
Single and Combined Toxicity Effects of Zinc Oxide Nanoparticles: Uptake and Accumulation in Marine Microalgae, Toxicity Mechanisms, and Their Fate in the Marine Environment


Department of Biology, College of Science, University of Bahrain, Zallaq P.O. Box 32038, Bahrain
Water 2022, 14(17), 2669; https://doi.org/10.3390/w14172669
Submission received: 14 June 2022
/
Revised: 16 August 2022
/
Accepted: 25 August 2022
/
Published: 29 August 2022
(This article belongs to the Special Issue Nanoparticles Toxicity to Marine Organisms—a Nanosized or a Giant Environmental Issue?)
Abstract
:Recently, there has been rapid growth in the production of zinc oxide nanoparticles (ZnO-NPs) due to their applications in household and cosmetic products. Over the last decade, considerable research was conducted to reveal the effect of ZnO-NPs on microalgae, which form the base of the aquatic food chain. This review discusses the fate and behavior of ZnO-NPs in the marine environment. Predominately, the toxicity mechanism of ZnO-NPs on marine microalgae could be attributed to three sources: the release of Zn2+ ions; the interaction between ZnO-NPs and algae cells; and the generation of reactive oxygen species (ROS). Most toxicity studies were carried out using single ZnO-NPs under conditions not often observed in natural ecosystems; however, organisms including microalgae are more likely exposed to ZnO-NPs mixed with other types of pollutants. This review highlights the importance of increasing the assessment studies of combined pollutants. Lastly, knowledge, research, gaps, and opportunities for further research in this field are presented.
1. Introduction
Nanoparticles (NPs) are objects comprising pure metals (for example, silver, gold, zinc, titanium, iron) or their compounds (oxides, sulfides, hydroxides, etc.) [1]. NPs are characterized by dimensions less than 100 nm [1]. The unique physiochemical properties of NPs are due to their high surface area and nanoscale size. They can go through several physiochemical changes in aquatic environments, for instance accumulation/dispersion, penetration/transformation, and aggregation/dissolution [2]. These changes are greatly affected by the fundamental characteristics of NPs such as size, shape, and structure [3]. Additionally, their activities depend on morphology, porosity, grain size, composition and crystallinity, group attached on the surface, charge of the surface [4], and other environmental exposure factors such as dissolved organic matter, pH, or ionic strength [5,6,7].
Due to their excellent properties, they are utilized in many applications such as medical fields [8], wastewater treatments [9], and industrial products [10] like optics, paints, coating, catalysis, medical products, cosmetics, and electronics; they can also be used in the degradation of organic pollutants. Nanotechnology has advanced significantly as a result of the widespread use of nanoparticles (NPs) in several fields. Metal-based nano-products are created in large quantities every year all over the world [11].
NPs can be obtained by natural or synthetic methods [3]. Natural NPs appear in nature via different processes including volcanic eruptions, photochemical reactions, dust storms from terrestrial environments, etc. [12]. In contrast, the anthropogenic sources of NPs consist of burning and food cooking, engine exhausts [13], the combustion of sewage sludge [14], the burning of fossil fuels, cosmetic materials, and many other consumer products [15]. Nano-metal, nano-oxide, carbon nanomaterials, quantum dots, and other NPs such as organic polymers are grouped into five categories based on their chemical composition [16]. The widespread combustion of a vast amount of fossil fuels throughout the Industrial Revolution resulted in a significant increase in the number of NPs, which has now become a possible environmental issue [17]. NPs will infiltrate the environment during manufacturing, transit, consumption, and disposal, eventually settling in bodies of water and soils [18]. Many large-scale chemical factories that produce NPs actually release their waste into oceans and rivers. Each year, a considerable number of NPs are dumped into water bodies [12]. In a water environment, NPs pose a risk to the aquatic ecosystem and human health through a series of complex processes such as adsorption, desorption, suspension, sedimentation, and interacting with minerals, organic matter, biofilm, and other complex aspects of the water environment [19].
Zinc oxide nanoparticles (ZnO-NPs) have a constantly expanding market powered primarily by growing demand for cosmetics and sunscreens [20,21,22]. UVA (320–400 nm) and UVB (290–320 nm) radiation protection are provided by ZnO-NPs, which are transparent ultraviolet (UV) absorbers [23]. The physicochemical properties of ZnO-NPs contribute to their effectiveness in several products. They can be employed in industrial applications such as paints, coatings, textiles, and electronics because of their antibacterial, anticorrosion, and catalytic qualities [23]. Wurtzite and zincite, two crystalline forms of ZnO-NPs, are generated [24,25]. Because the hexagonal wurtzite phase of ZnO-NPs is more stable, it is commonly used in designed NPs [24]. The annual output of ZnO-NPs to the environment is in the range of 550 to 5550 tons, and their production is 10 to 100 times higher than the manufacture of any further nanomaterial [11,26]. In Europe, in natural water and wastewater, ZnO-NPs concentrations were determined to be 10 ng/L and 430 ng/L, respectively [27].
Studies show that the production and consumption of NPs are increasing annually [23]. Currently, the aquatic environment, in particular marine ecosystems, faces huge challenges, and the expanding use of NPs is among them. A high percentage of NPs enters the marine environment via different means either intentionally or nonintentionally, which causes serious NP pollution. Most land wastes are discharged into essentially the marine environment. There is increasing concern regarding the implications of NPs’ presence in the marine environment due to their impacts on marine organisms at different trophic levels. The monitoring of the NPs’ characteristics and concentration should be continuous. Over the past twenty years, the pollution of the aquatic environment with NPs, as well as the effects of NPs on aquatic species such as phytoplankton, algae, zooplankton, fish, and other organisms, has been the subject of international research [15]. The existence of several NPs in the marine environment can lead them to interact with each other and modify their overall toxicity on marine organisms.
There are many applications of algae such as the treatment of wastewater, bioremediation, and the sequestration of carbon. They can uptake and accumulate different contaminants like heavy metals and some toxic organic compounds [28,29]. Algae are considered good sources of high-value metabolites that have medicinal and biotechnology applications and can be utilized for biogas production [29]. In addition, because algae are the base of aquatic food webs, any pollutants that damage them will be passed on to the rest of the food web. It is crucial to accurately distinguish the processes of NPs’ internalization in algal cells and consequent harmfulness to identify the risk of NPs in aquatic ecosystems.
Although the ultimate path of most contaminants will be in seawater, studies concerning the effect of NPs were conducted mostly on freshwater algae. For risk assessments of NPs, it is important to understand the mechanism of NPs’ uptake and internalization by cells and subsequent toxic effects. Furthermore, there are some contradictory results in the effect of ZnO-NPs on marine microalgae when tested under different environmental conditions. Therefore, the aim of this review is to summarize the impacts of ZnO-NPs on marine algae, address research gaps, and recommend future research in this field.
The following points and questions will be addressed using published literature from 2010 to 2022: (1) What is the state of the uptake and accumulation of ZnO-NPs in marine microalgae? (2) What are the toxic impacts of ZnO-NPs when used individually or when combined with other materials? (3) What are the toxicity mechanisms of ZnO-NPs?
This review is a pioneering article that summarizes and evaluates the effect of ZnO-NPs when used individually or when combined with other materials on marine microalgae.
2. Fate and Behavior of ZnO-NPs in the Marine Environment
Studies for the quantification of ZnO-NPs in the marine environment had been done, but these studies are few. The concentrations in the environment were mainly calculated using mathematical modeling approaches [30,31]. The employment of specialized and expensive analytical techniques is required for nanoparticle characterization in complex environments such as water as well as in the effluents from wastewater treatment plants, bio-solids, and sediment particles. As a result, the concentrations of these materials must be estimated based on nano-product market production and the known consumption of these items [32] using a life-cycle model to estimate ZnO-NPs released into the environment [27,31,33]. Table 1 highlights the modeled ZnO-NP concentrations in various environmental compartments. These expected ZnO-NP concentrations in the environment can be utilized to build analytical processes and assess the risk of ZnO-NPs [30].
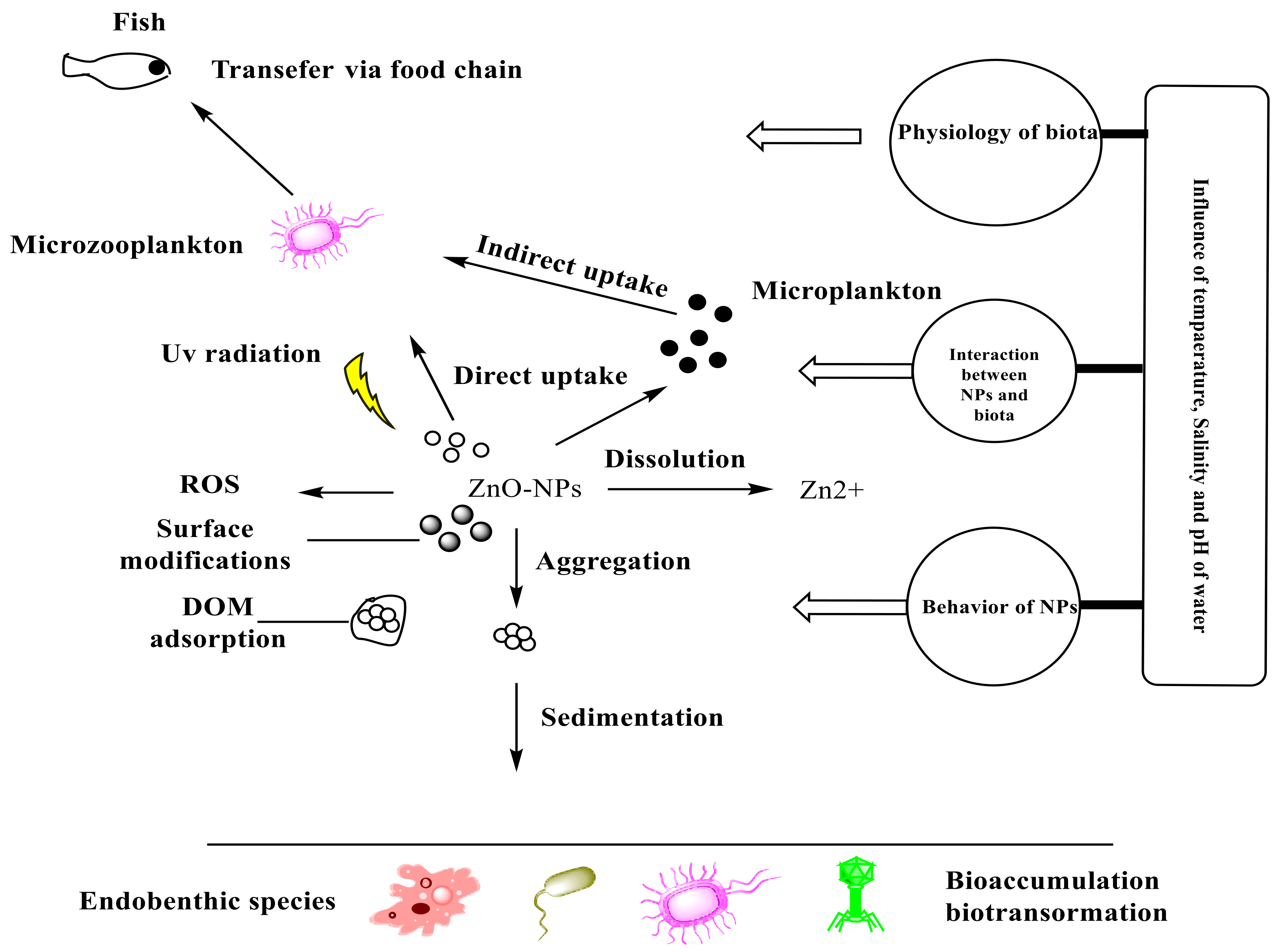
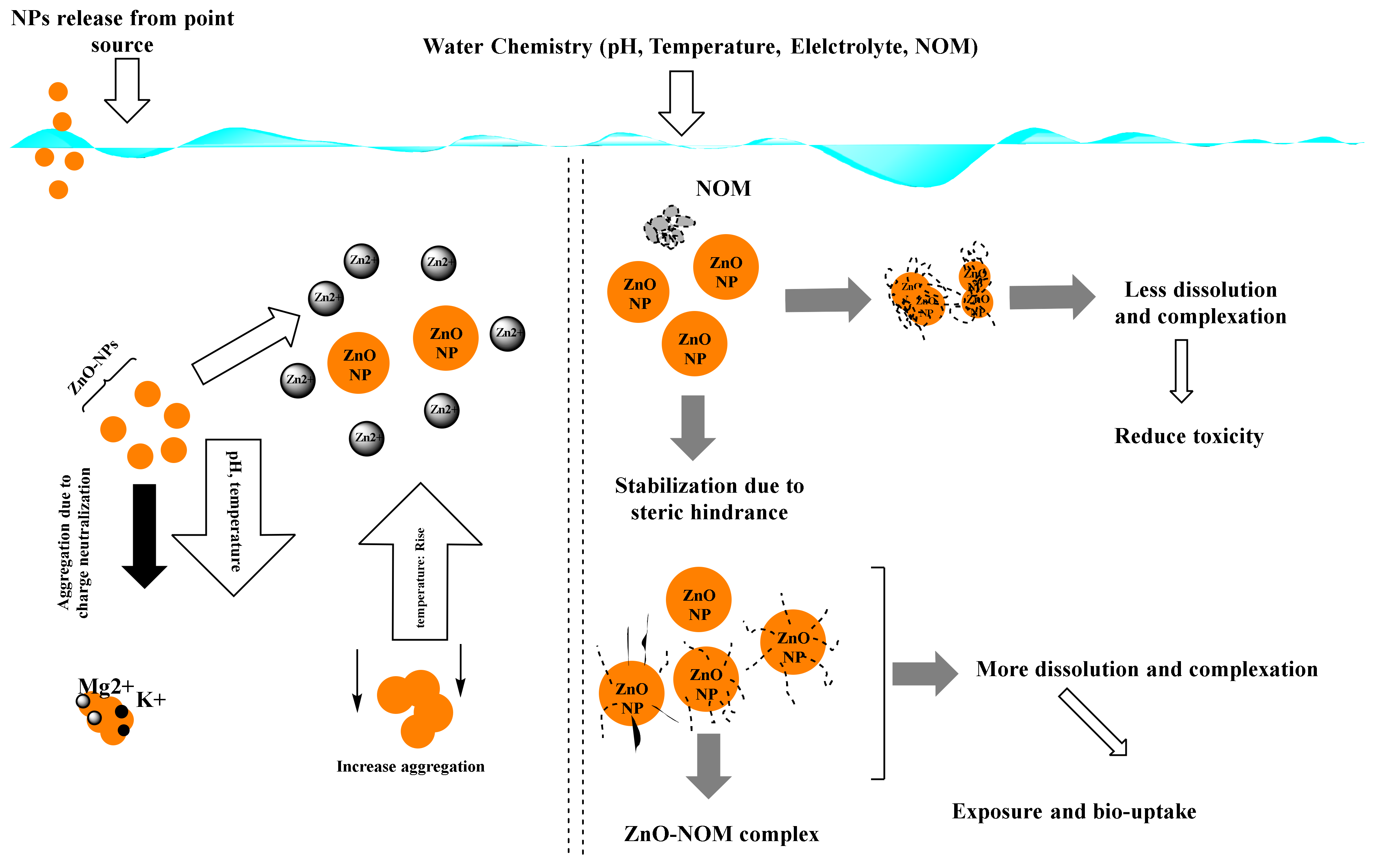
The behavior and movement of NPs in the environment are determined by two fundamental aspects. These factors are: (i) the NPs’ physicochemical properties, “such as particle size and shape, aggregation/agglomeration, surface chemistry (e.g., functionalization), chemical composition, and dissolution”, and (ii) The surrounding environment’s conditions, “for example, oxygen level, natural organic matter, ionic strength and temperature” [32]. Both exposure pathways and exposure concentrations of NPs can be predicted and estimated after understanding the persistence and distribution of these nanoparticles in the natural environment. ZnO-NPs are likely to take a variety of routes if released into the marine environment. NPs in suspensions as separate particles can be dissolved in seawater but also can be aggregated to form a large particle that can be deposited on sediment, in marine waters, it can adsorb onto the components, for instance, the dissolved organic matter (DOM), and can be chemically changed through reduction–oxidation (redox) reactions or physiologically in the existence of other microorganisms in the marine environment [34]. Figure 1 depicts a diagrammatic illustration of ZnO-NPs’ fate and behavior in the marine environment.
