
Non-Indigenous Cladocera (Crustacea: Branchiopoda): From a Few Notorious Cases to a Potential Global Faunal Mixing in Aquatic Ecosystems




{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods: Literature Study
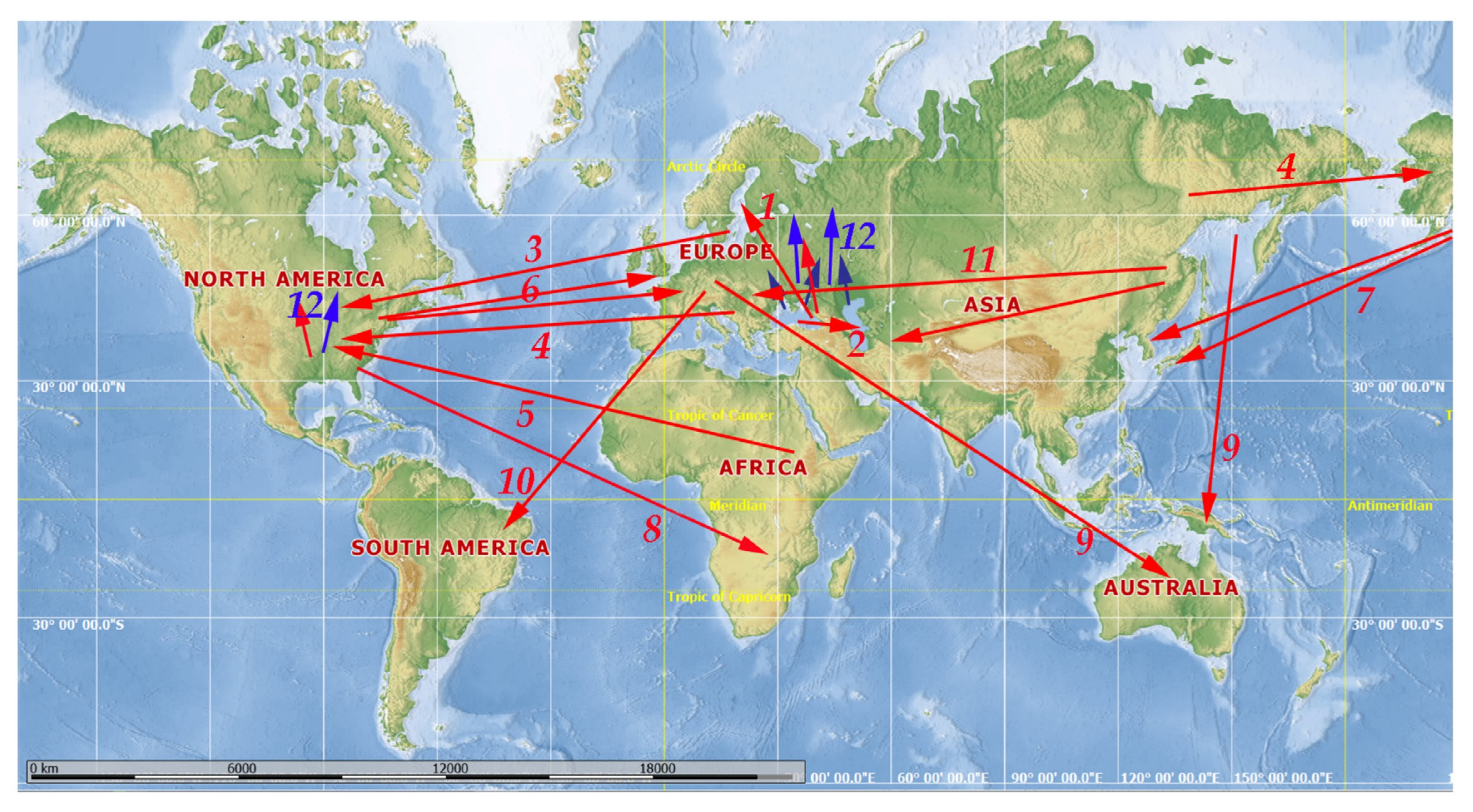
3. Results: Major Pathways of Transportations and Range Expansion
3.1. Introduction of Ponto–Caspian Onychopods in Europe
3.2. Transportation of Oceanic Cladocerans from the Black Sea to the Caspian Sea
3.3. The Baltic Sea Region as a Hub for Intercontinental Transportation and Further Expansion of Invaders in North America
3.4. Other European Invaders in North America
3.5. Transportation of African Cladocerans to North America
3.6. Transportation of North American Taxa to Europe
3.7. Transportation of North American Taxa to the Far East of Asia
3.8. American Taxa in Africa
3.9. Transportation of Several Taxa from the Holarctic to Australia/New Zealand and Subsequent Expansion of Their Ranges
3.10. European Taxa in South America
3.11. Transportation of Far Eastern Taxa to Europe and Middle Asia Due to Aquaculture and Rice Crops
3.12. General Range Expansions towards North and South of Europe and North America Due to Natural Factors Re-Enforced by Human Activity
3.13. Occasional Anthropogenic Transportation of Unpredictable, Chaotic Directionality
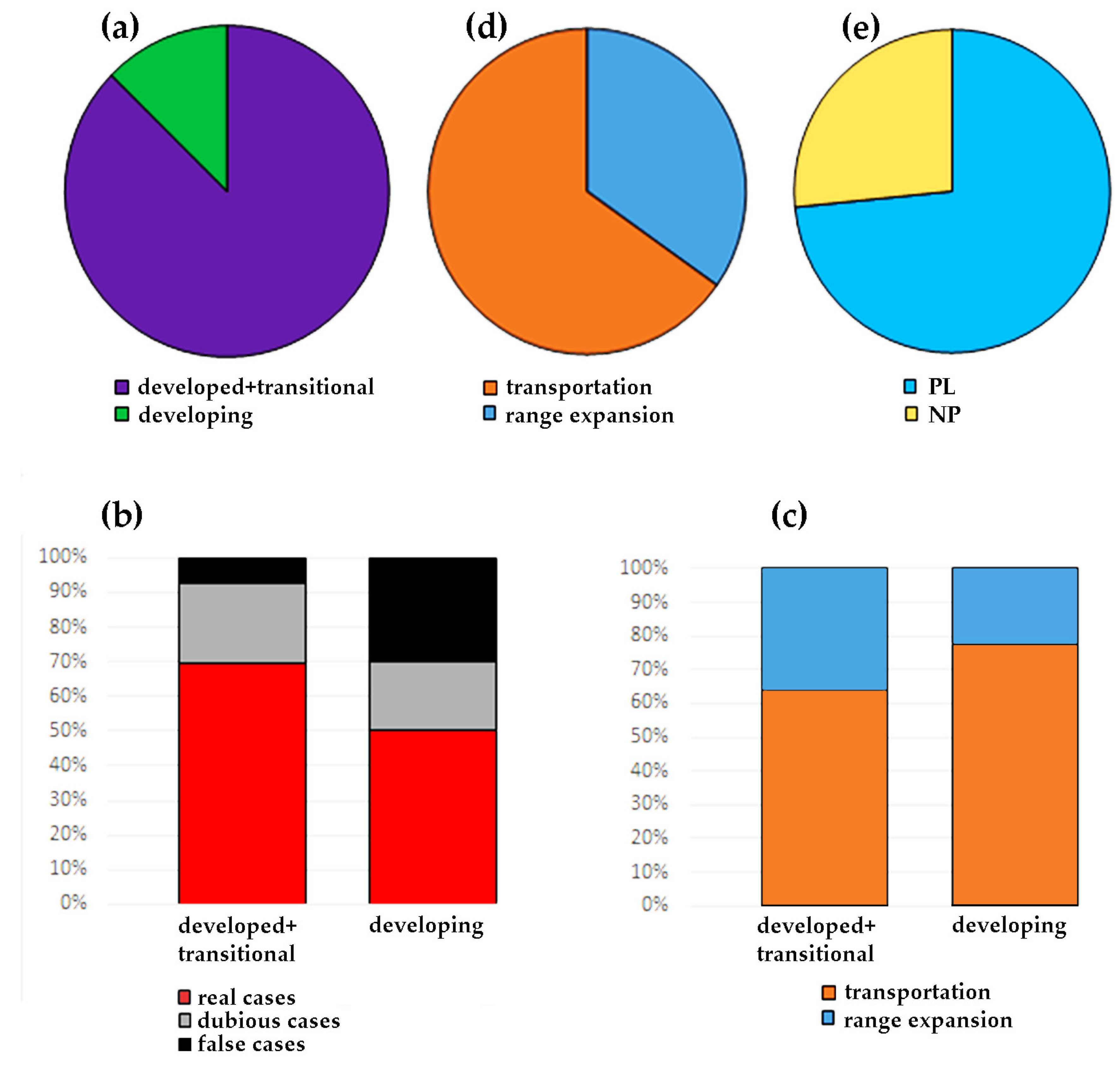
3.14. Developed vs. Developing Countries and More Cases of Planktonic Invaders
4. Discussion: Recent Insights in Cladoceran Exotics
4.1. Dispelling Myths: Some Cases May Not Be Exotics after All
4.2. Next Stage: Understanding Invasions Using Molecular Tools
4.3. Paleolimnological Study as the Final Evidence of a Past Invasion
4.4. General Analysis of Revealed Cases
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| № | TR/RE | PL/NP | Taxon | Region of Non-Indigenous Species Appearance (Recipient Region) | Donor Region | Fist Records of Invasion/Range Expansion | Last Reviews | Our Comments (with Reference to Last Source If We Agree with That Conclusion) |
| CTENOPODA—SIDIDAE | ||||||||
| 1 | TR | PL | Diaphanosoma brevireme Sars, 1901 | Great Lakes | Neotropics and Southern regions of North America | [122] | [130] | The taxon is widely distributed from Argentina to Florida [130], but its appearance in the Great Lakes seems to be a case of anthropogenic transportation (if the identification is correct). |
| 2 | TR | PL | Diaphanosoma dubium Manujlova, 1964 | Kazakhstan, Uzbekistan, lower reaches of Volga, Pre-Caucasus, Ukraine | East Asia | [129,131] | [130,132] | Hypothesis on a recent expansion of the distribution range in the south of European Russia, Ukraine, and Middle Asia is realistic [130,132]. |
| 3 | TR | PL | Diaphanosoma fluviatile Hansen, 1899 | Great Lakes, North America | Neotropical zone | [121] | Anthropogenic transportation (less likely, natural range expansion) [121]. | |
| 4 | TR | PL | Diaphanosoma macrophthalma Korovchinsky et Mirabdullaev, 1995 | Kazakhstan, Uzbekistan, Pre-Caucasus | East Asia | [113] | [130,132] | Possibly, occasionally introduced to Middle Asia due to fish introduction as this species is found predominantly in fishponds [130]. |
| 5 | RE | PL | ? Diaphanosoma mongolianum Ueno, 1938 | Volga basin | more southern regions | [128] | [130] | The ideas on distribution range changes could be a reflection of previous imperfect state of taxonomy. |
| 6 | RE | PL | ? Diaphanosoma orghidani Negrea, 1982 | Volga basin, European Russia | more southern regions | [127,232] | [130] | The ideas on distribution range changes could be a reflection of previous imperfect state of taxonomy. |
| 7 | RE | NP | ? Latonopsis australis Sars, 1888 | Italy | more southern regions | [124] | [84,90] | Conclusion on human introduction is dubious as this rare taxon had a good chance to be missed in previous studies. This “tropical” species is known from some localities in South Europe [130]. |
| 8 | RE | PL | ? Limnosida frontosa Sars, 1862 | Volga, Dniepr basins, and Czech Republic | more eastern regions | [84] | Conclusion on wide range expansion is dubious. The introduction of fish ponds in Czech Republic seems more realistic [130,233]. | |
| CTENOPODA—HOLOPEDIIDAE | ||||||||
| 9 | RE | PL | ? Holopedium gibberum Zaddach, 1855 | Lena Delta | ? | [181] | [179] | Misinterpretation [179]. A new record was made for the region [181], without interpreting the natural range expansion. H. gibberus is very common in North Eurasia. |
| ANOMOPODA—DAPHNIIDAE | ||||||||
| 10 | TR | PL | Daphnia ambigua Scourfield, 1947 | Europe | Eastern North America | [79] | [65,81,82,84] | Human-mediated transportation with subsequent range expansion confirmed genetically [82]. |
| 11 | TR | PL | Daphnia ambigua Scourfield, 1947 | Japan | North America | [93] | [94] | Possible invader from North America, or secondary introduction from Europe. |
| 12 | TR | PL | ? Daphnia barbata Weltner, 1897 = “D. brooksi Dodson, 1985” | Utah (USA) and Michoacán (Mexico) | Africa | [73] | [65] | Possibly an anthropogenic invasion initially from Africa and then North America [73], but this should be checked using molecular methods. |
| 13 | TR | PL | Daphnia curvirostris Eylmann, 1887 | Mexico and Atlantic Coast of USA | Eurasia | [62,63,64] | [65,67] | Anthropogenic transportation from Eurasia confirmed genetically [67]. |
| 14 | TR | PL | Daphnia exilis Herrick, 1895 | New York State (USA) | Southern USA and Mexico | [64,120] | [65] | Anthropogenic transportation ca. 1000 km north of the boundary of its natural distribution range. |
| 15 | RE | PL | ? Daphnia exilis Herrick, 1895 | Chile | North America? | [168,170] | [65,178] | In reality, South American D. spinulata Birabén, 1917 is a very common species in southern South America. It is a close congener of D. exilis [178]. Most probably, the authors mixed the two taxa. It is likely that they have observed colonisation of the single man-made water reservoir by a local species, which is a very common process. |
| 16 | TR | PL | Daphnia galeata Sars, 1863 | North America | Eurasia | [59] | [60] | Anthropogenic transportation and subsequent hybridisation with an indigenous congener confirmed genetically [59]. |
| 17 | TR | PL | Daphnia galeata Sars, 1863 | New Zealand | Eurasia | [101] | Anthropogenic transportation. Initially, the species was identified as D. dentifera Forbes, 1893, but then the identification was improved [15] and confirmed genetically [106]. | |
| 18 | TR | PL | Daphnia galeata Sars, 1863 | South Australia | Eurasia | [106] | Apparently, anthropogenic transportation from Eurasia to eutrophied lakes confirmed genetically [106]. | |
| 19 | TR | PL | Daphnia inopinata Popova et al., 2016 | Germany (single locality) | Unknown | [142] | It is, most probably, a non-indigenous taxon from an unknown locality as it was found in a military training area, but no other locality for this species is known worldwide [142]. | |
| 20 | TR | PL | Daphnia korovchinskyi Kotov et al., 2021 | Samara Area in European Russia | Far East of Russia | [234] | Most probably, transportation of ephippia attached to car wheels. Daphnia korovchinsky has produced hybrids with indigenous D. curvirostris, but the latter went locally extinct [234]. | |
| 21 | TR | PL | ? Daphnia longicephala Hebert, 1977 | Great Lakes (USA) | Australia? | [141] | Very dubious record of inadequately described Australian species from North America, most probably a misidentification. | |
| 22 | TR | PL | Daphnia lumholtzi Sars, 1885 | North America | Eastern hemisphere | [71,72] | [65,73] | Transportation with subsequent great range expansion (next entry), well-studied genetically [72]. |
| 23 | TR | PL | Daphnia lumholtzi Sars, 1885 | Brazil and Argentina (South America) | North America | [74,75] | [76] | Anthropogenic transportation from North American populations confirmed genetically [75,76]. |
| 24 | TR | PL | Daphnia magna Straus, 1820 | North America | Eurasia | [68] | Commercial clone escaped from a laboratory, confirmed genetically [68,192]. | |
| 25 | TR | PL | Daphnia magna Straus, 1820 | Sevan Lake (Armenia) | More southern regions | [143] | [68] | D. magna already existed in Sevan Lake (Armenia) in the past, but, indeed, together with an indigenous haplotype, this site was colonised by a haplotype from Central Russia—this is an apparent case of anthropogenic transportation confirmed genetically [68]. |
| 26 | TR | PL | Daphnia cf. obtusa Kurz, 1874 | Australia | ? | [105] | [65] | Transportation, but the exact donor region is unknown as many D. obtusa-like taxa are known in different continents [235]. |
| 27 | TR | PL | Daphnia parvula Fordyce, 1901 | Western and Southern Europe | North and South America | [83] | [65,81,84] | Transportation by military amphibian vehicles during WWII followed by range expansion in Europe [83,84]. |
| 28 | RE | PL | ? Daphnia pulex (Linnaeus, 1758) | A lake in Masurian LakeDistrict, Poland | ? | [182] | [179] | Misinterpretation by [179]. In reality, these specimens were artificially taken from another water body where they are indigenous and placed in the microcosms. |
| 29 | TR | PL | American Daphnia pulex × pulicaria clone | East Africa, then whole Africa | North America | [96,97] | [23] | Transportation to Rift Valley lakes and then range expansion confirmed genetically [23,97]. |
| 30 | TR | PL | American Daphnia pulex × pulicaria clone | New Zealand | North America | [102] | [103] | Transportation following a rapid range expansion; well-studied genetically. Likely imported by accompanying introduction of salmonid fishes, confirmed genetically [103]. |
| 31 | TR | PL | American Daphnia pulex × pulicaria clone | Japan | North America | [92] | Transportation confirmed by genetic methods [92]. | |
| 32 | RE | PL | American Daphnia pulex × pulicaria clone | Sardinia, North Italy, Spain | North America | [23,98,99] | [100,207] | Range expansion after transportation from North America confirmed by genetic methods [207]. |
| 33 | TR | PL | Daphnia cf. pulicaria Forbes, 1893 | Japan | North America | [91] | Transportation confirmed genetically [91]. | |
