
The Detrimental Effect of High Salinity on the Growth and Microcystins Contamination of Microcystis aeruginosa



Abstract
:1. Introduction
2. Materials and Methods
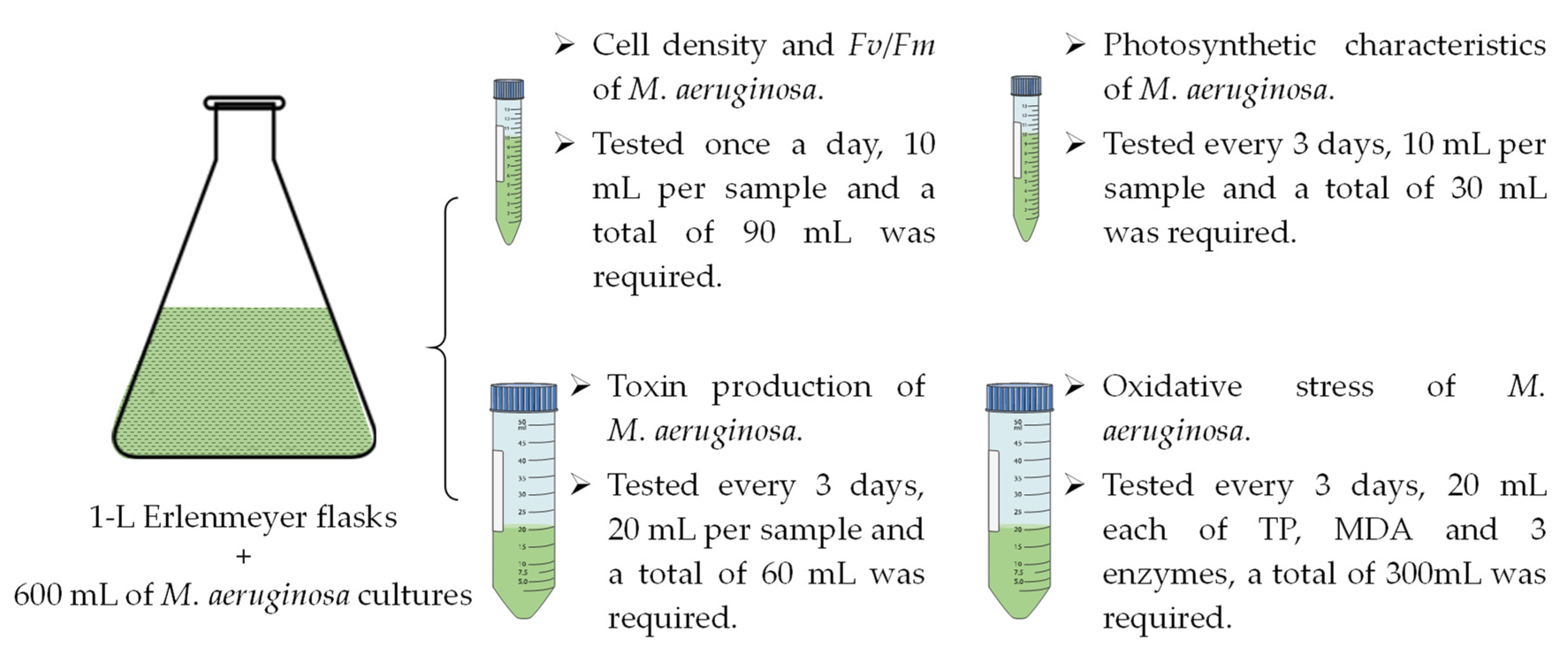
2.1. Culture Conditions and Experimental Design
2.2. Analysis Methods
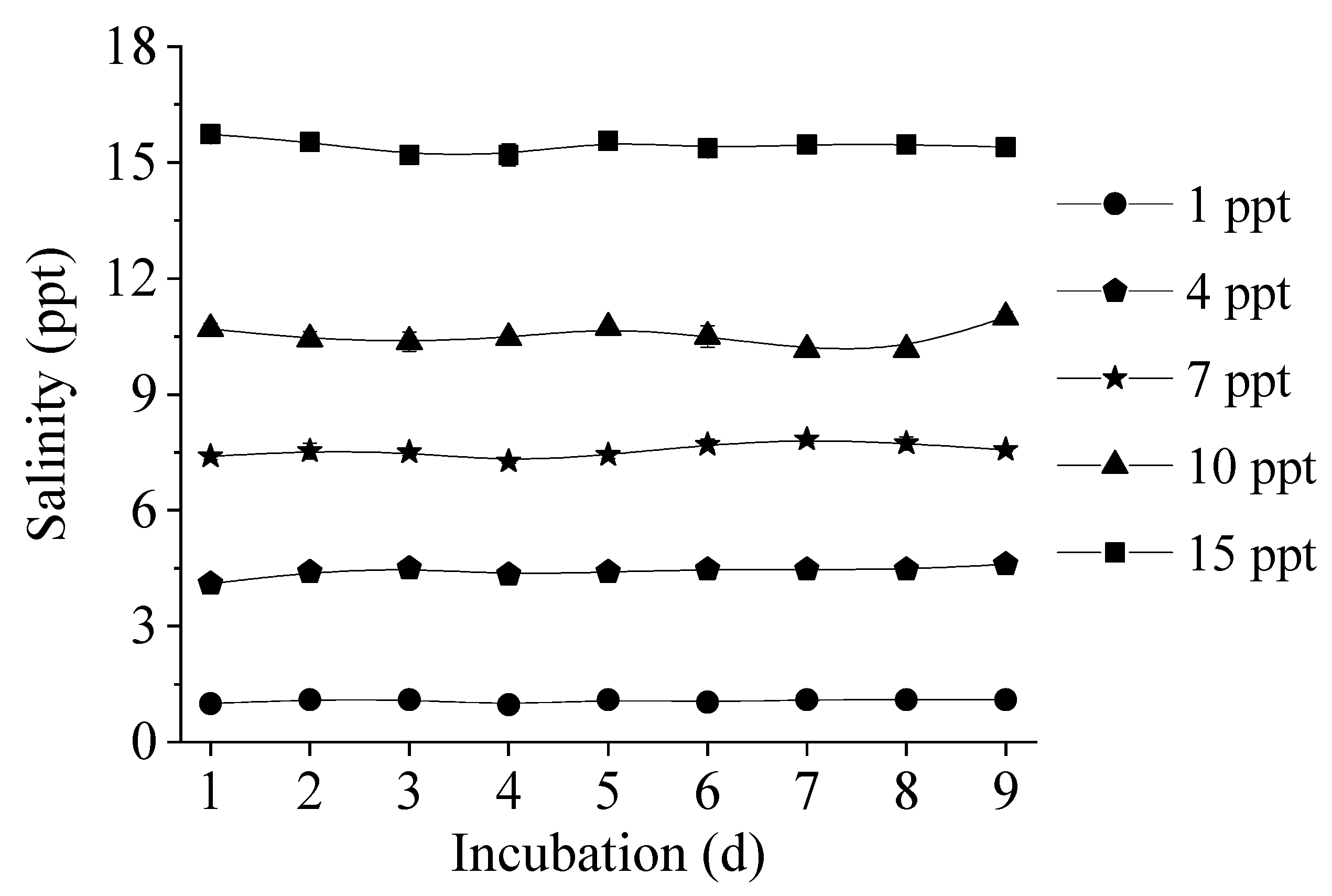
2.2.1. Salinity and Cell Density
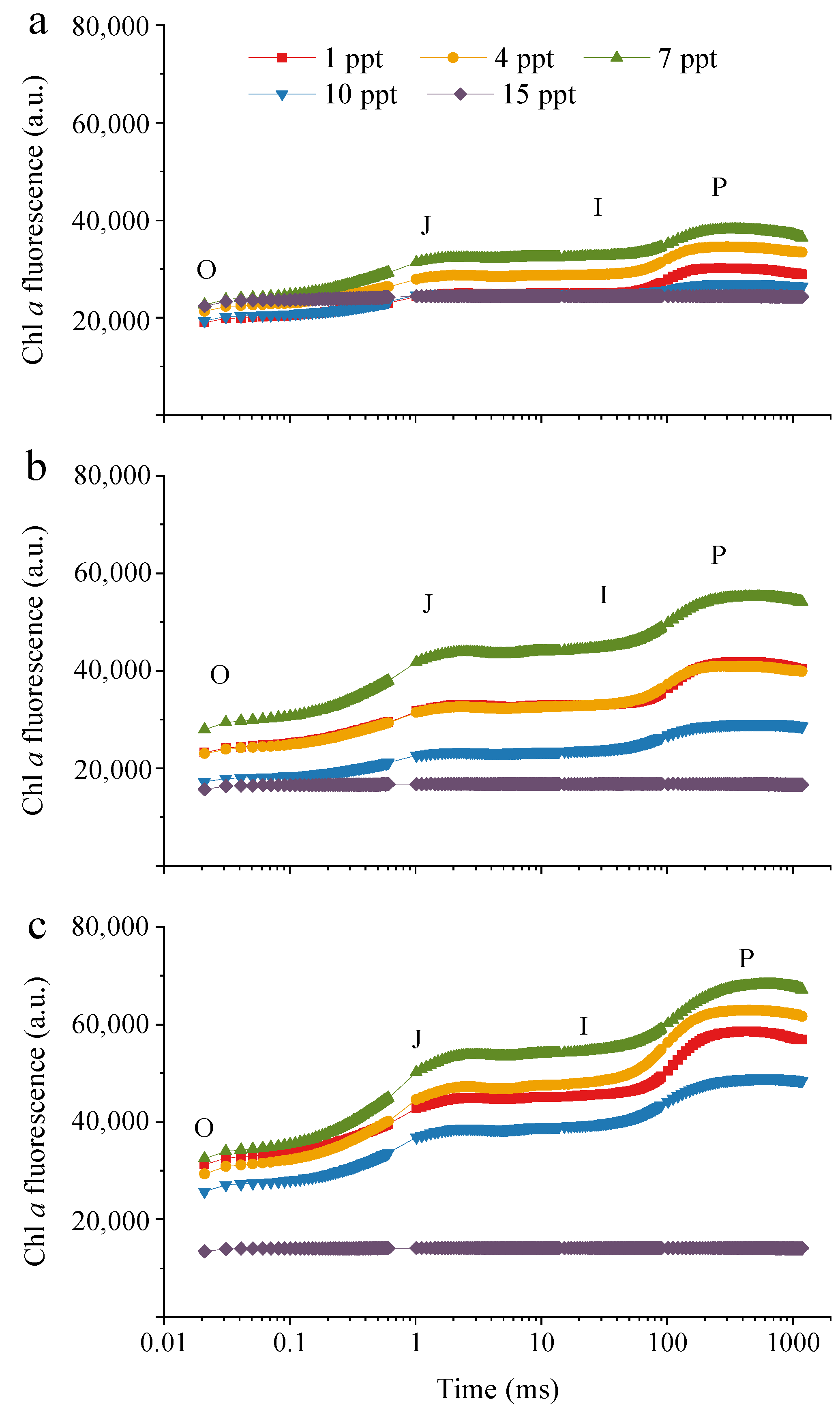
2.2.2. Fv/Fm and the Chlorophyll a Fluorescence Transient Analysis
2.2.3. Microcystins Quantification
2.2.4. Antioxidant Enzyme Activity and Malondialdehyde Content
2.3. Statistical Analysis
3. Results
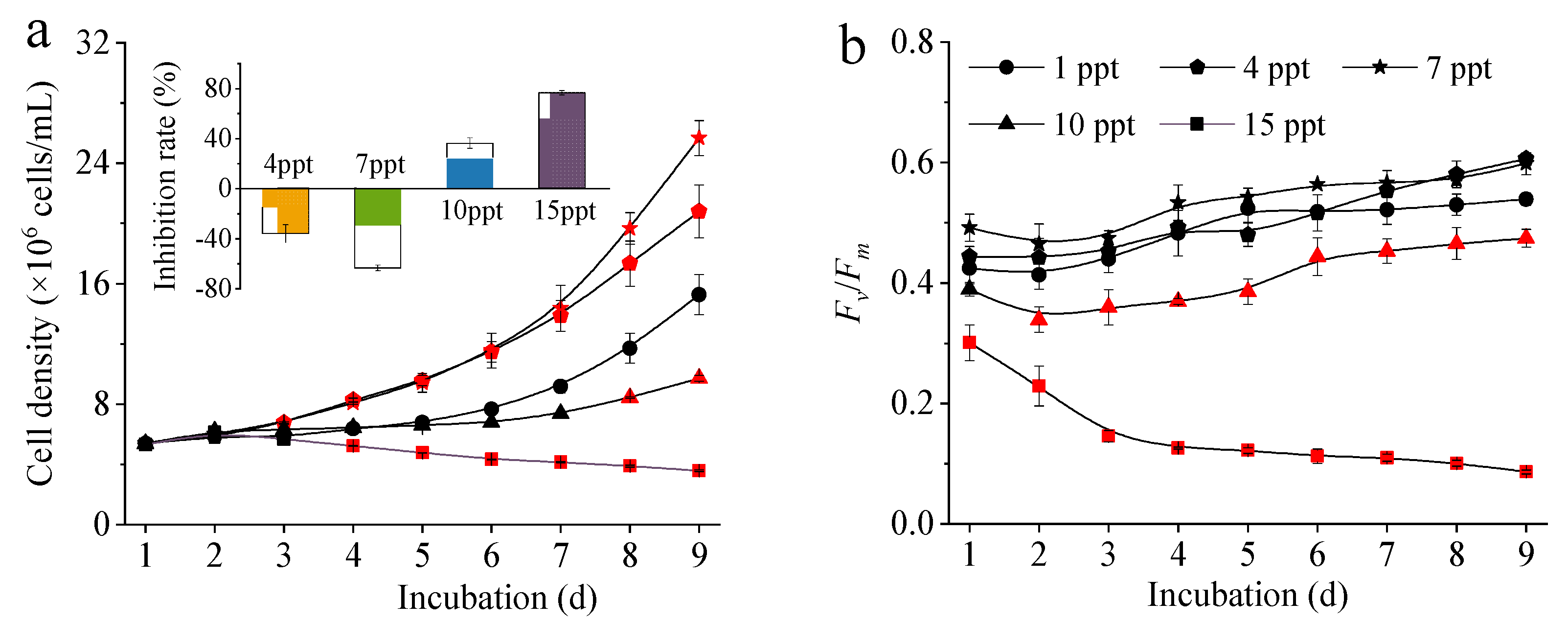
3.1. Cell Density and Fv/Fm of M. aeruginosa under Salinity Stress
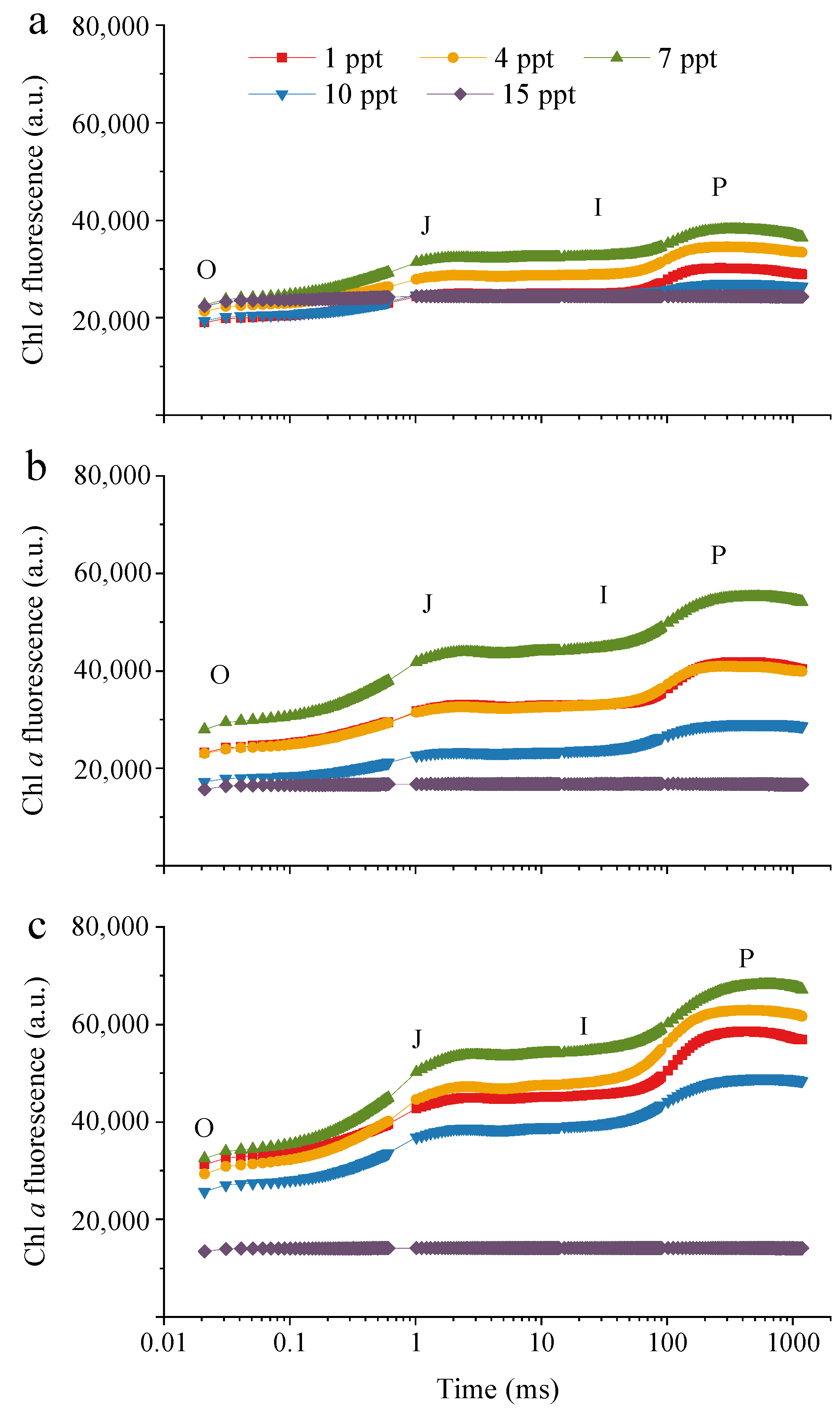
3.2. Photosynthetic Characteristics of M. aeruginosa under Salinity Stress
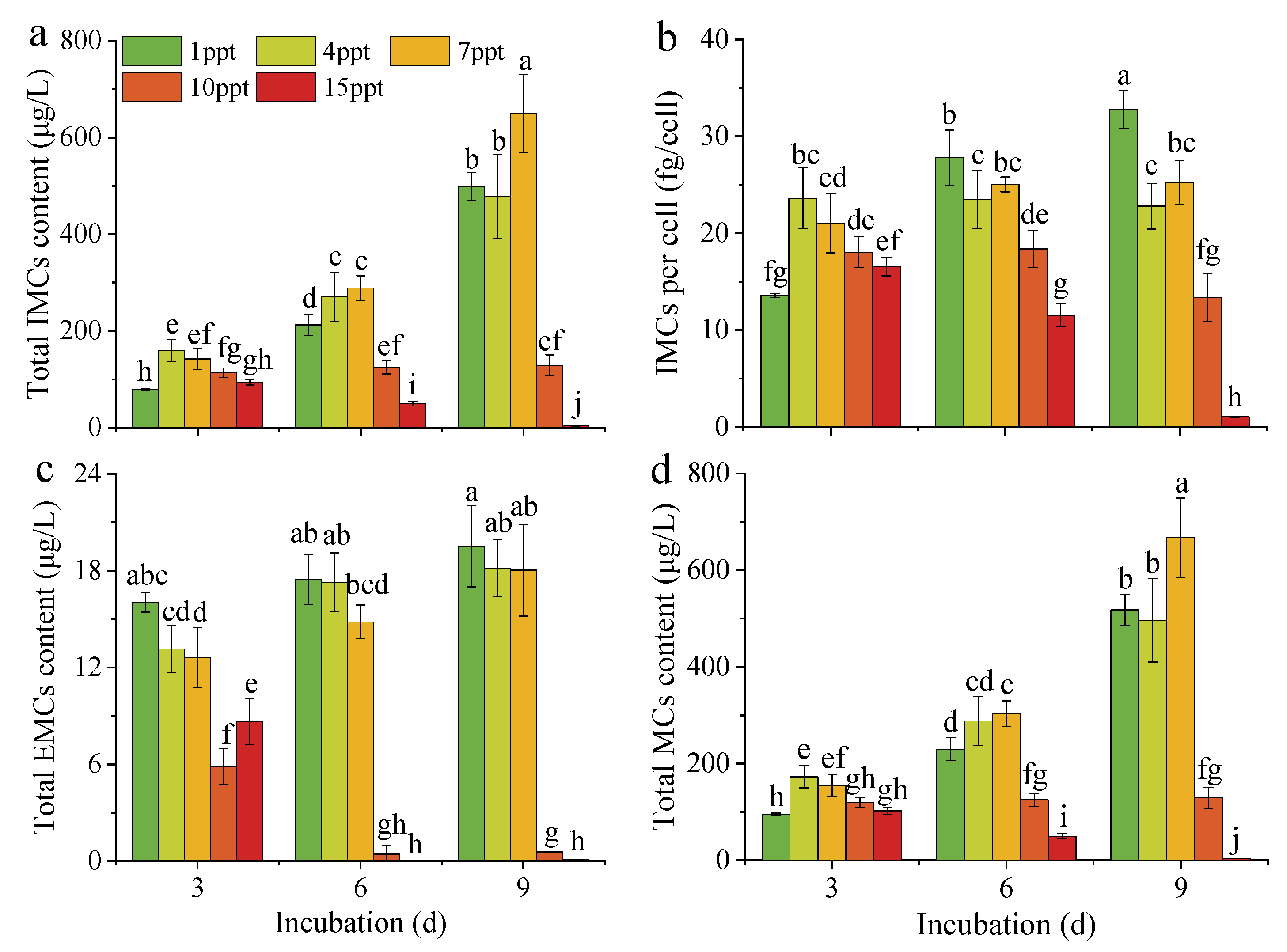
3.3. Toxin Production of M. aeruginosa under Salinity Stress
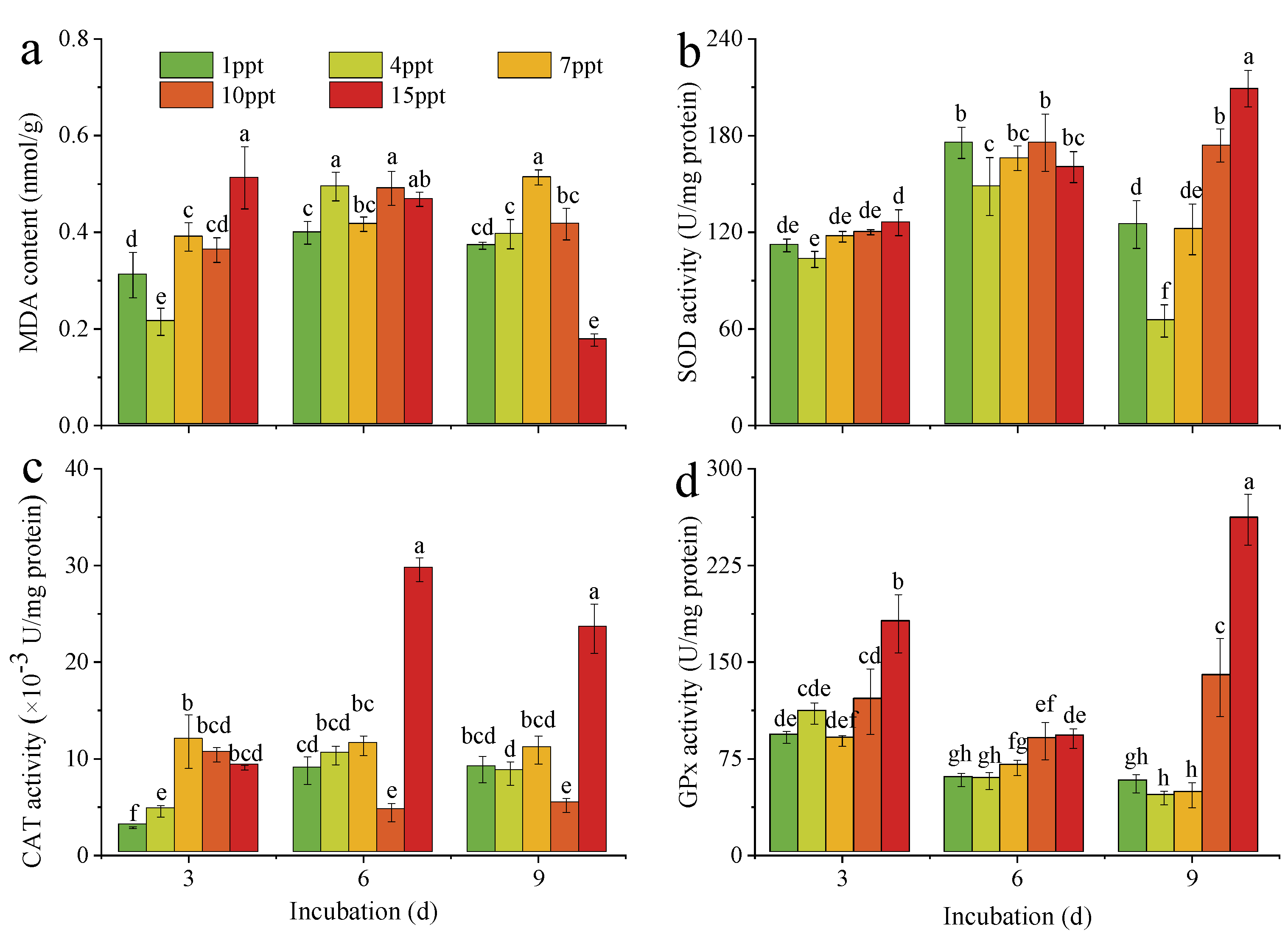
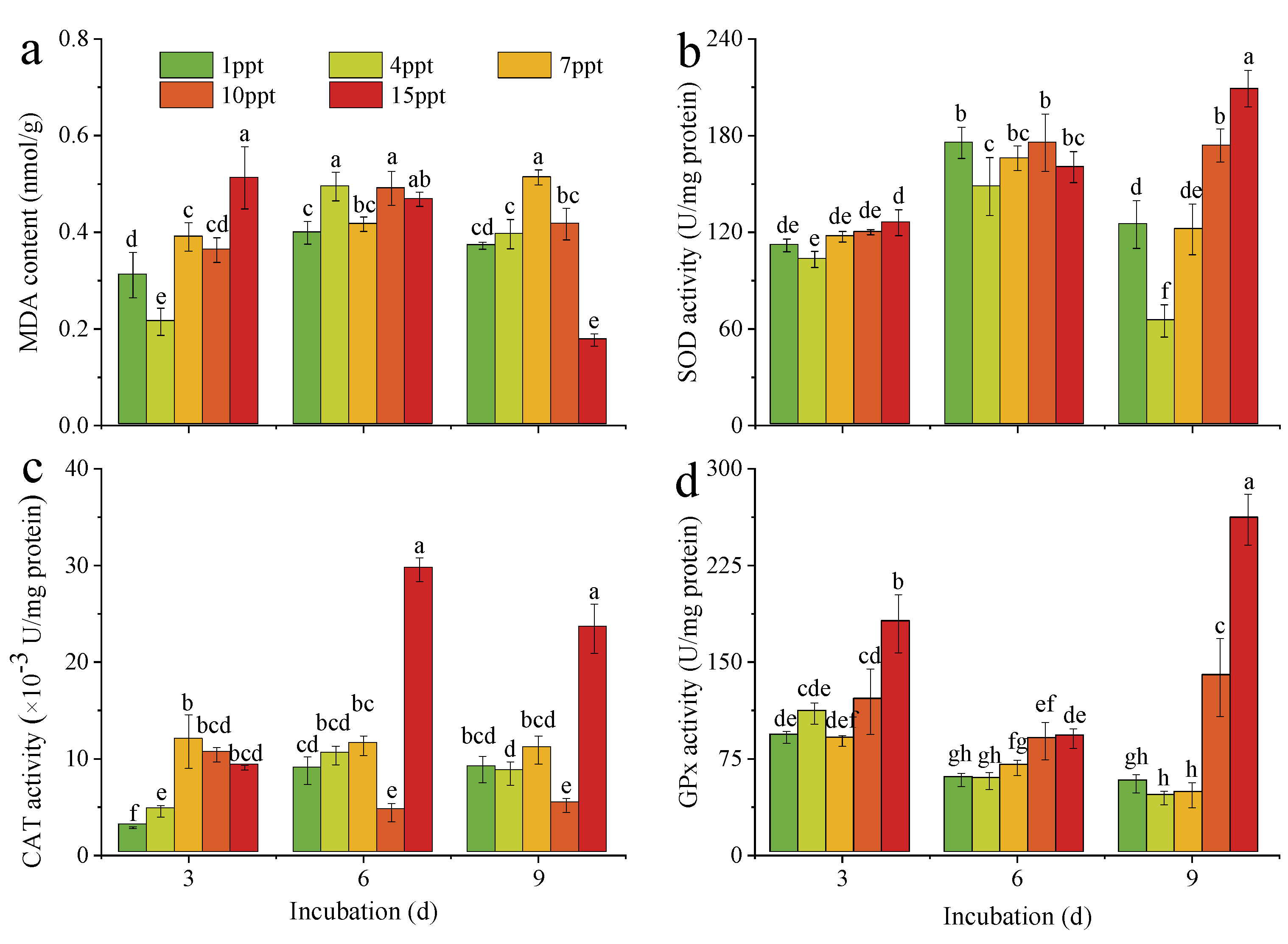
3.4. Oxidative Stress of M. aeruginosa under Salinity Stress
4. Discussion
4.1. Effects of Salinity Stress on the Growth and Photosynthetic Characteristics of M. aeruginosa
4.2. Effect of Salinity Stress on the Toxicity of M. aeruginosa
4.3. Effects of Salinity Stress on the Oxidative Stress of M. aeruginosa
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Vu, D.T.; Yamada, T.; Ishidaira, H. Assessing the impact of sea level rise due to climate change on seawater intrusion in Mekong Delta, Vietnam. Water Sci. Technol. 2018, 77, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Loáiciga, H.A.; Pingel, T.J.; Garcia, E.S. Sea water intrusion by sea-Level rise: Scenarios for the 21st century. Groundwater 2012, 50, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.; Warhurst, B.C.; Brown, A.; Huff, C.; Ochrietor, J.D. Mesohaline conditions represent the threshold for oxidative stress, cell death and toxin release in the cyanobacterium Microcystis aeruginosa. Aquat. Toxicol. 2019, 206, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Brodie, J.; Chan, C.X.; De Clerck, O.; Cock, J.M.; Coelho, S.M.; Gachon, C.; Grossman, A.R.; Mock, T.; Raven, J.A.; Smith, A.G.; et al. The algal revolution. Trends Plant Sci. 2017, 22, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, X.; Huang, Y.; Shen, Y.; Liu, S.; Lu, J.; Hu, J.; You, W. The salt tolerance of the freshwater cyanobacterium Microcystis depends on nitrogen availability. Sci. Total Environ. 2021, 777, 146186. [Google Scholar] [CrossRef]
- Robson, B.J.; Hamilton, D.P. Summer flow event induces a cyanobacterial bloom in a seasonal Western Australian estuary. Mar. Freshwater Res. 2003, 54, 139–151. [Google Scholar] [CrossRef]