2.1. Existence and Persistence
There are two forms of elemental zinc that can accumulate in water, sediment, and organisms: Zn2+ ions and ZnO-NPs. These forms are completely different in their persistence in water. In natural seawater, although Zn2+ ions can transform into other species, they are innately persistent, such as chloro-zinc complexes, hydroxo-zinc complexes, or Zn(OH)2. ZnO-NPs have a lower persistence ability than Zn2+ ions. They can aggregate or dissolve, which will lead to losing the features of the individual nanoparticle. In seawater, ZnO-NPs were soluble, and the amount of Zn2+ released from them varied with time and concentration [35]. The dissolution of ZnO-NPs reached 100% with an original concentration of 0.1 mg/L ZnO-NPs. When the initial concentration of ZnO-NPs was raised to 10 mg/L, dissolution dropped to 60% after 6 h. The release of dissolved Zn2+ ions in water increased after reducing the pH of the solution and in the existence of the organic material humic acid at pH 9–11 [12]. In addition, at 103 mg/L, smaller ZnO-NPs (4 nm) dissolved more rapidly than larger ZnO-NPs (15–240 nm) [36]. Coated ZnO-NPs, for example, silica and polymer in aqueous media, disperse better than uncoated ZnO-NPs. In comparison with uncoated NPs, coated NPs have a longer existence in the marine environment [37].
2.2. Aggregation
Once the nanoparticles find their way to the water, they can form aggregates. The collective effect of van der Waals forces and the double layer force (DLVO) theory produces these homo-aggregates. The interaction powers between the particles of charged surfaces in a liquid media are explained by this theory. The joint effects of van der Waals attraction and electrostatic repulsion due to the electric double layer of counterions that surround ZnO-NPs in an aqueous medium determine the constancy of the ZnO-NPs suspension. If the electrical charges on ZnO-NPs are large enough to overcome the van der Waals attraction force, they will remain separate and stable in suspension. The strong ionic strength of the marine environment causes the electric-double layer to compress. As a result, the electrostatic repulsion forces between the nanoparticles are minimized, and they are more likely to combine and settle out of suspension [36]. There are different factors that can affect the degree of aggregation of the nanoparticles: particle morphology; NP size, surface area, and surface charges; types of environmental structures; and exposure pathways. The transport of nanoparticles mainly depends on their size. Aggregates of nanoparticles usually have a greater probability of accumulation on sediments, and as a result, they are less mobile than individual nanoparticles. Other methods, such as the dissociation of nanoparticles or resuspension as a consequence of changes in the aquatic chemistry of the surrounding environment, as well as the activity of creatures in this habitat, might prevent aggregated nanoparticle deposition. The bioavailable ZnO-NP concentration for pelagic species is reduced when these aggregates are removed by sedimentation or deposition. The aggregated ZnO-NPs become less mobile, and the living organisms occupying the sediments can absorb them.
2.3. Adsorption
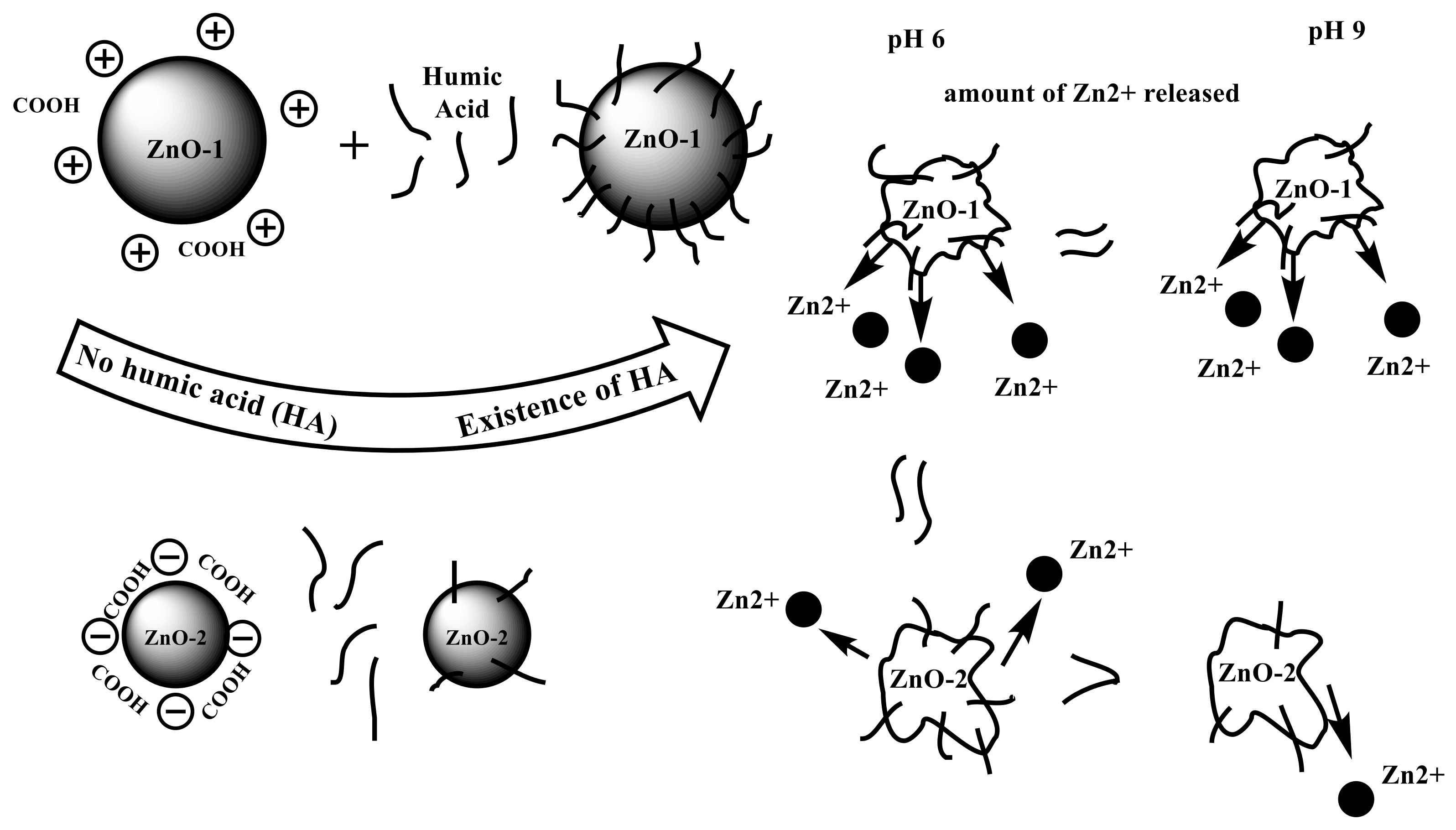
Nanoparticles are surface reactive due to their high surface-to-volume ratio. As a result, NPs in the marine environment can adsorb onto pollutants or dissolved organic matter (DOM). DOM generated from algae or higher plants, such as humic acids and fulvic acids, is abundant in the environment. These plant materials have different functional groups such as carboxylic acid and phenolic hydroxide. At neutral pH, these functional groups are usually negatively charged and may contain light-absorbing chromophores [38]. The light absorbed by these chromophores leads to the formation of reactive oxidative species (ROS). These species may increase the negative effects of ZnO-NPs or alter them chemically. When DOM is adsorbed onto the surface of ZnO-NPs, the surface charge of NPs is altered, and their aggregation/agglomeration is inhibited. As a result, their mobility in their surroundings improves. Humic acids (HAs), which are one of the most reactive types of natural organic matter (NOM), have a great effect on the biogeochemical behavior of NPs in different environments including aquatic ecosystems [39]. The ability of HAs to adsorb onto the surface of different NPs including ZnO-NPs can control the behavior of NPs in media such as aggregating [40], control their activity [41], and control their stability via hydrophobic interactions and the formation of hydrogen bonds [42]. Additionally, HAs might have an essential role in the bioavailability of NPs and the permeability of cell membranes [43]. NPs have the ability to adsorb molecules when in contact with biotic and abiotic environments due to their high surface energy. This feature leads to the formation of a biomolecule corona [44] which could be formed in the presence of HAs [45]. The adsorption of HAs onto ZnO-NPs is a good example of how to investigate the fate and behavior of ZnO-NPs in the environment. It was found that the adsorption of HAs resulted in a decrease in particle zeta potential. They can reduce the negative effects of ZnO-NPs on aquatic organisms [46]. Consequently, humic acid-coated ZnO-NPs were easier to disseminate and suspend than uncoated ZnO-NPs [47].
In the presence of HAs at pH 9.0, the dissolution level of nZnO−1 was higher than that of nZnO−2 (Figure 2). Although nZnO−2 is characterized by its small particle size and large surface area compared with nZnO−1, nZnO−1 adsorbed more HAs due to its lower hydrophilicity and the stronger electrostatic attraction between HAs and nZnO−1. This is caused by its lower water content and positive zeta potential [48]. The sorption of polycyclic aromatic hydrocarbons (PAHs) onto HA-coated ZnO-NPs is enhanced in a different study [49].
2.4. Transformation
ZnO-NPs can be transformed in the environment. This has a great influence on the size, shape, and surface chemistry of these nanoparticles. The process of transformation will have an effect on their persistence, distribution, and toxicity in the environment. ROS can be produced due to the presence of some redox reactions of organic species present in the environment on the outer surfaces of ZnO-NPs. These ROS species can stimulate oxidative stresses in vivo and in vitro on cultured cells of bacteria [50].
The balance of four processes determines the natural fate of ZnO-NPs: absorption, distribution, metabolism, and excretion. The process of ZnO-NP “biotransformation” by different living organisms in the biological environment and how this conversion impacts ZnO-NPs toxic effects are still unknown. In conclusion, the environmental fate of ZnO-NPs is controlled by their physical and chemical features as well as by ambient factors. The behavior and transport of ZnO-NPs, as well as their interactions with species in aquatic environments, will be dramatically influenced by modification.
3. Uptake and Accumulation of NPs in Microalgae
Different types of NPs such as carbon-based NPs (CNPs), metal oxide-based NPs (MONPs), and noble metal-based NPs (NMNPs) show different interaction pathways with microalgae. Several factors were found to control and affect the uptake and accumulation of NPs, such as the composition of cell wall and thickness, composition of the cell membrane and its hydrophobicity and permeability, cellular structure, NP aggregation, and the resulting sizes of particles [15]. Figure 3 presents a graphic illustration of the interactions between NPs and microalgae.
The first point of contact between NPs and cells is the cell wall of algal cells. Green algal cell walls are made up of different components including microfibrillar cellulose, homogalacturonan, rhamnogalacturonan-I, extensin, and arabinogalactan, and they have a comparable composition with that of higher plant cell walls [51]. The accumulation of NPs essentially requires the adsorption of NPs on the functional groups of the algae cell wall [1]. Electrostatic interaction will lead to the adherence of the charged NPs (positive or negative) in the adjacent environment or medium to the cell surface, which is negatively charged [52]. In contrast, uncharged NPs may interact with algae cells by other means. Many varieties of algae have glycoproteins in their cell walls, which are mostly made up of proteins bound to polysaccharides. Hydroxyl, carboxyl, sulfhydryl, amine, amide, imidazole, and others are negatively charged functional groups that can be found in polysaccharides and proteins [53,54], resulting in a high affinity for metal ions in microalgae cells [55,56]. These are the binding sites for metal ions that are transported through the cell membrane and into the cell.
Exopolysaccharides and glycoproteins, which are secreted by a variety of algae and cyanobacteria, may aid in the adsorption of uncharged or neutral NPs on the surfaces of cells that are negatively charged [15]. The nano-sized pores produced after breaking the cell wall allow the NPs to disperse passively into the area between the cell wall and the cell membrane following adsorption [1]. The second stage of intracellular NP uptake is transporting NPs via the cell membrane’s lipid bilayer. The NPs can be transported through the cell membrane by simple diffusion or by an unidentified protein-mediated transmembrane transport route [1]. The common mechanisms of the intracellular uptake of NPs in algae include the presence of pores in the cell membrane, endocytosis, protein-coupled transport, and carrier-mediated transport, all of which have yet to be confirmed [15] (Figure 3).
The pores in the cell walls of algae permit the bidirectional movements of NPs. The smaller the particles, the more easily they diffuse through the cell walls of algae [57]. The diffusion rate of NPs may vary through the cell walls of different algae due to the differences in structure and chemical composition of cell walls. The internalization of NPs into the cell is additionally affected by the aggregation of particles [58]. The uptake of huge aggregates of NPs in algae, on the other hand, has yet to be documented [15]. Uncharged NPs tend to agglomerate more rapidly than positively or negatively charged NPs [58,59]. Furthermore, the charges on the surface of NPs stimulate their adsorption on the surfaces of cells [15]. Positively charged NPs were shown to alter the potential of the cell membrane and eventually disturbed the lipid bilayer and increased the algae uptake of cationic NPs [60]. Many studies considered the effect of NP functionalization on the uptake mechanism. For instance, coating NPs with a polyethylene glycol group (PEG) was found to reduce the cellular uptake of NPs [61,62].
Djearamane et al. [62] used EDX analysis to show cellular accumulations of selenium, zinc, and ZnO-NPs in Spirulina platensis algal biomass [62]. In addition, ICP/OES studies of ZnO-NPs accumulated in algal cells revealed a dose- and time-dependent increase in the cellular accumulation of ZnO-NPs in H. pluvialis, with the greatest zinc accumulation of 6.27–18.36 pg/cell for 10–200 mg/L of ZnO-NPs at 96 h. Gunawan and his colleagues [63] showed that on day 8, when exposed to increasing concentrations of ZnO-NPs from one to 100 mg/L, the green alga Chlamydomonas reinhardtii showed a dose-dependent cellular absorption of zinc, with cellular uptake ranging from 5 fg/cell at 1 mg/L to 18 fg/cell at 100 mg/L [63]. In addition to that, on day 4, duckweed Spirodela polyrhiza was exposed to 1, 10, and 50 mg/L ZnO-NPs, which resulted in dose-dependent cellular accumulations of 2.8, 3.6, and 4.5 mg of zinc/g dry weight [64]. Shen et al. [65] reported that macrophages treated with 10, 50, and 100 mg/L of ZnO-NPs accumulated higher concentrations of zinc—13, 24.6, and 99.4 pg/cell, respectively—than monocytes, which accumulated 0.3, 5.1, and 22.9 pg/cell at 24 h [65].
4. Single and Combined Effects of ZnO-NPs on Marine Microalgae
During the last decade, the number of studies that revealed the effect of ZnO-NPs on organisms living in the marine environment, algae in particular, has increased. Studies have evaluated the effect of ZnO-NPs at various levels (for example, cellular, physiological, molecular, and behavioral). Figure 4 depicts the toxicity of ZnO-NPs on algae.