| 34 | RE | PL | ? Daphnia sinensis Gu, Xu, Li, Dumont et Han, 2013 | Ethiopia | Ethiopia | [142,171] | [179] | Misinterpretation by [179]. In reality, the authors have studied the colonisation of man-made reservoirs by a local species of the D. similis group, widely distributed in Africa [142]. This is a very common process in artificial water reservoirs due to colonisation by indigenous taxa. |
| 35 | RE | PL | ? Ceriodaphnia dubia Richard, 1894 | Two artificial reservoirs in Brazil | ? | [169] | We consider this a misidentification. Very likely, this is a common case of a Ceriodaphnia appearing in a man-made water reservoir. Ceriodaphnia cf. dubia is an unusual taxon in tropical South America [236]. The species of this genus are very hard to identify. | |
| 36 | RE | PL | ? Ceriodaphnia rotunda Sars, 1862 | Italy | Other countries of Europe? | [124] | [84,90] | Not likely an anthropogenic introduction; this rare European taxon [81] had a good chance to be missed in previous studies due to misidentification. Ceriodaphnia is not easy to identify. |
| 37 | TR | NP | Scapholeberis yahuarcaquensis Andrade-Sossa, Buitron-Caicedo et Elías-Gutiérrez, 2020 | Belgium | South America | [137] | Anthropogenic introduction, revealed genetically [137]. | |
| 38 | TR | NP | ?Simocephalus hejlongjiangensis Shi and Shi, 1994 | Italy | SE Asia | [124] | [90] | This can be a misidentification. Unfortunately, the taxonomy of Simocephalus is outdated now, and any conclusions on the non-indigenous status of this taxon seem premature. |
| 39 | TR | NP | ? Simocephalus vetulus (O.F. Müller, 1776) | New Zealand | ? | [15] | [179] | Misinterpretation by [179]. Ref. [15] originally stated that there is an “uncertain status in New Zealand” for S. vetulus, a species group with naturally occurring representatives in Australasia. |
| 40 | RE | NP | ? Simocephalus vetulus (O.F. Müller, 1776) | Poland | ? | [182] | [179] | Misinterpretation by [179]. In reality, specimens were artificially taken from one water body and placed in a microcosm of another water body. |
| ANOMOPODA—MOINIDAE | ||||||||
| 41 | TR? | PL | ? Moina affinis Birge, 1893 | Italy and Sicily | North America | [88,89] | [84,90] | European populations of M. affinis had a chance to appear due to transportation from North America – it is possible. However, note that M. cf. affinis is present in the Far East of Asia as well [237,238]. Records in Italy and Sicily could belong to a separate taxon or also be another, eastern invasive [239]. The non-indigenous status of European and Far Eastern populations of Moina need to be checked by molecular tools. |
| 42 | RE | PL | Moina micrura Kurz, 1875 | Volga basin, Baltic Sea | More southern regions? | [133] | [134] | Range expansion north. |
| 43 | TR | PL | Moina macrocopa (Straus, 1820) | South America | Eurasia | [109,110] | [69] | Intercontinental transportation confirmed genetically [69]. |
| 44 | TR | PL | Moina macrocopa (Straus, 1820) | Mexico | Eurasia | [69] | [69] | Multiple intercontinental transportations of M. macrocopa macrocopa confirmed genetically [69]. |
| 45 | TR? | PL | ? Moina weismanni Ishikawa, 1896 | Italy and then other countries of Southern Europe | Far East of Asia | [114,115,116] | [84] | European populations have appeared after transportation from East Asia. Numerous populations of this taxon were found in Central Asia [240] and southern portion of European Russia and authors concluded that it could have a very wide natural distribution range in South Asia, reaching South Europe. Moreover, [239] confirmed the status of some populations from European Russia as M. weismanni s.str. The taxon must be studied by phylogeographic/molecular methods. |
| ANOMOPODA—ILYOCRYPTIDAE | ||||||||
| 46 | TR? | NP | Ilyocryptus spinifer Herrick 1882 | Middle Volga River | Far East of Asia | [135] | Possible anthropogenic transportation or natural introduction of ephippia by waterfowl [135]. | |
| ANOMOPODA—MACROTHRICIDAE | ||||||||
| 47 | RE | NP | ? Wlassiscia pannonica Daday, 1904 | Italy | ? | [241] | [84,90] | This rare Macrothrix-like taxon had a good chance to be missed in previous studies. Italy is within its natural range in Europe. |
| ANOMOPODA—BOSMINIDAE | ||||||||
| 48 | TR | PL | Bosmina (Eubosmina) cf. coregoni Baird, 1857 (including B. maritima P. E. Müller, 1867) | North America | Europe (e.g., Baltic) | [55] | [56,57,58] | Transportation from Europe and then range expansion. Different species of B. (Eubosmina) were recorded from North America, but they form a lineage with recent speciation. Introduction is confirmed genetically [61]. |
| 49 | RE | PL | ? Bosmina (Eubosmina) coregoni Baird, 1857 | Slovakia, Volga and Dniepr Rivers basins | Europe | [84] | Such “range expansion” needs to be accurately studied genetically; this is a widespread taxon and different B. (Eubosmina) morphotypes occur in different water bodies independently [61]. | |
| 50 | RE | PL | Bosmina (Eubosmina) coregoni Baird, 1857 | Iberian Peninsula | Europe | [125] | Most probably, it is a true range expansion: penetration into the Iberian Peninsula where B. (Eubosmina) was apparently absent [126]. | |
| 51 | RE | PL | ? Bosmina longirostris (O. F. Müller, 1776) | Lake El Tobar (Spain) | ? | [180] | [179] | Misinterpretation. This is not an anthropogenic invasion, but colonisation of a new water body by a taxon indigenous to the region. |
| 52 | TR | PL | Bosmina longirostris (O. F. Müller, 1776) | New South Wales, Australia | Holarctic | [107] | Possible introduction from an unknown Holarctic locality mediated by fish transportation [107]. | |
| ANOMOPODA—EURYCERCIDAE | ||||||||
| 53 | TR | NP | Eurycercus lamellatus (O.F. Müller, 1776) | Brazil | North Eurasia | [111] | [112] | Transportation from North Eurasia [111]. |
| ANOMOPODA—CHYDORIDAE | ||||||||
| 54 | RE | NP | ? Acroperus harpae (Baird, 1834) | Lena Delta | ? | [181] | [179] | Misinterpretation by [179]. Ref. [181] made a new record for the Lena Delta, but A. harpae is very common in Eurasia [242]. |
| 55 | TR | NP | Alonella excisa (Fischer, 1854) clade I2 | Canada | Pacific Asia | [70] | Possible transportation revealed genetically [70]. | |
| 56 | TR | NP | Biapertura ossiani herricki (Sinev, 2013) | Korea | North America | [95] | Transportation, possibly during the Korean War [95]. | |
| 57 | RE | NP | ? Camptocercus uncinatus Smirnov, 1971 | Italy | Asia | [167] | [84,90] | This rare taxon had a good chance to be missed in previous studies. This taxon is known from Europe so range expansion is likely [218,243]. |
| 58 | TR | NP | Chydorus brevilabris Frey, 1980 | Belgium, Luxembourg, France | North America | [86] | Possible transportation from North America, followed by range expansion. | |
| 59 | TR | NP | Chydorus sphaericus (O.F. Müller, 1776) | Australia | North Atlantic (Iceland, Greenland) | [108] | Anthropogenic transportation confirmed genetically [108]. However, some populations could have appeared before the Europeans by means of long-term transportation by birds [7]. | |
| 60 | RE | NP | Disparalona cf. leei (Chen, 1970) | Slovakia and the Danube River basin | South Europe | [116,158] | [84] | We consider this a range expansion in Europe. Most probably, this is a separate Eurasian taxon instead of North American D. leei s.str. (needs molecular confirmation). |
| 61 | TR | NP | Disparalona striatoides (Šrámek-Hušek 1946) | Europe | Africa | [87,116,138] | [84,139] | The taxon was first described in the Czech Republic [138] but then found to be common in Africa [139]. A suggestion of its introduction from Africa [116] is realistic. |
| 62 | RE | NP | ? “Alona” rustica (Scott, 1895) | Italy | ? | [167] | [84,90] | This rare taxon had a good chance to be missed in previous studies. The species is quite common in Europe but in specific habitats [81,166]. Note: sometimes placed under Flavalona [166]. |
| 63 | RE | NP | ? Leberis diaphanus (King, 1853) | Italy | Tropics of the Old World | [244] | [90] | We consider this a misidentification. Name confusions in Leberis (former Alona diaphana group) are common; North African populations of naturally occurring Leberis can be expected in the Mediterranean; in Africa, L. diaphanus does not occur, but recently revised L. punctatus (Daday, 1898) is widely spread in the Old World [245]. |
| 64 | RE | NP | Pleuroxus cf. denticulatus Birge, 1879 | Slovakia, European Russia | Central Europe | More southern regions? | [158,246] | Ref. [158] speculated that the taxon was possibly introduced in Europe from North America, but this version is dubious as no revision of the P. denticulatus group was performed. Its appearance in Central Europe (i.e. Slovakia and relatively northern localities of European Russia) is most likely a local range expansion of a native species of the P. denticulatus group. |
| 65 | TR | NP | ? Ovalona weinecki (Studer, 1878) | Easter Island | Subantarctic islands | [150] | [156] | Misidentification. In reality, the population from Easter Island belongs to a separate endemic taxon, Ovalona pascua Van Damme, 2016 [156]. |
| 66 | RE | NP | Phreatalona protzi Hartwig, 1900 | Italy | ? | [124] | [84,90] | This rare taxon had a good chance to be missed in previous studies. The species is known from Europe [218] and it lives in unusual habitats; the genus was revised recently, allowing easier identification [165]; Italy is well within its natural range. |
| ONYCHOPODA—CERCOPAGIDIDAE | ||||||||
| 67 | TR | PL | Bythotrephes cederstroemii Schödler, 1877 (earlier misidentified as “B. longimanus”) | Great Lakes and then in other water bodies of North America | North-Western European Russia | [40,43,44] | [6,48,203] | Transportation following a range expansion. Korovchinsky and Arnott [48] concluded that invasive populations belong to a single taxon, B. cederstroemii (not B. longimanus). There are some data on earlier colonisation of the Great Lakes basin [203], but they must be rechecked, the recent invasion version is a more plausible hypothesis so far. |
