
Response of Aquatic Plants to Extreme Alterations in River Morphology


Abstract
:1. Introduction
2. Material and Methods
2.1. Hydromorphological Survey
2.2. Water Quality Survey
2.3. Macrophyte Survey
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EEA. European Waters—Assessment of Status and Pressures; European Environment Agency: Copenhagen, Denmark, 2012; p. 96. [Google Scholar]
- The European Parliament and the Council. Directive 2000/60/EC of the European Parliament and of the Council Establishing a Framework for Community Action in the Field of Water Policy. Available online: http://eur-lex.europa.eu/resource.html?uri=cellar:5c835afb-2ec6-4577-bdf8-756d3d694eeb.0004.02/DOC_1&format=PDF (accessed on 1 September 2022).
- Baattrup-Pedersen, A.; Larsen, S.E.; Rasmussen, J.J.; Riis, T. The future of European water management: Demonstration of a new WFD compliant framework to support sustainable management under multiple stress. Sci. Total Environ. 2018, 654, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.; Mackay, E.B.; Cardoso, A.C.; Baattrup-Pedersen, A.; Birk, S.; Blackstock, K.L.; Borics, G.; Borja, A.; Feld, C.K.; Ferreira, M.T.; et al. Protecting and restoring Europe’s waters: An analysis of the future development needs of the Water Framework Directive. Sci. Total Environ. 2018, 658, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Hering, D.; Borja, A.; Carstensen, J.; Carvalho, L.; Elliott, M.; Feld, C.K.; Heiskanen, A.-S.; Johnson, R.; Moe, J.; Pont, D. The European Water Framework Directive at the age of 10: A critical review of the achievements with recommendations for the future. Sci. Total Environ. 2010, 408, 4007–4019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, B. The Water Framework Directive: Total environment or political compromise? Sci. Total Environ. 2008, 400, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Dahm, V.; Hering, D.; Nemitz, D.; Graf, W.; Schmidt-Kloiber, A.; Leitner, P.; Melcher, A.; Feld, C.K. Effects of physico-chemistry, land use and hydromorphology on three riverine organism groups: A comparative analysis with monitoring data from Germany and Austria. Hydrobiologia 2013, 704, 389–415. [Google Scholar] [CrossRef]
- Turunen, J.; Muotka, T.; Vuori, K.-M.; Karjalainen, S.M.; Rääpysjärvi, J.; Sutela, T.; Aroviita, J. Disentangling the responses of boreal stream assemblages to low stressor levels of diffuse pollution and altered channel morphology. Sci. Total Environ. 2016, 544, 954–962. [Google Scholar] [CrossRef]
- Gebler, D.; Wiegleb, G.; Szoszkiewicz, K. Integrating river hydromorphology and water quality into ecological status modelling by artificial neural networks. Water Res. 2018, 139, 395–405. [Google Scholar] [CrossRef]
- Hachoł, J.; Bondar-Nowakowska, E.; Nowakowska, E. Factors Influencing Macrophyte Species Richness in Unmodified and Altered Watercourses. Pol. J. Environ. Stud. 2019, 28, 609–622. [Google Scholar] [CrossRef]