4.1. Toxicity Mechanisms of ZnO-NPs
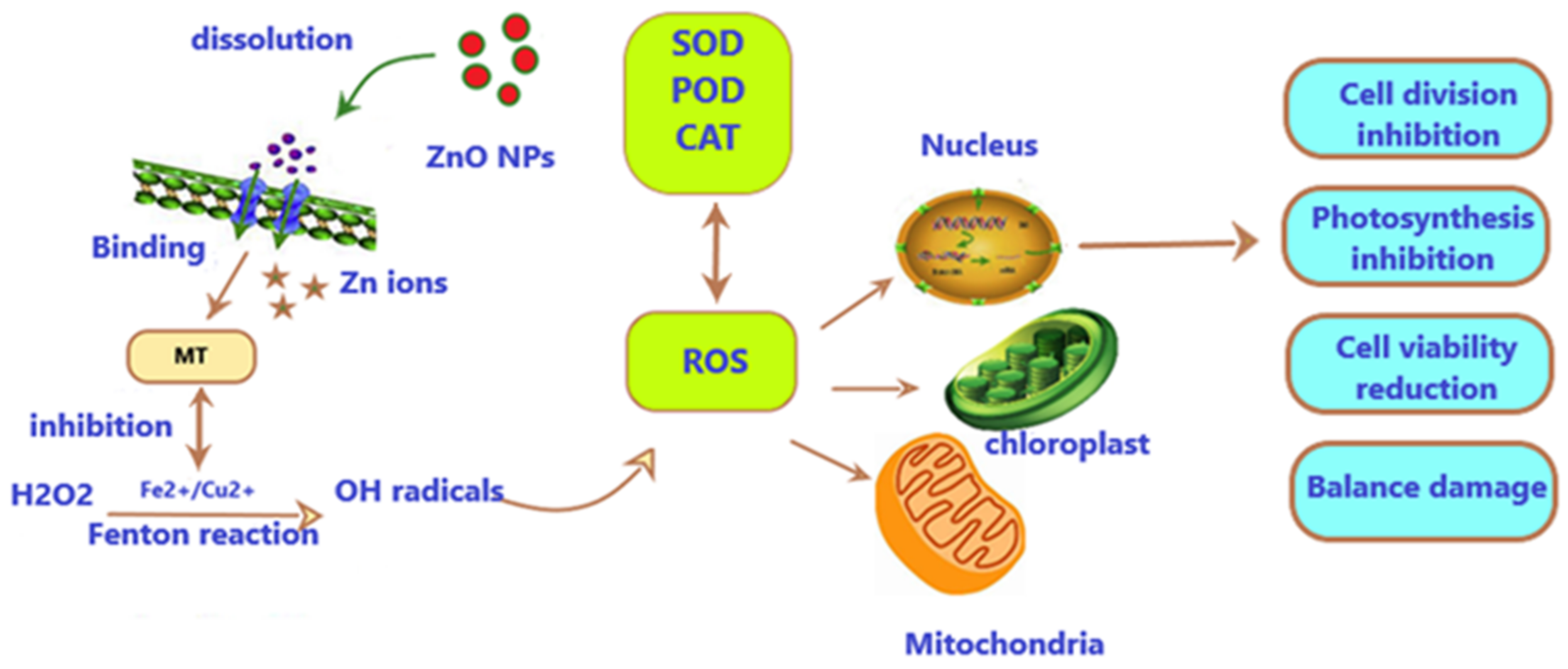
Three major mechanisms are suggested as describing the toxic effects of ZnO-NPs [26]. The first mechanism involves the release of Zn2+ ions (free or complexed with organic or inorganic ligands) in the medium of exposure. The release of Zn2+ ions is a significant factor in the uptake and accumulation of Zn inside cells. Zn2+ from NPs can be transported through ion/voltage-gated channels in the microalgae cell wall and enter the cell [67]. ZnO is more soluble In seawater, with high ionic strength and high pH [68,69,70,71]. Free Zn2+ ions were found to be toxic to many aquatic organisms [72], including marine phytoplankton [73], and dissolved Zn ions have been identified as the main mechanism driving the toxicity of ZnO-NPs in aqueous media [74,75,76]. Figure 5 shows the releasing mechanism of ZnO-NP, where Zn2+ ions (free or complexed with organic or inorganic ligands) are released in the exposure medium.
The second mechanism involves the interaction between NPs and the algae cells that may trigger mechanical damage. For example, the adsorption of NPs or their aggregates onto the cell membrane or the on-site release of ROS on the cell surface may lead to membrane injuries. The toxic effects may also be generated through the accumulation of NPs in the cell when the amount of NPs reaches a level that causes damage to cellular and physiological processes. The third mechanism involves the generation of reactive oxygen species (ROS) under ultraviolet (UV) radiation; this may cause the biological functions of living organisms to be disrupted [23,68,70,77], for instance, lipid peroxidation and DNA damage [78]. It was also indicated that ZnO-NPs pass through the nuclear envelope and enter the nucleus if they are small enough and can cause DNA damage, or they can be internalized through pores in the cell wall during cell division [79,80]. The effect of oxidative stress mediated by metal-based NPs is shown in Figure 6.
4.2. Single Effect of ZnO-NPs
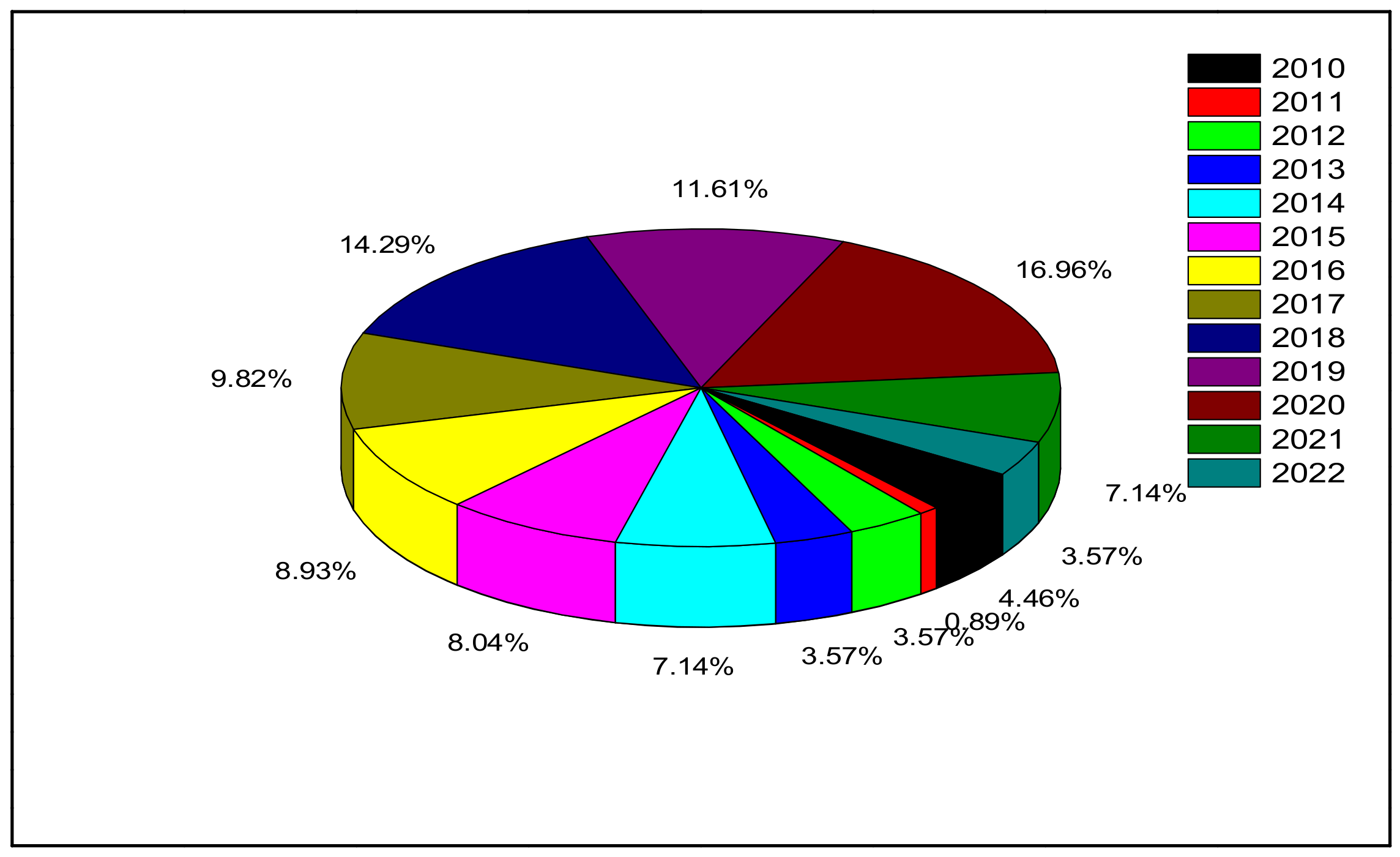
Many investigations of single (individual) cytotoxic effects of ZnO-NPs on marine microalgae have been conducted. Table 2 illustrates the toxicity effects of ZnO-NPs when used individually on different types of marine microalgae. Figure 7 shows the number of published articles in the period between 2010 and 2022.
Numerous studies have suggested that the toxic effects of ZnO-NPs are influenced by the algae species (i.e., species-dependent). For instance, Phaeodactylum tricornutum, a marine diatom, was observed to be less susceptible to ZnO-NPs than Thalassiosira pseudonana, another marine diatom [71]. When exposed to Zn2+ ions released from ZnO-NPs, T. pseudonana showed a considerable reduction in cell division, and the interruption of silicon uptake and frustule development was the trigger [71]. It was suggested that the toxic mechanism of ZnO-NPs was dependent on the physiochemical properties of NPs (e.g., size, shape [71], the amount of zinc ions [69,81], and the production of ROS) [70,77]. Another study by Li et al. [81] displayed that the diatom Phaeodactylum tricornutum was more susceptible than the green algae Tetraselmis suecica [81]. The motility of the green algae in culture media reduced the contact with the aggregates of ZnO-NPs, whereas when P. tricornutum entrapped cells on the test plate’s bottom (i.e., benthic diatom), the interaction could easily happen. Moreover, the shading effects caused a reduction in the light availability, thereby preventing their growth [81,82]. Gong et al. [83] and Li et al. [81] indicated that the effects of ZnO-NPs are linked to the physiochemical states of the NPs in the medium and the interactions between different algae and ZnO-NPs [81,82].
Several researchers ascribed the negative effects of ZnO-NPs to the released zinc ions from the NPs rather than the NPs themselves. For example, Wong et al. [70] found that soluble Zn2+ ions caused a reduction in algae growth and reduced the photosynthetic process in T. pseudonana (a marine diatom) after 48 h exposure [70]. Additionally, they found median inhibition concentrations (IC50) of ZnO-NPs of 4.56 and 2.36 mg/L in T. pseudonana and Skeletonema costatum, respectively. There was a significantly higher IC50 when the same microalgae were exposed to bulk ZnO, 6.65 and 2.97 mg/L for T. pseudonana and S. costatum, respectively, which reflects that ZnO-NPs were more lethal than their bulk counterparts [70].
The toxic effects of ZnO-NPs on the diatom T. pseudonana were linked to dissolved zinc ions by Miao et al. [57,69]. In their study, the release kinetics of Zn2+ were tested in a variety of situations, and tests revealed that pH has a negative relationship with ZnO solubility when the pH is less than 9. ZnO-NPs concentration was found to have little influence on the dissolution behavior of NPs, whereas natural organic compounds (NOC, fulvic acid, and cysteine) had different effects on Zn2+ release according to their chemical compositions and concentrations.
Peng et al. [71] showed that the morphology and aggregates of NPs contributed to the toxicity of ZnO-NPs in addition to the bioavailable Zn2+ ions when P. tricornutum exposed to different sizes and shapes of ZnO-NPs [71]. They identified that ZnO-NPs that are rod-shaped and one-dimensional were more harmful to microalgae than the spherical ZnO-NPs, despite the NPs’ size being less effective. The aggregates of ZnO-NPs present on the surfaces of cells may inhibit the growth of algae. For example, when the marine diatom P. tricornutum was exposed to ZnO-NPs at a concentration of 40 mg/L, there were visible attachments of ZnO-NPs aggregates to the algae cell wall, indicating a type of physicochemical interaction between the aggregates and the algal cell that inhibited growth [71]. The toxicity effect of free Zn2+ ions was also identified based on the adsorption of particles onto different marine algae cells (S. marinoi, T. pseudonana, Dunaliella tertiolecta, and Isochrysis galbana) membranes, which led to cellular damage due to the release of ROS and dissolution of Zn2+ ions, which caused a disruption in the cellular metabolic activity of these organisms [69].
It has also been found that the harmful effect of ZnO-NPs on algae is size dependent. For instance, with a 96-h lowest observed effect concentration (LOEC), of 0.50 and 3.00 mg/L, and 96-h median effective concentrations (EC50) of 2.42 and 4.45 mg/L for 100 nm and 200 nm ZnO, respectively. ZnO-NPs inhibited Dunaliella tertiolecta growth more than their bulk forms [77].
In contrast, Schiavo et al. [84] demonstrated that the toxic effects of ZnO-NPs increased with the increasing concentrations: After 72 h of exposure, ZnO particles hindered cell division (EC50: 2 mg Zn/L), which could be attributed to the release of harmful ions from the aggregate of ZnO particles near the cell membrane. After 72 h of exposure, the proportions of viable cells wer 32% and 26% at 50 and 100 mg Zn/L, respectively. Furthermore, ROS generation increased significantly upon exposure to values greater than 10 mg Zn/L [84].
Schiavo et al. [83] demonstrated that the toxicity of ZnO-NPs was mainly caused by the formation of aggregates larger than ZnO bulk particles and by the entrapping algal cells, leading to the inhibition of cell growth and photosynthetic processes and release of Zn2+ ions close to the cells of the marine microalgae Dunaliella tertiolecta [70,77]. The aggregation of ZnO-NPs in the medium-triggered mechanical damage in the cell wall or caused an indirect effect through shading and limiting nutrient uptake [85]. Moreover, they investigated the toxicity of ZnSO4 (a source of Zn ions) and ZnO-NPs with EC50 24 h (1.8 and 2.2 mg Zn/L for ZnSO4 and ZnO-NPs, respectively) and revealed that the toxicity of ZnO-NPs is not solely due to ion release and that the original nano size adds to the ultimate hazardous effect [70,86].
The toxicity of ZnO-NPs was also found to be concentration- and exposure-time-dependent. Suman et al. [87] examined the effects of about 40 nm ZnO-NPs using different concentrations (10–300 mg/L) on the marine microalgae Chlorella vulgaris [87]. The cytotoxicity of ZnO-NPs was evaluated using the MTT test (3-(4, 5-dimethylthiazol-2-yl) -2, 5-diphenyltetrazolium bromide (MTT) dye reduction assay) and was found to increase with increasing NP concentrations and exposure timse. The intactness of the cell membrane was also assessed using lactate dehydrogenase assay (LDH). An increase in the enzyme in the test solution was an indication of membrane damage and led to cell death. In that experiment, a significant amount of LDH was measured and found to increase over the incubation time and with increases in concentration. Oxidative stress and lipid peroxidation results confirmed the same pattern of ZnO-NPs’ toxic effect. Moreover, the conformation of surface chemical interaction indicated membrane disruption followed by attachment. Zhang et al. [88] found that the 24-, 48-, 72-, and 96-h EC50 of ZnO-NPs (50 nm) for microalgae Skeletonema costatum were 7.0, 6.4, 4.4, and 3.6 mg/L, respectively [88]. Furthermore, the ZnO-NPs were found to be more toxic than the bulk form (>100 nm) of ZnO on microalgae. In their experiment, there was a small variation in the concentration of Zn2+ ions released in the medium that could not explain the different toxic effects between nano-ZnO and bulk-ZnO on microalgae. The inhibition ratio (IR) and intracellular total Zn concentrations of microalgae were found to increase with increasing concentrations of both nano-ZnO and bulk-ZnO. The authors revealed that the intracellular total Zn was the dominant reason for algal growth inhibition. There was a significant increase in the malondialdehyde (MDA) level induced by nano-ZnO in the microalgae compared with the bulk-ZnO at 96 h.