- Kurobe, T.; Lehman, P.W.; Haque, M.E.; Sedda, T.; Lesmeister, S.; Teh, S. Evaluation of water quality during successive severe drought years within Microcystis blooms using fish embryo toxicity tests for the San Francisco Estuary, California. Sci. Total Environ. 2018, 610, 1029–1037. [Google Scholar] [CrossRef]
- Kruk, C.; Martínez, A.; De La Escalera, G.M.; Trinchin, R.; Manta, G.; Segura, A.M.; Piccini, C.; Brena, B.; Yannicelli, B.; Fabiano, G.; et al. Rapid freshwater discharge on the coastal ocean as a mean of long distance spreading of an unprecedented toxic cyanobacteria bloom. Sci. Total Environ. 2021, 754, 142362. [Google Scholar] [CrossRef]
- Kozdęba, M.; Borowczyk, J.; Zimolag, E.; Wasylewski, M.; Dziga, D.; Madeja, Z.; Drukala, J. Microcystin-LR affects properties of human epidermal skin cells crucial for regenerative processes. Toxicon 2014, 80, 38–46. [Google Scholar] [CrossRef]
- Lone, Y.; Koni, R.K.; Bhide, M. An overview of the toxic effect of potential human carcinogen Microcystin-LR on testis. Toxicol. Rep. 2015, 2, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.Y.; Xie, L.Q.; Yan, Y.J. Microcystin-LR impairs zebrafish reproduction by affecting oogenesis and endocrine system. Chemosphere 2015, 120, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Sneed, J.M.; Meickle, T.; Engene, N.; Reed, S.; Gunasekera, S.; Paul, V.J. Bloom dynamics and chemical defenses of benthic cyanobacteria in the Indian River Lagoon, Florida. Harmful Algae 2017, 69, 75–82. [Google Scholar] [CrossRef]
- Orr, P.T.; Jones, G.J.; Douglas, G.B. Response of cultured Microcystis aeruginosa from the Swan River, Australia, to elevated salt concentration and consequences for bloom and toxin management in estuaries. Mar. Freshw. Res. 2004, 55, 277–283. [Google Scholar] [CrossRef]
- Chen, L.; Mao, F.J.; Kirumba, G.C.; Jiang, C.; Manefield, M.; He, Y.L. Changes in metabolites, antioxidant system, and gene expression in Microcystis aeruginosa under sodium chloride stress. Ecotox. Environ. Safe 2015, 122, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.; Fathabad, S.G.; Tabatabai, B.; Jafar, S.; Sitther, V. Microcystin levels in selected cyanobacteria exposed to varying salinity. J. Water Resour. Prot. 2019, 11, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Tonk, L.; Bosch, K.; Visser, P.M.; Huisman, J. Salt tolerance of the harmful cyanobacterium Microcystis aeruginosa. Aquat. Microb. Ecol. 2007, 46, 117–123. [Google Scholar] [CrossRef]