- Jusik, S.; Macioł, A. The influence of hydromorphological modifications of the littoral zone in lakes on macrophytes. Oceanol. Hydrobiol. Stud. 2014, 43, 66–76. [Google Scholar] [CrossRef]
- O’Hare, M.T.; Baattrup-Pedersen, A.; Nijboer, R.; Szoszkiewicz, K.; Ferreira, T. Macrophyte communities of European streams with altered physical habitat. Hydrobiologia 2006, 566, 197–210. [Google Scholar] [CrossRef]
- Bączyk, A.; Wagner, M.; Okruszko, T.; Grygoruk, M. Influence of technical maintenance measures on ecological status of agricultural lowland rivers—Systematic review and implications for river management. Sci. Total Environ. 2018, 627, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Erba, S.; Terranova, L.; Cazzola, M.; Cason, M.; Buffagni, A.S. Defining Maximum Ecological Potential for heavily modified lowland streams of Northern Italy. Sci. Total Environ. 2019, 684, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Wyżga, B.; Amirowicz, A.; Oglęcki, P.; Hajdukiewicz, H.; Radecki-Pawlik, A.; Zawiejska, J.; Mikuś, P. Response of fish and benthic invertebrate communities to constrained channel conditions in a mountain river: Case study of the Biała, Polish Carpathians. Limnologica 2014, 46, 58–60. [Google Scholar] [CrossRef]
- Zelnik, I.; Muc, T. Relationship between Environmental Conditions and Structure of Macroinvertebrate Community in a Hydromorphologically Altered Pre-Alpine River. Water 2020, 12, 2987. [Google Scholar] [CrossRef]
- Haase, P.; Hering, D.; Jähnig, S.C.; Lorenz, A.W.; Sundermannet, A. The impact of hydromorphological restoration on river ecological status: A comparison of fish, benthic invertebrates, and macrophytes. Hydrobiologia 2012, 704, 475–488. [Google Scholar] [CrossRef]
- Marzin, A.; Archaimbault, V.; Belliard, J.; Chauvin, C.; Delmas, F.; Pont, D. Ecological assessment of running waters: Do macrophytes, macroinvertebrates, diatoms and fish show similar responses to human pressures? Ecol. Indic. 2012, 23, 56–65. [Google Scholar] [CrossRef]
- Horsák, M.; Bojková, J.; Zahrádková, S.; Omesová, M.; Helešic, J. Impact of reservoirs and channelization on lowland river macroinvertebrates: A case study from Central Europe. Limnologica 2009, 39, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Swales, S. Fish populations of a small lowland channelized river in England subject to long-term river maintenance and management works. Regul. Rivers Res. Manag. 1988, 2, 493–506. [Google Scholar] [CrossRef]
- Biggs, B.J.F. Patterns in Benthic Algae of Streams. In Algal Ecology: Freshwater Benthic Ecosystem; Stevenson, R.J., Bothwell, M.L., Lowe, R., Thorp, J., Eds.; Academic Press: Cambridge, MA, USA, 1996; pp. 31–56. [Google Scholar] [CrossRef]
- Buffagni, A.; Barca, E.; Erba, S.; Balestrini, R. In-stream microhabitat mosaic depicts the success of mitigation measures and controls the Ecological Potential of benthic communities in heavily modified rivers. Sci. Total Environ. 2019, 673, 489–501. [Google Scholar] [CrossRef]