Culture medium was also found to have an impact on the toxic effects of ZnO-NPs on different marine microalgae. Aravantinou et al. [89] used two types of culture medium commonly used for marine algae (salt ⅓N BG-11 and f/2) [89]. These media were chosen because of the differences in ZnSO4.7H2O concentration and, as a result, Zn2+ content. At 96 h, Dunaliella tertiolecta grew slower in f/2 medium than in salt 13N BG-11. In contrast, Tetraselmis suesica showed similar growth rates in the two media. However, the two species were not affected by ZnO-NPs at a concentration of 0.081 mg/L [89]. Total inhibition of algal growth was observed for concentrations ≥ 8.1 mg/L ZnO-NPs.
Some studies showed that the toxic effects of ZnO-NPs can be attributed to multiple factors. Wei and Wang [90] examined the impact of ZnO-NPs with various sizes (30, 90, and 200 nm) and Zn2+ ions on the marine microalgae Chlorella sp. for 96 h [90]. Chlorophyll a concentration was significantly reduced in algae exposed to ZnO-NPs compared with the untreated algae, though the lowest chlorophyll concentration was measured in microalgae treated with Zn2+ ions. This might be explained by the fact that ZnO-NPs aggregated on the algal cells after adsorbing them, and the aggregates adhered to and shielded the algae cells, reducing the amount of light available and limiting light usage [90]. Conversely, the aggregates of the different ZnO-NPs were found to have similar sizes after 96 h, and thus, there was no significant difference in chlorophyll a concentration. Additionally, the possibility of ZnO-NPs’ stimulating oxidative stress was assessed by determining the concentrations of superoxide dismutase (SOD) and catalase (CAT), which are intracellular antioxidant enzymes produced in algae cells under stress conditions. Small ZnO-NPs (30 nm) were found to cause more oxidative stress, which led to high concentrations of SOD and CAT. Lipid peroxidation is another factor that was examined to evaluate the toxicity of ZnO-NPs, specifically by measuring the level of malondialdehyde (MDA). MDA is the major product of the cell membrane’s lipid peroxidation activity, and its quantity indicates the exact degree of cell membrane injury. When a cell membrane is oxidized, its permeability and fluidity alter, potentially increasing Zn2+ ion transmembrane capacity [88]. The microalgae were most affected by ZnO-NPs of 30 nm, demonstrating that the smaller the particle, the more damage to the cell membranes. When there is a reduction in the size of NPs, there will be an increase in the surface area to mass or volume ratio, which has a cascading effect on other chemical and physical properties, such as surface atom reactivity [91]. ZnO-NPs can enter the nucleus through the nuclear pore or be internalized through the cell wall pores during cell division if they are small enough [80,92]. Dobretsov et al. [82] found that small ZnO-NPs (10–30 nm) resulted in greater Zn2+ ion dissolution, and the toxicity of NPs enhanced with increasing concentrations and time of exposure [92].
An unusual study was conducted by Du et al. [93], who evaluated the effects of ZnO-NPs at 10 and 100 mg/L on the swimming speed and locomotion style of Platymonas subcordiformis, a marine microalga, during 24 h [93]. The results of their study indicated that ZnO-NPs may inhibit the motility of P. subcordiformis by limiting the energy available for swimming, as specified by the significantly lower quantities of intracellular ATP and photosynthetic pigments and the reduced activities of enzymes catalyzing glycolysis. Moreover, the authors found an increase in reactive oxygen species (ROS), the accumulation of lipid peroxidation, and the production of antioxidant enzyme activities, which is an indication of oxidative stress, this could harm the structural basis for swimming and as a result could be a contributing factor in the observed motility loss.
ZnO-NPs cause cell damage by the generation of ROS and the expansion of oxidative stress (ROS), which is a well-recognized procedure [94]. By completely occupying oxidative stress defense molecules (e.g., metallothionein), ZnO-NPs might indirectly promote the formation of ROS by inhibiting them from binding Fe2+ or Cu2+, which stay free to catalyze Fenton-type reactions [95]. As a result, cells may experience significant decreases in mitochondrial membrane potential, as well as increases in H2O2 and OH concentrations [84,96]. However, when excessive exposure to ZnO-NPs occurs, antioxidant enzymes may not be sufficient to recover the ROS created. If they are not removed quickly, the balance will be destroyed, resulting in oxidative damage [70].
There is no direct indication of DNA damage in algae when exposed to NPs, although it was suggested that ROS production can cause DNA damage [15]. Schiavo et al. [84] is one of the few researchers who examined the genotoxicity of ZnO-NPs on the marine microalgae Dunaliella tertiolecta and found that up to 5 mg Zn/L, no effects were observed after 72 h of exposure [84]. The number of damaged nuclei rose between 10 and 100 mg Zn/L with the highest percentage of damaged nuclei observed at 25 mg Zn/L, and the highest mutagenic index (MI) of 3.64. Control algae (untreated D. tertiolecta) cells, on the other hand, had a compact nucleus with a well-defined head area and no tail. Instead, comets with unclear head regions and tails of DNA fragments appeared in damaged algal cells [84].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Toxic effects of several ZnO-NPs on various marine algae under various settings.
| Size of ZnO-NPs | Microalgae Species | Growth Conditions (h) | Endpoint (s) | Toxic Effect | Reference |
|---|---|---|---|---|---|
| “20 nm” | “Thalassiosira pseudonana” | “48” | “Growth rate”, | “IC50 = 0.39 µM” The dissolution of ZnO influenced its toxicity and the dissolution rate of ZnO-NPs was accelerated in seawater. | [69] |
| “20 nm” | “Skeletonema costatum” | “96” | “Growth inhibition” | “IC50 = 2.36 mg/L” | [76] |
| “Thalassiosira pseudonana” | IC50 = 4.56 mg/L | ||||
| “20–30 nm” | “Thalassiosira pseudonana” | “96” | “Growth rate” | ZnO-NPs At concentrations between 1 and 1000 µg L−1 reduced the growth rate of all four species | [68] |
| “Skeletonema marinoi” | |||||
| “Dunaliella tertiolecta” | |||||
| “Isochrysis galbana” | |||||
| “6.3–15.7 nm (sphere); 242–862 nm (rod)” | “Thalassiosira pseudonana” | “100” | “Growth inhibition test” | NPs at all concentrations stopped the growth of T. pseudonana and C. gracilis, whereas P. tricornutum was the least sensitive | [71] |
| “Thalassiosira pseudonana” | |||||
| “Phaeodactylum tricornatum” | Rod shaped ZnO with one-dimensional structures were more toxic than spheres NPs | ||||
| “100 nm 200 nm” | “Dunaliella tertiolecta” | “96” | “Growth inhibition” | EC50 = 1.94 mg/L (100 nm) EC50 = 3.57 mg/L (200 nm). The higher toxicity of nano ZnO is related to its physiochemical properties. ZnO-NPs significantly affected the growth rate of D. Tertiolecta. | [77] |
| “25nm” | “Phaeodactylum tricornutum” “Alexandrium minutum” “Tetraselmis suecica” | “Long term” | “Growth inhibition” | “The growth of microalgae was inhibited at concentration more than 10 mg/L. Decrease in chlorophyll fluorescence and high ROS” | [97] |
| “40–48 nm” | “Chlorella vulgaris” | “72” | “Cell viability” “LDH” “Oxidative stress” “Lipid peroxidation” “FTIR analysis” | “The toxicity was concentration and time dependent” | [87] |
| “<100 nm” | “96” | “Growth rate” | Marine species are more sensitive to ZnO-NPs compared to freshwater algae used in the same study. | [89] | |
| “Dunaliella Tertiolecta” | “96-h IC50: 1.5 mg/L (f/2) 96-h IC50: 1.33 mg/L (salt ⅓N BG-11)” | ||||
| “Tetraselmis suecica” | “96-h IC50: 2.1 mg/L (f/2) 96-h IC50: 2.57 mg/L (salt ⅓N BG-11)” | ||||
| “100 nm” | “Dunaliella Tertiolecta” | “72” | “Cell viability” “ROS” “COMET assay” | “EC50: 2.0 mg/L NOEC: 0.1 mg/L” “Significant increase of ROS at ZnO > 10 mg/L at 72-h” “ZnO-NPs was found to be genotoxic at 5 mg/L” | [84] |
| “50 nm” | “Skeletonema Costatum” | “96” | “Growth inhibition” “Lipid peroxidation” | “96-h EC50: 3.6 (50 nm)” “96-h EC50: 5.5 mg/L (5.5 (>100nm)” “High MDA level with the nano-ZnO” | [88] |
| “50 nm” | “Picochlorum sp.” | “32 days” | “Growth inhibition” “Chlorophyll a” | “The toxicity of NPs reduced due to aggregation and sedimentation of NPs after long incubation time” | [98] |
| “100 nm” “200 nm” “ZnSO4” | “Tetraselmis suecica” | “72” | “Growth inhibition” | “72-h EC50 (ZnO 100 nm): 3.91 mg/L 72-h EC50 (ZnO 200 nm): 7.12 mg/L 72-h EC50 (ZnSO4: 5.61 mg/L” | [81] |
| “Phaeodactylum triconutum” | “72-h EC50 (ZnO 100 nm): 1.09 mg/L 72-h EC50 (ZnO 200 nm): 3.47 mg/L 72-h EC50 (ZnSO4: 3.22 mg/L” P. triconatum showed a higher sensitivity than T. suecica | ||||
| “<100 nm” | “Dunaliella Tertiolecta” | “72” | “Growth inhibition” | Marine algae were more sensitive than bacteria and crustaceans used in the same study. “72-h EC50: 2.2 mg/L” | [82] |
| “<100 nm” | “Phaeodactylum tricornutum” | “72” | “Growth inhibition” | “72-h EC50: 2.5–5 mg/L” | [99] |
| “Isochrysis galbana” | “72-h EC50: 5 mg/L” | ||||
| “Tetraselmis suecica” | “72-h EC50: 2.5–5 mg/L” | ||||
| “30, 90 and 200 nm” | “Chlorella sp.” | “96” | “Chlorophyll a” “Oxidative stress (SOD and CAT) 1” “Lipid peroxidation (MDA) 2” | “96-h lowest chlorophyll a concentration was measured in zinc ions” “Highest oxidative stress was detected in cells exposed to 30 nm ZnO-NPs and Zn ions” “Highest MDA measured with 30 nm ZnO-NPs followed by Zn ions.” | [90] |
| “ZnO nano” “ZnO-Bulk” | “Platymonas subcordiformis” | “24” | “Motility of microalgae as a major endpoint in addition to other growth parameters” | “Significant reduction in motility parameters when microalgae exposed to ZnO-NPs”. “No negative effect recorded with bulk ZnO” | [93] |
1 (SOD): Superoxide dismutase; (CAT): Catalase. 2 (MDA): Malondialdehyde.
4.3. Combined Toxicity Effect of ZnO with Other Materials
Under natural environmental conditions, living organisms are subjected to multiple pollutants. Two or more pollutants may employ a synergistic or an antagonistic effect that depends largely on the type of organism, the concentrations, and the prevailing environmental factors. It is anticipated that the number of industrial NPs reaching the environment will further rise in the coming years. This increases the probability that different types of NPs could be found in the same medium; therefore, the harmful effects of each NP either could change, be hindered, or stay unaffected by the occurrence of the other NPs. It is well recognized that in the marine environment, pollutants coexist, and their combined effects on the marine organisms cannot be neglected. However, there is limited literature on the assessment of the combined effect of ZnO-NPs with other pollutants on marine microalgae.
Hazeem et al. [98] for example examined the combined effect of ZnO and TiO2 NPs on the marine microalgae Picochlorum sp. in addition to the individual effect of these NPs over 32 days [98]. It was found that the availability and toxicity of NPs to the microalgae were reduced, which was attributed to the aggregation of NPs. However, there were no significant difference between the individual and combined effects of the NPs during the late growth stages, which was explained by the fact that microalgae can reverse the harmful effects of NPs after long exposure through sedimentation of the large aggregates [98].
Schiavo et al. [100] evaluated the impacts of TiO2 and ZnO-NPs obtained from commercially produced sunscreen products (S2, S10, S11) on Dunaliella tertiolecta [100], the growth of algae was affected by the obtained NPs (“EC20 S2: 9.2 mg Ti L−1; EC20 S10: 0.49 mg Ti L−1; EC20 S11: 14 mg Ti L−1”); additionally, the microalgae were largely induced by the formulations of sunscreen, which could have been the result of certain mechanisms that increased the growth of algae. In the sunscreens that contained both ZnO and TiO2 (S10; S11), ROS was produced, and there was DNA damage in algae cells. Lower effects were noticed when using sunscreens and the resultant extracts in comparison with the impacts of manufactured NPs. The results of that study indicated that the formulations of products may alleviate the inhibitory influences of NPs by direct modifications of NP features or by supplying inorganic and organic nutrients that are essential in microalgae to promote their growth. The aging of sunscreens may also control the impact of NPs.
Gunasekaran et al. [101] examined the impacts of microplastics that are uncharged at 1 mg/L on the toxic impacts of bulk and nano-sized ZnO particles at 1.22, 12.28 and 122.88 μM with either dark or UV-A exposure conditions [101]. Viable cell concentration, the generation of ROS, lipid peroxidation, dissolution, and zinc salt toxicity were analyzed. Dunaliella salina was selected as a model marine microalga and the toxicity test was carried out for 72 h. The authors found that cell viability decreased with increasing doses of both the nano and bulk ZnO particles under the test conditions (UV-A illumination and dark conditions). The uncharged PS microplastic at 1 mg/L was found to enhance the cell viability of the microalgae with both nano and bulk ZnO at all test concentrations. Extracellular and intracellular reactive oxygen species (ROS) were found to increase and be well correlated with the concentrations of nano and bulk ZnO. When they were mixed with uncharged PS microplastics, there was a decrease in ROS production. The reduction in toxicity of nano and bulk ZnO in the presence of microplastic was attributed to the reduction in the hetero aggregation of ZnO with microalgae, and therefore, there was a reduction in algae entrapment. The uncharged PS microplastic functioned as a barrier and inhibited the interactions between negatively charged microalgae and the positively charged NPs, in turn reducing the formation of hetero aggregates. The reduction in lipid peroxidation production indicated that the presence of microplastic in the medium reduced the cell membrane damage from D. salina [101].
Baek et al. [102] evaluated the effects of ZnO with carbon-based nanohybrids (NHs) such as ZnO conjugated with graphene oxide (ZnO-GO) and ZnO-conjugated carbon nanotubes (ZnO-CNTs) on the marine diatom Thalassiosira pseudonana [102]. The microalgae were incubated with the materials for 72 h, and the test chemicals were used at concentrations between 50 and 100 mg/L. The results indicated that ZnO-GO was the most lethal NHs to the microalgae. The toxicity mechanism was ascribed to the production of reactive oxygen species and that graphene oxide (GO) caused dispersion that involved electrostatic repulsion and prevented particles from aggregation, and there was a rise in the surface area of NHs. The analysis using scanning electron microscopy (SEM) showed that NHs shielded microalgae cells and interacted with them (shading effects); this reduced the available light for photosynthetic processes. The toxicity of NHs might be attributed to damage in the cell membrane triggered by the attachment of NH [102].
Li et al. [103] evaluated the combined effects of ZnO-NPs with polyethylene microplastic (PEMP) on the marine microalga Tertraselmis helgolandica var. tsingtaoensis. It was found that the presence of PEMP decreased the inhibition effect of ZnO-NPs by 14.4%. They hypothesized that ZnO-NPs’ adsorption through weak attraction forces onto PEMP decreased its exposure level to the microalga, directly affecting cellular growth [103].