| 68 | RE | PL | Bythotrephes brevimanus Lilljeborg, 1901 | The Netherlands and Belgium | Circumbaltic region of Europe | [123] | [49,130] | The natural range of this species may have recently expanded westward and colonised water bodies of The Netherlands and Belgium. It may be a natural expansion but there may also be a contribution (or facilitation) by human-mediated transport. |
| 69 | TR | PL | Cercopagis pengoi (Ostroumov 1892) | Baltic Sea and the Neva River Estuary | Ponto-Caspian basin | [19,20] | [32,38,132] | Well-documented invasion. Transportation by ship (in ballast water) confirmed genetically [38]. A clear example of human introduction, also clear appearance of resting eggs in Baltic sediments in the 1990s using paleolimnology. |
| 70 | RE | PL | Cercopagis pengoi (Ostroumov 1892) | Volga, Don, and Dniepr Rivers basins | Black Sea | [9,27] | [32,38,132] | Range expansion confirmed genetically [38], i.e., after artificial connection of earlier isolated river basins. |
| 71 | TR | PL | Cercopagis pengoi (Ostroumov 1892) | North American Great Lakes | Baltic Sea (invasive range) | [54] | [38,132] | Transportation by ships (ballast water) from invasive range in the Baltic Sea confirmed genetically [38]. |
| ONYCHOPODA—PODONIDAE | ||||||||
| 72 | RE | PL | Cornigerius maeoticus (Pengo, 1880) | The Danube, Dnieper, Don, and Volga Rivers | Ponto-Caspian basin | [9,28] | [29,132] | Range expansion. |
| 73 | TR | PL | Cornigerius maeoticus (Pengo, 1880) | Baltic Sea | Ponto-Caspian basin | [25,50,51] | [29,132] | Transportation by ship (in ballast water). According to later records [247], this species has not been observed in the Baltic Sea for years. |
| 74 | TR | PL | Cornigerius bicornis (Zernov, 1901) | The lower reaches of the Dnieper, Don, and Volga Rivers | Ponto-Caspian basin | [29] | [132] | Range expansion. |
| 75 | TR | PL | Evadne anonyx Sars, 1897 | Baltic Sea | Ponto-Caspian basin | [52] | [29,132] | Recent introduction from Ponto–Caspian basin into the Gulf of Finland [52]. |
| 76 | TR | PL | Evadne nordmanni Loven, 1836 | Great Lakes (USA) | World Ocean | [140] | [141] | Transportation by ship (ballast water). This species is also native to the Baltic Sea. |
| 77 | TR | PL | Podon intermedius Lilljeborg, 1853 | Caspian Sea | Black Sea | [36] | Transportation through the Volga–Don Canal by ship (ballast water). | |
| 78 | TR | PL | Pleopis polyphemoides (Leuckart, 1859) | Caspian Sea | Black Sea | [8] | [132] | Transportation through the Volga–Don Canal by ship (ballast water). |
| 79 | TR | PL | Podonevadne trigona (Sars, 1897) | Lower reaches of the Dnieper, Don, and Volga Rivers | Ponto-Caspian basin | [9] | [29,132] | The appearance of this species in Eastern Europe was attributed to transportation, e.g., in Romania [228,248] combined with natural range expansion |
References
- Moyle, P.B.; Light, T. Biological invasions of fresh water: Empirical rules and assembly theory. Biol. Conserv. 1996, 78, 149–161. [Google Scholar] [CrossRef]
- Gallardo, B.; Clavero, M.; I Sánchez, M.; Vilà, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2015, 22, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Pullaiah, T.; Ielmini, M.R. (Eds.) Invasive Alien Species: Observations and Issues from Around the World; Wiley Blackwell: Hoboken, NJ, USA, 2021; ISBN 978-1-119-60702-1. [Google Scholar]
- Cuthbert, R.N.; Pattison, Z.; Taylor, N.G.; Verbrugge, L.; Diagne, C.; Ahmed, D.A.; Leroy, B.; Angulo, E.; Briski, E.; Capinha, C.; et al. Global economic costs of aquatic invasive alien species. Sci. Total Environ. 2021, 775, 145238. [Google Scholar] [CrossRef]
- Leppäkoski, E.; Gollasch, S.; Gruszka, P.; Ojaveer, H.; Olenin, S.; Panov, V. The Baltic—A sea of invaders. Can. J. Fish. Aquat. Sci. 2002, 59, 1175–1188. [Google Scholar] [CrossRef]
- Yan, N.D.; Leung, B.; Lewis, M.A.; Peacor, S.D. The spread, establishment and impacts of the spiny water flea, Bythotrephes longimanus, in temperate North America: A synopsis of the special issue. Biol. Invasions 2011, 13, 2423–2432. [Google Scholar] [CrossRef]
- Karabanov, D.P.; Bekker, E.I.; Garibian, P.G.; Shiel, R.J.; Kobayashi, T.; Taylor, D.J.; Kotov, A.A. Multiple recent colonizations of the Australian Region by the Chydorus sphaericus group (Crustacea: Cladocera). Water 2022, 14, 594. [Google Scholar] [CrossRef]
- Mordukhai-Boltovskoi, P.D. Appearance of a representative of Mediterranean polyphemids in the Caspian Sea. Zool. Zh. 1962, 41, 289–290. [Google Scholar]
- Mordukhai-Boltovskoi, P.D. Caspian Polyphemidae in the Don and Dniepr reservoirs. Tr. Inst. Biol. Vnutr. Vod. AN SSSR 1965, 8, 37–43. [Google Scholar]
- Banta, A.M. Studies on the physiology, genetics, and evolution of some Cladocera. Carnegie Inst. Washington 1939, 27, 1–285. [Google Scholar]
- Kotov, A.A. Morphology and Phylogeny of the Anomopoda (Crustacea: Cladocera); KMK Scientific Press Ltd.: Moscow, Russia, 2013; ISBN 9785873179237. [Google Scholar]
- Gollasch, S.; Macdonald, E.; Belson, S.; Botnen, H.; Christensen, J.T.; Hamer, J.P.; Houvenaghel, G.; Jelmert, A.; Lucas, I.; Masson, D.; et al. Life in ballast tanks. In Invasive Aquatic Species of Europe. Distribution, Impacts and Management; Leppakoski, E., Gollasch, S., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 217–231. ISBN 978-90-481-6111-9. [Google Scholar]
- Incagnone, G.; Marrone, F.; Barone, R.; Robba, L.; Naselli-Flores, L. How do freshwater organisms cross the “dry ocean”? A review on passive dispersal and colonization processes with a special focus on temporary ponds. Hydrobiologia 2014, 750, 103–123. [Google Scholar] [CrossRef]
- Figuerola, J.; Green, A.J.; Michot, T.C. Invertebrate Eggs Can Fly: Evidence of Waterfowl-Mediated Gene Flow in Aquatic Invertebrates. Am. Nat. 2005, 165, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Duggan, I.C.; Pullan, S.G. Do freshwater aquaculture facilities provide an invasion risk for zooplankton hitchhikers? Biol. Invasions 2017, 19, 307–314. [Google Scholar] [CrossRef]
- Swaffar, S.M.; O’Brien, W. Spines of Daphnia lumholtzi create feeding difficulties for juvenile bluegill sunfish (Lepomis macrochirus). J. Plankton Res. 1996, 18, 1055–1061. [Google Scholar] [CrossRef] [Green Version]
- Havens, K.E.; Beaver, J.R.; East, T.L.; Work, K.; Phlips, E.J.; Cichra, M.F.; Croteau, A.C.; Rodusky, A.J.; Fulton, R.S.; Rosati, T.C. The outcome of the invasion of Florida lakes by Daphnia lumholtzi. Freshw. Biol. 2012, 57, 552–562. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S. 100 of the World’s Worst Invasive Alien Species a Selection from the Global Invasive Species Database. Available online: http://www.iucngisd.org/gisd/100_worst.php (accessed on 7 June 2022).
- Ojaveer, H.; Lumberg, A. On the role of Cercopagis (Cercopagis) pengoi (Ostroumov) in Parnu Bay and the NE part of the Gulf of Riga ecosystem. Proc. Est. Acad. Sci. Ecol. 1995, 5, 20–25. [Google Scholar]
- Krylov, P.I.; Panov, V.E. Resting eggs in the life cycle of Cercopagis pengoi, a recent invader of the Baltic Sea. Arch. Hydrobiol. 1998, 52, 383–392. [Google Scholar]
- Antsulevich, A.; Valipakka, P. Cercopagis pengoi—New important food object of the Baltic Herring in the Gulf of Finland. Int. Rev. Hydrobiol. 2000, 85, 609–619. [Google Scholar] [CrossRef]
- Telesh, I.V.; Ojaveer, H. The predatory water flea Cercopagis pengoi in the Baltic Sea: Invasion history, distribution and implications to ecosystem dynamics. In Invasive Aquatic Species of Europe. Distribution, Impacts and Management; Leppakoski, E., Gollasch, S., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 62–65. ISBN 978-90-481-6111-9. [Google Scholar]
- Mergeay, J.; Verschuren, D.; De Meester, L. Invasion of an asexual American water flea clone throughout Africa and rapid displacement of a native sibling species. Proc. R. Soc. B Boil. Sci. 2006, 273, 2839–2844. [Google Scholar] [CrossRef]
- Marble Developers. Marble Windows Desktop v.2.2; KDE Family: 2022. Available online: https://marble.kde.org/ (accessed on 1 June 2022).
- Rivier, I.K. The success of the study of the family Polyphemoidae (Cladocera, Onychopoda) in the Laboratory of Ecology of Aquatic Invertebrates. In Biological Resources of Fresh Water: Invertebrates; Rivier, I.K., Shcherbina, G.K., Krylov, A.V., Eds.; Rybinsk Publishers House: Rybinsk, Russia, 2005. [Google Scholar]
- Cristescu, M.E.A.; Hebert, P.D.N. Phylogeny and adaptive radiation in the Onychopoda (Crustacea, Cladocera): Evidence from multiple gene sequences. J. Evol. Biol. 2002, 15, 838–849. [Google Scholar] [CrossRef]
- Valkanov, A. Investigation on Cercopagis pengoi (Ostr.) (Cladocera, Polyphemidae). Tr. Morsk. Biol. Sta. (Stalin) 1950, 16, 65–83. [Google Scholar]
- Pidgaiko, M.L. Zooplankton of Pre-Danube Water Bodies; Izdatelstvo AN SSSR: Kiev, Ukraine, 1957. [Google Scholar]
- Panov, V.E.; Rodionova, N.V.; Bolshagin, P.V.; Bychek, E.A. Invasion biology of Ponto-Caspian onychopod cladocerans (Crustacea: Cladocera: Onychopoda). Hydrobiologia 2007, 590, 3–14. [Google Scholar] [CrossRef]
- Lazareva, V.I. New Records of the Ponto-Caspian Predatory Cladoceran Cercopagis pengoi (Ostroumov, 1891) (Crustacea, Onychopoda) in the Water Reservoirs of the Kama and Volga Rivers. Inland Water Biol. 2019, 12, 290–297. [Google Scholar] [CrossRef]
- Dumonh, H.J.; Negrea, S. Introduction to the Class Branchiopoda. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J., Ed.; Backhuys Publ.: Leiden, The Netherlands, 2002; ISBN 9057821125. [Google Scholar]
- Dgebuadze, Y.Y.; Petrosyan, V.G.; Khlyap, L.A. (Eds.) The Most Dangerous Invasive Species in Russia (TOP-100); KMK Scientific press Ltd.: Russia, Moscow, 2018; ISBN 978-5-907099-53-1. [Google Scholar]
- Mineeva, N.; Lazareva, V.; Litvinov, A.; Stepanova, I.; Chuiko, G.; Papchenkov, V.; Korneva, L.; Scherbina, G.; Pryanichnikova, E.; Perova, S.; et al. The Volga River. In Rivers of Europe, 2nd ed.; Tockner, K., Zarfl, C., Robinson, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 27–79. ISBN 9780081026120. [Google Scholar]
- Soto, I.; Cuthbert, R.N.; Ricciardi, A.; Ahmed, D.A.; Altermatt, F.; Archambaud-Suard, G.; Bonada, N.; Canedo-Arguelles, M.; Csabai, Z.; Datry, T.; et al. The Faunal Ponto-Caspianization of European Waterways. Available online: https://www.researchsquare.com/article/rs-1702320/v1 (accessed on 5 July 2022).