- Jukonienė, I. The impact of anthropogenic habitats on rare bryophyte species in Lithuania. Folia Cryptogam. Est. 2008, 44, 55–62. [Google Scholar]
- Vieira, J.; Necchi, O. Microhabitat and plant structure of Characeae (Chlorophyta) populations in streams from São Paulo State, southeaster Brazil. Criptogam. Algol. 2002, 23, 51–63. [Google Scholar] [CrossRef]
- Jusik, S.; Staniszewski, R. Shading of River Channels as an Important Factor Reducing Macrophyte Biodiversity. Pol. J. Environ. Stud. 2019, 28, 1215–1222. [Google Scholar] [CrossRef]
- Köhler, J.; Hachoł, J.; Hilt, S. Regulation of submersed macrophyte biomass in a temperate lowland river: Interactions between shading by bank vegetation, epiphyton and water turbidity. Aquat. Bot. 2010, 92, 129–136. [Google Scholar] [CrossRef]
- Suren, A.M.; Riis, T.; Biggs, B.J.F.; McMurtrie, S.; Barker, R. Assessing the effectiveness of enhancement activities in urban streams: I. Habitat responses. River Res. Appl. 2005, 21, 381–401. [Google Scholar] [CrossRef]
- Dodkins, I.; Aguiar, F.; Rivaes, R.; Albuquerque, A.; Rodríguez-González, P.; Ferreira, M.T. Measuring ecological change of aquatic macrophytes in Mediterranean rivers. Limnologica 2012, 42, 95–107. [Google Scholar] [CrossRef]
- Ceschin, F.; Bini, L.M.; Padial, A.A. Correlates of fish and aquatic macrophyte beta diversity in the Upper Paraná River floodplain. Hydrobiologia 2017, 805, 377–389. [Google Scholar] [CrossRef]
- Gurnell, A.; O’Hare, J.; O’Hare, M.; Dunbar, M.; Scarlett, P. An exploration of associations between assemblages of aquatic plant morphotypes and channel geomorphological properties within British rivers. Geomorphology 2010, 116, 135–144. [Google Scholar] [CrossRef]
- Kovalenko, K.E.; Thomaz, S.M.; Warfe, D.M. Habitat complexity: Approaches and future directions. Hydrobiologia 2012, 685, 1–17. [Google Scholar] [CrossRef]
- Baattrup-Pedersen, A.; Göthe, E.; Riis, T.; O’Hare, M.T. Functional trait composition of aquatic plants can serve to disentangle multiple interacting stressors in lowland streams. Sci. Total Environ. 2016, 543, 230–238. [Google Scholar] [CrossRef]
- Gebler, D.; Szoszkiewicz, K.; Pietruczuk, K. Modeling of the river ecological status with macrophytes using artificial neural networks. Limnologica 2017, 65, 46–54. [Google Scholar] [CrossRef]
- Hilton, J.; O’Hare, M.; Bowes, M.J.; Jones, J.I. How green is my river? A new paradigm of eutrophication in rivers. Sci. Total Environ. 2006, 365, 66–83. [Google Scholar] [CrossRef] [PubMed]
- Kolada, A.; Willby, N.; Dudley, B.; Nõges, P.; Søndergaard, M.; Hellsten, S.; Mjelde, M.; Penning, E.; van Geest, G.; Bertrin, V.; et al. The applicability of macrophyte compositional metrics for assessing eutrophication in European lakes. Ecol. Indic. 2014, 45, 407–415. [Google Scholar] [CrossRef]