- Qian, H.F.; Yu, S.Q.; Sun, Z.Q.; Xie, X.C.; Liu, W.P.; Fu, Z.W. Effects of copper sulfate, hydrogen peroxide and N-phenyl-2-naphthylamine on oxidative stress and the expression of genes involved photosynthesis and microcystin disposition in Microcystis aeruginosa. Aquat. Toxicol. 2010, 99, 405–412. [Google Scholar] [CrossRef]
- Sun, X.; Cao, Y.; Xu, H.; Liu, Y.; Sun, J.; Qiao, D.; Cao, Y. Effect of nitrogen-starvation, light intensity and iron on triacylglyceride/carbohydrate production and fatty acid profile of Neochloris oleoabundans HK-129 by a two-stage process. Bioresour. Technol. 2014, 155, 204–212. [Google Scholar] [CrossRef]
- Verslues, P.E. Time to grow: Factors that control plant growth during mild to moderate drought stress. Plant Cell Environ. 2017, 40, 177–179. [Google Scholar] [CrossRef]
- Almeida, A.C.; Gomes, T.; Langford, K.; Thomas, K.V.; Tollefsen, K.E. Oxidative stress in the algae Chlamydomonas reinhardtii exposed to biocides. Aquat. Toxicol. 2017, 189, 50–59. [Google Scholar] [CrossRef]
- Singh, R.; Upadhyay, A.K.; Singh, D.P. Regulation of oxidative stress and mineral nutrient status by selenium in arsenic treated crop plant Oryza sativa. Ecotox. Environ. Safe 2018, 148, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Barros, A.; Delgadillo, I.; Coimbra, M.; Santos, C. Effects of fungus inoculation and salt stress on physiology and biochemistry of in vitro grapevines: Emphasis on sugar composition changes by FT-IR analyses. Environ. Exp. Bot. 2009, 65, 1–10. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, S.K.; Govindjee, G.; Srivastava, A.; Strasser, R. Polyphasic chlorophyll a fluorescence transcient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Lazár, D. Chlorophyll a fluorescence induction. BBA-Bioenergetics 1999, 1412, 1–28. [Google Scholar] [CrossRef]
- Wang, C.X.; Wang, M.; Chen, B.B.; Qin, W.L.; Lin, L.D.; Dai, C.J.; Yu, H.G.; Li, R.H.; Zhao, M.; Ma, Z.L. Harmful algal bloom-forming dinofl agellate Prorocentrum donghaiense inhibits the growth and photosynthesis of seaweed Sargassum fusiformis embryos. J. Oceano. Limnol. 2021, 39, 2237–2251. [Google Scholar] [CrossRef]
- Preece, E.P.; Hardy, F.J.; Moore, B.C.; Bryan, M. A review of microcystin detections in Estuarine and Marine waters: Environmental implications and human health risk. Harmful Algae 2017, 61, 31–45. [Google Scholar] [CrossRef]
- Phooprong, S.; Ogawa, H.; Hayashizaki, K. Photosynthetic and respiratory responses of Gracilaria salicornia (C. Ag.) Dawson (Gracilariales, Rhodophyta) from Thailand and Japan. J. Appl. Phycol. 2007, 19, 795–801. [Google Scholar] [CrossRef]