- Lazareva, V.I. Spreading of Alien Zooplankton Species of Ponto-Caspian Origin in the Reservoirs of the Volga and Kama Rivers. Russ. J. Biol. Invasions 2019, 10, 328–348. [Google Scholar] [CrossRef]
- Kurashova, E.K.; Tinenkova, D.H.; Elyzarenko, M.M. Podon intermedius (Cladocera, Podonodae) in the Caspian Sea. Zoologičeskij Žurnal 1992, 71, 135–137. [Google Scholar]
- Kodukhova, Y.V.; Karabanov, D.P. Finding of Longtail Dwarf Goby Knipowitschia longecaudata (Actinopterygii: Gobiidae) in the Upstream Part of Unregulated Section of the Volga River. Inland Water Biol. 2021, 14, 620–625. [Google Scholar] [CrossRef]
- Cristescu, M.; Hebert, P.; Witt, J.D.S.; MacIsaac, H.J.; Grigorovich, I.A. An invasion history for Cercopagis pengoi based on mitochondrial gene sequences. Limnol. Oceanogr. 2001, 46, 224–229. [Google Scholar] [CrossRef]
- Bailey, S.A. An overview of thirty years of research on ballast water as a vector for aquatic invasive species to freshwater and marine environments. Aquat. Ecosyst. Health Manag. 2015, 18, 261–268. [Google Scholar] [CrossRef]
- Bur, M.T.; Klarer, D.M.; Krieger, K. First Records of a European Cladoceran, Bythotrephes cederstroemi, in Lakes Erie and Huron. J. Great Lakes Res. 1986, 12, 144–146. [Google Scholar] [CrossRef]
- NAS. Nonindigenous Aquatic Species Database. Available online: http://nas.er.usgs.gov (accessed on 7 June 2022).
- Berg, D.J.; Garton, D.W.; Macisaac, H.J.; Panov, V.E.; Telesh, I.V. Changes in genetic structure of North American Bythotrephes populations following invasion from Lake Ladoga, Russia. Freshw. Biol. 2002, 47, 275–282. [Google Scholar] [CrossRef]
- Johannsson, O.E.; Mills, E.L.; O’Gorman, R. Changes in the Nearshore and Offshore Zooplankton Communities in Lake Ontario: 1981–88. Can. J. Fish. Aquat. Sci. 1991, 48, 1546–1557. [Google Scholar] [CrossRef]
- Lange, C.; Cap, R. Bythotrephes cederstroemi (Schödler). (Cercopagidae: Cladocera): A New Record for Lake Ontario. J. Great Lakes Res. 1986, 12, 142–143. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Redescription of Bythotrephes longimanus Leydig, 1860 and B. cederströmii Schödler, 1877 (Crustacea: Cladocera: Onychopoda), with notes on the morphology and systematics of the genus Bythotrephes Leydig, 1860. Zootaxa 2015, 3955, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Korovchinsky, N.M. Further revision of the genus Bythotrephes Leydig (Crustacea: Cladocera: Onychopoda): Redescription of B. brevimanus Lilljeborg, reevaluation of B. cederströmii Schödler, and description of a new species of the genus. Zootaxa 2018, 4379, 347–387. [Google Scholar] [CrossRef] [PubMed]
- Korovchinsky, N.M. Morphological assessment of the North Eurasian interspecific hybrid forms of the genus Bythotrephes Leydig, 1860 (Crustacea: Cladocera: Cercopagididae). Zootaxa 2019, 4550, 340–356. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; E Arnott, S. Taxonomic resolution of the North American invasive species of the genus Bythotrephes Leydig, 1860 (Crustacea: Cladocera: Cercopagididae). Zootaxa 2019, 4691, 125–138. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Description of a new species in the genus Bythotrephes Leydig, 1860 (Crustacea: Cladocera: Onychopoda), supplements to selected species, and concluding remarks on the genus. Zootaxa 2020, 4789, 441–465. [Google Scholar] [CrossRef] [PubMed]
- Litvinchuk, L.F.; Maksimova, O.B. On the discovery of the Ponto-Azov crustacean Cornigerius maeoticus maeoticus (Pengo, 1879) (Cladocera, Polyphemoidea, Podonidae) in the Gulf of Finland of the Baltic Sea. In Biological Resources of Fresh Water: Invertebrates; Rivier, I.K., Shcherbina, G.K., Krylov, A.V., Eds.; Rybinsk Publishers House: Rybinsk, Russia, 2005; pp. 249–251. [Google Scholar]
- Rodionova, N.V.; Krylov, P.I.; Panov, V.E. Invasion of the Ponto-Caspian predatory Cladoceran Cornigerius maeoticus maeoticus (Pengo, 1879) into the Baltic Sea. Oceanology 2005, 45, 73–75. [Google Scholar]
- Litvinchuk, L.F. Evadne anonyx Sars, 1897 (Cladocera, Polyphemoidea, Podonidae)—A new representative of the Baltic Sea fauna. In Biological Resources of Fresh Water: Invertebrates; Rivier, I.K., Shcherbina, G.K., Krylov, A.V., Eds.; Rybinsk Publishers House: Rybinsk, Russia, 2005; pp. 240–248. [Google Scholar]
- Korovchinsky, N.M.; Kotov, A.A.; Boikova, O.S.; Smirnov, N.N. Water Fleas (Crustacea: Cladocera) of Northern Eurasia; KMK Scientific Press Ltd.: Moscow, Russia, 2021; Volume 1, ISBN 978-5-907372-27-6. [Google Scholar]
- MacIsaac, H.J.; Grigorovich, I.A.; Hoyle, J.A.; Yan, N.D.; Panov, V.E. Invasion of Lake Ontario by the Ponto–Caspian predatory cladoceran Cercopagis pengoi. Can. J. Fish. Aquat. Sci. 1999, 56, 1–5. [Google Scholar] [CrossRef]
- Lieder, U. The Bosmina kessleri-like morphotype of Eubosmina in Lake Muskoka, Ontario, Canada, as putative interspecific hybrids. Hydrobiologia 1991, 225, 71–80. [Google Scholar] [CrossRef]
- De Melo, R.; Hebert, P.D.N. A taxonomic reevaluation of North American Bosminidae. Can. J. Zool. 1994, 72, 1808–1825. [Google Scholar] [CrossRef]
- Taylor, D.J.; Ishikane, C.R.; Haney, R.A. The systematics of Holarctic bosminids and a revision that reconciles molecular and morphological evolution. Limnol. Oceanogr. 2002, 47, 1486–1495. [Google Scholar] [CrossRef]
- Beaver, J.R.; Renicker, T.R.; Tausz, C.E.; Vitanye, B.T. Distribution of six taxa in the family Bosminidae Baird (Crustacea: Branchiopoda: Anomopoda) in the plankton of lakes and reservoirs within the continental United States, including expanded range of the invasive cladoceran Bosmina (Eubosmina) coregoni Baird. Zootaxa 2018, 4407, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.J.; Hebert, P.D.N. Cryptic intercontinental hybridization in Daphnia (Crustacea): The ghost of introductions past. Proc. R. Soc. B Boil. Sci. 1993, 254, 163–168. [Google Scholar] [CrossRef]
- Ishida, S.; Taylor, D.J. Quaternary diversification in a sexual Holarctic zooplankter, Daphnia galeata. Mol. Ecol. 2007, 16, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Faustova, M.; Sacherovà, V.; Svensson, J.-E.; Taylor, D.J. Radiation of European Eubosmina (Cladocera) from Bosmina (E) longispina -concordance of multipopulation molecular data with paleolimnology. Limnol. Oceanogr. 2011, 56, 440–450. [Google Scholar] [CrossRef]
- Duffy, M.; Perry, L.J.; Kearns, C.M.; Weider, L.J.; Hairston, N.G. Paleogenetic evidence for a past invasion of Onondaga Lake, New York, by exotic Daphnia curvirostris using mtDNA from dormant eggs. Limnol. Oceanogr. 2000, 45, 1409–1414. [Google Scholar] [CrossRef]
- Nandini, S.; Silva-Briano, M.; García, G.G.; Sarma, S.; Adabache-Ortiz, A.; de la Rosa, R.G. First record of the temperate species Daphnia curvirostris Eylmann, 1887 emend. Johnson, 1952 (Cladocera: Daphniidae) in Mexico and its demographic characteristics in relation to algal food density. Limnology 2009, 10, 87–94. [Google Scholar] [CrossRef]
- Hairston, N.G.; Kearns, C.M.; Demma, L.P.; Effler, S.W. Species-specific daphnia phenotypes: A history of industrial pollution and pelagic ecosystem response. Ecology 2005, 86, 1669–1678. [Google Scholar] [CrossRef] [Green Version]
- Benzie, J.A.H. The Genus Daphnia (Including Daphniopsis): Anomopoda: Daphniidae; Kenobi Productions: Ghent, Belgium, 2005; ISBN 9057821516. [Google Scholar]
- Kotov, A.A.; Taylor, D.J. Contrasting endemism in pond-dwelling cyclic parthenogens: The Daphnia curvirostris species group (Crustacea: Cladocera). Sci. Rep. 2019, 9, 6812. [Google Scholar] [CrossRef]
- Kotov, A.A.; Garibian, P.G.; Bekker, E.I.; Taylor, D.J.; Karabanov, D.P. A new species group from the Daphnia curvirostris species complex (Cladocera: Anomopoda) from the eastern Palaearctic: Taxonomy, phylogeny and phylogeography. Zool. J. Linn. Soc. 2020, 191, 772–822. [Google Scholar] [CrossRef]
- Bekker, E.I.; Karabanov, D.P.; Galimov, Y.R.; Haag, C.R.; Neretina, T.V.; Kotov, A.A. Phylogeography of Daphnia magna Straus (Crustacea: Cladocera) in Northern Eurasia: Evidence for a deep longitudinal split between mitochondrial lineages. PLoS ONE 2018, 13, e0194045. [Google Scholar] [CrossRef]
- Montoliu-Elena, L.; Elías-Gutiérrez, M.; Silva-Briano, M. Moina macrocopa (Straus, 1820): A species complex of a common Cladocera, highlighted by morphology and DNA barcodes. Limnetica 2019, 38, 253–277. [Google Scholar] [CrossRef]
- Neretina, A.N.; Karabanov, D.P.; Sacherova, V.; Kotov, A.A. Unexpected mitochondrial lineage diversity within the genus Alonella Sars, 1862 (Crustacea: Cladocera) across the Northern Hemisphere. PeerJ 2021, 9, e10804. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, K.H.; Sterner, R. Extreme cyclomorphosis in Daphnia lumholtzi. Freshw. Biol. 1992, 28, 257–262. [Google Scholar] [CrossRef]
- Havel, J.E.; Hebert, P.D.N. Daphnia lumholtzi in North America: Another exotic zooplankter. Limnol. Oceanogr. 1993, 38, 1823–1827. [Google Scholar] [CrossRef]
- Silva-Briano, M.; Arroyo-Bustos, G.; Beltran-Alvarez, R.; Adabache-Ortiz, A.; Galvan de la Rosa, R. Daphnia Ctenodaphnia lumholtzi G.O.Sars, 1885 (Crustacea: Cladocera); an exotic cladoceran in Mexico. Hidrobiologica 2010, 20, 275–280. [Google Scholar]
- Zanata, L.H.; Espíndola, E.L.G.; Rocha, O.; Pereira, R.H.G. First record of Daphnia lumholtzi (Sars, 1885), exotic cladoceran, in São Paulo State (Brazil). Braz. J. Biol. 2003, 63, 717–720. [Google Scholar] [CrossRef]
- Kotov, A.A.; Taylor, D.J. Daphnia lumholtzi Sars, 1885 (Cladocera: Daphniidae) invades Argentina. J. Limnol. 2014, 73. [Google Scholar] [CrossRef]
- Nunes, A.H.; Miracle, M.R.; Dias, J.; Fabrin, T.M.C.; De Braghin, L.S.M.; Bonecker, C.C. First genetic characterization of non-native Daphnia lumholtzi Sars, 1885 in Brazil confirms North American origin. Int. Rev. Hydrobiol. 2018, 103, 48–53. [Google Scholar] [CrossRef]
- Dobberfuhl, D.R.; Elser, J.J. Distribution and Potential Competitive Effects of an Exotic zooplankter (Daphnia lumholtzi) in Arizona reservoirs. J. Arizona-Nevada Acad. Sci. 2002, 34, 89–94. [Google Scholar]
- Kolar, C.S.; Boase, J.C.; Clapp, D.F.; Wahl, D.H. Potential Effect of Invasion by an Exotic Zooplankter, Daphnia Iumholtzi. J. Freshw. Ecol. 1997, 12, 521–530. [Google Scholar] [CrossRef]
- Scourfield, D.J. A short-spined Daphnia presumably belonging to the ‘longispina’ group—D. ambigua sp. J. Quekett Microsc. Club 1947, 4, 127–131. [Google Scholar]
- Harding, J.P. The Genus Daphnia. Nature 1959, 183, 93–94. [Google Scholar] [CrossRef]
- Flossner, D. Die Haplopoda Und Cladocera (Ohne Bosminidae) Mitteleuropas; Backhuys: Leiden, The Netherlands, 2000; ISBN 9057820579. [Google Scholar]
- Hebert, P.; Witt, J.D.S.; Adamowicz, S.J. Phylogeographical patterning in Daphnia ambigua: Regional divergence and intercontinental cohesion. Limnol. Oceanogr. 2003, 48, 261–268. [Google Scholar] [CrossRef]
- Kraus, K.; Flössner, D. Zwei Fur Mitteleuropa Neue Cladoceren-Arten (Daphnia ambigua Scourfield, 1946, Und Daphnia parvula Fordyce, 1901) Aus Süddeutschland. Crustaceana 1976, 30, 301–309. [Google Scholar] [CrossRef]
- Bledzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe: Cladocera & Copepoda (Calanoida, Cyclopoida). Key to Species Identification, with Notes on Ecology, Distribution, Methods and Introduction to Data Analysis; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-29870-2. [Google Scholar]
- CABI. Invasive Species Compendium: Daphnia parvula. Available online: https://www.cabi.org/isc/datasheet/113798 (accessed on 7 January 2022).
