
Colonization Dynamics of Potential Stowaways Inhabiting Marinas: Lessons from Caprellid Crustaceans


Abstract
:1. Introduction
2. Materials and Methods
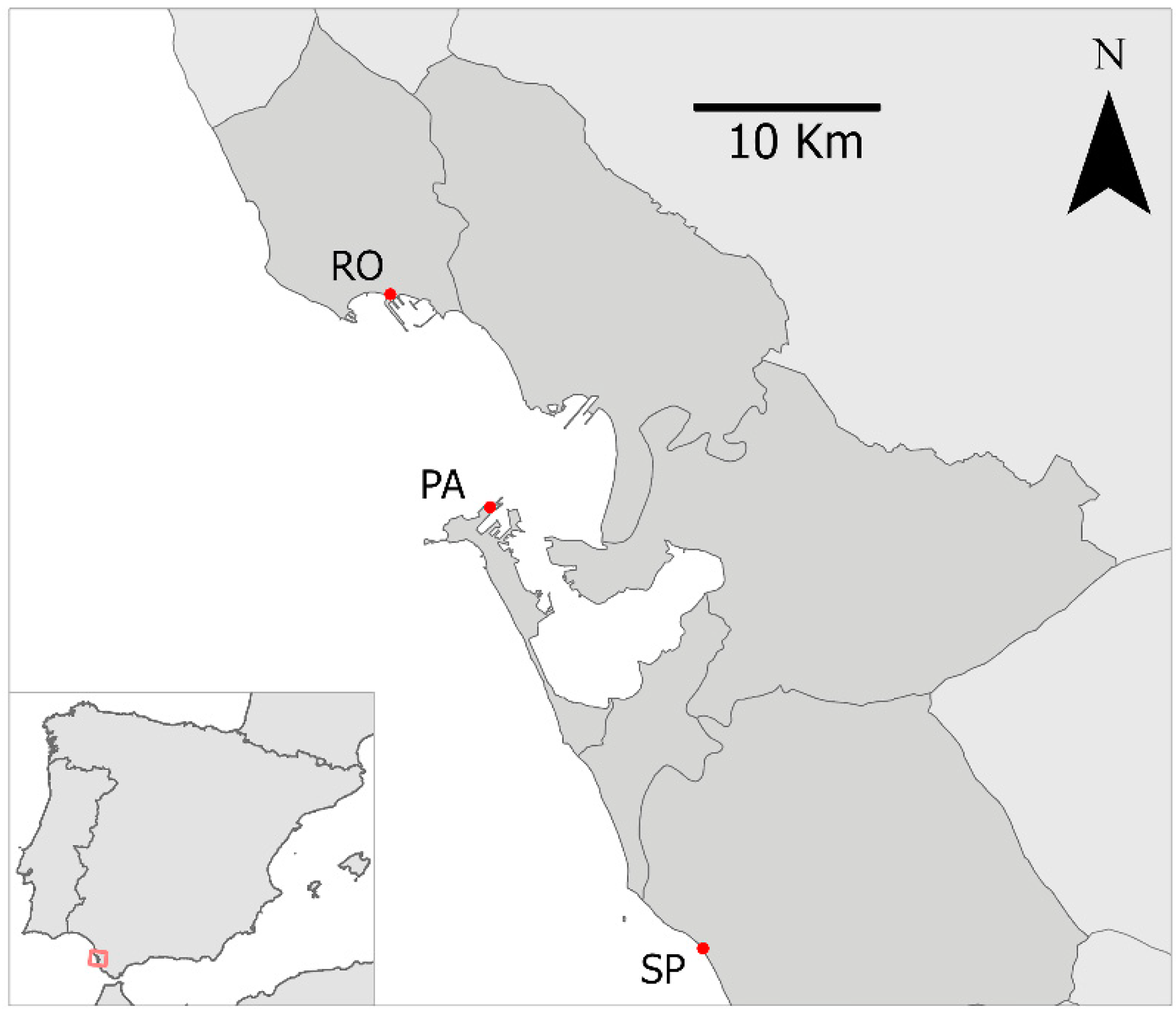
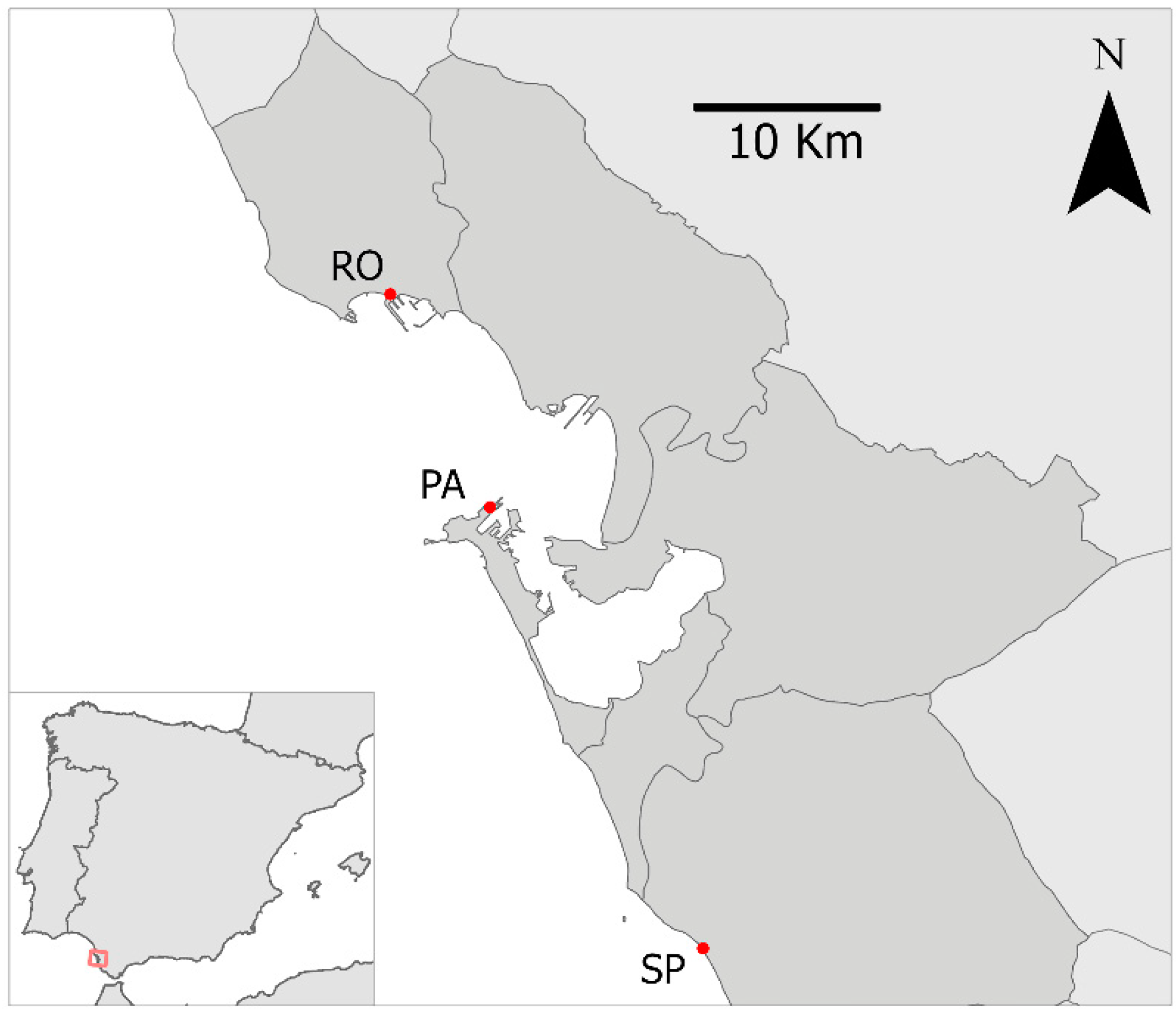
2.1. Study Area
2.2. Experimental Design and Field Procedure
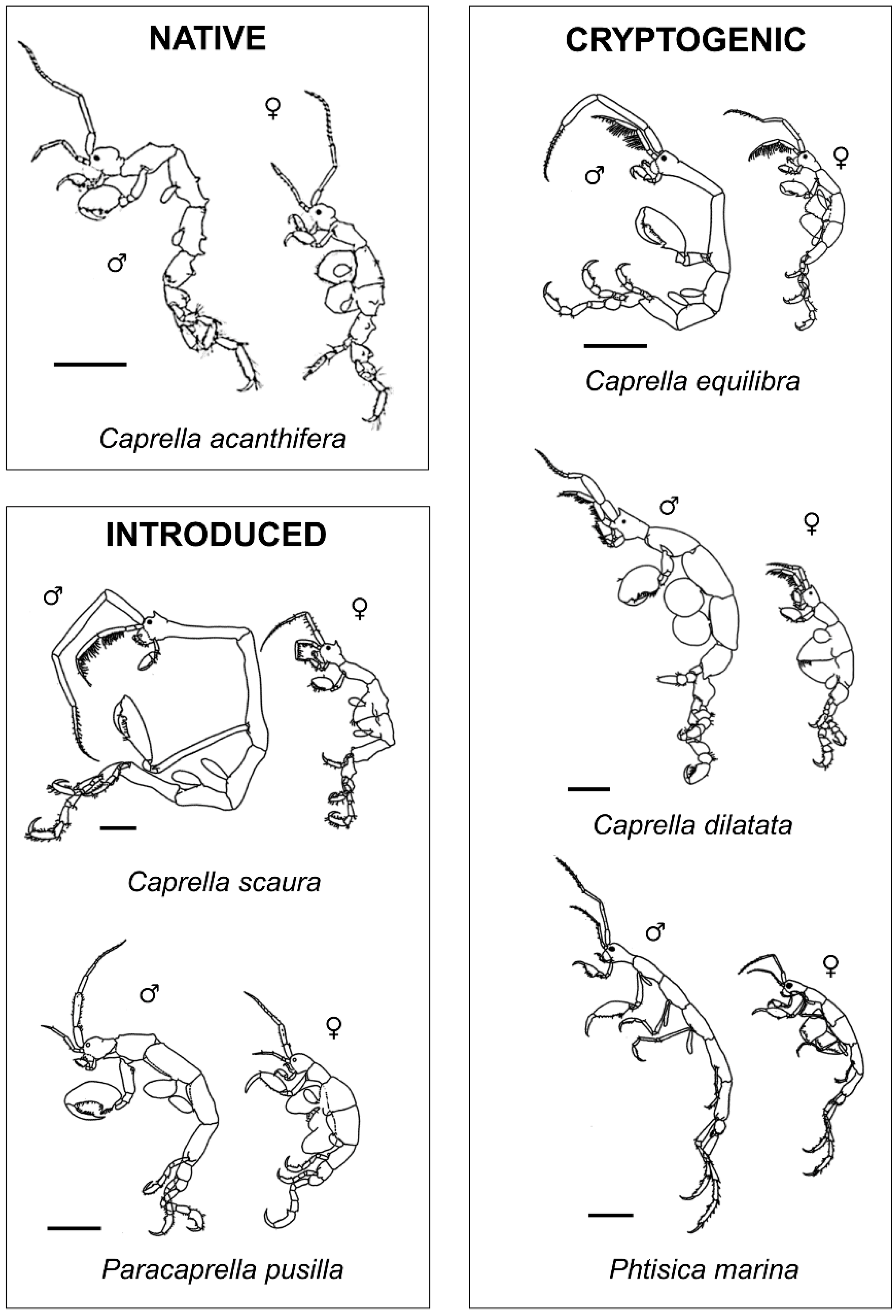
2.3. Laboratory Processing and Classification of Collected Individuals
2.4. Statistical Analyses
3. Results
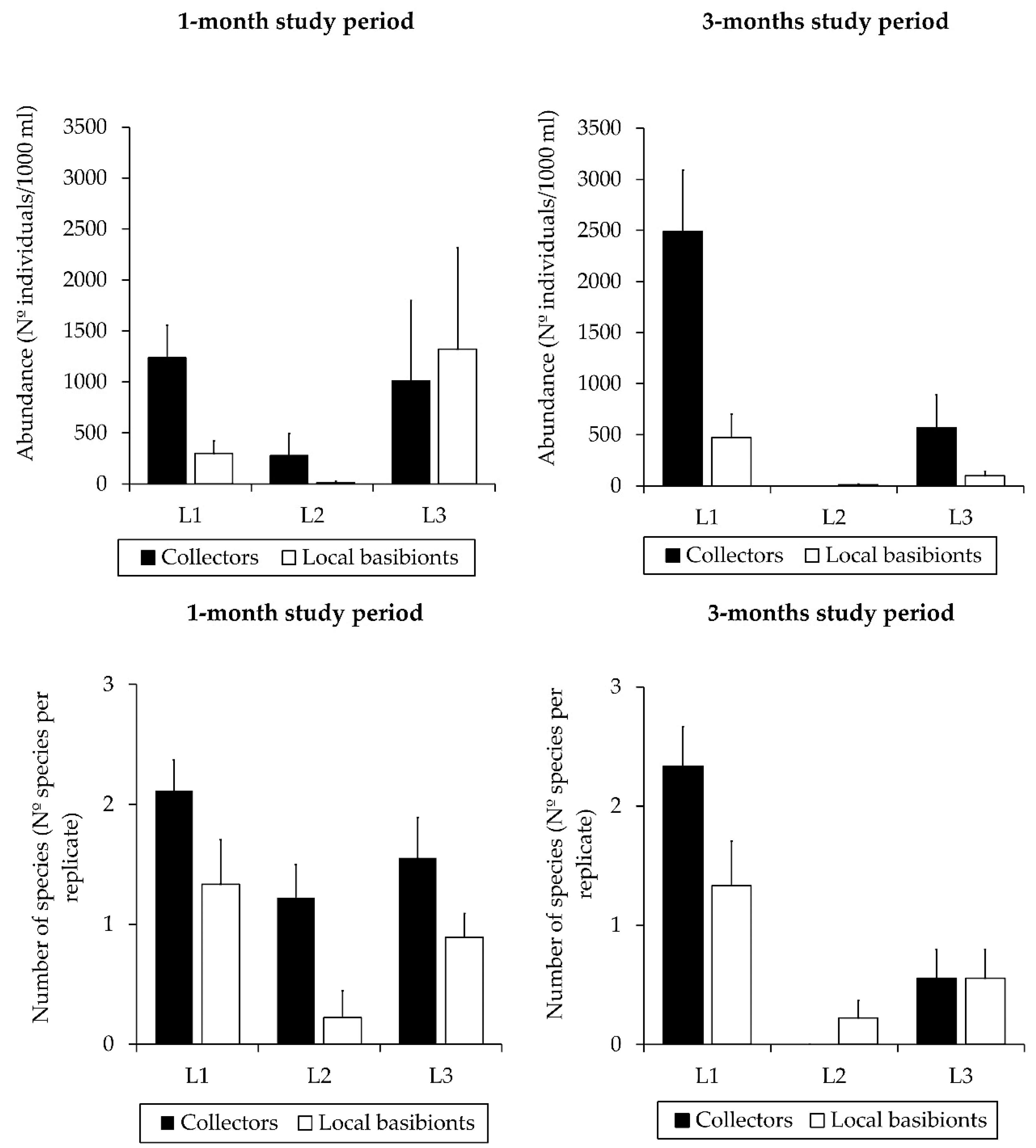
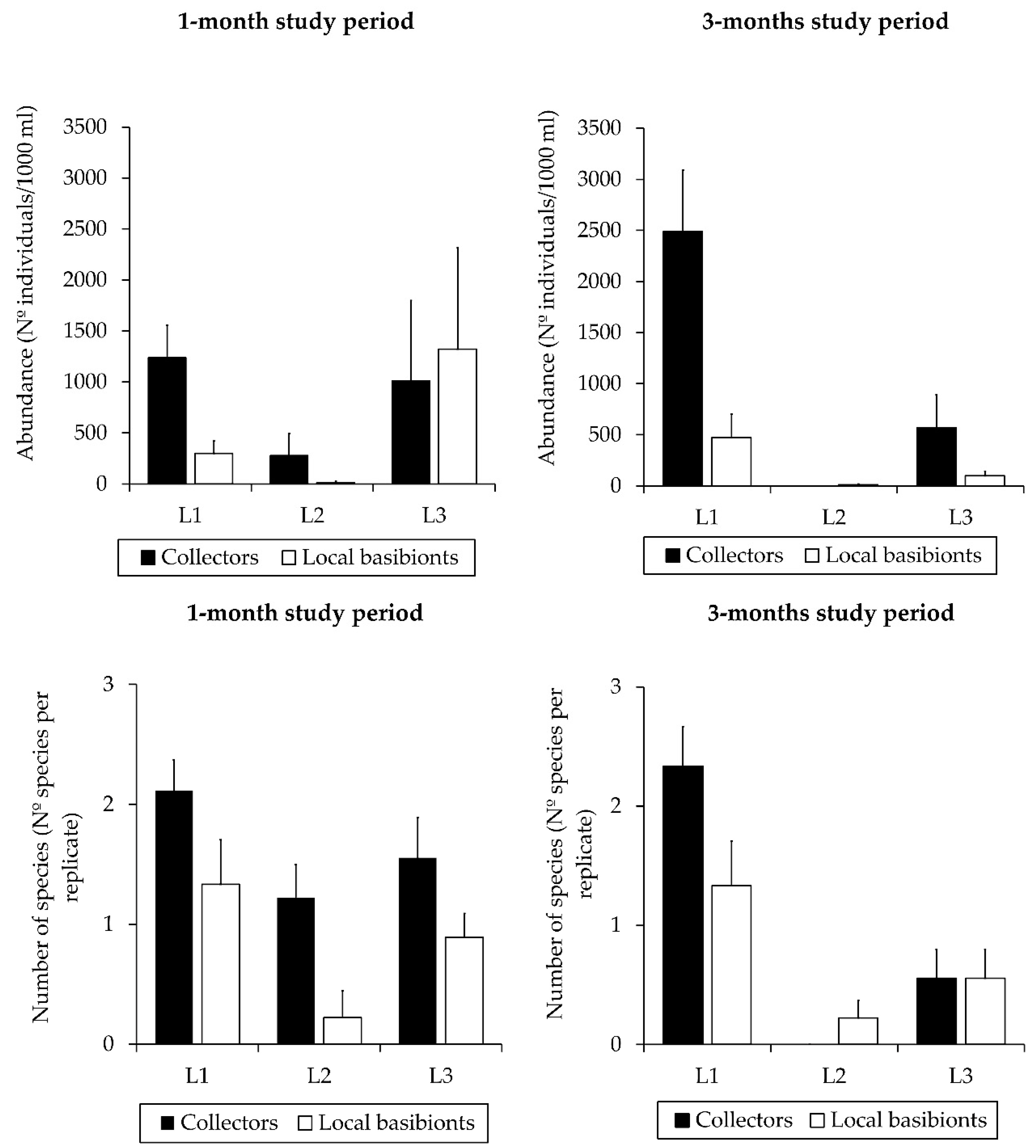
3.1. Influence of Factors on Caprellid Community Assemblages
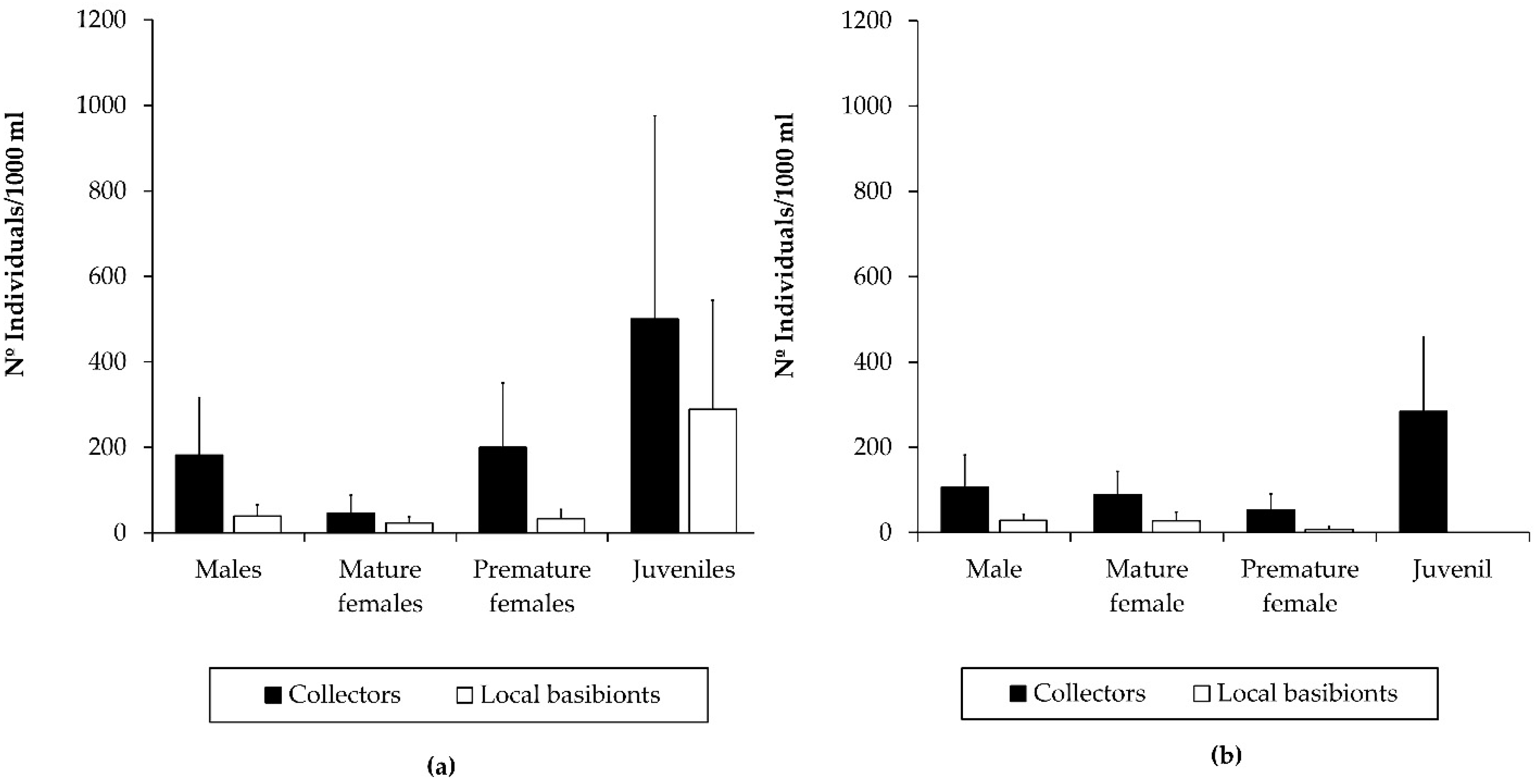
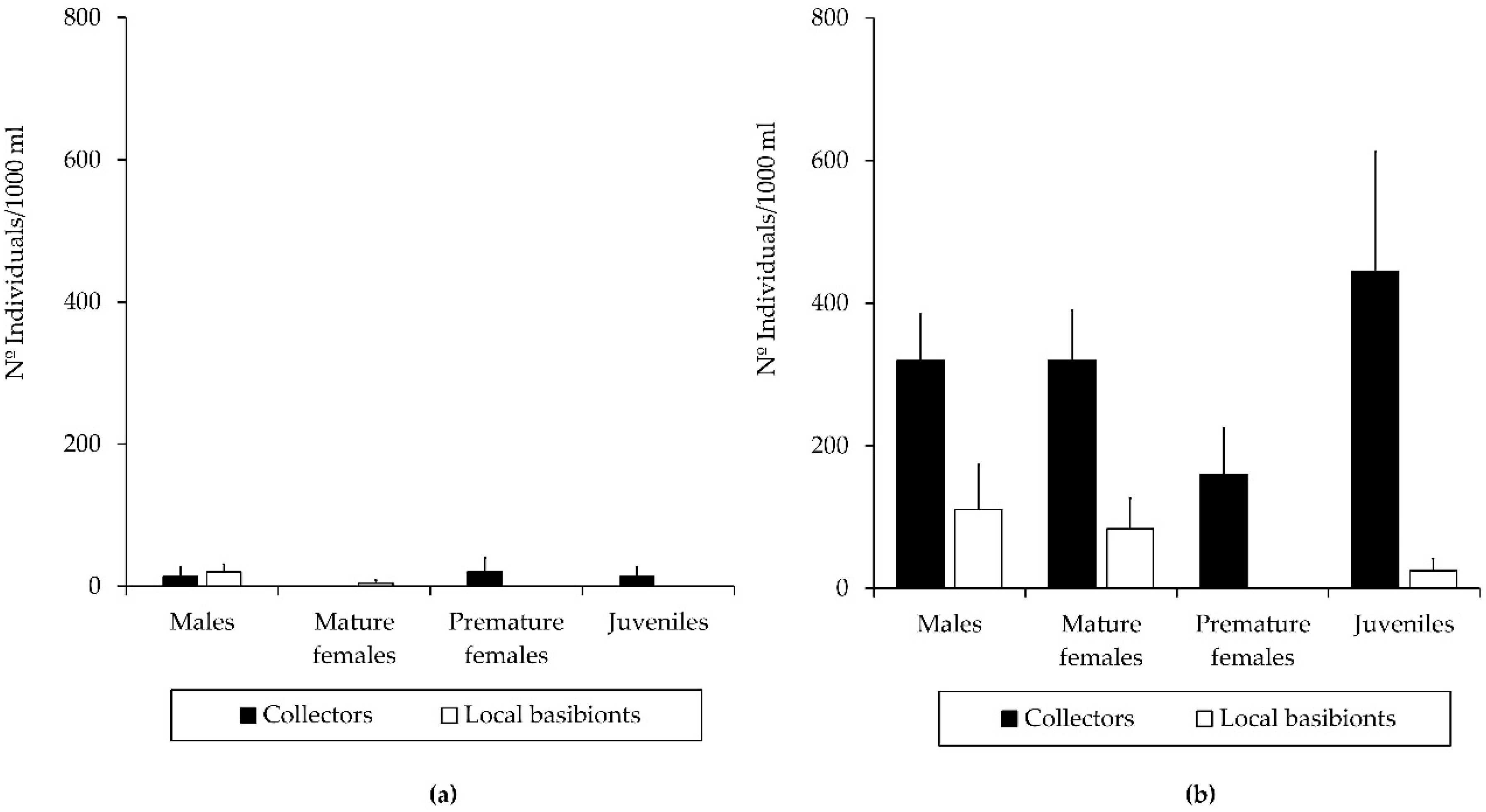
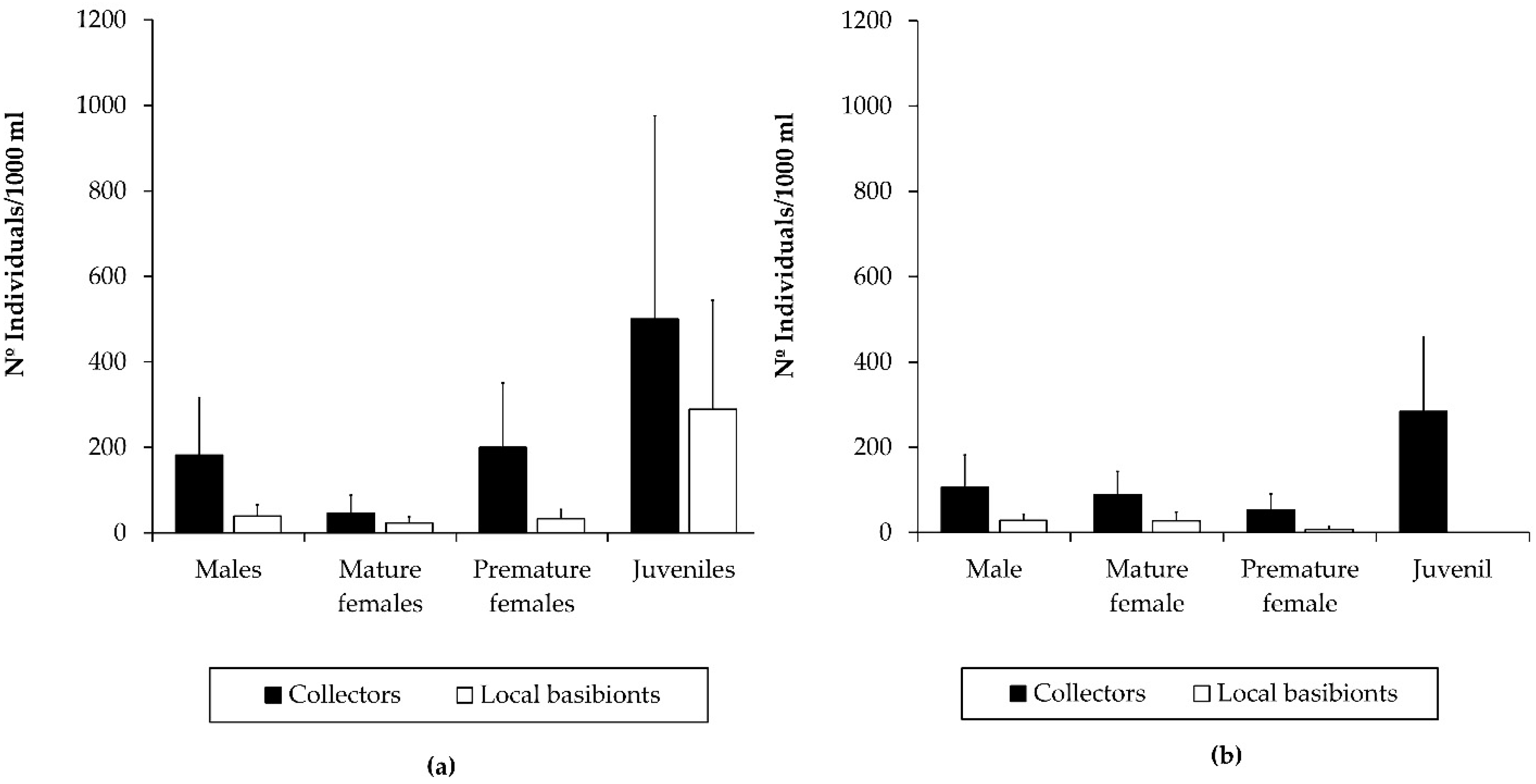
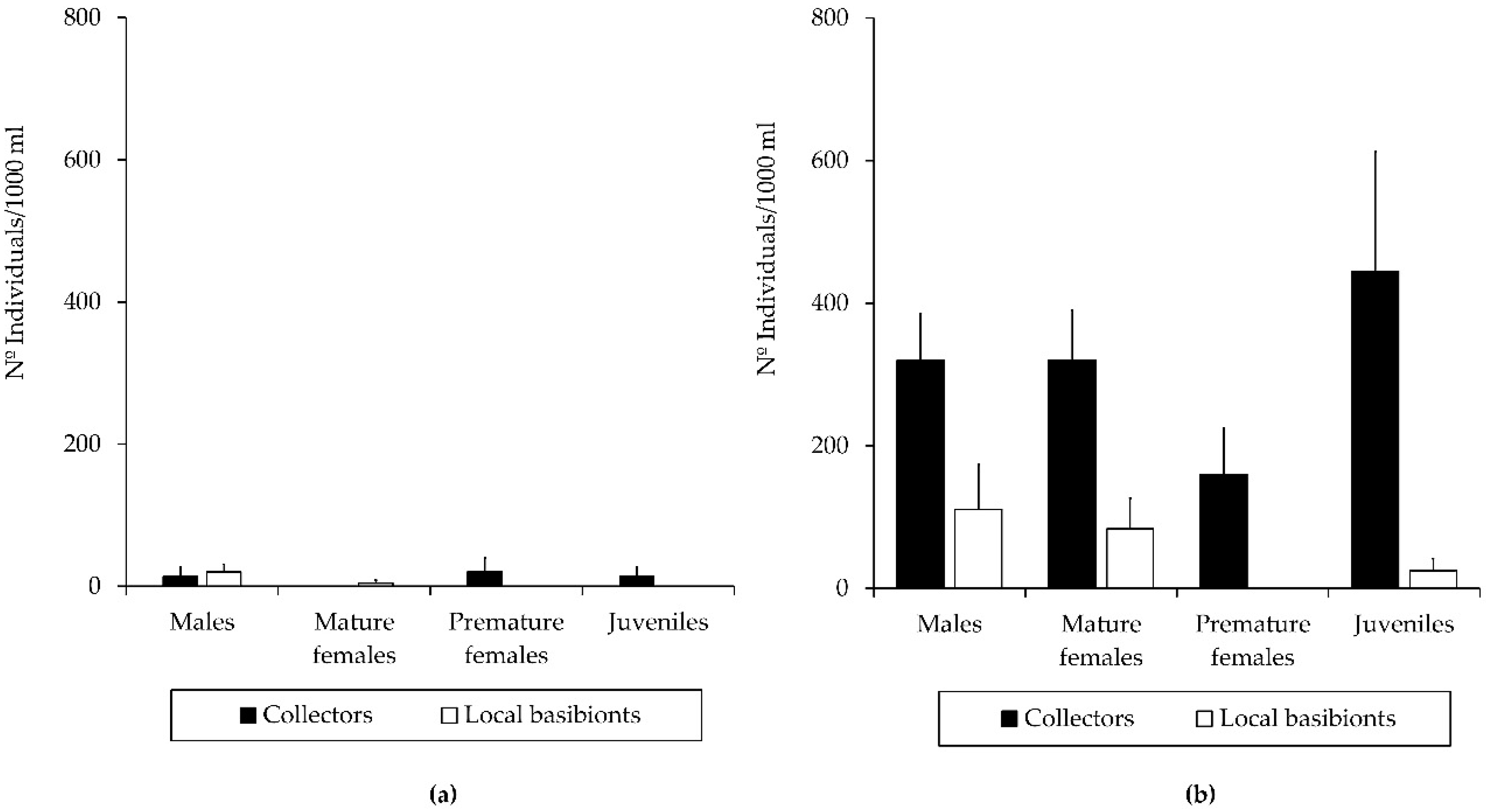
3.2. Population Structure of the Introduced Caprellid Species
4. Discussion
4.1. Factors Affecting the Colonization Patterns of Caprellid Community Assemblages
4.2. Colonization Abilities of Potential Stowaways Inhabiting Marinas
4.3. The Colonization Dynamics of the Introduced Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulme, P.E.; Bacher, S.; Kenis, M.; Klotz, S.; Kühn, I.; Minchin, D.; Nentwig, W.; Olenin, S.; Panov, J.; Pergl, J.; et al. Grasping at the routes of biological invasions: A framework for integrating pathways into policy. J. App. Ecol. 2008, 45, 403–414. [Google Scholar] [CrossRef]
- Banks, N.C.; Paini, D.R.; Bayliss, K.L.; Hodda, M. The role of global trade and transport network topology in the human-mediated dispersal of alien species. Ecol. Lett. 2015, 18, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Tingley, R.; García-Díaz, P.; Arantes, C.R.R.; Cassey, P. Integrating transport pressure data and species distribution models to estimate invasion risk for alien stowaways. Ecography 2018, 41, 635–646. [Google Scholar] [CrossRef]
- Fofonoff, P.W.; Ruiz, G.M.; Hines, A.H.; Steves, B.D.; Carlton, J.T. Four centuries of biological invasions in tidal waters of the Chesapeake Bay region. In Biological Invasions in Marine Ecosystems; Rilov, G., Crooks, A., Eds.; Springer: Heidelberg, Germany, 2009; pp. 479–506. ISBN 978-3-5407-9235-2. [Google Scholar]
- Carlton, J.T. Bioinvasion ecology: Assessing invasion impact and scale. In Invasive Aquatic Species of Europe. Distribution, impacts, and management; Leppäkoski, E., Gollasch, S., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 7–19. ISBN 978-90-481-6111-9. [Google Scholar]