- Szoszkiewicz, K.; Jusik, S.; Pietruczuk, K.; Gebler, D. The Macrophyte Index for Rivers (MIR) as an Advantageous Approach to Running Water Assessment in Local Geographical Conditions. Water 2019, 12, 108. [Google Scholar] [CrossRef] [Green Version]
- O’Hare, M.T.; Baattrup-Pedersen, A.; Baumgarte, I.; Freeman, A.; Gunn, I.D.M.; Lázár, A.N.; Sinclair, R.; Wade, A.J.; Bowes, M.J. Responses of Aquatic Plants to Eutrophication in Rivers: A Revised Conceptual Model. Front. Plant Sci. 2018, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Schinegger, R.; Trautwein, C.; Melcher, A.; Schmutz, S. Multiple human pressures and their spatial patterns in E uropean running waters. Water Environ. J. 2011, 26, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Hering, D.; Carvalho, L.; Argillier, C.; Beklioglu, M.; Borja, A.; Cardoso, A.C.; Duel, H.; Ferreira, T.; Globevnik, L.; Hanganu, J.; et al. Managing aquatic ecosystems and water resources under multiple stress—An introduction to the MARS project. Sci. Total Environ. 2015, 503–504, 10–21. [Google Scholar] [CrossRef]
- Raven, P.; Holmes, N.T.H.; Dawson, F.H.; Everard, M. Quality assessment using river habitat survey data. Aquat. Conserv. Mar. Freshw. Ecosyst. 1998, 8, 477–499. [Google Scholar] [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Dell Inc. Dell Statistica; Version 13; Data Analysis Software System; Dell Inc.: Round Rock, TX, USA, 2016. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco for Windows, Version 4.0; CPRO-DLO: Wageningen, The Netherlands, 1998. [Google Scholar]
- Budka, A.; Łacka, A.; Szoszkiewicz, K. The use of rarefaction and extrapolation as methods of estimating the effects of river eutrophication on macrophyte diversity. Biodivers. Conserv. 2018, 28, 385–400. [Google Scholar] [CrossRef] [Green Version]
- Tomczyk, P.; Wiatkowski, M.; Gruss, Ł. Application of Macrophytes to the Assessment and Classification of Ecological Status above and below the Barrage with Hydroelectric Buildings. Water 2019, 11, 1028. [Google Scholar] [CrossRef] [Green Version]
- Szoszkiewicz, K.; Ciecierska, H.; Kolada, A.; Schneider, S.C.; Szwabinska, M.; Ruszczyńska, J. Parameters structuring macrophyte communities in rivers and lakes—Results from a case study in North-Central Poland. Knowl. Manag. Aquat. Ecosyst. 2014, 415, 8. [Google Scholar] [CrossRef] [Green Version]
- Jusik, S.; Szoszkiewicz, K.; Kupiec, J.M.; Lewin, I.; Samecka-Cymerman, A. Development of comprehensive river typology based on macrophytes in the mountain-lowland gradient of different Central European ecoregions. Hydrobiologia 2014, 745, 241–262. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.-T.; Herricks, E.E.; Tsai, W.-P.; Chang, F.-J. Assessing the natural and anthropogenic influences on basin-wide fish species richness. Sci. Total Environ. 2016, 572, 825–836. [Google Scholar] [CrossRef]
- Lorenz, A.W.; Korte, T.; Sundermann, A.; Januschke, K.; Haase, P. Macrophytes respond to reach-scale river restorations. J. Appl. Ecol. 2012, 49, 202–212. [Google Scholar] [CrossRef]