- Benavente-Valdes, J.R.; Aguilar, C.; Contreras-Esquivel, J.C.; Mendez-Zavala, A.; Montanez, J. Strategies to enhance the production of photosynthetic pigments and lipids in chlorophycae species. Biotechnol. Rep. 2016, 10, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Upadhyay, A.K.; Chandra, P.; Singh, D.P. Sodium chloride incites reactive oxygen species in green algae Chlorococcum humicola and Chlorella vulgaris: Implication on lipid synthesis, mineral nutrients and antioxidant system. Bioresour. Technol. 2018, 270, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M.; Erdmann, N.; Wittenburg, E. Synthesis of glucosylglycerol in salt-stressed cells of the cyanobacterium Microcystis firma. Arch. Microbiol. 1987, 148, 275–279. [Google Scholar] [CrossRef]
- Georges des Aulnois, M.; Roux, P.; Caruana, A.; Réveillon, D.; Briand, E.; Hervé, F.; Savar, V.; Bormans, M.; Amzil, Z. Physiological and metabolic responses of freshwater and brackish-water strains of Microcystis aeruginosa acclimated to a salinity gradient: Insight into salt tolerance. Appl. Environ. Microb. 2019, 85, e01614-19. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Xu, Q.J.; Xi, B.D. Effect of NaCl salinity on the growth, metabolites, and antioxidant system of Microcystis aeruginosa. J. Freshw. Ecol. 2013, 28, 477–487. [Google Scholar] [CrossRef]
- Couée, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Potts, M.; Slaughter, S.M.; Hunneke, F.U.; Garst, J.F.; Helm, R.F. Desiccation tolerance of prokaryotes: Application of principles to human cells. Integr. Comp. Biol. 2005, 45, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Ördög, V.; Stirk, W.A.; Bálint, P.; Staden, J.; Lovász, C. Changes in lipid, protein and pigment concentrations in nitrogen-stressed Chlorella minutissima cultures. J. Appl. Phycol. 2012, 24, 907–914. [Google Scholar] [CrossRef]
- Tanabe, Y.; Hodoki, Y.; Sano, T.; Tada, K.; Watanabe, M.M. Adaptation of the freshwater bloom-forming cyanobacterium Microcystis aeruginosa to brackish water is driven by recent horizontal transfer of sucrose genes. Front. Microbiol. 2018, 9, 1150. [Google Scholar] [CrossRef]
- Lu, C.; Vonshak, A. Effects of salinity stress on photosystem II function in cyanobacterial Spirulina platensis cells. Physiol. Plant. 2002, 114, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Swapnil, P.; Yadav, A.K.; Srivastav, S.; Sharma, N.K.; Srikrishna, S.; Rai, A.K. Biphasic ROS accumulation and programmed cell death in a cyanobacterium exposed to salinity (NaCl and Na2SO4). Algal Res. 2017, 23, 88–95. [Google Scholar] [CrossRef]
- Rai, A.K. Biochemical characteristics of photosynthetic response to various external salanities in halotolerant and fresh water cyanobacteria. FEMS Microbiol. Lett. 1990, 69, 177–180. [Google Scholar] [CrossRef]