- Floerl, O.; Inglis, G.J. Starting the invasion pathway: The interaction between source populations and human transport vectors. Biol. Invasions. 2005, 7, 589–606. [Google Scholar] [CrossRef]
- Carlton, J.T. Man’s role in changing the face of the ocean: Biological invasions and implications for conservation of near-shore environments. Conserv. Biol. 1989, 3, 265–273. [Google Scholar] [CrossRef]
- Williamson, M. Biological Invasions; Springer: Dordrecht, The Netherlands, 1996; ISBN 978-0-412-59190-7. [Google Scholar]
- Ruiz, G.M.; Carlton, J.T.; Grosholz, E.D.; Hines, A.H. Global invasions of marine and estuarine habitats by non-indigenous species: Mechanisms, extent, and consequences. Am. Zool. 1997, 37, 621–632. [Google Scholar] [CrossRef]
- Puth, L.M.; Post, D.M. Studying invasion: Have we missed the boat? Ecol. Lett. 2005, 8, 715–721. [Google Scholar] [CrossRef]
- Floerl, O.; Inglis, G.J.; Dey, K.; Smith, A. The importance of transport hubs in stepping-stone invasions. J. App. Ecol. 2009, 46, 7–45. [Google Scholar] [CrossRef]
- Minchin, D.; Floerl, O.; Savini, D.; Occhipinti-Ambrogi, A. Small craft and the spread of exotic species. In The Ecology of Transportation: Managing Mobility for the Environment; Davenport, J., Davenport, J.L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 99–118. ISBN 978-1-4020-4504-2. [Google Scholar]
- Murray, C.C.; Pakhomov, E.A.; Therriault, T.W. Recreational boating: A large unregulated vector transporting marine invasive species. Divers. Distrib. 2011, 17, 1161–1172. [Google Scholar] [CrossRef]
- Ros, M.; Navarro-Barranco, C.; González-Sánchez, M.; Ostalé-Valriberas, E.; Cervera-Currado, L.; Guerra-García, J.M. Starting the stowaway pathway: The role of dispersal behaviour in the invasion success of low-mobile marine species. Biol. Invasions. 2020, 22, 2797–2812. [Google Scholar] [CrossRef]
- Chapple, D.G.; Simmonds, S.M.; Wong, B.B. Can behavioral and personality traits influence the success of unintentional species introductions? Trends. Ecol. Evol. 2012, 27, 57–64. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; ISBN 978-0-6910-8836-5. [Google Scholar]
- Ebenhard, T. Colonization in metapopulations: A review of theory and observations. Biol. J. Linn. Soc. 1991, 42, 105–121. [Google Scholar] [CrossRef]