- Schröder, M.; Kiesel, J.; Schattmann, A.; Jähnig, S.C.; Lorenz, A.W.; Kramm, S.; Keizer-Vlek, H.; Rolauffs, P.; Graf, W.; Leitner, P.; et al. Substratum associations of benthic invertebrates in lowland and mountain streams. Ecol. Indic. 2013, 30, 178–189. [Google Scholar] [CrossRef]
- Szoszkiewicz, K.; Budka, A.; Pietruczuk, K.; Kayzer, D.; Gebler, D. Is the macrophyte diversification along the trophic gradient distinct enough for river monitoring? Environ. Monit. Assess. 2016, 189, 4. [Google Scholar] [CrossRef] [Green Version]
- Thiebaut, G.; Guérold, F.; Muller, S. Are trophic and diversity indices based on macrophyte communities pertinent tools to monitor water quality? Water Res. 2002, 36, 3602–3610. [Google Scholar] [CrossRef]
- Willby, N.; Pitt, J.-A.; Phillips, G. The Ecological Classification of UK Rivers Using Aquatic Macrophytes; Report—SC010080/R1; Environment Agency: Bristol, UK, 2009; p. 221. [Google Scholar]
- Haury, J.; Peltre, M.-C.; Trémolières, M.; Barbe, J.; Thiébaut, G.; Bernez, I.; Daniel, H.; Chatenet, P.; Haan-Archipof, G.; Muller, S.; et al. A new method to assess water trophy and organic pollution—The Macrophyte Biological Index for Rivers (IBMR): Its application to different types of river and pollution. Hydrobiologia 2006, 570, 153–158. [Google Scholar] [CrossRef]
- Wiegleb, G.; Herr, W.; Zander, B.; Bröring, U.; Brux, H.; van de Weyer, K. Natural variation of macrophyte vegetation of lowland streams at the regional level. Limnologica 2015, 51, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Holmes, N.T.H.; Raven, P. Rivers. A Natural and Not-So-Natural History; Bloomsbury Publishing: London, UK, 2014; p. 432. [Google Scholar]
- Gerardo, R.; de Lima, I.P. Monitoring Duckweeds (Lemna minor) in Small Rivers Using Sentinel-2 Satellite Imagery: Application of Vegetation and Water Indices to the Lis River (Portugal). Water 2022, 14, 2284. [Google Scholar] [CrossRef]
- Van Dyck, I.; Vanhoudt, N.; i Batlle, J.V.; Horemans, N.; Nauts, R.; Van Gompel, A.; Claesen, J.; Vangronsveld, J. Effects of environmental parameters on Lemna minor growth: An integrated experimental and modelling approach. J. Environ. Manag. 2021, 300, 113705. [Google Scholar] [CrossRef]
- Dawson, F.H.; Raven, P.J.; Gravelle, M.J. Distribution of the morphological groups of aquatic plants for rivers in the U.K. In Biology, Ecology and Management of Aquatic Plants. Developments in Hydrobiology; Caffrey, J., Barrett, P.R.F., Ferreira, M.T., Moreira, I.S., Murphy, K.J., Wade, P.M., Eds.; Springer: Dordrecht, The Netherlands, 1999; Volume 147, pp. 123–130. [Google Scholar] [CrossRef]
- Lang, P.; Murphy, K.J. Environmental drivers, life strategies and bioindicator capacity of bryophyte communities in high-latitude headwater streams. Hydrobiologia 2011, 679, 1–17. [Google Scholar] [CrossRef]
