


Reoligotrophication of a High-Nitrogen Reservoir with Phosphorus Removal and Implications for Management

Abstract
:1. Introduction
2. Materials and Methods
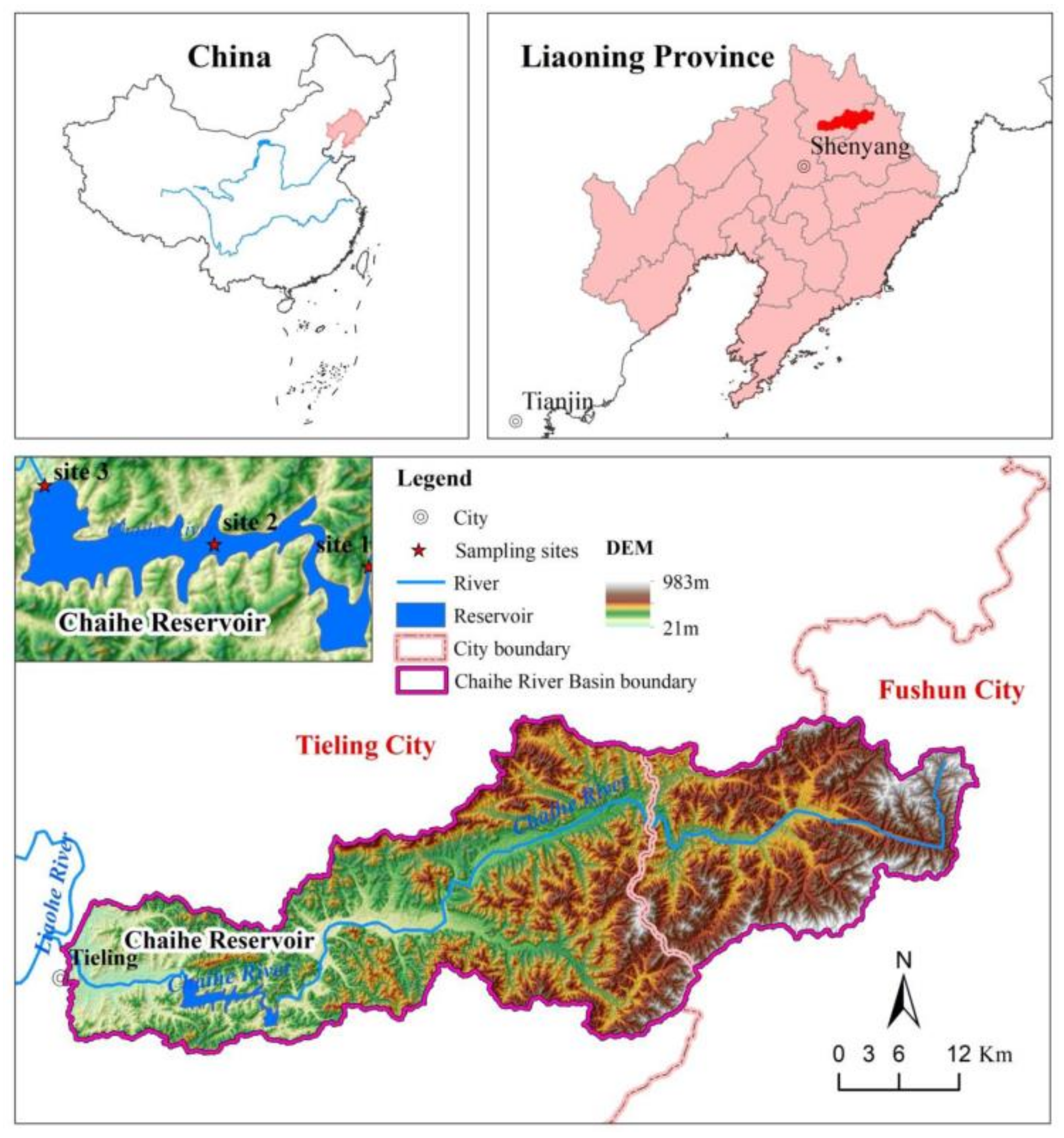