- Cáceres, C.E.; Soluk, D.A. Blowing in the wind: A field test of overland dispersal and colonization by aquatic invertebrates. Oecologia 2002, 131, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Buschbaum, C.; Gutow, L. Mass occurrence of an introduced crustacean (Caprella mutica) in south-eastern North Sea. Helgol. Mar. Res. 2005, 59, 252–253. [Google Scholar] [CrossRef]
- Ros, M. Invasion Dynamic of Introduced Caprellids (Crustacea: Amphipoda) in the Mediterranean Sea and Adjacent Waters. Global Distribution, Ecology, and Vectors of Spread. Ph.D. Dissertation, University of Seville, Seville, Spain, 2015. [Google Scholar]
- Cabezas, M.P.; Navarro-Barranco, C.; Ros, M.; Guerra-García, J.M. Long-distance dispersal, low connectivity, and molecular evidence of a new cryptic species in the obligate rafter Caprella andreae Mayer, 1890 (Crustacea: Amphipoda: Caprellidae). Helgol. Mar. Res. 2013, 67, 483–497. [Google Scholar] [CrossRef]
- Ros, M.; Vázquez-Luis, M.; Guerra-Garcia, J.M. Environmental factors modulating the extent of impact in coastal invasions: The case of a widespread invasive caprellid (Crustacea: Amphipoda) in the Iberian Peninsula. Mar. Poll. Bull. 2015, 98, 247–258. [Google Scholar] [CrossRef]
- Drake, J.M.; Lodge, D.M. Global hot spots of biological invasions: Evaluating options for ballast–water management. Proc. R. Soc. Lond. B. Biol. Sci. 2004, 271, 575–580. [Google Scholar] [CrossRef]
- Ros, M.; López De La Rosa, V.L.; Revanales, T.; Spinelli, A.; Fernández-Romero, A.; Cervera-Currado, L.; Guerra-García, J.M. Towards a standardized methodology for monitoring and assessing marine mobile epibenthic communities across spatio-temporal scales. In Proceedings of the XX Iberian Symposium on Marine Biology Studies (SIEBM XX), Braga, Portugal, 9–12 September 2019. [Google Scholar] [CrossRef]
- Scribano, G.; Marchini, A.; Ros, M. Population dynamics and life history traits of the non-indigenous Paranthura japonica Richardson (1909) in a recently invaded Mediterranean site. Reg. Stud. Mar. Sci. 2021, 46, 101883. [Google Scholar] [CrossRef]
- Dafforn, K.A.; Johnston, E.L.; Glasby, T.M. Shallow moving structures promote marine invader dominance. Biofouling 2009, 25, 277–287. [Google Scholar] [CrossRef]
- Thiel, M.; Guerra-García, J.M.; Lancellotti, D.A.; Vásquez, N. The distribution of littoral caprellids (Crustacea: Amphipoda: Caprellidae) along the Pacific coast of continental Chile. Rev. Chil. Hist. Nat. 2003, 76, 203–218. [Google Scholar] [CrossRef]