- Shevock, J.R.; Ma, W.-Z.; Akiyama, H. Diversity of the rheophytic condition in bryophytes: Field observations from multiple continents. Bryophyt. Divers. Evol. 2017, 39, 75–93. [Google Scholar] [CrossRef] [Green Version]
- Vieira, C.; Aguiar, F.C.; Portela, A.P.; Monteiro, J.; Raven, P.J.; Holmes, N.T.H.; Cambra, J.; Flor-Arnau, N.; Chauvin, C.; Loriot, S.; et al. Bryophyte communities of Mediterranean Europe: A first approach to model their potential distribution in highly seasonal rivers. Hydrobiologia 2016, 812, 27–43. [Google Scholar] [CrossRef]
- Wharfe, J.; Taylor, K.; Montgomery, H. The growth of cladophora glomerata in a river receiving sewage effluent. Water Res. 1984, 18, 971–979. [Google Scholar] [CrossRef]
- Whitton, B.A.; Kelly, M.G. Use of algae and other plants for monitoring rivers. Austral Ecol. 1995, 20, 45–56. [Google Scholar] [CrossRef]
- Pikosz, M.; Messyasz, B. Characteristics of Cladophora and coexisting filamentous algae in relation to environmental factors in freshwater ecosystems in Poland. Oceanol. Hydrobiol. Stud. 2016, 45, 202–215. [Google Scholar] [CrossRef]
- Whitton, B. Biology of Cladophora in freshwaters. Water Res. 1970, 4, 457–476. [Google Scholar] [CrossRef]
- Birk, S.; Willby, N. Towards harmonization of ecological quality classification: Establishing common grounds in European macrophyte assessment for rivers. Hydrobiologia 2010, 652, 149–163. [Google Scholar] [CrossRef]
- Lange, K.; Townsend, C.R.; Matthaei, C.D. A trait-based framework for stream algal communities. Ecol. Evol. 2015, 6, 23–36. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variable | Shortcut | Unit | Minimum | Maximum | Mean | Median |
|---|---|---|---|---|---|---|
| Conductivity | Cond. | µS/cm | 150 | 2250 | 606 | 531 |
| Alkalinity | Alkal. | mg CaCO3/L | 50 | 490 | 192 | 175 |
| Reactive phosphorus | PO4-P | mg PO4/L | 0.02 | 4.40 | 0.34 | 0.19 |
| Total phosphorus | TP | mg P/L | 0.03 | 2.56 | 0.38 | 0.28 |
| Nitrate nitrogen | NO3-N | mg N/L | 0.02 | 5.74 | 0.92 | 0.60 |
| Ammonia nitrogen | NH4-N | mg N/L | 0.01 | 7.75 | 0.45 | 0.21 |
| Total nitrogen | TN | mg N/L | 0.71 | 26.77 | 3.85 | 3.23 |
| Dissolved oxygen | DO | mg 02/L | 0.80 | 22.32 | 8.17 | 8.18 |
| Channel width | Width | m | 0.65 | 15.00 | 5.12 | 4.00 |
| Channel depth | Depth | m | 0.05 | 3.50 | 0.66 | 0.50 |
| Habitat Quality Assessment | HQA | - | 6 | 52 | 27 | 27 |
| Habitat Modification Score | HMS | - | 11 | 108 | 62 | 59 |
| Taxon | Group of Macrophytes | Number of Sites | Frequency |
|---|---|---|---|
| Phalaris arundinacea L. | emergent | 138 | 73% |
| Lemna minor L. | free-floating | 125 | 66% |
| Sparganium emersum Rehmann | emergent, submerged, floating leaved | 105 | 55% |
| Cladophora sp. Kütz. | macroscopic algae | 102 | 54% |
| Agrostis stolonifera L. | emergent, amphibious | 86 | 45% |
| Myosotis palustris (L.) L. em. Rchb. | emergent, amphibious | 84 | 44% |
| Sparganium erectum L. em. Rchb. | emergent | 84 | 44% |