Another study examined the combined toxicity effects of ZnO-NPs and graphene quantum dots (GQDs) using the marine dinoflagellate Gymnodinium sp. Zhu et al. [104] exposed microalgae to 1, 10, and 20 mg/L ZnO-NPs and GQDs and demonstrated that the growth inhibition rates of the microalgae increased with increases in GQD concentration. The combined exposure of both materials reduced the inhibitory effect and showed antagonistic effects. ZnO-NPs were found to decrease the toxicity of GQDs, which should precede the aggregation and sedimentation between NPs [104]. Table 3 is a summary of the published articles on the combined effects of ZnO-NPs with other pollutants using different types of marine microalgae.
5. Critical Evaluation of the Existing Literature on the Single and Combined Effects of ZnO-NPs on Marine Microalgae
Living organisms are usually exposed to a mixture of pollutants in any natural ecosystem. The presence of two or more contaminants at the same time may alter the impact of each contaminant, and this depends on multiple environmental factors and features of the living organisms under examination [15]. According to the literature collected between 2010 and 2022 that was available during the writing of this review, most toxicity studies were conducted on single ZnO-NPs with different particle sizes (~6 nm–100 nm); in a few cases, findings were compared with the bulk form of ZnO (>100 nm). Different marine microalgae species were tested using standard growth toxicity conditions. One research group, Hazeem et al. [98], was among the first (Table 2) to use long-term toxicity tests (32 days growth inhibition test), and the authors found that the toxicity of ZnO-NPs was reduced due to aggregation and sedimentation after long incubation times. In contrast, the acute toxicity tests revealed that ZnO-NPs were toxic throughout the incubation period.
Research studies on the potential threats of a mixture of contaminants are still limited. Acute toxicity tests were carried out by most investigators (96 h incubation tests) using ZnO-NPs mixed with only one type of contaminant (e.g., TiO2 NPs, polystyrene/polyethylene microplastic and GO) (Table 3). There are contradictory results that reveal the need for more risk assessments of different mixtures of contaminants using different organisms under relevant environmental conditions. It is essential to conduct more comparative studies between the single effect of ZnO-NPs and its effects when mixed with other contaminants under the same experimental conditions. Additionally, most toxicity tests use cellular, physiological, and behavioral responses as the main endpoints, but it is important to assess the impact of contaminants at the molecular level.
6. Research, Knowledge Gap and Recommendations:
Based on this extensive evaluation of related literature, numerous knowledge gaps have been identified that must be addressed in order to improve our current understanding of the toxicity of ZnO-NPs for environmental risk assessment and the management of this widely marketed product category.
- Toxic mechanisms of ZnO-NPs at the molecular level in marine organisms at different trophic levels should be examined using advanced techniques including transcriptomic, proteomic, and metabolomic methods to discriminate the diverse modes of toxicity of ZnO-NPs when used separately or combined with other materials.
- Research should be performed on the toxicity of ZnO-NPs under environmentally relevant UV radiation because of its photoactive properties. The assessments should be applied using different marine organisms at different trophic levels. ROS released due to oxidative stress imposed by the exposure to ZnO-NPs should also be investigated.
- Acute and chronic toxicity studies are needed to better understand the effects of ZnO-NPs on various marine organisms at various stages of development.
- Trophic transfer of ZnO-NPs should be examined for the accumulation, cellular localization, or biotransformation of NPs in a well-defined marine food chain.
- There have been few investigations of the combined effect of ZnO-NPs and other contaminants. It should be addressed properly in a variety of environmental conditions, including temperature, pH, UV radiation, and the presence of dissolved organic material. The data could be used to improve water quality criteria and thereby conserve marine life by limiting the usage and release of ZnO-NPs.
- Evaluating surface modification of ZnO-NPs is required. Methods should be developed to maintain the initial chemical features of ZnO-NPs while decreasing their biological toxicity.
- There is to date a substantial lack of characterization of ZnO-NPs and theirs exposure system in toxicity studies. The effects of ZnO-NPs depend on exposure conditions, for instance, water chemistry and irradiation. It is essential to have a detailed characterization of the exposure conditions to effectively explain the toxicity data.
- Certain international regulations and policies should be applied for the production and use of NPs. Furthermore, wastes containing NPs should be well treated before disposal. Public awareness programs should alert people to take extra care in the use and disposal of NPs.
- Cost-effective methods for the elimination and retrieval of NPs from waste before disposal should be established to retrieve and reuse the original forms of the NPs.
7. Conclusions
The continuous discharge of ZnO-NPs into aquatic environments due to their extensive application has vital impacts on living organisms. In this review, the existing literature on the toxicity effects of ZnO-NPs on marine microalgae revealed that several factors affect the toxicity of ZnO-NPs when assessed singly or when mixed with other contaminants. Some of these factors are related to the physiochemical properties of ZnO-NPs, and others are related to the organisms exposed to NPs and the surrounding environments. Consequently, ZnO-NPs might not follow the same toxicity mechanism under different conditions. Hence, research should consider multiple factors to determine precise and consistent toxicity assessment results. Most studies were carried out by examining single effects of ZnO-NPs; therefore, more research is required to compare the single and combined effects of ZnO-NPs with other contaminants under relevant environmental conditions. Additionally, there are adequate data on the toxicity of ZnO-NPs at cellular, physiological, and behavioral levels of marine microalgae. However, to explain the internal mechanism of action, the toxicity effect of ZnO-NPs should be well oriented at the molecular level. Lastly, to improve our evaluation and understanding of the toxicity mechanism of ZnO-NPs, long term toxicity test should be employed.
Funding
The author would like to thank the generous fund from the University of Bahrain for supporting this part of the research.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
I am grateful to my colleague Suad Rashdan for her valuable comments on the manuscript. I also thank Ayman Helmi for his generosity and extensive professional expertise and guidance and for proofreading the final version of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Nguyen, M.K.; Moon, J.Y.; Lee, Y.C. Microalgal ecotoxicity of nanoparticles: An updated review. Ecotoxic. Environ. Saf. 2020, 201, 110781. [Google Scholar] [CrossRef] [PubMed]
- Shevlin, D.; O’Brien, N.; Cummins, E. Silver engineered nanoparticles in freshwater systems-likely fate and behavior through natural attenuation processes. Sci. Total. Environ. 2018, 621, 1033–1046. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, Applications and Toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Adochite, C.; Andronic, L. Aquatic Toxicity of Photocatalyst Nanoparticles to Green Microalgae Chlorella vulgaris. Water 2021, 13, 77. [Google Scholar] [CrossRef]
- Ma, S.; Zhou, K.; Yang, K.; Lin, D. Hetero agglomeration of oxide nanoparticles with algal cells: Effects of particle type, ionic strength and pH. Environ. Sci. Technol. 2015, 49, 932–939. [Google Scholar] [CrossRef]
- Li, S.; Ma, H.; Wallis, L.K.; Etterson, M.A.; Riley, B.; Hoff, D.J.; Diamond, S.A. Impact of natural organic matter on particle behavior and phototoxicity of titanium dioxide nanoparticles. Sci. Total. Environ. 2016, 542, 324–333. [Google Scholar] [CrossRef]
- Nolte, T.M.; Hartmann, N.B.; Kleijn, J.M.; Garnæs, J.; van de Meent, D.; Jan Hendriks, A.; Baun, A. The toxicity of plastic nanoparticles on green algae as influenced by surface modification, medium hardness and cellular adsorption. Aquat. Toxicol. 2017, 183, 11–20. [Google Scholar] [CrossRef]
- Sangchay, W. The Self-cleaning and Photocatalytic Properties of TiO2 Doped with SnO2, Thin Films Preparation by Sol-gel Method. Energy Procedia 2016, 89, 170–176. [Google Scholar] [CrossRef]
- Ali, I.; Alharbi, O.M.L.; Tkachev, A.; Galunin, E.; Burakov, A.; Grachev, V.A. Water treatment by new-generation graphene materials: Hope for bright future. Environ. Sci. Pollut. Res. 2018, 25, 7315–7329. [Google Scholar] [CrossRef]
- Corma, A.; Atienzar, P.; García, H.; Chane-Ching, J.-Y. Hierarchically mesostructured doped CeO2 with potential for solar-cell use. Nat. Mater. 2004, 3, 394–397. [Google Scholar] [CrossRef]
- Piccinno, F.; Gottschalk, F.; Seeger, S.; Nowack, B. Industrial production quantities and uses of ten engineered nanomaterials in Europe and the world. J. Nanopart. Res. 2012, 14, 1109. [Google Scholar] [CrossRef]
- Handy, R.D.; Owen, R.; Valsamijones, E. The ecotoxicology of nanoparticles and nanomaterials: Current status, knowledge gaps, challenges, and future needs. Ecotoxicology 2008, 17, 315–325. [Google Scholar] [CrossRef]
- Rogers, F.; Arnott, P.; Zielinska, B.; Sagebiel, J.; Kelly, K.E.; Wagner, D.; Lighty, J.S.; Sarofim, A.F. Real-Time Measurements of Jet Aircraft Engine Exhaust. J. Air Waste Manag. Assoc. 2005, 55, 583–593. [Google Scholar] [CrossRef]
- Seames, W.S.; Fernandez, A.; Wendt, J.O.L. A study of fine particulate emissions from combustion of treated pulverized municipal sewage sludge. Environ. Sci. Technol. 2002, 36, 2772–2776. [Google Scholar] [CrossRef]
- Mahana, A.; Guliy, O.I.; Mehta, S.K. Accumulation and cellular toxicity of engineered metallic nanoparticle in freshwater microalgae: Current status and future challenges. Ecotoxicol. Environ. Saf. 2021, 208, 111662. [Google Scholar] [CrossRef]
- Junam, Y.; Lead, J.R. Manufactured nanoparticles: An overview of their chemistry, interactions and potential environmental implications. Sci. Total Environ. 2008, 400, 396–414. [Google Scholar] [CrossRef]
- Trojanowski, R.; Fthenakis, V. Nanoparticle emissions from residential wood combustion: A critical literature review, characterization, and recommendations. Renew. Sustain. Energy Rev. 2019, 103, 515–528. [Google Scholar] [CrossRef]
- Chen, C.; Huang, W. Aggregation kinetics of diesel soot nanoparticles in wet environments. Environ. Sci. Technol. 2017, 51, 2077–2086. [Google Scholar] [CrossRef]
- Somasundaran, P.; Fang, X.; Ponnurangam, S.; Li, B. Nanoparticles: Characteristics, mechanisms, and modulation of biotoxicity. KONA Power Part. J. 2010, 28, 38–49. [Google Scholar] [CrossRef]
- Lu, P.J.; Huang, S.C.; Chen, Y.P.; Chiueh, L.C.; Shih, D.Y.C. Analysis of titanium dioxide and zinc oxide nanoparticles in cosmetics. J. Food Drug Anal. 2015, 23, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Bairi, V.G.; Lim, J.-H.; Fong, A.; Linder, S.W. Size characterization of metal oxide nanoparticles in commercial sunscreen products. J. Nanopart. Res. 2017, 19, 256. [Google Scholar] [CrossRef]
- Fatehah, M.O.; Aziz, H.A.; Stoll, S. Nanoparticle Properties, behavior, Fate in Aquatic Systems and Characterization Methods. J. Colloid Sci. Biotechnol. 2014, 3, 111–1140. [Google Scholar] [CrossRef]
- McIntyre, R.A. Common nanomaterials and their use in real world applications. Sci. Prog. 2012, 95, 1–22. [Google Scholar] [CrossRef]
- Smijs, T.G.; Pavel, S. Titanium dioxide and zinc oxide nanoparticles in sunscreens: Focus on their safety and effectiveness. Nanotechnol. Sci. Appl. 2011, 4, 95–112. [Google Scholar] [CrossRef]
- Mirzaei, H.; Darroudi, M. Zinc oxide nanoparticles: Biological synthesis and biomedical applications. Ceram. Int. 2017, 43, 907–914. [Google Scholar] [CrossRef]
- Ma, H.; Williams, P.L.; Diamond, S.A. Ecotoxicity of manufactured ZnO nanoparticles—A review. Environ. Pollut. 2013, 172, 76–85. [Google Scholar] [CrossRef]
- Gottschalk, F.; Sonderer, T.; Scholz, R.W.; Nowack, B. Modeled Environmental Concentrations of Engineered Nanomaterials (TiO2, ZnO, Ag, CNT, Fullerenes) for Different Regions. Environ. Sci. Technol. 2009, 43, 9216–9222. [Google Scholar] [CrossRef]
- Mehta, S.K.; Gaur, P. Use of Algae for Removing Heavy Metal Ions From Wastewater: Progress and Prospect. Crit. Rev. Biotechnol. 2005, 25, 113–152. [Google Scholar] [CrossRef]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B.M. Microalgae and wastewater treatment. Saudi J. Biol. Sci. 2012, 19, 257–275. [Google Scholar] [CrossRef]
- Nowack, B. The occurrence, behavior, and effects of engineered nanomaterials in the environment. In Advances in Nanotechnology and the Environment; Kim, J., Ed.; Pan Stanford: Singapore, 2012; pp. 197–207. [Google Scholar]
- Boxall, A.B.A.; Chaudhry, Q.; Sinclair, C.; Jones, A.D.; Aitken, R.; Jefferson, B.; Watts, C. Current and Future Predicted Environmental Exposure to Engineered Nanoparticles; Central Science Laboratory, Department of the Environment and Rural Affairs: London, UK, 2007; 89p.