- Guerra-Garcia, J.M.; Cabezas, P.; Baeza-Rojano, E.; Espinosa, F.; García-Gómez, J.C. Is the north side of the Strait of Gibraltar more diverse than the south side? A case study using the intertidal peracarids (Crustacea: Malacostraca) associated to the seaweed Corallina elongata. J. Mar. Biolog. 2009, 89, 387–397. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Ganesh, T.; Jaikumr, M.; Rama, A.V. Caprellids (Crustacea: Amphipoda) from India. Helgol. Mar. Res. 2010, 64, 297–310. [Google Scholar] [CrossRef]
- Pereira, S.G.; Lima, F.P.; Queiroz, N.C.; Ribeiro, P.A.; Santos, A.M. Biogeographic patterns of intertidal macroinvertebrates and their association with macroalgae distribution along the Portuguese coast. Hydrobiologia 2006, 555, 185–192. [Google Scholar] [CrossRef]
- Carlton, J.T. Biological Invasions and Cryptogenic Species. Ecology 1996, 77, 1653–1655. [Google Scholar] [CrossRef]
- Ros, M.; Guerra-García, J.M.; González-Macías, M.; Saavedra, Á.; López-Fe, C.M. Influence of fouling communities on the establishment success of alien caprellids (Crustacea: Amphipoda) in Southern Spain. Mar. Biol. Res. 2013, 9, 261–273. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997; ISBN 978-0-5118-0640-7. [Google Scholar]
- Underwood, A.J.; Chapman, M.G.; Richards, S. GMAV-5 for Windows. An Analysis of Variance Programme; University of Sydney: Sydney, Australia, 2002. [Google Scholar]
- Anderson, M.J.; Legendre, P. An empirical comparison of permutation methods for tests of partial regression coefficients in a linear model. J. Stat. Comput. Simul. 1999, 62, 271–303. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Guerra-García, J.M.; Tierno de Figueroa, J.T. What do caprellids (Crustacea: Amphipoda) feed on? Mar. Biol. 2009, 156, 1881–1890. [Google Scholar] [CrossRef]
- Ros, M.; Vázquez-Luis, M.; Guerra-García, J.M. The role of marinas and recreational boating in the occurrence and distribution of exotic caprellids (Crustacea: Amphipoda) in the Western Mediterranean: Mallorca Island as a case study. J. Sea Res. 2013, 83, 94–103. [Google Scholar] [CrossRef]
- De Rivera, C.E.; Ruiz, G.; Crooks, J.; Wasson, K.; Lonhart, S. Broadscale non-indigenous species monitoring along the west coast in national marine sanctuaries and national estuarine research reserves. Environ. Sci. Manag. 2005, 76, 1–56. [Google Scholar]