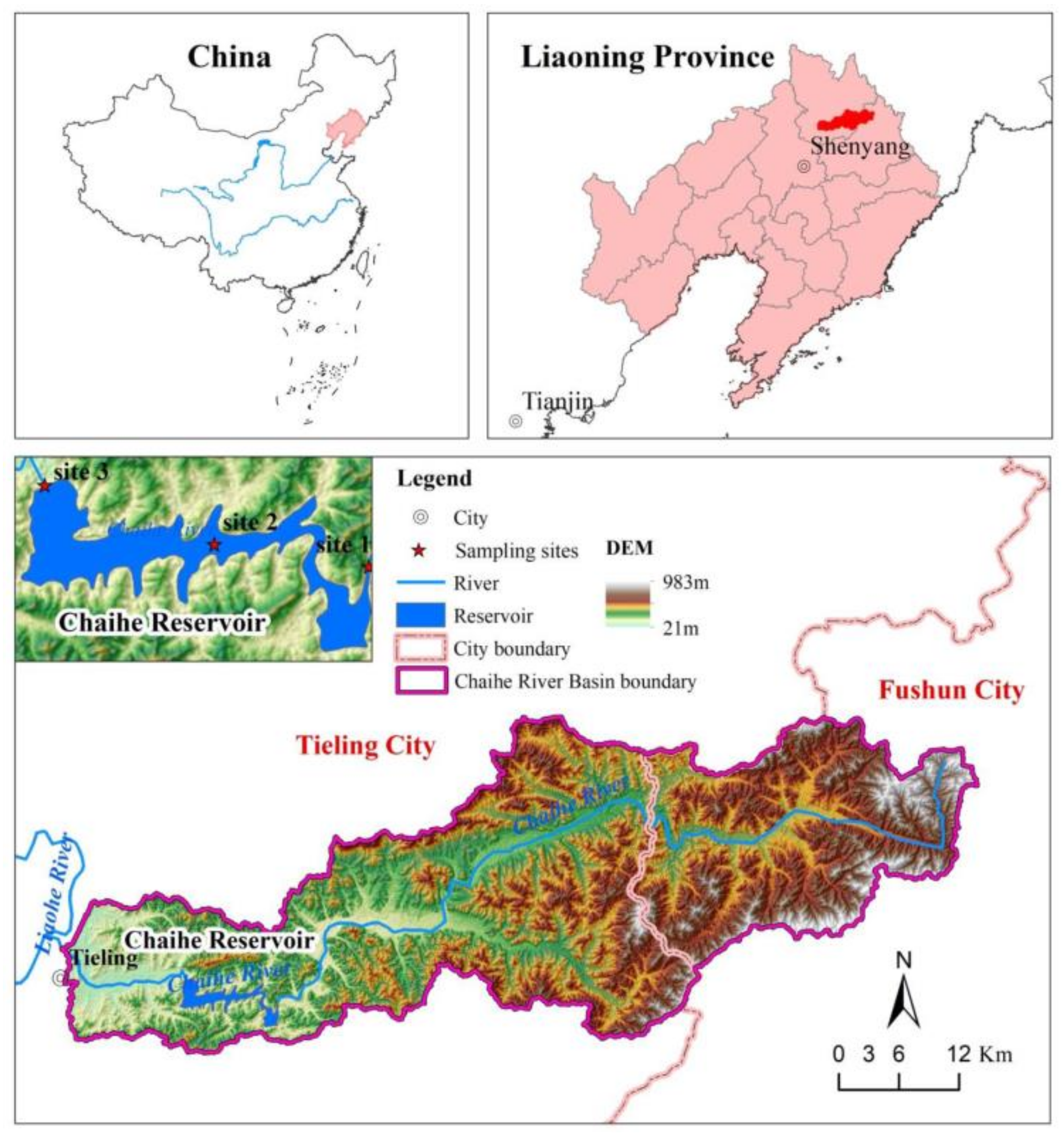
2.1. Study Area
2.2. Sampling and Procedures