- Wang, G.; Jin, L.; Wang, X.; Hu, J.; Liu, Y.; Cai, W. Response of photosynthetic systems to salinity stress in the desert cyanobacterium Scytonema javanicum. Adv. Space Res. 2014, 53, 30–36. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, Z.S.; Gao, H.Y.; Fan, X.L.; Liu, M.J.; Li, X.D. The mechanism by which NaCl treatment alleviates PSI photoinhibition under chilling-light treatment. J. Photoch. Photobio. B 2014, 140, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Swapnil, P.; Singh, M.; Singh, S.; Sharma, N.K.; Rai, A.K. Recombinant glycinebetaine improves metabolic activities, ionic balance and salt tolerance in diazotrophic freshwater cyanobacteria. Algal Res. 2015, 11, 194–203. [Google Scholar] [CrossRef]
- Wiedner, C.; Visser, P.M.; Fastner, J.; Metcalf, J.S.; Codd, G.A.; Mur, L.R. Effects of light on the microcystin content of Microcystis strain PCC 7806. Appl. Environ. Microbiol. 2003, 69, 1475–1481. [Google Scholar] [CrossRef]
- Dolman, A.M.; Rucker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and cyanotoxins: The influence of nitrogen versus phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef]
- Black, K.; Yilmaz, M.; Phlips, E.J. Growth and toxin production by Microcystis aeruginosa PCC 7806 (Kutzing) Lemmerman at elevated salt concentrations. J. Environ. Prot. 2011, 2, 669–674. [Google Scholar] [CrossRef]
- Paldaviciene, A.; Mazur-Marzec, H.; Razinkovas, A. Toxic cyanobacteria blooms in the Lithuanian part of the Curonian Lagoon. Oceanologia 2009, 51, 203–216. [Google Scholar] [CrossRef]
- de la Escalera, G.M.; Kruk, C.; Segura, A.M.; Nogueira, L.; Alcántara, I.; Piccini, C. Dynamics of toxic genotypes of Microcystis aeruginosa complex (MAC) through a wide freshwater to marine environmental gradient. Harmful Algae 2017, 62, 73–83. [Google Scholar] [CrossRef]
- Engström-Öst, J.; Repka, S.; Mikkonen, M. Interactions between plankton and cyanobacterium Anabaena with focus on salinity, growth and toxin production. Harmful Algae 2011, 10, 530–535. [Google Scholar] [CrossRef]
- Kehr, J.C.; Zilliges, Y.; Springer, A.; Disney, M.D.; Ratner, D.D.; Bouchier, C.; Seeberger, P.H.; De Marsac, N.T.; Dittmann, E. A mannan binding lectin is involved in cell–cell attachment in a toxic strain of Microcystis aeruginosa. Mol. Microbiol. 2006, 59, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Schatz, D.; Keren, Y.; Vardi, A.; Sukenik, A.; Carmeli, S.; Brner, T.; Dittmann, E.; Kaplan, A. Towards clarification of the biological role of microcystins, a family of cyanobacterial toxins. Environ. Microbiol. 2010, 9, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Martín-Luna, B.; Sevilla, E.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Variation in the synthesis of microcystin in response to saline and osmotic stress in Microcystis ruginosa PCC7806. Limnetica 2015, 34, 205–214. [Google Scholar] [CrossRef]