- Dohet, A.; Hoffmann, L. Seasonal succession and spatial distribution of the zooplankton community in the reservoir of Esch-sur-Sure (Luxembourg). Belg. J. Zool. 1995, 125, 109–123. [Google Scholar]
- Van Damme, K.; Cart, J.F. Extra vigilance required for an exotic chydorid in Europe. Cladocera News 2011, 1, 12–14. [Google Scholar]
- Morroni, A. L’Ecosistema di Risaia; Ente Nazionale Risi: Milano, Italy, 1962. [Google Scholar]
- Goulden, C.E. The Systematics and Evolution of the Moinidae. Trans. Am. Philos. Soc. 1968, 58, 1–101. [Google Scholar] [CrossRef]
- Gherardi, F.; Bertolino, S.; Bodon, M.; Casellato, S.; Cianfanelli, S.; Ferraguti, M.; Lori, E.; Mura, G.; Nocita, A.; Riccardi, N.; et al. Animal xenodiversity in Italian inland waters: Distribution, modes of arrival, and pathways. Biol. Invasions 2007, 10, 435–454. [Google Scholar] [CrossRef]
- Urabe, J.; Ishida, S.; Nishimoto, M.; Weider, L.J. Daphnia pulicaria, a zooplankton species that suddenly appeared in 1999 in the offshore zone of Lake Biwa. Limnology 2003, 4, 35–41. [Google Scholar] [CrossRef]
- So, M.; Ohtsuki, H.; Makino, W.; Ishida, S.; Kumagai, H.; Yamaki, K.G.; Urabe, J. Invasion and molecular evolution of Daphnia pulex in Japan. Limnol. Oceanogr. 2015, 60, 1129–1138. [Google Scholar] [CrossRef]
- Hanazato, T.; Yasuno, M. Occurrence of Daphnia ambigua Scourfield in Lake Kasumigaura. Jpn. J. Limnol. (Rikusuigaku Zasshi) 1985, 46, 212–214. [Google Scholar] [CrossRef]
- Makino, W.; Maruoka, N.; Nakagawa, M.; Takamura, N. DNA barcoding of freshwater zooplankton in Lake Kasumigaura, Japan. Ecol. Res. 2017, 32, 481–493. [Google Scholar] [CrossRef]
- Kotov, A.A.; Sinev, A.Y.; Garibian, P.G.; Neretina, A.N.; Jeong, H.G.; Lee, W.; Chae, K.-S.; Min, G.-S. Recent progress in studies of the Cladocera (Crustacea: Branchiopoda) of South Korea with seven new records for the Korean Peninsula. J. Species Res. 2017, 6, 227–246. [Google Scholar] [CrossRef]
- Mergeay, J.; Verschuren, D.; De Meester, L. Daphnia species diversity in Kenya, and a key to the identification of their ephippia. Hydrobiologia 2005, 542, 261–274. [Google Scholar] [CrossRef]
- Mergeay, J.; Verschuren, D.; De Meester, L. Cryptic invasion and dispersal of an American Daphnia in East Africa. Limnol. Oceanogr. 2005, 50, 1278–1283. [Google Scholar] [CrossRef]
- Fadda, A.; Marková, S.; Kotlík, P.; Lugliè, A.; Padedda, B.M.; Buscarinu, P.; Sechi, N.; Manca, M. First record of planktonic crustaceans in Sardinian reservoirs. Biologia 2011, 66, 856–865. [Google Scholar] [CrossRef]
- Vergilino, R.; Markova, S.; Ventura, M.; Manca, M.; Dufresne, F. Reticulate evolution of the Daphnia pulex complex as revealed by nuclear markers. Mol. Ecol. 2011, 20, 1191–1207. [Google Scholar] [CrossRef]
- Burillo, J.P.; Jiménez, L.; Pérez-Martínez, C. Identifying invasive Daphnia species by morphological analysis of postabdominal claws in Sierra Nevada alpine lakes. J. Paleolimnol. 2019, 62, 121–135. [Google Scholar] [CrossRef]
- Duggan, I.C.; Green, J.D.; Burger, D.F. First New Zealand records of three non-indigenous Zooplankton species: Skistodiaptomus pallidus, Sinodiaptomus valkanovi, and Daphnia dentifera. N. Z. J. Mar. Freshw. Res. 2006, 40, 561–569. [Google Scholar] [CrossRef]
- Duggan, I.; Robinson, K.; Burns, C.; Banks, J.; Hogg, I.D. Identifying invertebrate invasions using morphological and molecular analyses: North American Daphnia ‘pulex’ in New Zealand fresh waters. Aquat. Invasions 2012, 7, 585–590. [Google Scholar] [CrossRef]
- Ye, Z.; Williams, E.; Zhao, C.; Burns, C.W.; Lynch, M. The rapid, mass invasion of New Zealand by North American Daphnia “pulex”. Limnol. Oceanogr. 2021, 66, 2672–2683. [Google Scholar] [CrossRef]
- Duggan, I.C.; Özkundakci, D.; David, B.O. Long-term zooplankton composition data reveal impacts of invasions on community composition in the Waikato lakes, New Zealand. Aquat. Ecol. 2020, 55, 1127–1142. [Google Scholar] [CrossRef]
- Benzie, J.A.H.; Hodges, A.M.A. Daphnia obtusa Kurz, 1874 emend Scourfield, 1942 from Australia. Hydrobiologia 1996, 333, 195–199. [Google Scholar] [CrossRef]
- Karabanov, D.P.; Bekker, E.I.; Shiel, R.J.; Kotov, A.A. Invasion of a Holarctic planktonic cladoceran Daphnia galeata Sars (Crustacea: Cladocera) in the Lower Lakes of South Australia. Zootaxa 2018, 4402, 136–148. [Google Scholar] [CrossRef]
- Garibian, P.G.; Kotov, A.A. Bosmina longirostris (O.F. Müller, 1785) (Anomopoda: Cladocera) in Australia: A new case of the anthropogenic introduction of a boreal waterflea. Invertebr. Zool. 2021, 18, 240–246. [Google Scholar] [CrossRef]
- Sharma, P.; Kotov, A.A. Establishment of Chydorus sphaericus (O.F. Muller, 1785) (Crustacea: Cladocera) in Australia: Consequences of mass fish stocking from Northern Europe? J. Limnol. 2015, 74, 225–233. [Google Scholar] [CrossRef]
- Elias-Gutierrez, M.; Zamuriano-Claros, R. Primer registro de Moina macrocopa (Daphniiformes: Moinidae) en Bolivia. Rev. Biol. Trop. 1994, 42, 385. [Google Scholar]
- Paggi, J.C. Moina Macrocopa (Straus, 1820) (Branchiopoda, Anomopoda) in South America: Another Case of Species Introduction? Crustaceana 1997, 70, 886–893. [Google Scholar] [CrossRef]
- Bekker, E.I.; Kotov, A.; Elmoor-Loureiro, L. The genus Eurycercus Baird, 1843 (Cladocera: Eurycercidae) in the Neotropics. J. Nat. Hist. 2010, 44, 2481–2508. [Google Scholar] [CrossRef]
- Kotov, A.A.; Bekker, E.I. Cladocera: Family Eurycercidae (Branchiopoda. Cladocera. Anomopoda); Backhuys Publishers-Margraf Publishers GmbH: Weikersheim, Germany, 2016; ISBN 9783823617310. [Google Scholar]
- Korovchinsky, N.M.; Mirabdullaev, I.M. A new species of the genus Diaphanosoma Fischer, 1850 (Crustacea: Daphniiformes: Sididae) from Central Asia and China. Hydrobiologia 1995, 304, 235–242. [Google Scholar] [CrossRef]
- Margaritora, F.G.; Ferrari, I.; Crosetti, D. A Far East Moina, M. weismanni Ishikawa, 1896 found in an Italian ricefield. Hydrobiologia 1987, 145, 93–103. [Google Scholar] [CrossRef]
- Hudec, I. Moina weismanni Ishikawa, 1896 (Cladocera, Moinidae) in Central Europe. Hydrobiologia 1990, 190, 33–42. [Google Scholar] [CrossRef]
- Hudec, I. Anomopoda, Ctenopoda, Haplopoda, Onychopoda (Crustacea: Branchiopoda); VEDA: vydavateľtvo Slovenskej akademie vied: Bratislava, Slovakia, 2010; ISBN 9788022411417. [Google Scholar]
- Karabanov, D.P.; Kodukhova, Y.V.; Pashkov, A.N.; Reshetnikov, A.N.; Makhrov, A.A. “Journey to the West”: Three Phylogenetic Lineages Contributed to the Invasion of Stone Moroko, Pseudorasbora parva (Actinopterygii: Cyprinidae). Russ. J. Biol. Invasions 2021, 12, 67–78. [Google Scholar] [CrossRef]
- Karabanov, D.P.; Bekker, E.I.; Pavlov, D.D.; Borovikova, E.A.; Kodukhova, Y.V.; Kotov, A.A. New Sets of Primers for DNA Identification of Non-Indigenous Fish Species in the Volga-Kama Basin (European Russia). Water 2022, 14, e437. [Google Scholar] [CrossRef]
- Voroshilova, I.S.; Pryanichnikova, E.G.; Prokin, A.A.; Sabitova, R.Z.; Karabanov, D.P.; Pavlov, D.D.; Kurina, E.M. Morphological and Genetic Traits of the First Invasive Population of the Asiatic Clam Corbicula fluminea (O.F. Müller, 1774) Naturalized in the Volga River Basin. Russ. J. Biol. Invasions 2021, 12, 36–43. [Google Scholar] [CrossRef]
- Hairston, N.G.; Perry, L.J.; Bohonak, A.J.; Fellows, M.Q.; Kearns, C.M.; Engstrom, D.R. Population biology of a failed invasion: Paleolimnology of Daphnia exilis in upstate New York. Limnol. Oceanogr. 1999, 44, 477–486. [Google Scholar] [CrossRef]
- Whitmore, E.A.; Connolly, J.K.; Van Damme, K.; Watkins, J.M.; Hinchey, E.K.; Rudstam, L.G. First record of the Neotropical cladoceran Diaphanosoma fluviatile in the Great Lakes basin. BioInvasions Rec. 2019, 8, 614–622. [Google Scholar] [CrossRef]
- U.S. Geological Survey. Diaphanosoma Brevireme. Available online: https://nas.er.usgs.gov/queries/collectioninfo.aspx?SpeciesID=2649 (accessed on 4 June 2022).