2.3. Data Analysis
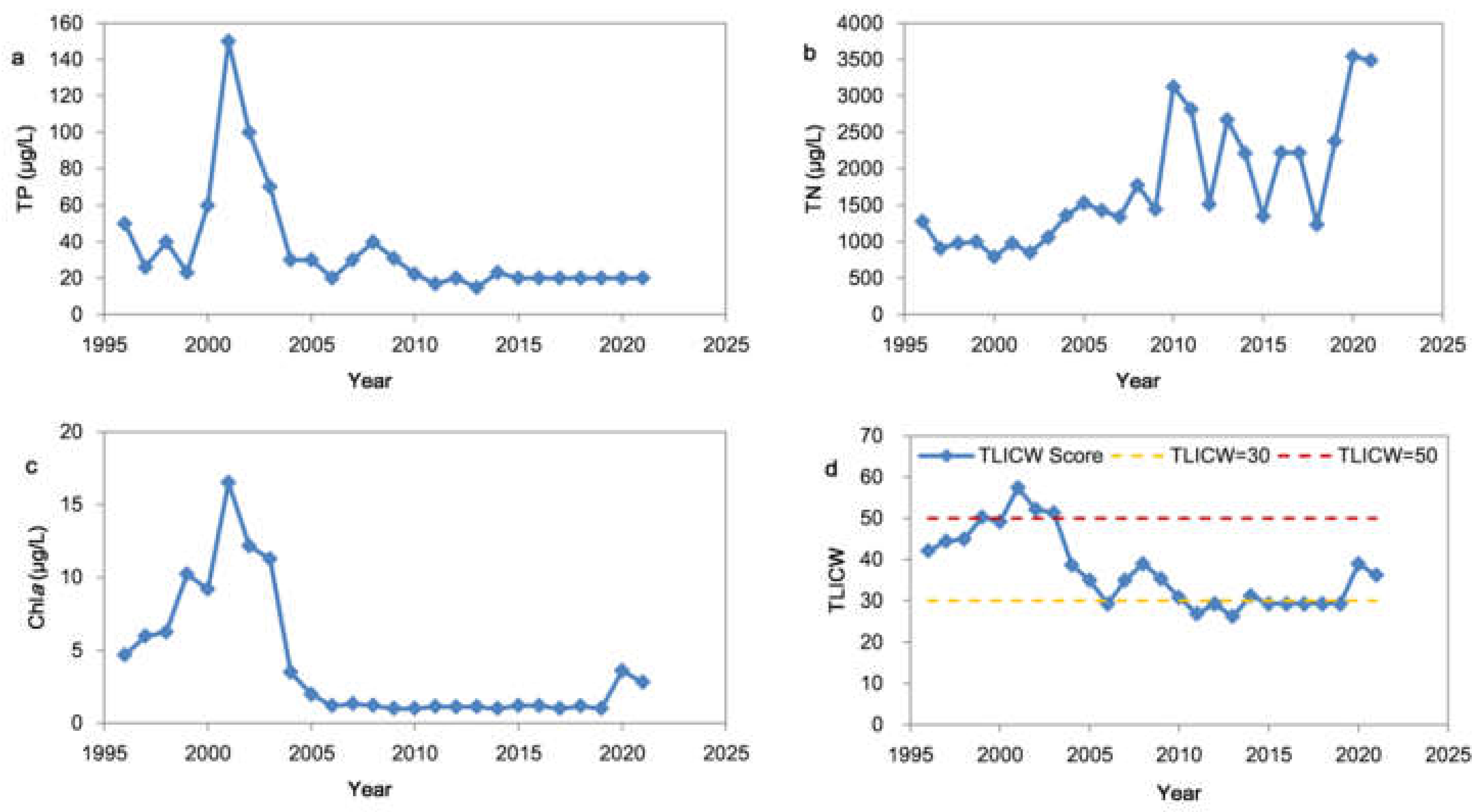
2.3.1. Changed Weight Trophic Level Index