- Gottschalk, F.; Sun, T.Y.; Nowack, B. Environmental concentrations of engineered nanomaterials: Review of modeling and analytical studies. Environ. Pollut. 2013, 181, 287–300. [Google Scholar] [CrossRef]
- Gottschalk, F.; Sonderer, T.; Scholz, R.W.; Nowack, B. Possibility and limitations of modeling environmental exposure to engineered nanomaterials by probabilistic material flow analysis. Environ. Toxicol. Chem. 2010, 29, 1036–1048. [Google Scholar] [CrossRef]
- Tiede, K.; Hassellöv, M.; Breitbarth, E.; Chaudhry, Q.; Boxall, A.B.A. Considerations for environmental fate and ecotoxicity testing to support environmental risk assessments for engineered nanoparticles. J. Chromatogr. A 2009, 1216, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Fairbairn, E.A.; Keller, A.A.; Madler, L.; Zhou, D.; Pokhrel, S. Metal oxide nanoparticles in seawater: Linking physiochemical characteristics with biological response in sea urchin development. J. Hazard. Mater. 2011, 192, 1565–1571. [Google Scholar] [CrossRef] [PubMed]
- Bian, S.W.; Mudunkotuwa, I.A.; Rupasinghe, T.; Grassian, V.H. Aggregation and dissolution of 4nm ZnO nanoparticles in aqueous environments: Influence of pH, ionic strength, size, and adsorption of humic acid. Langmuir 2011, 27, 6059–6068. [Google Scholar] [CrossRef]
- Tang, E.; Cheng, G.; Ma, X.; Pang, X.; Zhao, Q. Surface modification of zinc oxide nanoparticle by PMAA and its dispersion in aqueous system. Appl. Surf. Sci. 2006, 252, 5227–5232. [Google Scholar] [CrossRef]
- Lowry, G.V.; Wiesner, M.R. Environmental considerations: Occurrences, fate, and characterization of nanoparticles in the environment. In Nanotoxicology: Characterization, Dosing and Health Effects; Monteiro-Riviere, N.A., Lang Tran, C., Eds.; Informa Healthcare: New York, NY, USA, 2007; pp. 369–389. [Google Scholar]
- Morales, J.; Manso, J.A.; Cid, A.; Mejuto, J.C. Degradation of carbofuran and carbofuran derivatives in presence of humic substances under basic conditions. Chemosphere 2012, 89, 1267–1271. [Google Scholar] [CrossRef]
- Thio, B.J.R.; Zhou, D.; Keller, A.A. Influence of natural organic matter on the aggregation and deposition of titanium dioxide nanoparticles. J. Hazard. Mater. 2011, 189, 556–563. [Google Scholar] [CrossRef]
- Litvin, V.A.; Galagan, R.L.; Minaev, B.F. Kinetic and mechanism formation of silver nanoparticles coated by synthetic humic substances. Colloids Surf. A Physicochem. Eng. Asp. 2012, 414, 234–243. [Google Scholar] [CrossRef]
- Sutton, R.; Sposito, G. Molecular structure in soil humic substances: The new view. Environ. Sci. Technol. 2005, 39, 9009–9015. [Google Scholar] [CrossRef]
- Ojwang’, L.M.; Cook, R.L. Environmental conditions that influence the ability of humic acids to induce permeability in model bio membranes. Environ. Sci. Technol. 2013, 47, 8280–8287. [Google Scholar]
- Tenzer, S.; Doctor, D.; Kuharev, J.; Musyanovych, A.; Fetz, V.; Hecht, R.; Schlenk, F.; Fischer, D.; Kiouptsi, K.; Reindhardt, C.; et al. Rapid formation of plasma protein corona critically affects nanoparticle pathophysiology. Nat. Nanotechnol. 2013, 8, 772–781. [Google Scholar] [CrossRef]
- Nowak, B.; Bucheli, T.D. Occurrence, behavior and effects of nanoparticles in the environment. Environ. Pollut. 2007, 150, 5–22. [Google Scholar] [CrossRef]
- Efthimiou, I.; Georgiou, Y.; Vlastos, D.; Dailianis, S.; Deligiannakis, Y. Assessing the cyto-genotoxic potential of model zinc oxide nanoparticles in the presence of humic-acid-like-poly condensate (HALP) and the leaonardite HA (LHA). Sci. Total Environ. 2020, 721, 137625. [Google Scholar] [CrossRef]
- Yunga, M.M.N.; Mouneyracb, C.; Leung, K.M.Y. Ecotoxicity of Zinc Oxide Nanoparticles in the Marine Environment. In Encyclopedia of Nanotechnology; Bhushan, B., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 1–17. [Google Scholar]
- Han, Y.; Kim, D.; Hwang, G.; Lee, B.; Eom, I.; Kim, P.J.; Tong, M.; Kim, H. Aggregation and dissolution of ZnO nanoparticles synthesized by different methods: Influence of ionic strength and humic acid. Colloids Surf. A 2014, 451, 7–15. [Google Scholar] [CrossRef]
- Wang, X.; Lu, J.; Xu, M.; Xing, B. Sorption of pyrene by regular and nanoscaled metal oxide particles: Influence of adsorbed organic matter. Environ. Sci. Technol. 2008, 42, 7267–7272. [Google Scholar] [CrossRef]
- Lipovsky, A.; Tzitrinovich, Z.; Friedmann, H.; Applerot, G.; Gedanken, A.; Lubart, R. EPR study of visible light-induced ROS generation by nanoparticles of ZnO. J. Phys. Chem C 2009, 113, 15997–16001. [Google Scholar] [CrossRef]
- Estevez, J.M.; Kasuli, L.; Fernandez, P.V.; Dupree, P.; Ciancia, M. Chemical in situ characterization of macromolecular components of the complex cell walls from the coenocytic green alga Codium fragile. Glycobiology 2009, 18, 250–259. [Google Scholar]
- Shankar, P.D.; Shobana, S.; Karuppusamy, I.; Pugazhendhi, A.; Ramkumar, V.S.; Arvindnarayan, S.; Kumar, G. A review on the biosynthesis of metallic nanoparticles (gold and silver) using bio-components of microalgae: Formation mechanism and applications. Enzyme Microb. Technol. 2016, 95, 28–44. [Google Scholar] [CrossRef]
- Ting, Y.P.; Prince, I.G.; Lawson, F. Uptake of cadmium and zinc by the alga Chlorella vulgaris: II. Multi-ion situation. Biotechnol. Bioeng. 1991, 37, 445–455. [Google Scholar] [CrossRef]
- Mehta, S.K.; Gaur, J.P. Heavy-metal-induced proline accumulation and its role in ameliorating metal toxicity in Chlorella vulgaris. New Phytol. 1999, 143, 253–259. [Google Scholar] [CrossRef]
- Crist, R.H.; Martin, J.R.; Guptill, P.W.; Eslinger, J.M.; Crist, D.R. Interaction of Metal Ions with Acid Sites of Bio sorbents Peat Moss and Vaucheria and Model Substances Alginic and Humic Acids. Environ. Sci. Technol. 1990, 24, 337–342. [Google Scholar] [CrossRef]
- Crist, R.H.; Martin, J.R.; Carr, D.; Watson, J.R.; Clarke, H.J.; Crist, D.R. Interaction of metals and protons with algae. Ion-exchange vs adsorption models and a reassessment of Scatchard plots; ion-exchange rates and equilibria compared with calcium alginate. Environ. Sci. Technol. 1994, 28, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, N.P.; Hurst, G.B.; Wang, W.; Foster, C.M.; Nallathamby, P.D.; Retterer, S.T. Dynamic development of the protein corona on silica nanoparticles: Composition and role in toxicity. Nanoscale 2013, 5, 6372–6380. [Google Scholar] [CrossRef] [PubMed]
- Izak-Nau, E.; Voetz, M.; Eiden, S.; Duschl, A.; Puntes, V.F. Altered characteristics of silica nanoparticles in bovine and human serum: The importance of nanomaterial characterization prior to its toxicological evaluation. Part. Fibre Toxicol. 2013, 10, 56. [Google Scholar] [CrossRef]
- Kim, S.T.; Saha, K.; Kim, C.; Rotello, V.M. The role of surface functionality in determining nanoparticle cytotoxicity. Acc. Chem. Res. 2013, 46, 681–691. [Google Scholar] [CrossRef]
- Ehrenberg, M.S.; Friedman, A.E.; Finkelstein, J.N.; Oberdörster, G.; Mcgrath, J.L. The influence of protein adsorption on nanoparticle association with cultured endothelial cells. Biomaterials 2009, 30, 603–610. [Google Scholar] [CrossRef]
- Verma, A.; Stellacci, F. Effect of surface properties on nanoparticle-cell interactions. Small 2010, 6, 12–21. [Google Scholar] [CrossRef]
- Djearamane, S.; Wong, L.S.; Mooi, L.Y.; Lee, P.F. Cytotoxic effects of zinc oxide nanoparticles on cyanobacterium Spirulina (Arthrospira) platensis. PeerJ 2018, 6, e4682. [Google Scholar] [CrossRef]
- Gunawan, C.; Sirimanoonphan, A.; Teoh, W.Y.; Marquis, C.P.; Amal, R. Submicron and nano formulations of titanium dioxide and zinc oxide stimulate unique cellular toxicological responses in the green microalga Chlamydomonas reinhardtii. J. Hazard. Mater. 2013, 260, 984–992. [Google Scholar] [CrossRef]
- Hu, C.; Liu, Y.; Li, X.; Li, M. Biochemical responses of duckweed (Spirodela polyrhiza) to zinc oxide nanoparticles. Arch. Environ. Contam. Toxicol. 2013, 64, 643–651. [Google Scholar] [CrossRef]
- Shen, C.; James, S.A.; De Jonge, M.D.; Turney, T.W.; Wright, P.F.A.; Feltis, B.N. Relating cytotoxicity, zinc ions, and reactive oxygen in ZnO nanoparticle-exposed human immune cells. Toxicol. Sci. 2013, 136, 120–130. [Google Scholar] [CrossRef]
- Hou, J.; Wu, Y.; Li, X.; Wei, B.; Li, S.; Wang, X. Toxic effects of different types of zinc oxide nanoparticles on algae, plants, invertebrates, vertebrates and microorganisms. Chemosphere 2018, 193, 852–860. [Google Scholar] [CrossRef]
- Colvin, R.A.; Fontaine, C.P.; Laskowski, M.; Thomas, D. Zn2+ transporters and Zn2+ homeostasis in neurons. Eur. J. Pharmacol. 2003, 479, 171–185. [Google Scholar] [CrossRef]
- Miller, R.J.; Lenihan, H.S.; Muller, E.B.; Tseng, N.; Hanna, S.K.; Keller, A.A. Impact of metal oxide nanoparticles on marine phytoplankton. Environ. Sci. Technol. 2010, 44, 7329–7334. [Google Scholar] [CrossRef]
- Miao, A.J.; Zhang, X.Y.; Luo, Z.; Chen, C.S.; Chin, W.C.; Santschi, P.H.; Quigg, A. Zinc oxide engineered nanoparticles: Dissolution and toxicity to marine phytoplankton. Environ. Toxicol. Chem. 2010, 29, 2814–2822. [Google Scholar] [CrossRef]
- Wong, S.W.Y.; Leung, P.T.Y.; Djurišić, A.B.; Leung, K.M.Y. Toxicities of nano zinc oxide to five marine organisms: Influences of aggregate size and ion solubility. Anal. Bioanal. Chem. 2010, 396, 609–618. [Google Scholar] [CrossRef]
- Peng, X.; Palma, S.; Fisher, N.S.; Wong, S.S. Effect of morphology of ZnO nanostructures on their toxicity to marine algae. Aquat. Toxicol. 2011, 102, 186–196. [Google Scholar] [CrossRef]
- Eisler, R. Zinc Hazards to Fish, Wildlife, and Invertebrates: A Synoptic Review; No. 26; U.S. Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 1993; p. 106.
- Brand, L.E.; Sunda, W.G.; Guillard, R.R.L. Limitation of Marine Phytoplankton Reproductive Rates by Zinc, Manganese, and Iron. Limnol. Oceanogr. 1983, 28, 1182–1198. [Google Scholar] [CrossRef]
- Franklin, N.M.; Rogers, N.J.; Apte, S.C.; Batley, G.E.; Gadd, G.E.; Casey, P.S. Comparative toxicity of nanoparticulate ZnO, bulk ZnO, and ZnCl2 to a freshwater microalga (Pseudokirchneriella subcapitata): The importance of particle solubility. Environ. Sci. Technol. 2007, 41, 8484–8490. [Google Scholar] [CrossRef]
- Aruoja, V.; Kahru, A.; Dubourguier, H.C. Toxicity of ZnO, TiO2 and CuO nanoparticles to microalgae Pseudokirchneriella subcapitata. Toxicol. Lett. 2008, 180, S220. [Google Scholar] [CrossRef]
- Xia, T.; Kovochich, M.; Liong, M.; Madler, L.; Gilbert, B.; Shi, H.; Yeh, J.; Zink, J.I.; Nel, A.E. Comparison of the mechanism of toxicity of zinc oxide and cerium oxide nanoparticles based on dissolution and oxidative stress properties. ACS Nano 2008, 2, 2124–2134. [Google Scholar] [CrossRef]
- Manzo, S.; Miglietta, M.L.; Rametta, G.; Buono, S.; Di Francia, G. Toxic effects of ZnO nanoparticles towards marine algae Dunaliella tertiolecta. Sci. Total Environ. 2013, 445, 371–376. [Google Scholar] [CrossRef]
- Maness, P.C.; Huang, Z.; Smolinski, S.; Jacoby, W.; Blake, D.; Wolfrum, E. Photosterilization and photominerization of microbial cells with titanium dioxide. Photochem. Photobiol. 1999, 69, 64s–65s. [Google Scholar]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of silver nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 2008, 42, 8959–8964. [Google Scholar] [CrossRef]
- Magdolenova, Z.; Collins, A.; Kumar, A.; Dhawan, A.; Stone, V.; Dusinska, M. Mechanisms of genotoxicity. A review of in vitro and in vivo studies with engineered nanoparticles. Nanotoxicology 2014, 8, 233–278. [Google Scholar] [CrossRef]
- Li, J.; Schiavo, S.; Rametta, G.; Miglietta, M.L.; Ferrara, V.L.; Wu, C.; Manzo, S. Comparative toxicity of nano ZnO and bulk ZnO towards marine algae Tetraselmis suecica and Phaeodactylum tricornutum. Environ. Sci. Pollut. Res. 2017, 24, 6543–6553. [Google Scholar] [CrossRef]
- Schiavo, S.; Oliviero, M.; Li, J.; Manzo, S. Testing ZnO nanoparticle ecotoxicity: Linking time variable exposure to effect on different marine model organisms. Environ. Sci. Pollut. Res. 2018, 25, 4871–4880. [Google Scholar] [CrossRef]
- Gong, N.; Shao, K.; Feng, W.; Lin, Z.; Liang, C.; Sun, Y. Biotoxicity of nickel oxide nanoparticles and bio-remediation by microalgae Chlorella vulgaris. Chemosphere 2011, 83, 510–516. [Google Scholar] [CrossRef]
- Schiavo, S.; Oliviero, M.; Miglietta, M.; Rametta, G.; Manzo, S. Genotoxic and cytotoxic effects of ZnO nanoparticles for Dunaliella tertiolecta and comparison with SiO2 and TiO2 effects at population growth inhibition levels. Sci. Total Environ. 2016, 550, 619–627. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, X.; Lao, Y.; Lv, X.; Tao, Y.; Huang, B.; Wange, J.; Zhou, J.; Cai, Z. TiO2 nanoparticles in the marine environment: Physical effects responsible for the toxicity on algae Phaeodactylum tricornutum. Sci. Total Environ. 2016, 565, 818–826. [Google Scholar] [CrossRef]
- Ji, J.; Long, Z.; Lin, D. Toxicity of oxide nanoparticles to the green algae Chlorella sp. Chem. Eng. J. 2011, 170, 525–530. [Google Scholar] [CrossRef]
- Suman, T.Y.; Rajasree, S.R.R.; Kirubagaran, R. Evaluation of zinc oxide nanoparticles toxicity on marine algae Chlorella vulgaris through flow cytometric, cytotoxicity and oxidative stress analysis. Ecotoxicol. Environ. Saf. 2015, 113, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, J.; Tan, L.; Chen, X. Toxic effects of nano-ZnO on marine microalgae Skeletonema costatum: Attention to the accumulation of intracellular Zn. Aquat. Toxicol. 2016, 178, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Aravantinou, A.F.; Tsarpali, V.; Dailianis, S.; Manariotis, I.D. Effect of cultivation media on the toxicity of ZnO nanoparticles to freshwater and marine microalgae. Ecotoxicol. Environ. Saf. 2015, 114, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Wang, Y. Toxic effects of different particle size ZnO NPs on marine microalgae Chlorella sp. IOP Conf. Ser. Earth Environ. Sci. 2021, 770, 012022. [Google Scholar] [CrossRef]
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of Nanoparticle Toxicity on Their Physical and Chemical Properties. Nanoscale Res. Lett. 2018, 13, 44. [Google Scholar] [CrossRef]
- Dobretsov, S.; Sathe, P.; Bora, T.; Barry, M.; Myint, M.T.Z.; Al Abri, M. Toxicity of Different Zinc Oxide Nanomaterials at 3 Trophic Levels: Implications for Development of Low-Toxicity Antifouling Agents. Environ. Toxicol. Chem. 2020, 39, 1343–1354. [Google Scholar] [CrossRef]
- Du, X.; Zhou, W.; Zhang, W.; Sun, S.; Han, Y.; Tang, Y.; Shi, W.; Liu, G. Toxicities of three metal oxide nanoparticles to a marine microalga: Impacts on the motility and potential affecting mechanisms. Environ. Pollut. 2021, 290, 118027. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Root Uptake and Phytotoxicity of ZnO Nanoparticles. Environ. Sci. Technol. 2008, 42, 5580–5585. [Google Scholar] [CrossRef]
- Chevion, M.A. site-specific mechanism for free radical induced biological damage: The essential role of redox-active transition metals. Free Radic. Biol. Med. 1988, 5, 27–37. [Google Scholar] [CrossRef]
- Guo, D.; Bi, H.; Liu, B.; Wu, Q.; Wang, D.; Cui, Y. Reactive oxygen species induced cytotoxic effects of zinc oxide nanoparticles in rat retinal ganglion cells. Toxicol. Vitro. 2013, 27, 731–738. [Google Scholar] [CrossRef]
- Castro-Bugallo, A.; González-Fernández, Á.; Guisande, C.; Barreiro, A. Comparative Responses to Metal Oxide Nanoparticles in Marine Phytoplankton. Arch. Environ. Contam. Toxicol. 2014, 67, 483–493. [Google Scholar] [CrossRef]
- Hazeem, L.J.; Bououdina, M.; Rashdan, S.; Brunet, L.; Slomianny, C.; Boukherroub, R. Cumulative effect of zinc oxide and titanium oxide nanoparticles on growth and chlorophyll a content of Picochlorum sp. Environ. Sci. Pollut. Res. 2016, 23, 2821–2830. [Google Scholar] [CrossRef]
- Broccoli, A.; Anselmi, S.; Cavallo, A.; Ferrari, V.; Prevedelli, D.; Pastorino, P.; Renzi, M. Ecotoxicological effects of new generation pollutants (nanoparticles, amoxicillin and white musk) on freshwater and marine phytoplankton species. Chemosphere 2021, 279, 130623. [Google Scholar] [CrossRef]
- Schiavo, S.; Oliviero, M.; Philippe, A.; Sonia Manzo, S. Nanoparticles based sunscreens provoke adverse effects on marine microalgae Dunaliella tertiolecta. Environ. Sci. Nano 2018, 5, 3011–3022. [Google Scholar] [CrossRef]
- Gunasekarana, D.; Chandrasekarana, N.; Jenkinsb, D.; Mukherjeea. Plain polystyrene microplastics reduce the toxic effects of ZnO particles on marine microalgae Dunaliella salina. J. Environ. Chem. Eng. 2020, 8, 104250. [Google Scholar] [CrossRef]
- Baek, S.; Joo, S.H.; Su, C.; Toborek, M. Toxicity of ZnO/TiO2-conjugated carbon-based nanohybrids on the coastal marine alga Thalassiosira pseudonana. Environ. Toxicol. 2020, 35, 87–96. [Google Scholar] [CrossRef]
- Li, J.; Mao, S.; Ye, Y.; Lü, J.; Jing, F.; Guo, Y.; Liu, H.; Wang, P.; Ma, W.; Qi, P.; et al. Micro-polyethylene particles reduce the toxicity of nano zinc oxide in marine microalgae by adsorption. Environ. Pollut. 2021, 290, 118042. [Google Scholar] [CrossRef]
- Zhu, X.; Tan, L.; Zhao, T.; Huang, W.; Guo, X.; Wang, J.; Wang, J. Alone and combined toxicity of ZnO nanoparticles and graphene quantum dots on microalgae Gymnodinium. Environ. Sci. Pollut. Res. 2022, 29, 47310–47322. [Google Scholar] [CrossRef]
Figure 1.