- Ketelaars, H.A.M.; Gille, L. Range extension of the predatory cladoceran Bythotrephes longimanus Leydig 1860 (Crustacea, Onychopoda) in Western Europe. Neth. J. Aquat. Ecol. 1994, 28, 175–180. [Google Scholar] [CrossRef]
- Margaritora, F.G. Crustacea, Branchiopoda, Cladocera. In Checklist e Distribuzione Della Fauna Italiana: 10.000 Specie Terrestri e Delle Acque Interne Memorie del Museo Civico di Storia Naturale di Verona; Serie 2: Sezione scienze della, vita; Ruffo, S., Stoch, F., Eds.; Museo civico di storia naturale di Verona: Verona, Italy, 2005; pp. 87–90. ISBN 8889230037. [Google Scholar]
- Geraldes, A.M.A.; Alonso, M. Bosmina (Eubosmina) coregoni Baird, 1857 (Crustacea, Branchiopoda, Anomopoda): New planktonic invader in the Iberian peninsula. Graellsia 2014, 70, e015. [Google Scholar] [CrossRef]
- Alonso, M. Crustacea, Branchiopoda: Fauna Iberica; Museo Nacional de Ciencias Naturales: Madrid, Spain, 1996; Volume 7, ISBN 9788400075712. [Google Scholar]
- Lazareva, V.I. New and rare zooplankton species expansion and particular features of their naturalization in waterbodies of the Upper Volga Basin. Inland Water Biol. 2008, 1, 76–83. [Google Scholar] [CrossRef]
- Zhdanova, S.M. Diaphanosoma mongolianum Ueno, 1938 (Cladocera: Sididae) in Lakes of Yaroslavl Oblast (Russia). Inland Water Biol. 2018, 11, 145–152. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; Mirabdullaev, I.M. Diaphanosoma dubium Manuilova, 1964 and D. excisum Sars, 1885, new species of hydrofauna from Central Asia and Kazakhstan. Arthropoda Sel. 1994, 3, 7–11. [Google Scholar]
- Korovchinsky, N.M. Cladocera: Ctenopoda. Families Sididae, Holopediidae and Pseudopenilidae (Branchiopoda: Cladocera); Backhuys Publishers: Weikersheim, Germany; Margraf Publishers GmbH: Weikersheim, Germany, 2018; ISBN 978-3-8236-1756-3. [Google Scholar]
- Gromova, Y.F.; Protasov, A.A.; Guleykova, L.V. Cladocera of Genus Diaphanosoma (Crustacea: Cladocera) of the Water Bodies and Water Courses of Ukraine. Hydrobiol. J. 2018, 54, 27–33. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; Kotov, A.A.; Sinev, A.Y.; Neretina, A.N.; Garibian, P.G. Water Fleas (Crustacea: Cladocera) of Northern Eurasia; KMK Scientific press Ltd.: Moscow, Russia, 2021; Volume 2, ISBN 978-5-907372-50-4. [Google Scholar]
- Stolbunova, V.N.; Rivier, I.K.; Pidgaiko, M.L. New species of Cladocera for the Ivankovskoje water reservoir. Inland Water Biol. 1975, 28, 33–36. [Google Scholar]
- Semenova, A.S.; Tchougounov, V.K. The Distribution of Moina micrura Kurz, 1875 (Crustacea: Moinidae) in the Russian Part of the Vistula Lagoon (Baltic Sea). Russ. J. Biol. Invasions 2018, 9, 175–183. [Google Scholar] [CrossRef]
- Zhikharev, V.S.; Neretina, A.N.; Zolotareva, T.V.; Gavrilko, D.E.; Shurganova, G.V. Ilyocryptus spinifer Herrick 1882 (Crustacea, Branchiopoda, Cladocera): первая нахoдка вида для фауны Еврoпы. Zool. Zh. 2020, 99, 138–145. [Google Scholar] [CrossRef]
- Lazareva, V.I.; Bolotov, S.E. Analysis of coexistence of the recent invader Diaphanosoma orghidani Negrea with the aboriginal species D. brachyurum (Lievin) (Crustacea, Cladocera) in the Rybinsk reservoir. Russ. J. Biol. Invasions 2013, 4, 161–173. [Google Scholar] [CrossRef]
- Taylor, D.J.; Connelly, S.J.; Kotov, A. The Intercontinental phylogeography of neustonic daphniids. Sci. Rep. 2020, 10, 1818. [Google Scholar] [CrossRef]
- Sramek-Husek, R. O perloockach Pleuroxus striatoides n. sp. and P. striatus Schoedler. Acta Soc. Zool. Bohem. 1946, 10, 232–240. [Google Scholar]
- Neretina, A.N.; Garibian, P.G.; Sinev, A.Y.; Kotov, A.A. Diversity of the subgenus Disparalona (Mixopleuroxus) Hudec, 2010 (Crustacea: Cladocera) in the New and Old World. J. Nat. Hist. 2018, 52, 155–205. [Google Scholar] [CrossRef]
- Adebayo, A.A.; Zhan, A.; Bailey, S.A.; MacIsaac, H.J. Domestic ships as a potential pathway of nonindigenous species from the Saint Lawrence River to the Great Lakes. Biol. Invasions 2013, 16, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Stanislawczyk, K.; Johansson, M.L.; MacIsaac, H.J. Microscopy versus automated imaging flow cytometry for detecting and identifying rare zooplankton. Hydrobiologia 2017, 807, 53–65. [Google Scholar] [CrossRef]
- Popova, E.V.; Petrusek, A.; Kořínek, V.; Mergeay, J.; Bekker, E.I.; Karabanov, D.P.; Galimov, Y.R.; Neretina, T.V.; Taylor, D.J.; Kotov, A.A. Revision of the Old World Daphnia (Ctenodaphnia) similis group Cladocera: Daphniidae). Zootaxa 2016, 4161, 1–40. [Google Scholar] [CrossRef]
- Krylov, A.V.; Kosolapov, D.B.; Kosolapova, N.G.; Hovsepyan, A.A.; Gerasimov, Y.V. The Plankton Community of Sevan Lake (Armenia) after Invasion of Daphnia (Ctenodaphnia) magna Straus, 1820. Biol. Bull. 2018, 45, 505–511. [Google Scholar] [CrossRef]
- Krylov, А.V.; Hayrapetyan, А.O.; Kosolapov, D.B.; Sakharova, Е.G.; Kosolapova, N.G.; Sabitova, R.Z.; Malin, M.I.; Malina, I.P.; Gerasimov, Y.V.; Hovsepyan, А.А.; et al. Features of Structural Changes in the Plankton Community of an alpine lake with increasing fish density in summer and autumn. Zool. Zh. 2021, 100, 147–158. [Google Scholar] [CrossRef]
- Smirnov, A.K.; Pavlov, D.D.; Kodukhova, Y.V.; Karabanov, D.P. Impact of Zebra Mussel Dreissena polymorpha Pallas 1771 (Bivalvia) Appearance on fish populations in Lake Pleshcheevo, European Russia. Zool. Zh. 2020, 99, 1363–1374. [Google Scholar] [CrossRef]
- Kubersky, E.S. Worldwide distribution and ecology of Alonopsis (Cladocera: Chydoridae) with a description of Alonopsis americana sp. nov. Int. Rev. Hydrobiol. 1977, 62, 649–685. [Google Scholar] [CrossRef]
- Sinev, A.Y.; Atroschenko, M.M. Revision of the genus Alonopsis Sars, 1862 and its position within Aloninae (Cladocera: Anomopoda: Chydoridae). Zootaxa 2011, 2800, 1. [Google Scholar] [CrossRef]
- Frey, D.G. Alona weinecki Studer on the subantarctic islands, not Alona rectangula Sars (Chydoridae, Cladocera)1. Limnol. Oceanogr. 1988, 33, 1386–1411. [Google Scholar] [CrossRef]
- Dumont, H.J.; Martens, K. The freshwater microcrustacea of Easter Island. Hydrobiologia 1996, 325, 83–99. [Google Scholar] [CrossRef]
- Dumont, H.J.; Cocquyt, C.; Fontugne, M.; Arnold, M.; Reyss, J.-L.; Bloemendal, J.; Oldfield, F.; Steenbergen, C.L.; Korthals, H.J.; Zeeb, B.A. The end of moai quarrying and its effect on Lake Rano Raraku, Easter Island. J. Paleolimnol. 1998, 20, 409–422. [Google Scholar] [CrossRef]
- Havel, J.E.; Medley, K.A. Biological Invasions Across Spatial Scales: Intercontinental, Regional, and Local Dispersal of Cladoceran Zooplankton. Biol. Invasions 2006, 8, 459–473. [Google Scholar] [CrossRef]
- Bailey, S.A.; Brown, L.; Campbell, M.L.; Canning-Clode, J.; Carlton, J.T.; Castro, N.; Chainho, P.; Chan, F.T.; Creed, J.C.; Curd, A.; et al. Trends in the detection of aquatic non-indigenous species across global marine, estuarine and freshwater ecosystems: A 50-year perspective. Divers. Distrib. 2020, 26, 1780–1797. [Google Scholar] [CrossRef]
- Van Damme, K.; Kotov, A.A.; Dumont, H.J. A checklist of names in Alona Baird 1843 (Crustacea: Cladocera: Chydoridae) and their current status: An analysis of the taxonomy of a lump genus. Zootaxa 2010, 2330, 1–63. [Google Scholar] [CrossRef]
- Van Damme, K.; Kotov, A.A. The fossil record of the Cladocera (Crustacea: Branchiopoda): Evidence and hypotheses. Earth-Science Rev. 2016, 163, 162–189. [Google Scholar] [CrossRef]
- Van Damme, K.; Dumont, H.J. Further division of Alona Baird, 1843: Separation and position of Coronatella Dybowski & Grochowski and Ovalona gen.n. (Crustacea: Cladocera). Zootaxa 2008, 1960, 1–44. [Google Scholar] [CrossRef]
- Van Damme, K. Endemism and long distance dispersal in the waterfleas of Easter Island. Zootaxa 2016, 4154, 251–272. [Google Scholar] [CrossRef]
- Louette, G.; De Meester, L. High dispersal capacity of cladoceran zooplankton in newly founded communities. Ecology 2005, 86, 353–359. [Google Scholar] [CrossRef]
- Hudec, I.; Illyová, M. Pleuroxus denticulatus (Crustacea: Anomopoda: Chydoridae) a new invader in the Danube Basin. Hydrobiologia 1998, 368, 65–73. [Google Scholar] [CrossRef]
- Vad, C.F.; Horvath, Z.; Kiss, K.T.; Acs, E. Microcrustacean (Cladocera, Copepoda) communities in artificial lakes in the region of the North Hungarian mountains, with special reference to the adventive species. Acta Zool. Academ. Sci. Hung. 2012, 58, 47–61. [Google Scholar]
- Frey, D.G. Changing attitudes toward chydorid anomopods since 1769. Hydrobiologia 1995, 307, 43–55. [Google Scholar] [CrossRef]
- Klimovsky, A.I.; Sinev, A.Y.; Bekker, E.I.; Kotov, A.A. Cladocera (Crustacea, Branchiopoda) of Central Yakutia. 2. Some representatives of the families Bosminidae, Eurycercidae and Chydoridae. Zool. Zh. 2015, 94, 1009–1022. [Google Scholar] [CrossRef]
- Sarmaja-Korjonen, K.; Hakojärvi, M.; Korhola, A. Subfossil remains of an unknown chydorid (Anomopoda: Chydoridae) from Finland. Hydrobiologia 2000, 436, 165–169. [Google Scholar] [CrossRef]
- Van Damme, K.; Nevalainen, L. The most latent cladoceran in the Holarctic revealed-sinking Unapertura Sarmaja-Korjonen, Hakojärvi amp; Korhola, 2000 into the genus Rhynchotalona Norman, 1903 (Branchiopoda: Cladocera: Chydoridae). Zootaxa 2019, 4613, 463–476. [Google Scholar] [CrossRef]
- Nevalainen, L.; Kivilä, E.H.; Luoto, T.P.; Rantala, M.V.; Van Damme, K. A hidden species becoming visible: Biogeography and ecology of Rhynchotalona latens (Cladocera, Anomopoda, Chydoridae). Hydrobiologia 2019, 837, 47–59. [Google Scholar] [CrossRef]
- Van Damme, K.; Brancelj, A.; Dumont, H.J. Adaptations to the hyporheic in Aloninae (Crustacea: Cladocera): Allocation of Alona protzi Hartwig, 1900 and related species to Phreatalona gen. nov. Hydrobiologia 2008, 618, 1–34. [Google Scholar] [CrossRef]
