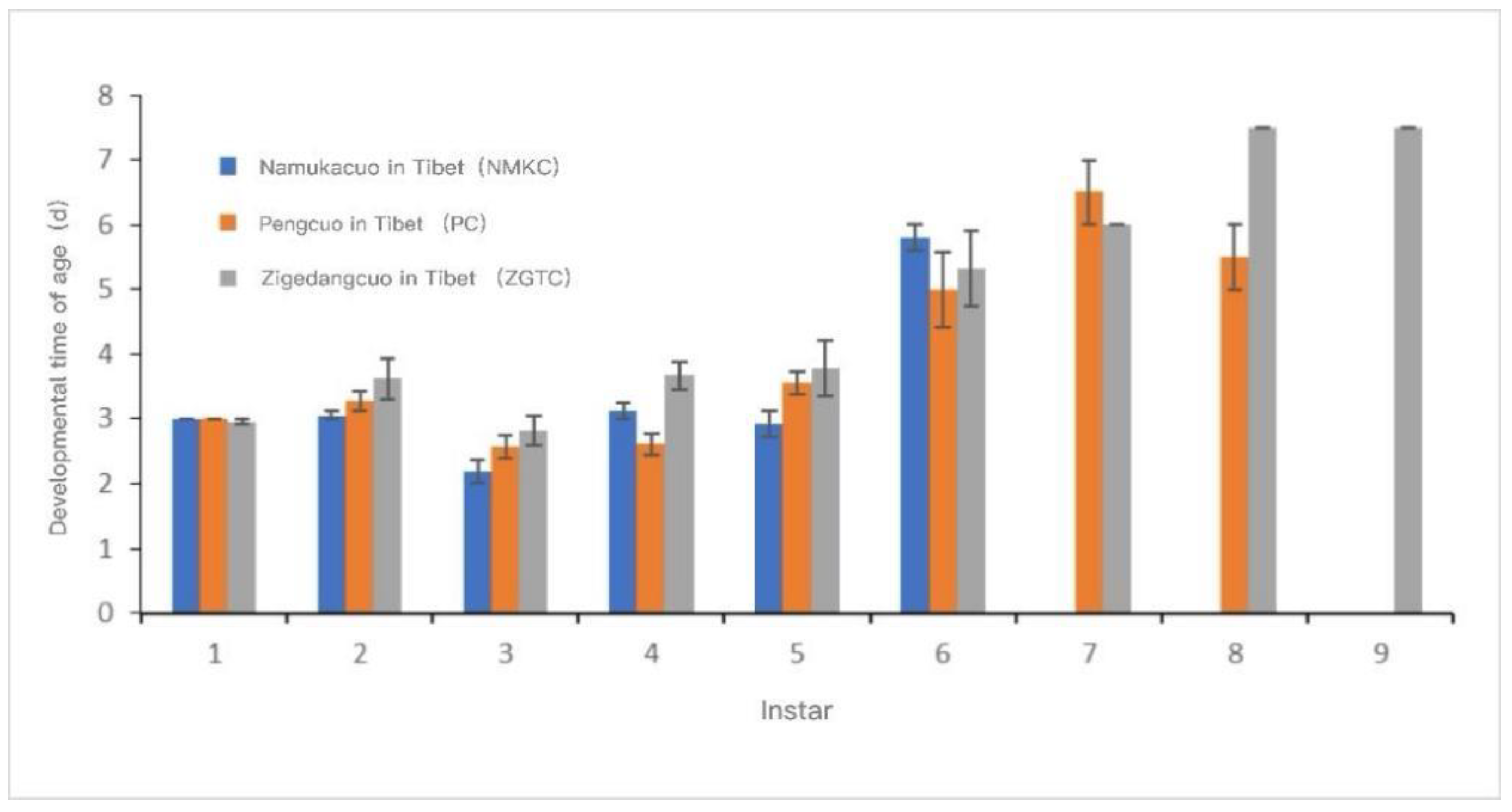
Figure 8.
Cumulative development time of Daphniopsis tibetana from different origins.