- Chen, L.; Giesy, J.P.; Adamovsky, O.; Svircev, Z.; Xie, P. Challenges of using blooms of Microcystis spp. in animal feeds: A comprehensive review of nutritional, toxicological and microbial health evaluation. Sci. Total Environ. 2021, 764, 142319. [Google Scholar] [CrossRef]
- Tang, Q.M.; Chen, F.; Xiang, W.Z.; Peng, J.; He, H. Effect of ammonium nitrogen on the growth, biochemical constitutes and toxin production of Microcystis aeruginosa FACHB-905. J. Jinan Univ. 2008, 3, 290–294, (In Chinese with English abstract). [Google Scholar]
- Kuniyoshi, T.M.; Sevilla, E.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Phosphate deficiency (N/P 40:1) induces mcyD transcription and microcystin synthesis in Microcystis aeruginosa PCC7806. Plant Physiol. Bioch. 2013, 65, 120–124. [Google Scholar] [CrossRef]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Iron availability affects mcyD expression and microcystin-LR synthesis in Microcystis aeruginosa PCC7806. Environ. Microbiol. 2008, 10, 2476–2483. [Google Scholar] [CrossRef]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Bes, M.T.; Peleato, M.L.; Fillat, M.F. Microcystin-LR synthesis as response to nitrogen: Transcriptional analysis of the mcyD gene in Microcystis aeruginosa PCC7806. Ecotoxicology 2010, 19, 1167–1173. [Google Scholar] [CrossRef]
- Sevilla, E.; Martin-Luna, B.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. An active photosynthetic electron transfer chain required for mcyD transcription and microcystin synthesis in Microcystis aeruginosa PCC7806. Ecotoxicology 2012, 21, 811–819. [Google Scholar] [CrossRef]
- Choudhury, S.; Panda, P.; Sahoo, L.; Panda, S.K. Reactive oxygen species signaling in plants under abiotic stress. Plant Signal. Behav. 2013, 8, e23681. [Google Scholar] [CrossRef]
- Abogadallah, G.M. Antioxidative defense under salt stress. Plant Signal. Behav. 2010, 5, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, C.W. Co-existence of photosynthetic and respiratory activities in cyanobacterial thylakoid membranes. BBA-Bioenergetics 2014, 1837, 503–511. [Google Scholar] [CrossRef]
- Mikula, P.; Zezulka, S.; Jancula, D.; Marsalek, B. Metabolic activity and membrane integrity changes in Microcystis aeruginosa -new findings on hydrogen peroxide toxicity in cyanobacteria. Eur. J. Phycol. 2012, 47, 195–206. [Google Scholar] [CrossRef]