| Rorippa amphibia (L.) Besser | emergent, amphibious | 84 | 44% |
| Spirodela polyrhiza (L.) Schleid. | free-floating | 82 | 43% |
| Glyceria maxima (Hartm.) Holmb. | emergent | 80 | 42% |
| Callitriche cophocarpa Sendtn. | submerged | 62 | 33% |
| Elodea canadensis Michx. | submerged | 62 | 33% |
| Mentha aquatica L. | emergent, amphibious | 62 | 33% |
| Potamogeton pectinatus L. | submerged | 61 | 32% |
| Ranunculus repens L. | emergent, amphibious | 61 | 32% |
| Berula erecta (Huds.) Coville | emergent, submerged | 59 | 31% |
| Sagittaria sagittifolia L. | emergent, submerged, floating leaved | 55 | 29% |
| Veronica anagallis-aquatica L. | emergent, submerged | 51 | 27% |
| Glyceria fluitans (L.) R.Br. | emergent, amphibious | 47 | 25% |
| Leptodictyum riparium (Hedw.) Warnst. | bryophyte | 47 | 25% |
| Variable | Symbol | Minimum | Maximum | Mean | Median |
|---|---|---|---|---|---|
| Abundance (cover) | pi (%) | 0.25 | 100.00 | 34.48 | 28.75 |
| Shannon exponential | 1H | 1.04 | 15.83 | 4.96 | 4.38 |
| Simpson concentration | 2D | 1.01 | 12.16 | 3.47 | 2.86 |
| Species richness | N | 2 | 40 | 16 | 16 |
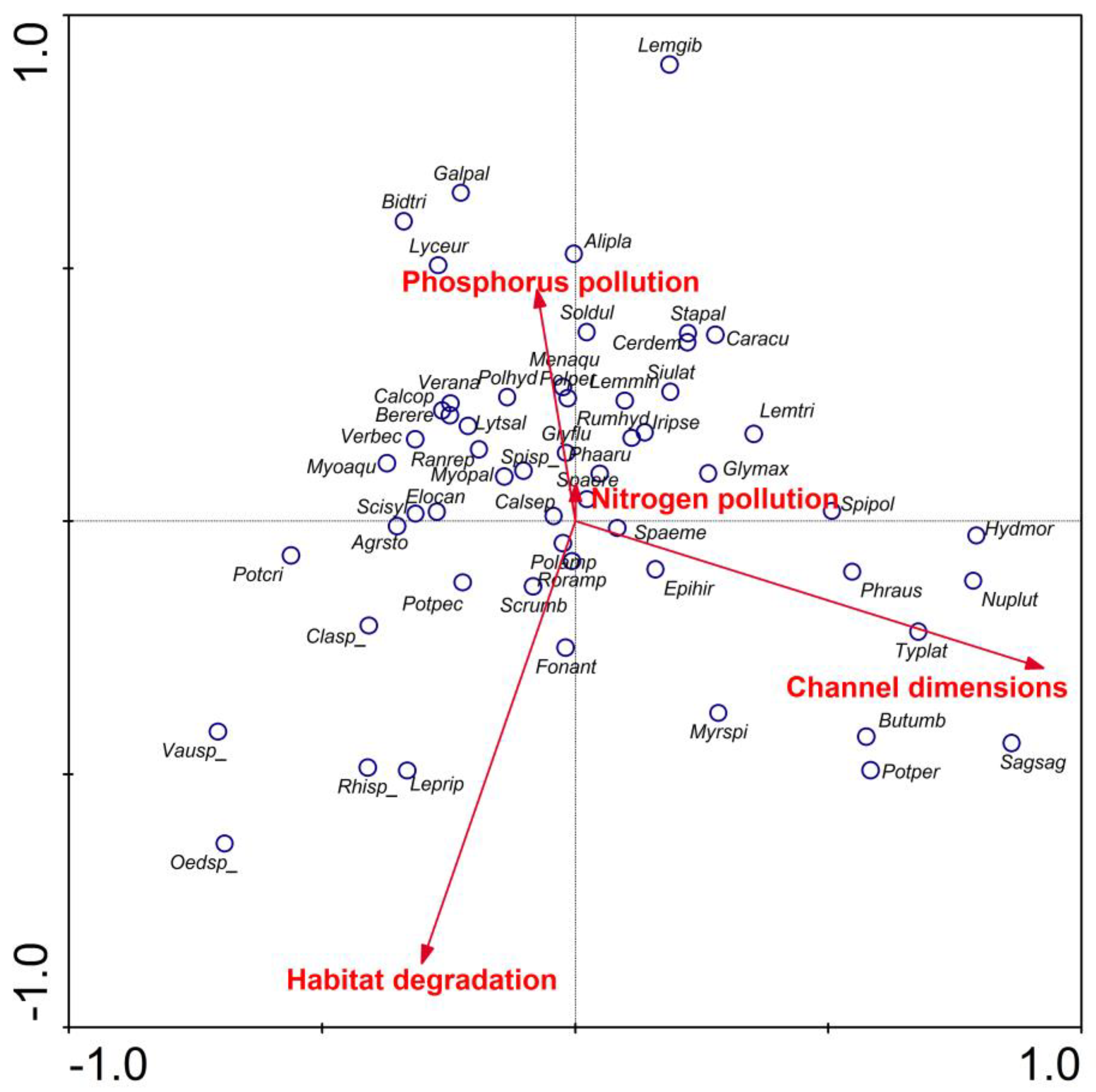
| Variable | 1st Factor | 2nd Factor | 3rd Factor | 4th Factor |
|---|---|---|---|---|
| Phosphorus Pollution | Channel Dimensions | Nitrogen Pollution | Habitat Degradation | |
| Habitat Quality Assessment | −0.172 | −0.095 | −0.022 | −0.797 |
| Habitat Modification Score | −0.089 | −0.162 | −0.122 | 0.762 |
| Conductivity | 0.648 | −0.199 | 0.113 | 0.037 |
| Alkalinity | 0.726 | −0.115 | −0.117 | −0.027 |
| Reactive phosphorus | 0.806 | 0.028 | −0.028 | −0.047 |
| Total phosphorus | 0.752 | 0.090 | 0.082 | 0.113 |
| Nitrate | −0.018 | −0.047 | 0.855 | −0.053 |
| Ammonia | 0.600 | −0.020 | 0.285 | 0.009 |
| Total nitrogen | 0.336 | 0.069 | 0.653 | −0.221 |
| Dissolved oxygen | −0.042 | −0.152 | 0.544 | 0.423 |
| Channel width | −0.139 | 0.893 | −0.057 | −0.031 |
| Channel depth | −0.008 | 0.912 | −0.023 | −0.066 |
| % of explained variance | 22.5% | 14.6% | 13.2% | 12.2% |
| Axes | 1 | 2 | 3 | 4 | Total Interia |
|---|---|---|---|---|---|
| Eigenvalues | 0.132 | 0.070 | 0.047 | 0.018 | 3.653 |
| Species–environment correlations | 0.716 | 0.583 | 0.610 | 0.441 | |
| Cumulative percentage variance | |||||
| of species data | 3.6 | 5.5 | 6.8 | 7.3 | |
| of species–environment relation | 49.5 | 75.6 | 93.3 | 100.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebler, D.; Szoszkiewicz, K. Response of Aquatic Plants to Extreme Alterations in River Morphology. Water 2022, 14, 3746. https://doi.org/10.3390/w14223746
Gebler D, Szoszkiewicz K. Response of Aquatic Plants to Extreme Alterations in River Morphology. Water. 2022; 14(22):3746. https://doi.org/10.3390/w14223746
Chicago/Turabian StyleGebler, Daniel, and Krzysztof Szoszkiewicz. 2022. "Response of Aquatic Plants to Extreme Alterations in River Morphology" Water 14, no. 22: 3746. https://doi.org/10.3390/w14223746