- Tamburini, M.; Keppel, E.; Marchini, A.; Repetto, M.F.; Ruiz, G.M.; Ferrario, J.; Occhipinti-Ambrogi, A. Monitoring non-indigenous species in port habitats: First application of a standardized North American protocol in the Mediterranean Sea. Front. Mar. Sci. 2021, 8, 700730. [Google Scholar] [CrossRef]
- Floerl, O.; Inglis, G.J. Boat harbour design can exacerbate hull fouling. Austral. Ecol. 2003, 28, 116–127. [Google Scholar] [CrossRef]
- Koehl, M.R.A. Mini review: Hydrodynamics of larval settlement into fouling communities. Biofouling 2007, 23, 357–368. [Google Scholar] [CrossRef] [PubMed]
- McCain, J.C.; Steinberg, J.E. Amphipoda-I, Caprellidea-I. In Crustaceorum Catalogus; Gruner, H.E., Holthuis, L.B., Eds.; W. Junk: The Hague, The Netherlands, 1970; Volume 2, pp. 1–78. [Google Scholar]
- Bellan-Santini, D.; Ruffo, S. Faunistics and Zoogeography. In The Amphipoda of the Mediterranean, Part 4. Mémoires de l’Institut Océanographique (Monaco); Ruffo, S., Ed.; Institut Océanographique: Monaco, 1998; Volume 13, pp. 895–911. ISBN 2-7260-0201-3. [Google Scholar]
- Katsanevakis, S.; Crocetta, F. Pathways of introduction of marine alien species in European waters and the Mediterranean—A possible undermined role of marine litter. In Proceedings of the Marine Litter in the Mediterranean and Black Seas, CIESM Workshop, Tirana, Albania, 18–21 June 2014; CIESM Publisher: Monaco, Monaco, 2014; Volume 46, pp. 6–68. [Google Scholar]
- Fernandez-Gonzalez, V.; Sanchez-Jerez, P. Fouling assemblages associated with off-coast aquaculture facilities: An overall assessment of the Mediterranean Sea. Mediterr. Mar. Sci. 2017, 18, 87–96. [Google Scholar] [CrossRef]
- Leclerc, J.C.; Viard, F. Habitat formation prevails over predation in influencing fouling communities. Ecol. Evol. 2018, 8, 477–492. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, T.; Sukumaran, S.; Dias, H.Q. Extended distribution of Phtisica marina Slabber, 1769 (Crustacea: Amphipoda): First observation of alien Caprellid in the coastal waters of Indian subcontinent. Bioinvasions Rec. 2019, 8, 96–107. [Google Scholar] [CrossRef]