- Verma, E.; Chakraborty, S.; Tiwari, B.; Singh, S.; Mishra, A.K. Alleviation of NaCl toxicity in the cyanobacterium Synechococcus sp. PCC 7942 by exogenous calcium supplementation. J. Appl. Phycol. 2018, 30, 1465–1482. [Google Scholar] [CrossRef]
- Mallik, S.; Nayak, M.; Sahu, B.B.; Panigrahi, A.K.; Shaw, B.P. Response of antioxidant enzymes to high NaCl concentration in different salt-tolerant plants. Biol. Plant. 2011, 55, 191–195. [Google Scholar] [CrossRef]
- Chakravarty, D.; Banerjee, M.; Bihani, S.C.; Ballal, A. A salt-inducible Mn-catalase (KatB) protects cyanobacterium from oxidative stress. Plant Physiol. 2016, 170, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Arias-Moreno, D.M.; Jimenez-Bremont, J.F.; Maruri-Lopez, I.; Delgado-Sanchez, P. Effects of catalase on chloroplast arrangement in Opuntia streptacantha chlorenchyma cells under salt stress. Sci. Rep. UK 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vardi, A.; Berman-Frank, I.; Rozenberg, T.; Hadas, O.; Kaplan, A.; Levine, A. Programmed cell death of the dinoflagellate Peridinium gatunense is mediated by CO2 limitation and oxidative stress. Curr. Biol. 1999, 9, 1061–1064. [Google Scholar] [CrossRef]
- Alfonso, M.; Perewoska, I.; Kirilovsky, D. Redox control of psbA gene expression in the cyanobacterium Synechocystis PCC 6803. Involvement of the cytochrome b6/f complex. Plant Physiol. 2000, 122, 505–516. [Google Scholar] [CrossRef]
- Zhang, L.; Pei, H.; Chen, S.; Jiang, L.; Hou, Q.; Yang, Z.; Yu, Z. Salinity-induced cellular cross-talk in carbon partitioning reveals starch-to-lipid biosynthesis switching in low-starch freshwater algae. Bioresour. Technol. 2018, 250, 449–456. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Time (d) | 1 ppt | 4 ppt | 7 ppt | 10 ppt | 15 ppt |
|---|---|---|---|---|---|---|
| VJ | 3 | 0.495 ± 0.010 | 0.519 ± 0.017 | 0.589 ± 0.011 * | 0.735 ± 0.035 * | 1.025 ± 0.122 * |
| 6 | 0.489 ± 0.007 | 0.507 ± 0.000 | 0.552 ± 0.011 * | 0.496 ± 0.028 | 0.988 ± 0.016 * | |
| 9 | 0.460 ± 0.008 | 0.498 ± 0.009 * | 0.563 ± 0.009 * | 0.518 ± 0.010 * | −0.061 ± 0.892 * | |
| Mo | 3 | 0.696 ± 0.045 | 0.725 ± 0.038 | 0.792 ± 0.008 * | 0.940 ± 0.090 * | 2.111 ± 0.343 * |
| 6 | 0.659 ± 0.015 | 0.711 ± 0.018 | 0.672 ± 0.005 | 0.621 ± 0.012 | 2.939 ± 0.876 * | |
| 9 | 0.576 ± 0.013 | 0.605 ± 0.009 | 0.655 ± 0.019 * | 0.619 ± 0.034 | −0.463 ± 1.916 * | |
| φPo | 3 | 0.340 ± 0.022 | 0.352 ± 0.013 | 0.374 ± 0.013 | 0.306 ± 0.005 | 0.046 ± 0.036 * |
| 6 | 0.418 ± 0.006 | 0.401 ± 0.014 | 0.464 ± 0.003 * | 0.443 ± 0.050 | 0.012 ± 0.005 * | |
| 9 | 0.4390.010 | 0.506 ± 0.008 * | 0.498 ± 0.018 * | 0.520 ± 0.004 * | 0.011 ± 0.003 * | |