- Sinev, A.Y.; Dumont, H.J. Revision of the costata-group of Alona s. lato (Cladocera: Anomopoda: Chydoridae) confirms its generic status. Eur. J. Taxon. 2016, 2016, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Margaritora, F.G.; Vagaggini, D.; Stoch, F. Two Chydoridae species (Crustacea, Cladocera) new to Italy: Alona rustica and Camptocercus uncinatus. Ital. J. Zool. 2002, 69, 59–63. [Google Scholar] [CrossRef]
- Heine-Fuster, I.; Vega-Retter, C.; Sabat, P.; Ramos-Jiliberto, R. Osmoregulatory and demographic responses to salinity of the exotic cladoceran Daphnia exilis. J. Plankton Res. 2010, 32, 1405–1411. [Google Scholar] [CrossRef]
- Rocha, O.; Espíndola, E.L.G.; Rietzler, A.C.; Fenerich-Verani, N.; Verani, J.R. Animal Invaders in São Paulo State Reservoirs. Oecologia Aust. 2011, 15, 631–642. [Google Scholar] [CrossRef]
- Carter, M.J.; Silva-Flores, P.; Oyanedel, J.P.; Ramos-Jiliberto, R. Morphological and life-history shifts of the exotic cladoceran Daphnia exilis in response to predation risk and food availability. Limnologica 2013, 43, 203–209. [Google Scholar] [CrossRef]
- Haileselasie, T.H.; Mergeay, J.; Vanoverbeke, J.; Orsini, L.; De Meester, L. Founder effects determine the genetic structure of the water flea Daphnia in Ethiopian reservoirs. Limnol. Oceanogr. 2017, 63, 915–926. [Google Scholar] [CrossRef]
- Smirnov, N.N. On inshore Cladocera of the Volga Water Reservoirs. Hydrobiologia 1963, 21, 166–176. [Google Scholar] [CrossRef]
- Pecorari, S.; De Paggi, S.J.; Paggi, J.C. Assessment of the urbanization effect on a lake by zooplankton. Water Resour. 2006, 33, 677–685. [Google Scholar] [CrossRef]
- Liu, P.; Xu, S.; Lin, J.; Li, H.; Lin, Q.; Han, B.-P. Urbanization increases biotic homogenization of zooplankton communities in tropical reservoirs. Ecol. Indic. 2019, 110, 105899. [Google Scholar] [CrossRef]
- Kinzelbach, R. Neozoans in european waters—Exemplifying the worldwide process of invasion and species mixing. Experientia 1995, 51, 526–538. [Google Scholar] [CrossRef]
- Armengol, J. Los crustaceos del plancton de los embalses espanoles. Oecol. Aquat. 1978, 3, 3–96. [Google Scholar]
- Alonso, M. Review of Iberian Cladocera with Remarks on Ecology and Biogeography; Springer: Dordrecht, The Netherlands, 1991; ISBN 978-90-481-4101-2. [Google Scholar]
- Adamowicz, S.J.; Hebert, P.D.N.; Marinone, M.C. Species diversity and endemism in the Daphnia of Argentina: A genetic investigation. Zool. J. Linn. Soc. 2004, 140, 171–205. [Google Scholar] [CrossRef] [Green Version]
- Dexter, E.; Bollens, S.M. Zooplankton invasions in the early 21st century: A global survey of recent studies and recommendations for future research. Hydrobiologia 2019, 847, 309–319. [Google Scholar] [CrossRef]
- López-Blanco, C.; Miracle, M.R.; Vicente, E. Cladoceran assemblages in a karstic lake as indicators of hydrological alterations. Hydrobiologia 2011, 676, 249–261. [Google Scholar] [CrossRef]
- Abramova, E.; Vishnyakova, I.; Boike, J.; Solovyev, G.; Martynov, F. Structure of freshwater zooplankton communities from tundra waterbodies in the Lena River Delta, Russian Arctic, with a discussion on new records of glacial relict copepods. Polar Biol. 2017, 40, 1629–1643. [Google Scholar] [CrossRef]
- Kalinowska, K.; Ejsmont-Karabin, J.; Rzepecki, M.; Kostrzewska-Szlakowska, I.; Feniova, I.Y.; Palash, A.; Dzialowski, A.R. Impacts of large-bodied crustaceans on the microbial loop. Hydrobiologia 2014, 744, 115–125. [Google Scholar] [CrossRef]
- Havel, J.E.; Colbourne, J.K.; Hebert, P. Reconstructing the history of intercontinental dispersal in Daphnia lumholtzi by use of genetic markers. Limnol. Oceanogr. 2000, 45, 1414–1419. [Google Scholar] [CrossRef]
- Briski, E.; Cristescu, M.; Bailey, S.; MacIsaac, H.J. Use of DNA barcoding to detect invertebrate invasive species from diapausing eggs. Biol. Invasions 2010, 13, 1325–1340. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, H.; Wu, H.; Xue, J.; Liu, Y.; Jiang, X. Invasion risk to Yangtze River Estuary posed by resting eggs in ballast sediments from transoceanic ships. J. Exp. Mar. Biol. Ecol. 2021, 545, 151627. [Google Scholar] [CrossRef]
- Briski, E.; Wiley, C.J.; Bailey, S.A. Role of domestic shipping in the introduction or secondary spread of nonindigenous species: Biological invasions within the Laurentian Great Lakes. J. Appl. Ecol. 2012, 49, 1124–1130. [Google Scholar] [CrossRef]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Ostell, J.; Pruitt, K.; Mizrachi, I.K. GenBank. Nucleic Acids Res. 2018, 47, D94–D99. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, A.; Makino, W.; Urabe, J. The taxonomic position of Asian Holopedium (Crustacea: Cladocera) confirmed by morphological and genetic analyses. Limnology 2019, 21, 97–106. [Google Scholar] [CrossRef]
- Ma, X.; Ni, Y.; Wang, X.; Hu, W.; Yin, M. Lineage diversity, morphological and genetic divergence in Daphnia magna (Crustacea) among Chinese lakes at different altitudes. Contrib. Zool. 2020, 89, 450–470. [Google Scholar] [CrossRef]
- Garibian, P.G.; Neretina, A.N.; Taylor, D.J.; Kotov, A.A. Partial revision of the neustonic genus Scapholeberis Schoedler, 1858 (Crustacea: Cladocera): Decoding of the barcoding results. PeerJ 2020, 8, e10410. [Google Scholar] [CrossRef] [PubMed]
- Karabanov, D.P.; Bekker, E.I.; Kotov, A.A. Underestimated consequences of biological invasions in phylogeographic reconstructions as seen in Daphnia magna (Crustacea, Cladocera). Zool. Zh. 2020, 99, 1232–1241. [Google Scholar] [CrossRef]
- Comtet, T.; Sandionigi, A.; Viard, F.; Casiraghi, M. DNA (meta)barcoding of biological invasions: A powerful tool to elucidate invasion processes and help managing aliens. Biol. Invasions 2015, 17, 905–922. [Google Scholar] [CrossRef]
- Hernandez, C.; Bougas, B.; Perreault-Payette, A.; Simard, A.; Côté, G.; Bernatchez, L. 60 specific eDNA qPCR assays to detect invasive, threatened, and exploited freshwater vertebrates and invertebrates in Eastern Canada. Environ. DNA 2020, 2, 373–386. [Google Scholar] [CrossRef]
- Yates, M.C.; Derry, A.M.; Cristescu, M.E. Environmental RNA: A Revolution in Ecological Resolution? Trends Ecol. Evol. 2021, 36, 601–609. [Google Scholar] [CrossRef]
- Walsh, J.R.; Spear, M.J.; Shannon, T.P.; Krysan, P.J.; Zanden, M.J.V. Using eDNA, sediment subfossils, and zooplankton nets to detect invasive spiny water flea (Bythotrephes longimanus). Biol. Invasions 2018, 21, 377–389. [Google Scholar] [CrossRef]
- Mychek-Londer, J.G.; Chaganti, S.R.; Heath, D.D. Metabarcoding of native and invasive species in stomach contents of Great Lakes fishes. PLoS ONE 2020, 15, e0236077. [Google Scholar] [CrossRef]
- Smirnov, N.N. Historical Ecology of Freshwater Zoocenozes; KMK Scientific Press Ltd.: Moscow, Russia, 2010; ISBN 978-5-87317-646-5. [Google Scholar]
- Smirnov, N.N. First record of Pleuroxus pigroides (Lilljeborg, 1901) in Russia. Arthropoda Sel. 2004, 13, 109–112. [Google Scholar]
- Kotov, A.A. The first record of Daphnia magna Straus, 1820 (Cladocera: Anomopoda) in Lake Glubokoe. Tr. Gidrobiol. Stantsii Na Glubokom Ozere 2005, 9, 191–194. [Google Scholar]
- Suchy, K.D.; Hann, B.J. Using Microfossil Remains in Lake Sediments to Examine the Invasion of Eubosmina coregoni (Cladocera, Bosminidae) in Lake of the Woods, Ontario, Canada. J. Great Lakes Res. 2007, 33, 867–874. [Google Scholar] [CrossRef]
- Suchy, K.D.; Salki, A.; Hann, B.J. Investigating the invasion of the nonindigenous zooplankter, Eubosmina coregoni, in Lake Winnipeg, Manitoba, Canada. J. Great Lakes Res. 2010, 36, 159–166. [Google Scholar] [CrossRef]
- DeWeese, N.E.; Favot, E.J.; Branstrator, D.K.; Reavie, E.D.; Smol, J.P.; Engstrom, D.R.; Rantala, H.M.; Schottler, S.P.; Paterson, A.M. Early presence of Bythotrephes cederströmii (Cladocera: Cercopagidae) in lake sediments in North America: Evidence or artifact? J. Paleolimnol. 2021, 66, 389–405. [Google Scholar] [CrossRef] [PubMed]
- Frey, D.G. The Taxonomic and Phylogenetic Significance Of the Head Pores of the Chydoridae (Cladoceha). Int. Rev. Hydrobiol. 1959, 44, 27–50. [Google Scholar] [CrossRef]
- Szeroczynska, K.; Sarmaja-Korjonen, K. Atlas of Subfossil Cladocera from Central and Northern Europe; Friends of the Lower Vistula Society: Swiecie, Poland, 2007; ISBN 9788392491965. [Google Scholar]
- Ohtsuki, H.; Awano, T.; Tsugeki, N.K.; Ishida, S.; Oda, H.; Makino, W.; Urabe, J. Historical Changes in the Ecosystem Condition of a Small Mountain Lake over the Past 60 Years as Revealed by Plankton Remains and Daphnia Ephippial Carapaces Stored in Lake Sediments. PLoS ONE 2015, 10, e0119767. [Google Scholar] [CrossRef]
- Conde-Porcuna, J.M.; Veiga, J.; Moreno, E.; Jiménez, L.; Ramos-Rodríguez, E.; Pérez-Martínez, C. Spatiotemporal genetic structure in the Daphnia pulex complex from Sierra Nevada lakes (Spain): Reproductive mode and first record of North American D. cf. pulex in European alpine lakes. J. Plankton Res. 2021, 43, 380–395. [Google Scholar] [CrossRef]
- Ellegaard, M.; Clokie, M.R.J.; Czypionka, T.; Frisch, D.; Godhe, A.; Kremp, A.; Letarov, A.; McGenity, T.J.; Ribeiro, S.; Anderson, N.J. Dead or alive: Sediment DNA archives as tools for tracking aquatic evolution and adaptation. Commun. Biol. 2020, 3, 169. [Google Scholar] [CrossRef]
- Capo, E.; Giguet-Covex, C.; Rouillard, A.; Nota, K.; Heintzman, P.; Vuillemin, A.; Ariztegui, D.; Arnaud, F.; Belle, S.; Bertilsson, S.; et al. Lake Sedimentary DNA Research on Past Terrestrial and Aquatic Biodiversity: Overview and Recommendations. Quaternary 2021, 4, 6. [Google Scholar] [CrossRef]
- Lazareva, V.I. (Ed.) Structure and Functioning of the Ecosystem in the Rybinsk Reservoir at the Beginning of the 21st Century; Russian Academy of Sciences: Moscow, Russia, 2018; ISBN 978-5-907036-18-5. [Google Scholar]