- Cabezas, M.P.; Ros, M.; Dos Santos, A.M.; Martinez-Laiz, G.; Xavier, R.; Montelli, L.; Hoffman, R.; Fersi, A.; Claude-Davin, J.; Guerra-Garcia, J.M. Unravelling the origin and introduction pattern of the tropical species Paracaprella pusilla Mayer, 1890 (Crustacea, Amphipoda, Caprellidae) in temperate European waters: First molecular insights from a spatial and temporal perspective. NeoBiota 2019, 47, 43–80. [Google Scholar] [CrossRef]
- Ashton, G.V. Distribution and Dispersal of the Non-Native Caprellid Amphipod, Caprella mutica Schurin, 1935. Ph.D. Thesis, University of Aberdeen, Aberdeen, UK, 2006. [Google Scholar]
- Ros, M.; Guerra-García, J.M. On the occurrence of the tropical caprellid Paracaprella pusilla Mayer, 1890 (Crustacea: Amphipoda) in Europe. Mediterr. Mar. Sci. 2012, 13, 134–139. [Google Scholar] [CrossRef]
- Martínez-Laiz, G.; Guerra-García, J.M.; Ros, M.; Fenwick, D.; Bishop, J.D.; Horton, T.; Faase, M.A.; Cabezas, M.P. Hitchhiking northwards: On the presence of the invasive skeleton shrimp Caprella scaura in the UK. Mar. Biodiv. 2021, 51, 78. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Ros, M.; Gordillo, I.; Cabezas, M.D.P.; Baeza-Rojano, E.; Izquierdo, D.; Corzo, J.; Domínguez, J.; Varona, S. Distribution patterns of intertidal and shallow water caprellids associated with macroalgae along the Iberian Peninsula. Zool. Baetica. 2011, 21, 101–129. [Google Scholar]
- Baeza-Rojano, E.; Guerra-García, J.M. Ciclo de vida bajo condiciones de laboratorio de los caprélidos (Crustacea: Amphipoda) del sur de la Península Ibérica: Caprella equilibra y Caprella dilatata (Caprellidae) y Phtisica marina (Phtisicidae). Zool. Baetica. 2013, 24, 155–186. [Google Scholar]
- Conradi, M.; López-González, P.J.; Cervera, J.L.; García-Gómez, J.C. Seasonality and spatial distribution of peracarids associated with the bryozoan Bugula neritina in Algeciras Bay, Spain. J. Crust. Biol. 2000, 20, 334–349. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Ros, M.; Dugo-Cota, A.; Burgos, V.; Flores-León, A.M.; Baeza-Rojano, E.; Núñez, J. Geographical expansion of the invader Caprella scaura (Crustacea: Amphipoda: Caprellidae) to the East Atlantic coast. Marine Biol. 2011, 158, 2617–2622. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A.; Minchin, D.; Narščius, A.; Ojaveer, H.; Olenin, S. International arrivals: Widespread bioinvasions in European Seas. Ethol. Ecol. Evol. 2014, 26, 152–171. [Google Scholar] [CrossRef]