| ψo | 3 | 0.521 ± 0.034 | 0.481 ± 0.017 * | 0.411 ± 0.011 * | 0.331 ± 0.013 * | 0.023 ± 0.002 * |
| 6 | 0.511 ± 0.007 | 0.493 ± 0.000 | 0.448 ± 0.011 * | 0.504 ± 0.028 | 0.009 ± 0.002 * | |
| 9 | 0.540 ± 0.008 | 0.502 ± 0.009 * | 0.437 ± 0.009 * | 0.482 ± 0.010 * | 1.061 ± 0.892 * | |
| φEo | 3 | 0.177 ± 0.019 | 0.169 ± 0.012 | 0.154 ± 0.003 | 0.104 ± 0.002 * | 0.033 ± 0.024 * |
| 6 | 0.214 ± 0.003 | 0.198 ± 0.007 | 0.208 ± 0.004 | 0.245 ± 0.008 * | 0.001 ± 0.000 * | |
| 9 | 0.238 ± 0.009 | 0.254 ± 0.005 | 0.218 ± 0.010 | 0.258 ± 0.009 * | 0.012 ± 0.012 * | |
| ABS/RC | 3 | 4.313 ± 0.425 | 3.968 ± 0.231 | 3.604 ± 0.215 * | 4.742 ± 0.313 * | 16.721 ± 2.611 * |
| 6 | 3.216 ± 0.013 | 3.245 ± 0.165 | 2.627 ± 0.077 * | 2.721 ± 0.050 * | N.D. | |
| 9 | 2.856 ± 0.082 | 2.397 ± 0.084 * | 2.335 ± 0.116 * | 2.164 ± 0.068 * | N.D. | |
| TR0/RC | 3 | 1.456 ± 0.057 | 1.395 ± 0.029 | 1.345 ± 0.039 * | 1.341 ± 0.021 * | 2.050 ± 0.097 * |
| 6 | 1.346 ± 0.016 | 1.408 ± 0.027 | 1.219 ± 0.029 * | 1.279 ± 0.009 * | 1.974 ± 0.036 * | |
| 9 | 1.254 ± 0.016 | 1.214 ± 0.024 | 1.162 ± 0.017 * | 1.195 ± 0.55 | 0.006 ± 0.006 * | |
| ET0/RC | 3 | 0.760 ± 0.068 | 0.670 ± 0.015 * | 0.553 ± 0.031 * | 0.435 ± 0.023 * | 0.203 ± 0.162 * |
| 6 | 0.687 ± 0.008 | 0.709 ± 0.012 | 0.547 ± 0.027 * | 0.692 ± 0.026 | N.D. | |
| 9 | 0.678 ± 0.012 | 0.609 ± 0.022 * | 0.507 ± 0.008 * | 0.576 ± 0.026 * | 0.448 ± 0.633 * | |
| DI0/RC | 3 | 2.857 ± 0.379 | 2.573 ± 0.204 | 2.259 ± 0.179 * | 3.034 ± 0.117 | 64.672 ± 5.485 * |
| 6 | 1.870 ± 0.025 | 1.866 ± 0.158 | 1.408 ± 0.049 * | 1.342 ± 0.086 * | N.D. | |
| 9 | 1.602 ± 0.074 | 1.184 ± 0.060 * | 1.173 ± 0.101 * | 1.302 ± 0.351 | N.D. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Y.; Ma, Z.; Liu, X.; Zheng, R.; Xiao, Y.; Wang, M. The Detrimental Effect of High Salinity on the Growth and Microcystins Contamination of Microcystis aeruginosa. Water 2022, 14, 2871. https://doi.org/10.3390/w14182871
Qiu Y, Ma Z, Liu X, Zheng R, Xiao Y, Wang M. The Detrimental Effect of High Salinity on the Growth and Microcystins Contamination of Microcystis aeruginosa. Water. 2022; 14(18):2871. https://doi.org/10.3390/w14182871
Chicago/Turabian StyleQiu, Yu, Zengling Ma, Xiang Liu, Ranran Zheng, Yang Xiao, and Min Wang. 2022. "The Detrimental Effect of High Salinity on the Growth and Microcystins Contamination of Microcystis aeruginosa" Water 14, no. 18: 2871. https://doi.org/10.3390/w14182871
APA StyleQiu, Y., Ma, Z., Liu, X., Zheng, R., Xiao, Y., & Wang, M. (2022). The Detrimental Effect of High Salinity on the Growth and Microcystins Contamination of Microcystis aeruginosa. Water, 14(18), 2871. https://doi.org/10.3390/w14182871