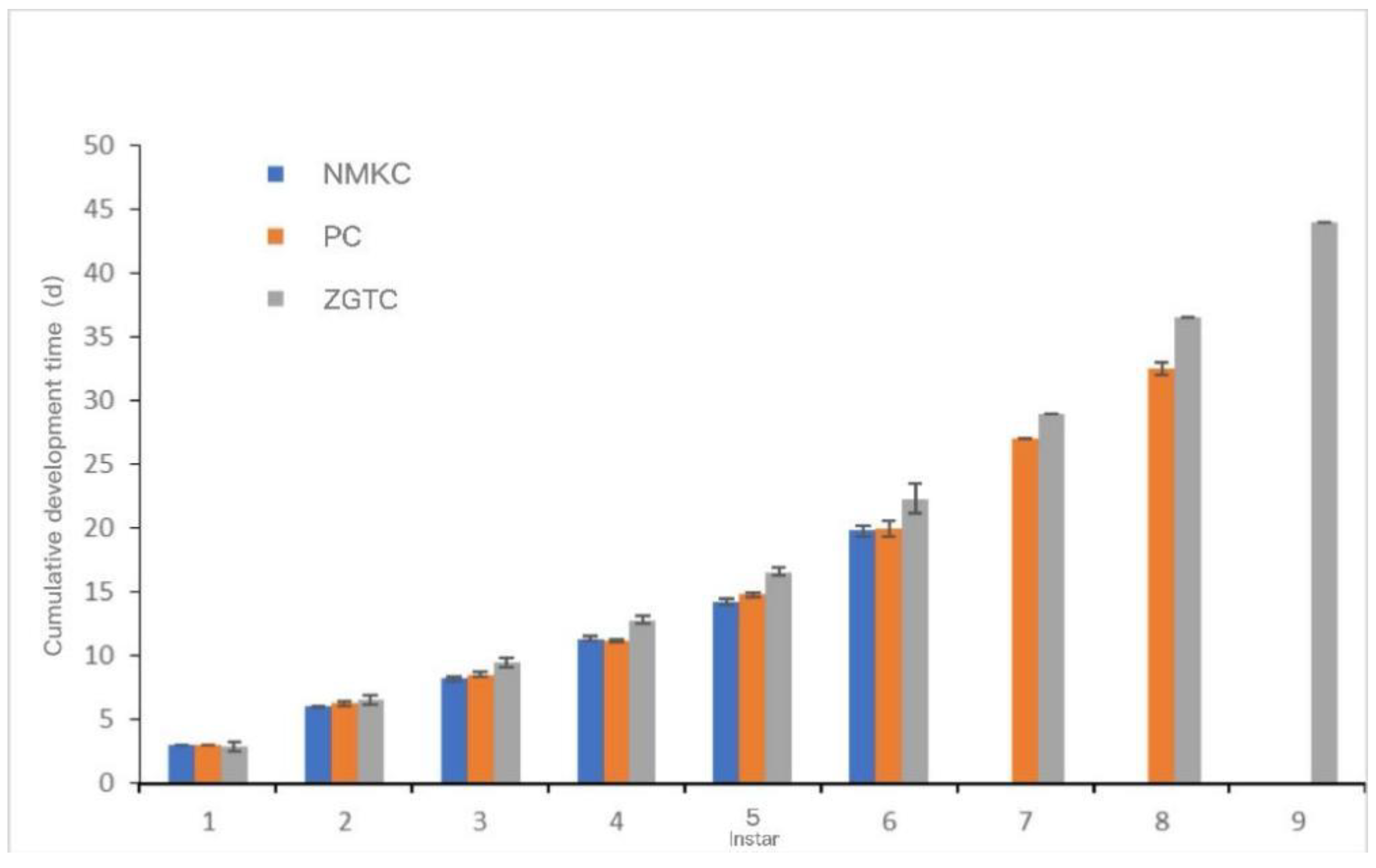
Figure 9.
Gene expression differences between wild-type and domesticated Daphniopsis tibetana.

Figure 10.
GO enrichment analysis of DEGs between wild-type and domesticated Daphniopsis.

Figure 11.
Scatterplot of DEGs between wild-type and domesticated Daphniopsis tibetana enriched in the KEGG pathways.
Figure 11.
Scatterplot of DEGs between wild-type and domesticated Daphniopsis tibetana enriched in the KEGG pathways.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of population growth parameters of Daphniopsis tibetana from different origins.
| Population Growth | Origin | ||
|---|---|---|---|
| Parameter | NMKC | PC | ZGTC |
| Intrinsic growth rate (d−1) | 0.11 ± 0.033 a | 0.13 ± 0.017 a | 0.04 ± 0.021 b |
| Weekly growth rate (d−1) | 1.12 ± 0.036 a | 1.13 ± 0.019 a | 1.04 ± 0.022 b |
| Generation cycle (d) | 19.40 ± 0.55 a | 22.37 ± 2.74 a | 31.01 ± 26.88 a |
| Net reproductive rate (ind.) | 9.20 ± 4.00 a | 19.00 ± 11.83 a | 3.00 ± 1.41 a |
Note: a: no significant difference; b: significant difference.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, W.; Zhao, W.; Zou, J.; Wei, J.; Wang, S.; Yin, D. Comparative Biology of Daphniopsis tibetana from Different Habitats under Seawater Acclimation. Water 2023, 15, 34. https://doi.org/10.3390/w15010034
AMA Style
Zhang W, Zhao W, Zou J, Wei J, Wang S, Yin D. Comparative Biology of Daphniopsis tibetana from Different Habitats under Seawater Acclimation. Water. 2023; 15(1):34. https://doi.org/10.3390/w15010034
Chicago/Turabian StyleZhang, Wan, Wen Zhao, Jingjing Zou, Jie Wei, Shan Wang, and Dongpeng Yin. 2023. "Comparative Biology of Daphniopsis tibetana from Different Habitats under Seawater Acclimation" Water 15, no. 1: 34. https://doi.org/10.3390/w15010034
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.