- Cabezas, M.P.; Xaiver, R.; Branco, M.; Santos, A.M.; Guerra-García, J.M. Invasion history of Caprella scaura Templeton, 1836 (Amphipoda: Caprellidae) in the Iberian Peninsula: Multiple introductions revealed by mitochondrial sequence data. Biol. Invasions 2014, 16, 2221–2245. [Google Scholar] [CrossRef]
- Ros, M.; Guerra-García, J.M.; Navarro-Barranco, C.; Cabezas, M.P.; Vázquez-Luis, M.P. The spreading of the non-native caprellid (Crustacea: Amphipoda) Caprella scaura Templeton, 1836 into southern Europe and northen Africa: A complicated taxonomic history. Mediterr. Mar. Sci. 2014, 15, 145–155. [Google Scholar] [CrossRef]
- Martínez-Laiz, G.; Ros, M.; Guerra-García, J.; Faasse, M.; Santos, A.; Cabezas, M. Using molecular data to monitor the post establishment evolution of the invasive skeleton shrimp Caprella scaura. Mar. Environ. Res. 2021, 166, 105266. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.; Adarraga, I. First record of invasive caprellid Caprella scaura Templeton, 1836 sensu lato (Crustacea: Amphipoda: Caprellidae) from the Iberian Peninsula. Aquat. Invasions 2008, 3, 165–171. [Google Scholar] [CrossRef]
- Fofonoff, P.W.; Ruiz, G.M.; Steves, B.; Simkanin, C.; Carlton, J.T. National Exotic Marine and Estuarine Species Information System. Available online: http://invasions.si.edu/nemesis (accessed on 12 December 2021).
- Lo Brutto, S.; Iaciofano, D.; Guerra-García, J.M.; Lubinevsky, H.; Galil, B.S. Desalination effluents and the establishment of the non-indigenous skeleton shrimp Paracaprella pusilla Mayer, 1890 in the south-eastern Mediterranean. BioInvasions Rec. 2019, 8, 661–669. [Google Scholar] [CrossRef]
- Fersi, A.; Davin, J.; Pezy, J.; Neifar, L. Amphipods from tidal channels of the Gulf of Gabès (central Mediterranean Sea). Mediterra. Mar. Sci. 2018, 19, 430–443. [Google Scholar] [CrossRef]
- Ros, M.; Vázquez-Luis, M.; Guerra-García, J.M. The tropical caprellid amphipod Paracaprella pusilla: A new alien crustacean in the Mediterranean Sea. Helgol. Mar. Res. 2013, 67, 675–685. [Google Scholar] [CrossRef]
- Ros, M.; Guerra-García, J.M.; Hoffman, R. First record of the exotic caprellid amphipod Paracaprella pusilla Mayer, 1890 in the eastern Mediterranean. Mar. Biodivers. 2016, 46, 281–284. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | df | MS | F | p | Source of Variation | df | MS | F | p |
|---|---|---|---|---|---|---|---|---|---|