Description of the fate and behavior of ZnO-NPs in the marine environment.
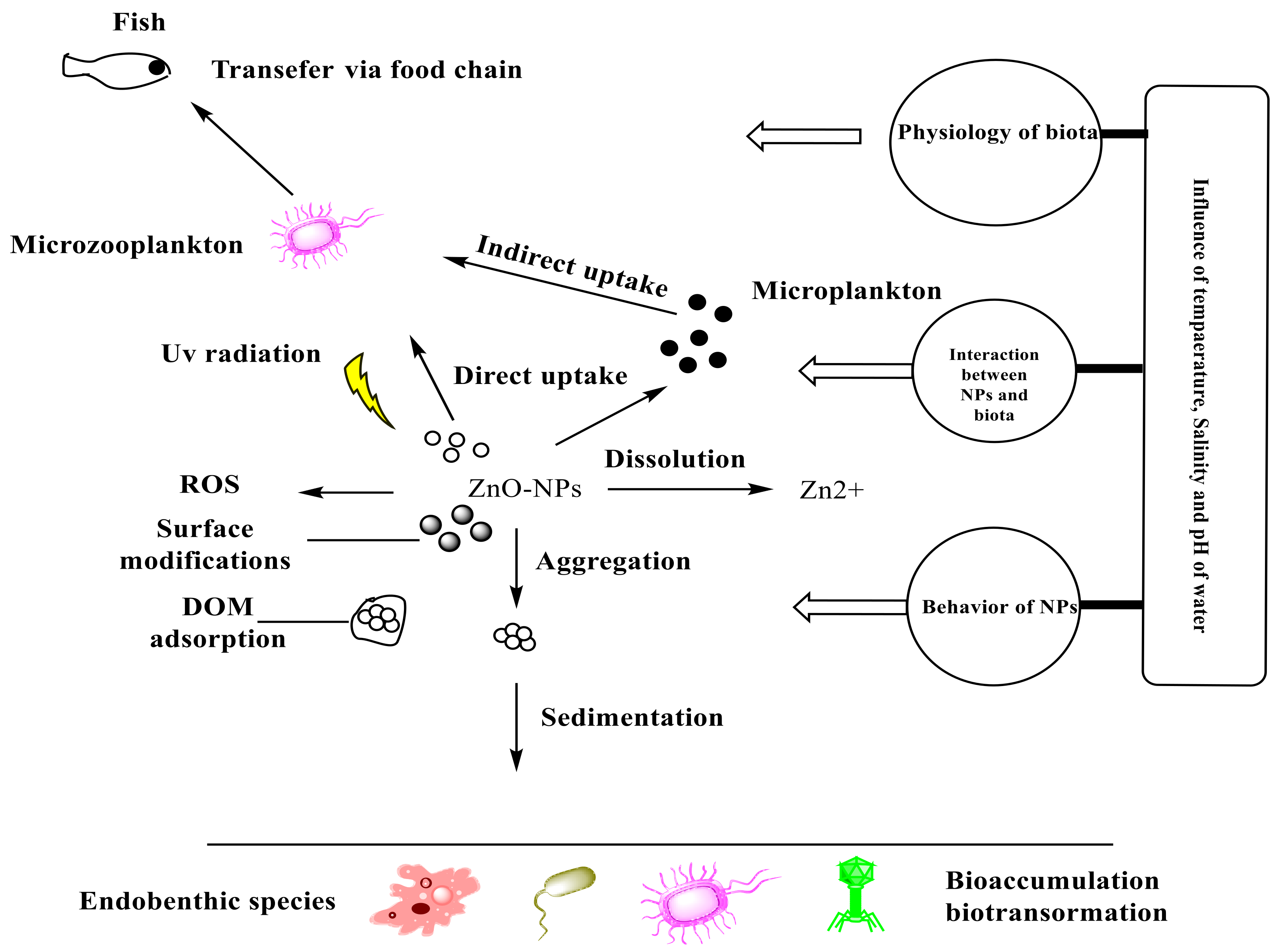
Figure 2.
Effects of humic acid and pH on the release of Zn2+ in the marine system.
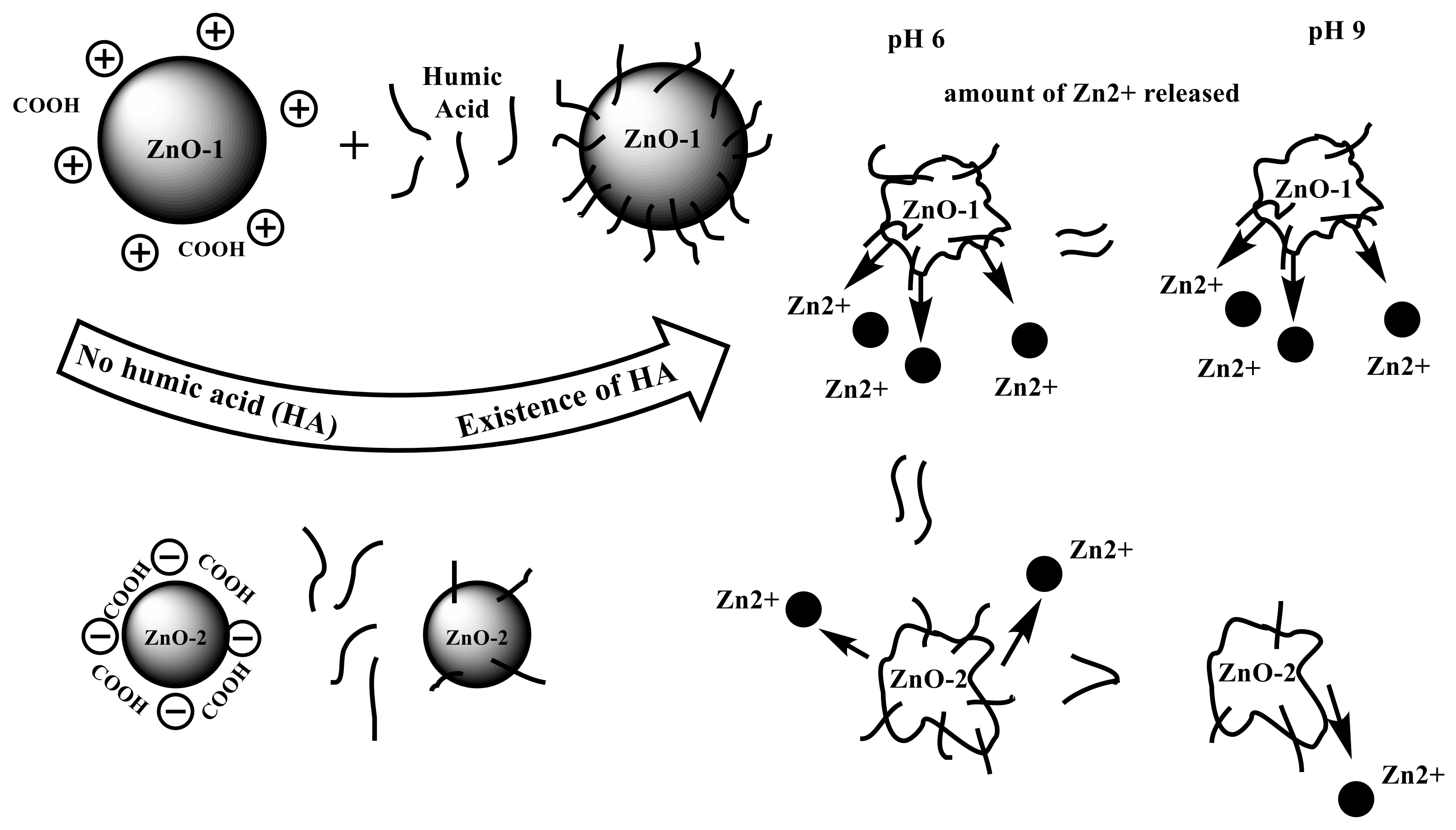
Figure 3.
Schematic illustration of potential pathways of intracellular NP uptake in the algal cell. NPs can accumulate in a gap between the cell wall and the cell membrane in algal cells. Metal ions enter the cells through ion channels and carrier proteins after NPs are dispersed in the external media. The transfer of NPs is then aided by transport proteins that move NPs from one location to another [15].
Figure 3.
Schematic illustration of potential pathways of intracellular NP uptake in the algal cell. NPs can accumulate in a gap between the cell wall and the cell membrane in algal cells. Metal ions enter the cells through ion channels and carrier proteins after NPs are dispersed in the external media. The transfer of NPs is then aided by transport proteins that move NPs from one location to another [15].
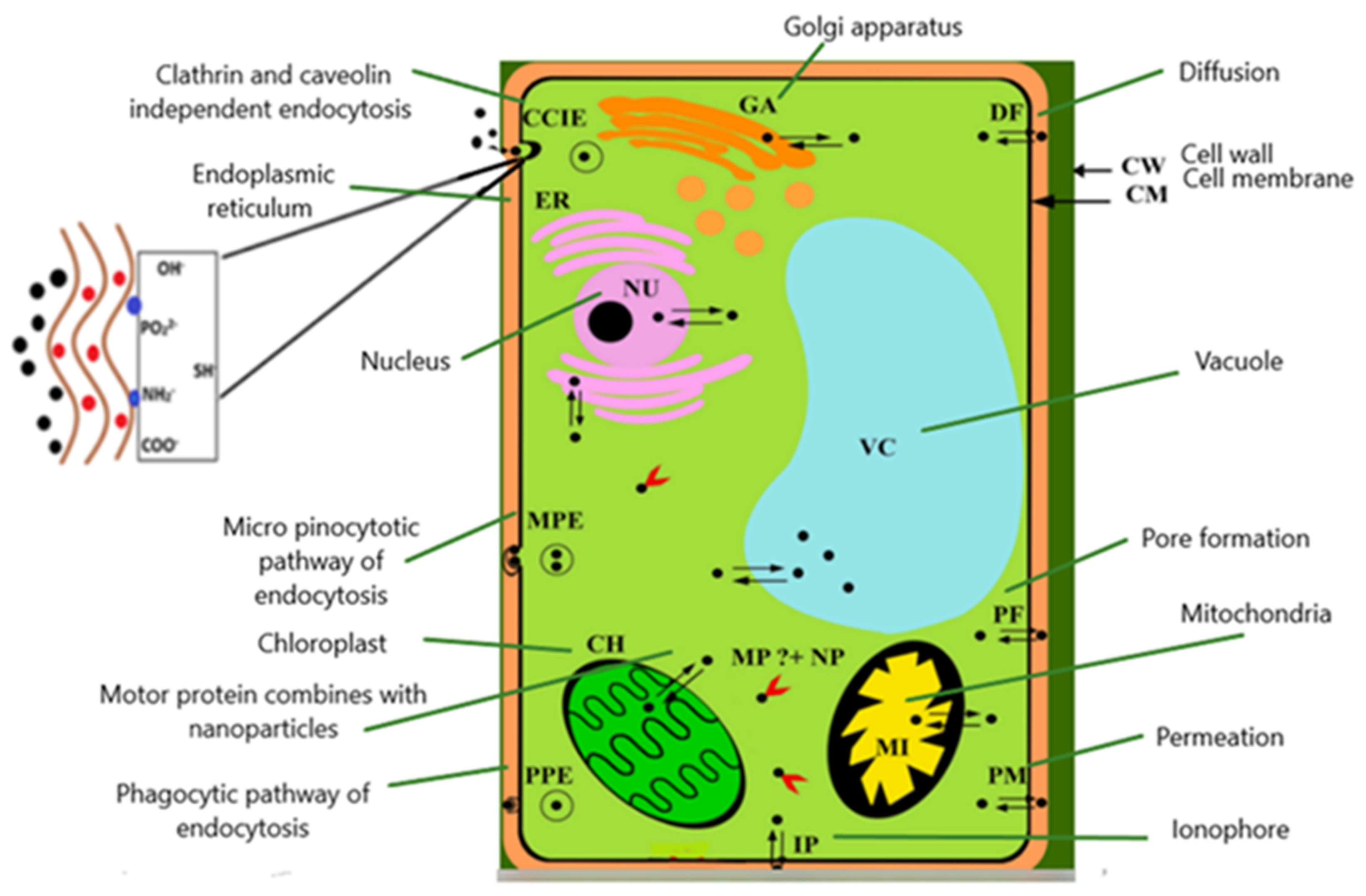
Figure 4.
The toxicity of ZnO-NPs on algae [66].
Figure 4.
The toxicity of ZnO-NPs on algae [66].