- Manujlova, E.F. The Cladocerans of Fauna of the USSR; Nauka: Moscow, Russia, 1964. [Google Scholar]
- Glagolev, S.M. Genus Daphnia. In Key to Freshwater Invertebrates of Russia and Adjacent Lands: 2. Crustacea; Alekseev, V.R., Ed.; Zoological Institute of Russian Academy of Sciences: St. Petersburg, Russia, 1995; pp. 48–58. [Google Scholar]
- Korovchinsky, N.M. Cladocerans of the Order Ctenopoda of the World Fauna (Morphology, Systematics, Ecology, Biogeography); KMK Press: Moscow, Russia, 2004. [Google Scholar]
- Elias-Gutierrez, M.; Suarez Morales, E.; Gutierrez Aguirre, M.; Silva Briano, M.; Granados Ramirez, J.G.; Garfias Espejo, T. Cladocera y Copepoda de Las Aguas Continentales de Mexico: Guia Ilustrada; Universidad Nacional Autonoma de Mexico: Mexico City, Mexico, 2008. [Google Scholar]
- Sousa, F.D.R.; Elmoor-Loureiro, L.M.A. Identification key of Ctenopoda (Cladocera, Holopedidae, Sididae) taxa from Brazil with remarks on taxonomy and geographic distribution. Zootaxa 2021, 5047, 53–67. [Google Scholar] [CrossRef]
- Macêdo, R.L.; Sousa, F.D.R.; Dumont, H.J.; Rietzler, A.C.; Rocha, O.; Elmoor-Loureiro, L.M.A. Climate change and niche unfilling tend to favor range expansion of Moina macrocopa Straus 1820, a potentially invasive cladoceran in temporary waters. Hydrobiologia 2022. [Google Scholar] [CrossRef]
- Frey, D.G. The non-cosmopolitanism of Chydorid Cladocera: Implications for biogeography and evolution. In Crustacean Biogeography; Gore, R.H., Robert, H., Kenneth, L., Eds.; Routledge: Rotterdam, The Netherlands, 1986; pp. 237–256. ISBN 9781315140674. [Google Scholar]
- Smirnov, N.N. Chydoridae Fauni Mira. Fauna SSSR. Rakoobraznie. [Chydoridae of the World’s Fauna. Fauna of the USSR. Crustacea]; Nauka: Leningrad, Russia, 1971. [Google Scholar]
- Frey, D.G. On the plurality of Chydorus sphaericus (O. F. Müller) (Cladocera, Chydoridae), and designation of a neotype from Sjaelsø, Denmark. Hydrobiologia 1980, 69, 83–123. [Google Scholar] [CrossRef]
- Kotov, A.A. Faunistic complexes of the Cladocera (Crustacea, Branchiopoda) of Eastern Siberia and the Far East of Russia. Zool. Zh. 2016, 95, 748–768. [Google Scholar] [CrossRef]
- Wang, J.; Ni, Y.; Hu, W.; Yin, M. Lineage diversity and gene introgression in freshwater cladoceran crustaceans of the Chydorus sphaericus species complex. Limnol. Oceanogr. 2020, 66, 95–107. [Google Scholar] [CrossRef]
- Lin, W.; Zhou, G.; Cheng, X.; Xu, R. Fast Economic Development Accelerates Biological Invasions in China. PLoS ONE 2007, 2, e1208. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, P.R.; Pringle, R.M. Where does biodiversity go from here? A grim business-as-usual forecast and a hopeful portfolio of partial solutions. Proc. Natl. Acad. Sci. USA 2008, 105, 11579–11586. [Google Scholar] [CrossRef]
- Barber, P.H.; Ablan-Lagman, M.C.A.; Ambariyanto; Berlinck, R.G.S.; Cahyani, D.; Crandall, E.D.; Ravago-Gotanco, R.; Juinio-Meñez, M.A.; Mahardika, I.G.N.; Shanker, K.; et al. Advancing biodiversity research in developing countries: The need for changing paradigms. Bull. Mar. Sci. 2014, 90, 187–210. [Google Scholar] [CrossRef]
- Fryer, G.; Frey, D.G. Two-egged ephippia in the chydorid Cladocera. Freshw. Biol. 1981, 11, 391–394. [Google Scholar] [CrossRef]
- Fryer, G. Crustacean diversity in relation to the size of water bodies: Some facts and problems. Freshw. Biol. 1985, 15, 347–361. [Google Scholar] [CrossRef]
- Makrushin, A.V. Resistence to drying of latent eggs of Bosmina obtusirostris (Cladocera: Crustacea) and their reactivation. Zool. Zh. 1989, 68, 132–134. [Google Scholar]
- Rivier, I.K. The predatory Cladocera, (Onychopoda: Podonidae, Polyphemidae, Cercopagidae), and Leptodorida of the World; Backhuys: Leiden, The Netherlands, 1998; ISBN 9789073348851. [Google Scholar]
- Branstrator, D.K. Ecological Interactions Between Bythotrephes cederstroemi and Leptodora kindtii and the Implications for Species Replacement in Lake Michigan. J. Great Lakes Res. 1995, 21, 670–679. [Google Scholar] [CrossRef]
- Enz, C.A.; Heller, C.; Muller, R.; Bürgi, H.-R. Investigations on fecundity of Bythotrephes longimanus in Lake Lucerne (Switzerland) and on Niche Segregation of Leptodora kindti and Bythotrephes longimanus in Swiss lakes. Hydrobiologia 2001, 464, 143–151. [Google Scholar] [CrossRef]
- Kiehnau, E.L.; Weider, L.J. The great escape: Daphnia pulicaria from post-Bythotrephes invasion time periods have an enhanced escape ability. Hydrobiologia 2022, 849, 1083–1094. [Google Scholar] [CrossRef]
- Lazareva, V.I. The Distribution of species of the genus Diaphanosoma (Crustacea, Cladocera) in reservoirs of the Volga and Sheksna Rivers: Impact of environmental factors. Inland Water Biol. 2012, 5, 257–265. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Redescription of Limnosida frontosa Sars, 1862 (Crustacea: Cladocera: Sididae), with evidence of possible incipient species differentiation. Zootaxa 2014, 3764, 61–80. [Google Scholar] [CrossRef]
- Karabanov, D.P.; Garibian, P.G.; Bekker, E.I.; Sabitova, R.Z.; Kotov, A.A. Genetic Signature of a Past Anthropogenic Transportation of a Far-Eastern Endemic Cladoceran (Crustacea: Daphniidae) to the Volga Basin. Water 2021, 13, 2589. [Google Scholar] [CrossRef]
- Adamowicz, S.J.; Petrusek, A.; Colbourne, J.K.; Hebert, P.D.; Witt, J.D. The scale of divergence: A phylogenetic appraisal of intercontinental allopatric speciation in a passively dispersed freshwater zooplankton genus. Mol. Phylogenetics Evol. 2009, 50, 423–436. [Google Scholar] [CrossRef] [PubMed]
- López, C.; Mosquera, P.V.; Hampel, H.; Neretina, A.N.; Alonso, M.; Van Damme, K.; Kotov, A.A. An annotated checklist of the freshwater cladocerans (Crustacea: Branchiopoda: Cladocera) of Ecuador and the Galapagos Islands. Invert. Zool. 2018, 15, 277–291. [Google Scholar] [CrossRef]
- Yoon, S.M. Arthropoda: Branchiopoda: Anostraca, Notostraca, Spinicaudata, Laevicaudata, Ctenopoda, Anomopoda, Haplopoda Branchiopods. Invertebr. Fauna Korea 2010, 21, 1–156. [Google Scholar]
- Xiang, X.-F.; Ji, G.-H.; Chen, S.-Z.; Yu, G.-L.; Xu, L.; Han, B.-P.; Kotov, A.A.; Dumont, H.J. Annotated Checklist of Chinese Cladocera (Crustacea: Branchiopoda). Part I. Haplopoda, Ctenopoda, Onychopoda and Anomopoda (families Daphniidae, Moinidae, Bosminidae, Ilyocryptidae). Zootaxa 2015, 3904, 1–27. [Google Scholar] [CrossRef]
- Makino, W.; Machida, R.J.; Okitsu, J.; Usio, N. Underestimated species diversity and hidden habitat preference in Moina (Crustacea, Cladocera) revealed by integrative taxonomy. Hydrobiologia 2019, 847, 857–878. [Google Scholar] [CrossRef]
- Mirabdullaev, I.M. Moina weismanni (Crustacea, Cladocera): A species new for Russia and central Asia. Zool. Zh. 1992, 71, 136–139. [Google Scholar]
- Leoni, B.; Cotta-Ramusino, M.; Margaritora, F.G. Seasonal succession of Cladocerans in a ricefield in Italy. Hydrobiologia 1998, 391, 239–245. [Google Scholar] [CrossRef]
- Sinev, A.Y. Discrimination between two sibling species of Acroperus (Baird, 1843) from the Palearctic (Cladocera: Anomopoda: Chydoridae). Zootaxa 2009, 2176, 1–21. [Google Scholar] [CrossRef]
- Sinev, A.Y. A new and a rare species of Chydorus Leach, 1816 (Branchiopoda: Cladocera: Anomopoda) from Cat Tien National Park, Vietnam. Zootaxa 2014, 3861, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Margaritora, F.G. Cladocera. Fauna d’ltalia; Edizioni Calderini: Bologna, Italy, 1985; Volume XXIII, ISBN 8870192512. [Google Scholar]
- Neretina, A.N.; Sinev, A.Y. A revision of the genus Leberis Smirnov, 1989 (Cladocera: Chydoridae) in the Old World and Australia. Zootaxa 2016, 4079, 501–533. [Google Scholar] [CrossRef]
- Zhikharev, V.S.; Sinev, A.Y.; Shurganova, G.V. New data on the distribution of the alian species Pleuroxus denticulatus Birge, 1879 (Cladocera: Chydoridae) in European Russia. In Invasion of Alien Species in Holarctic. Borok-VI; The Sixth International Symposium “Invasion of Alien Species in Holarctic. Borok-VI”, Borok—Uglich, Russia; Dgebuadze, Y.Y., Krylov, A.V., Petrosyan, V.G., Karabanov, D.P., Eds.; Buk Press: Kazan, Russia, 2021; p. 245. ISBN 978-5-00118-788-2. [Google Scholar]
- Dumont, H.J.; Vierstraete, A.; Akbulut, N. Cornigerius lacustris of Lake Hazar, Turkey, a synonym of Cornigerius maeoticus (Pengo) of the Ponto-Caspian (Cladocera: Onychopoda). Zootaxa 2019, 4619, 177–183. [Google Scholar] [CrossRef]
- Zinevici, V.; Moldoveanu, M.; Florescu, L. Podonevadne trigona ovum (Zernov, 1901) (Cladocera), an immigrant subspecies with Caspian origin, newly penetrated in Romanian waters. Rom. J. Biol. Zool. 2011, 56, 3–15. [Google Scholar]
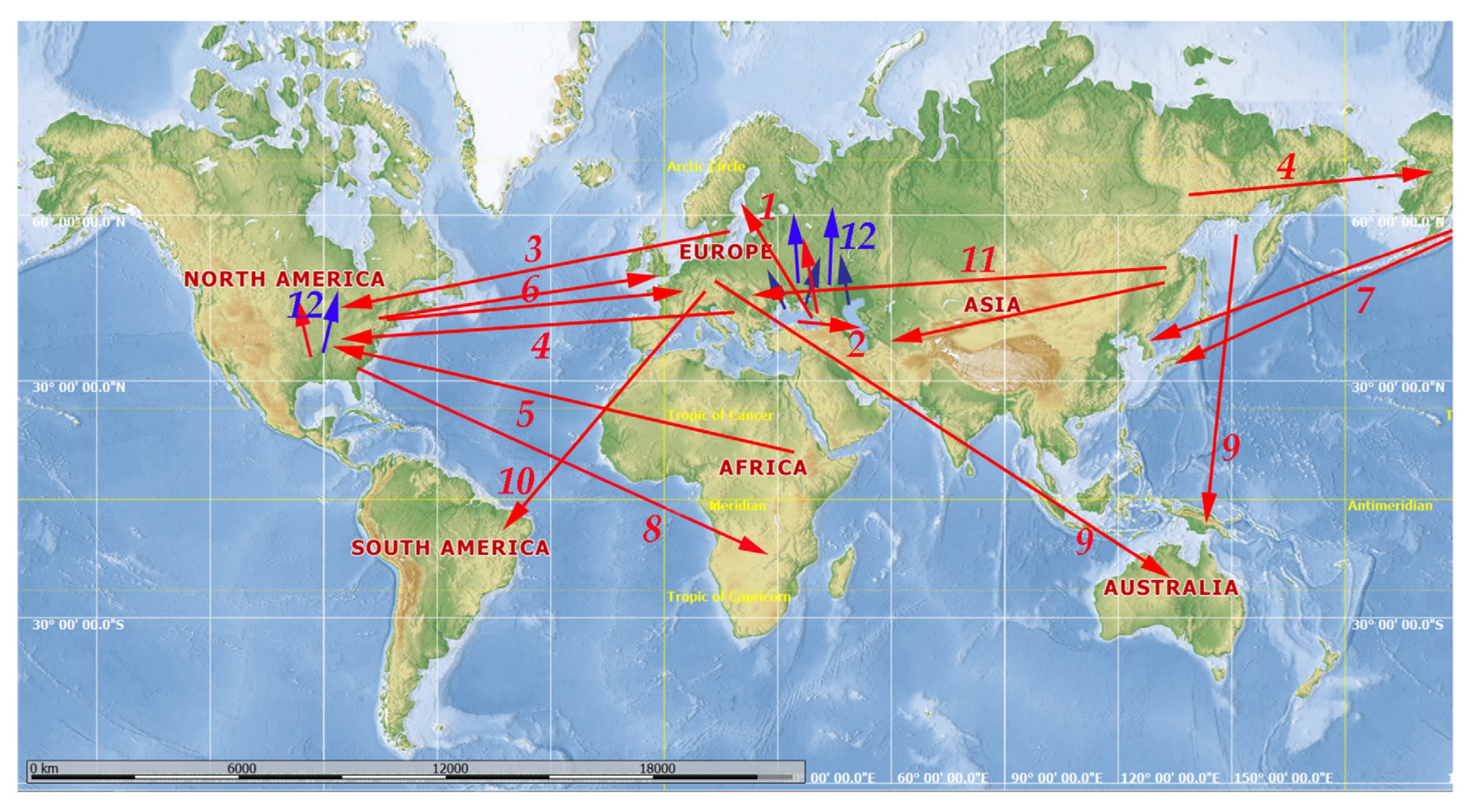

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotov, A.A.; Karabanov, D.P.; Van Damme, K. Non-Indigenous Cladocera (Crustacea: Branchiopoda): From a Few Notorious Cases to a Potential Global Faunal Mixing in Aquatic Ecosystems. Water 2022, 14, 2806. https://doi.org/10.3390/w14182806
Kotov AA, Karabanov DP, Van Damme K. Non-Indigenous Cladocera (Crustacea: Branchiopoda): From a Few Notorious Cases to a Potential Global Faunal Mixing in Aquatic Ecosystems. Water. 2022; 14(18):2806. https://doi.org/10.3390/w14182806
Chicago/Turabian StyleKotov, Alexey A., Dmitry P. Karabanov, and Kay Van Damme. 2022. "Non-Indigenous Cladocera (Crustacea: Branchiopoda): From a Few Notorious Cases to a Potential Global Faunal Mixing in Aquatic Ecosystems" Water 14, no. 18: 2806. https://doi.org/10.3390/w14182806
APA StyleKotov, A. A., Karabanov, D. P., & Van Damme, K. (2022). Non-Indigenous Cladocera (Crustacea: Branchiopoda): From a Few Notorious Cases to a Potential Global Faunal Mixing in Aquatic Ecosystems. Water, 14(18), 2806. https://doi.org/10.3390/w14182806