| Total Abundance | Number of Species | ||||||||
| Time = TI | 1 | 23.3316 | 1.67 | 0.3258 | Time = TI | 1 | 4.0833 | 2.41 | 0.2608 |
| Locality = LO | 2 | 181.3196 | 33.82 | 0.0000 *** | Locality = LO | 2 | 17.1944 | 26.15 | 0.0000 *** |
| Substrate = SU | 1 | 62.3382 | 5.50 | 0.1437 | Substrate = SU | 1 | 7.7870 | 9.24 | 0.0933 |
| TI × LO | 2 | 14.0017 | 2.61 | 0.0786 | TI × LO | 2 | 1.6944 | 2.58 | 0.0812 |
| TI × SU | 1 | 9.7162 | 0.76 | 0.4757 | TI × SU | 1 | 2.0833 | 1.74 | 0.3175 |
| LO × SU | 2 | 11.3397 | 2.12 | 0.1262 | LO × SU | 2 | 0.8426 | 1.28 | 0.2823 |
| TI × LO × SU | 2 | 12.8171 | 2.39 | 0.0970 | TI × LO × SU | 2 | 1.1944 | 1.82 | 0.1681 |
| Residual | 96 | 5.3607 | Residual | 96 | 0.6574 | ||||
| Total | 107 | Total | 107 | ||||||
| Cochran’s C-test | C = 0.2018 | Cochran’s C-test | C = 0.1585 | ||||||
| Transformation | Ln (X + 1) | Transformation | None | ||||||
| SNK | LO: L1 > L3 > L2 | SNK | LO: L1 > L3 > L2 | ||||||
 |

| Source of Variation | df | MS | Pseudo-F | P(perm) | P(MC) |
|---|---|---|---|---|---|
| Abundance (Non-Transformed) | |||||
| Substrata = SU | 1 | 11,273 | 1.3592 | 0.2146 | 0.1639 |
| Time = TI | 1 | 17,597 | 1.8889 | 0.0488 | 0.0225 * |
| Locality = LO | 2 | 18,252 | 4.5744 | 0.0001 | 0.0001 *** |
| SU × TI | 1 | 9403.5 | 1.5467 | 0.112 | 0.0856 |
| SU × LO | 2 | 8293.7 | 2.0786 | 0.0001 | 0.0003 *** |
| TI × LO | 2 | 9315.8 | 2.3347 | 0.0001 | 0.0001 *** |
| SU × TI × LO | 2 | 6079.5 | 1.5237 | 0.0069 | 0.0141 * |
| Residual | 96 | 3990.1 | |||
| Total | 107 | ||||
| Pair-Wise tests a | SU × TI × LO | ||||
| T1 × L1, L2: CO ≠ LB; T1 × L3: CO = LB. | |||||
| T3 × L1: CO ≠ LB; T3 × L2, L3: CO = LB. | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Revanales, T.; Guerra-García, J.M.; Ros, M. Colonization Dynamics of Potential Stowaways Inhabiting Marinas: Lessons from Caprellid Crustaceans. Water 2022, 14, 2659. https://doi.org/10.3390/w14172659
Revanales T, Guerra-García JM, Ros M. Colonization Dynamics of Potential Stowaways Inhabiting Marinas: Lessons from Caprellid Crustaceans. Water. 2022; 14(17):2659. https://doi.org/10.3390/w14172659
Chicago/Turabian StyleRevanales, Triana, José M. Guerra-García, and Macarena Ros. 2022. "Colonization Dynamics of Potential Stowaways Inhabiting Marinas: Lessons from Caprellid Crustaceans" Water 14, no. 17: 2659. https://doi.org/10.3390/w14172659