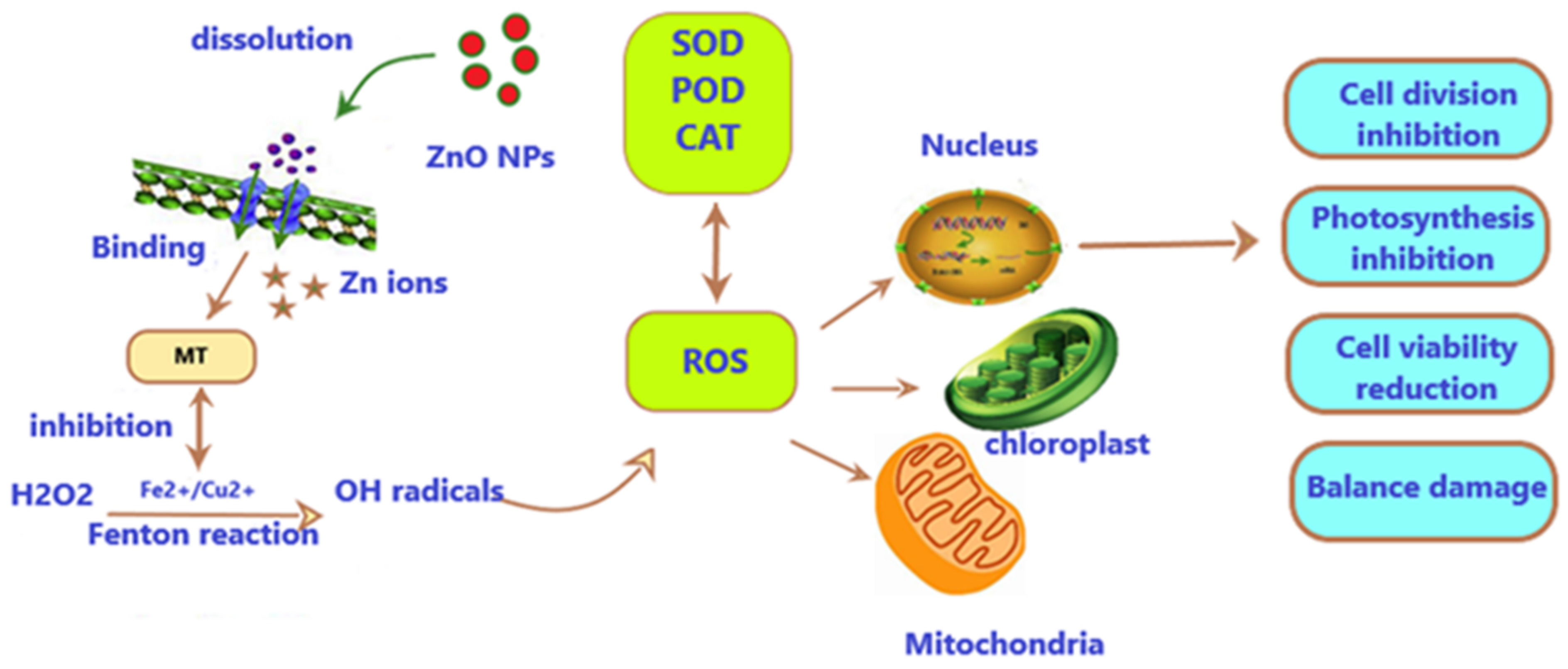
Figure 5.
Releasing mechanism of ZnO-NP. Release of Zn2+ ions (in the exposure medium, whether free or complexed with organic or inorganic ligands).
Figure 5.
Releasing mechanism of ZnO-NP. Release of Zn2+ ions (in the exposure medium, whether free or complexed with organic or inorganic ligands).

Figure 6.
The role of metal-based NPs in oxidative stress.
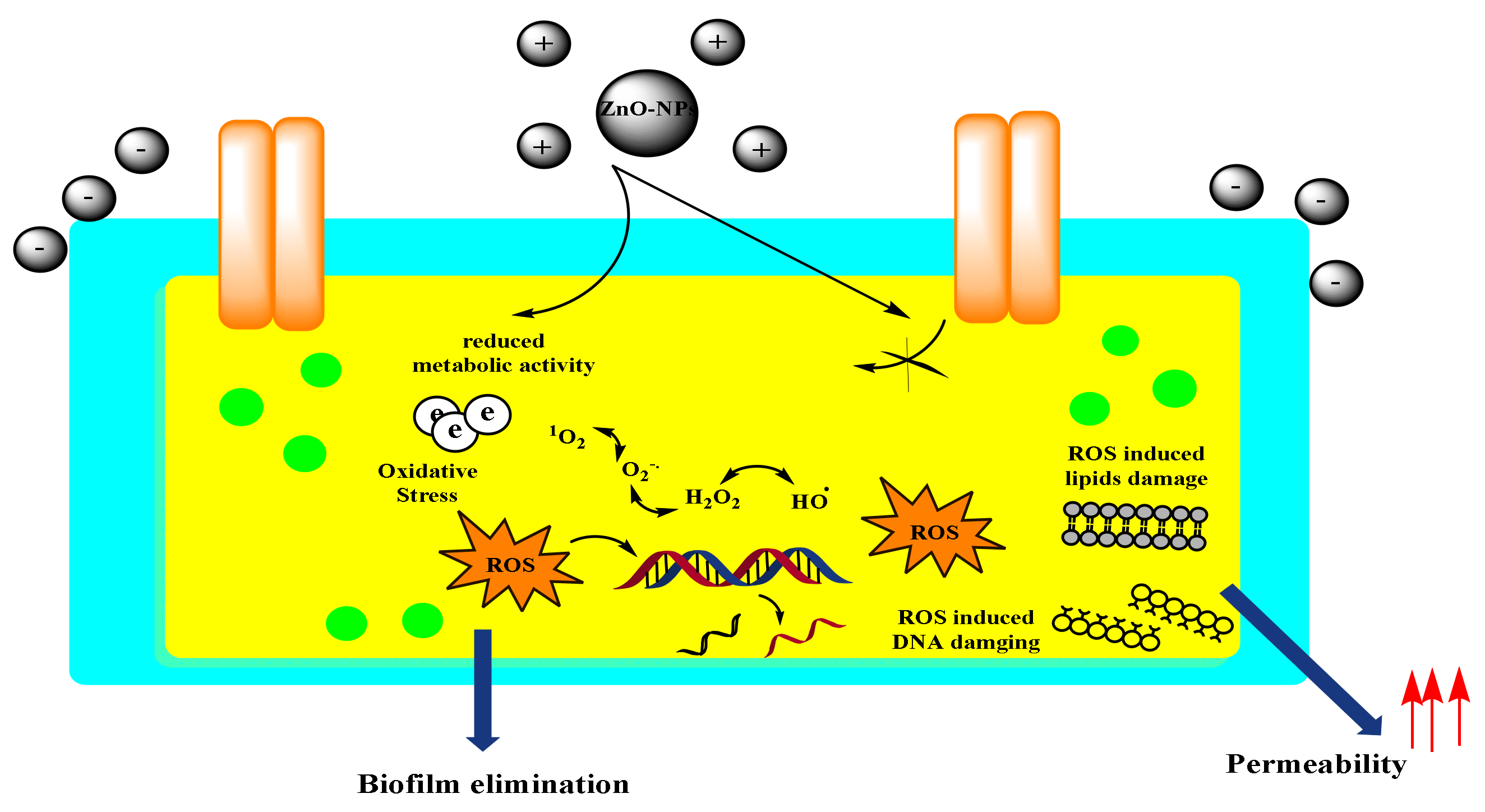
Figure 7.
The number of published articles in the period between 2010 and 2022.
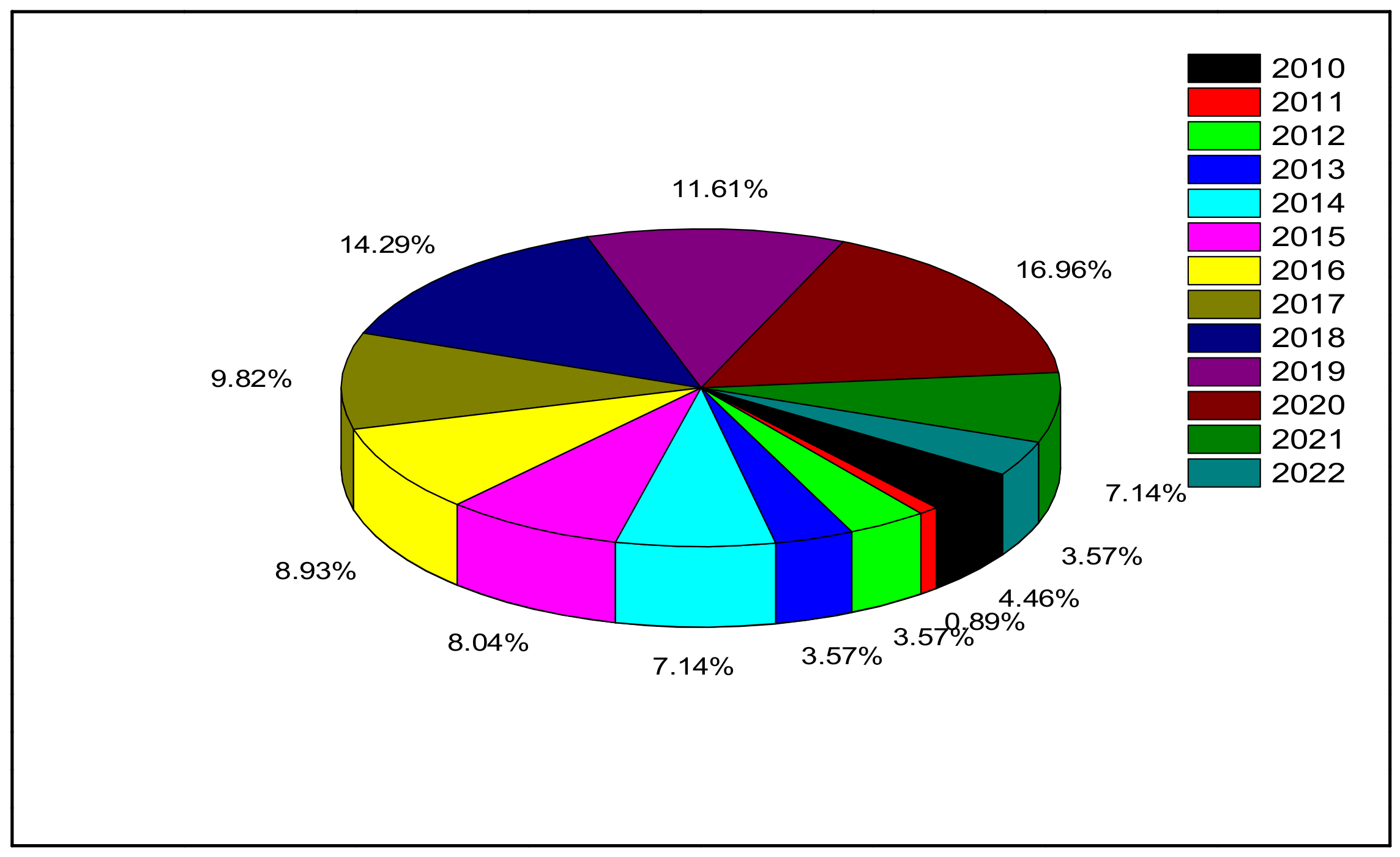
Table 1.
Modeled ZnO-NP concentrations in various environmental compartments in different countries presented as mode (most frequent value) and as a range of the lower and upper quantiles (Q0.15 and Q0.85) a.
Table 1.
Modeled ZnO-NP concentrations in various environmental compartments in different countries presented as mode (most frequent value) and as a range of the lower and upper quantiles (Q0.15 and Q0.85) a.
| Compartment | Predicted Environmental Concentration Mode (Q0.15, Q0.85) (µg/L, µg/m3 or µg/Kg) | ||
|---|---|---|---|
| Europe | U.S. | Switzerland | |
| Surface water | 0.010 (0.008, 0.055) | 0.001 (0.001, 0.003) | 0.013 (0.011, 0.058) |
| STP effluent | 0.432 (0.340, 1.42) | 0.3 (0.22, 0.74) | 0.441 (0.343, 1.32) |
| STP sludge | 17.1 (13.6, 57.0) | 23.2 (17.4, 57.7) | 21.4 (16.8, 64.7) |
| Sediment | 2.9 (2.65, 51.7) | 0.51 (0.49, 8.36) | 3.33 (3.30, 56.0) |
| Soil | 0.093 (0.085, 0.661) | 0.050 (0.041, 0.274) | 0.032 (0.026, 0.127) |
| Sludge treated soil | 3.25 (2.98, 23.1) | 1.99 (1.62, 10.9) | na |
| Air | <0.0005 | <0.0005 | <0.0005 |
Table 3.
Combined toxicity effects of ZnO-NPs with different materials of marine microalgae.
| Tested Materials | NP Size and Characteristics | Concentration | Microalgae Species | Growth Conditions | Toxic Effect | References |
|---|---|---|---|---|---|---|
| ZnO and TiO2 | “ZnO (50 nm) TiO2 (21 nm; Anatase and rutile phases)” | “10 mg/L” | “Picochlorumsp.” | “5 weeks incubation experiment” | “No significant difference between control and treated cells” | [98] |
| ZnO and TiO2 | - | - | “Dunaliella tertiolecta” | - | “ROS and DNA damage was evidenced in algae cells exposed to sunscreen containing both ZnO and TiO2” | [100] |
| ZnO and polystyrene (PS) microplastic | “ZnO < 50 nm ZnO-Bulk > 100 nm” | “ZnO ((1.22, 12.28 and 122.88 μM) Microplastic (1 mg/L)” | “Dunaliella salina” | “72 h” | “The harmful effects of bulk and nano-sized ZnO particles were considerably reduced in the presence of plain PS microplastics” | [101] |
| ZnO-GO ZnO-CNTs | “GO (1–5 μm diameter, and 0.8–1.2 nm thickness)” “CNT (6–13 nm [O.D.] × 2.5−20 μm [L]” | “50 and 100 mg/L” | “Thalassiosira pseudonana” | “72 h” | “ZnO-GO was the most inhibitory material. The primary mechanism identified was the generation of ROS” | [102] |
| ZnO-NPs Polyethylene Microplastic (PEMP) | “ZnO (20–30 nm)” “150 µm” | “ZnO (2.7 mg/L)” “PEMP (1g/L)” | “Tetraselmis helgolandica var. tsingtaoensis” | “96 h” | “The inhibitory effect of ZnO reduced by 14.4% in the presence of PEMP” | [103] |
| ZnO-NPs Graphene Quantum Dots (GQDs) | “1–20 mg/L” | “Gymnodinium sp. | “The inhibitory effect was decreased compared to the single GQDs and showed antagonistic effect” | [104] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hazeem, L. Single and Combined Toxicity Effects of Zinc Oxide Nanoparticles: Uptake and Accumulation in Marine Microalgae, Toxicity Mechanisms, and Their Fate in the Marine Environment. Water 2022, 14, 2669. https://doi.org/10.3390/w14172669
AMA Style
Hazeem L. Single and Combined Toxicity Effects of Zinc Oxide Nanoparticles: Uptake and Accumulation in Marine Microalgae, Toxicity Mechanisms, and Their Fate in the Marine Environment. Water. 2022; 14(17):2669. https://doi.org/10.3390/w14172669
Chicago/Turabian StyleHazeem, Layla. 2022. "Single and Combined Toxicity Effects of Zinc Oxide Nanoparticles: Uptake and Accumulation in Marine Microalgae, Toxicity Mechanisms, and Their Fate in the Marine Environment" Water 14, no. 17: 2669. https://doi.org/10.3390/w14172669
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.