Method (TLICW)
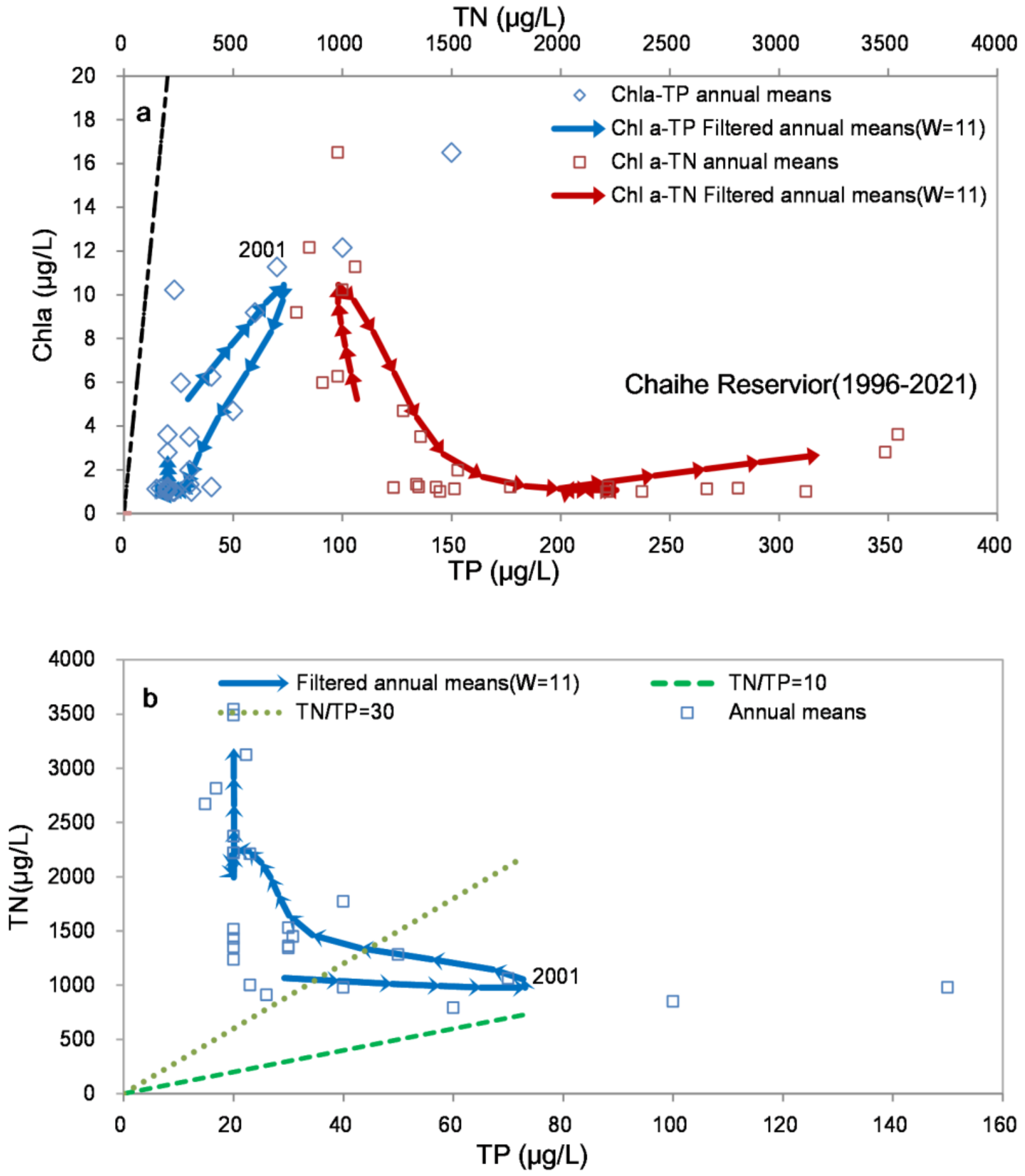
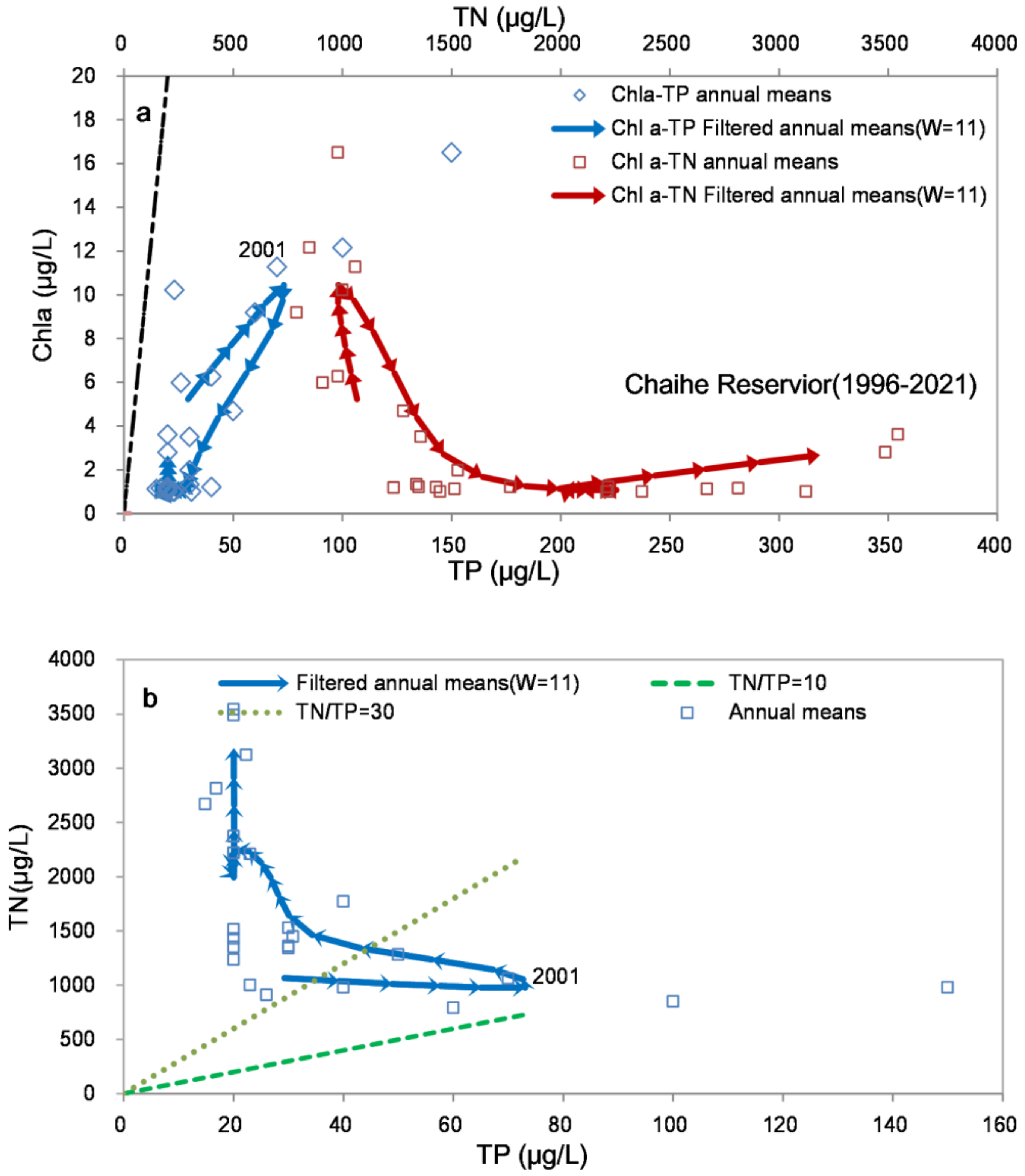
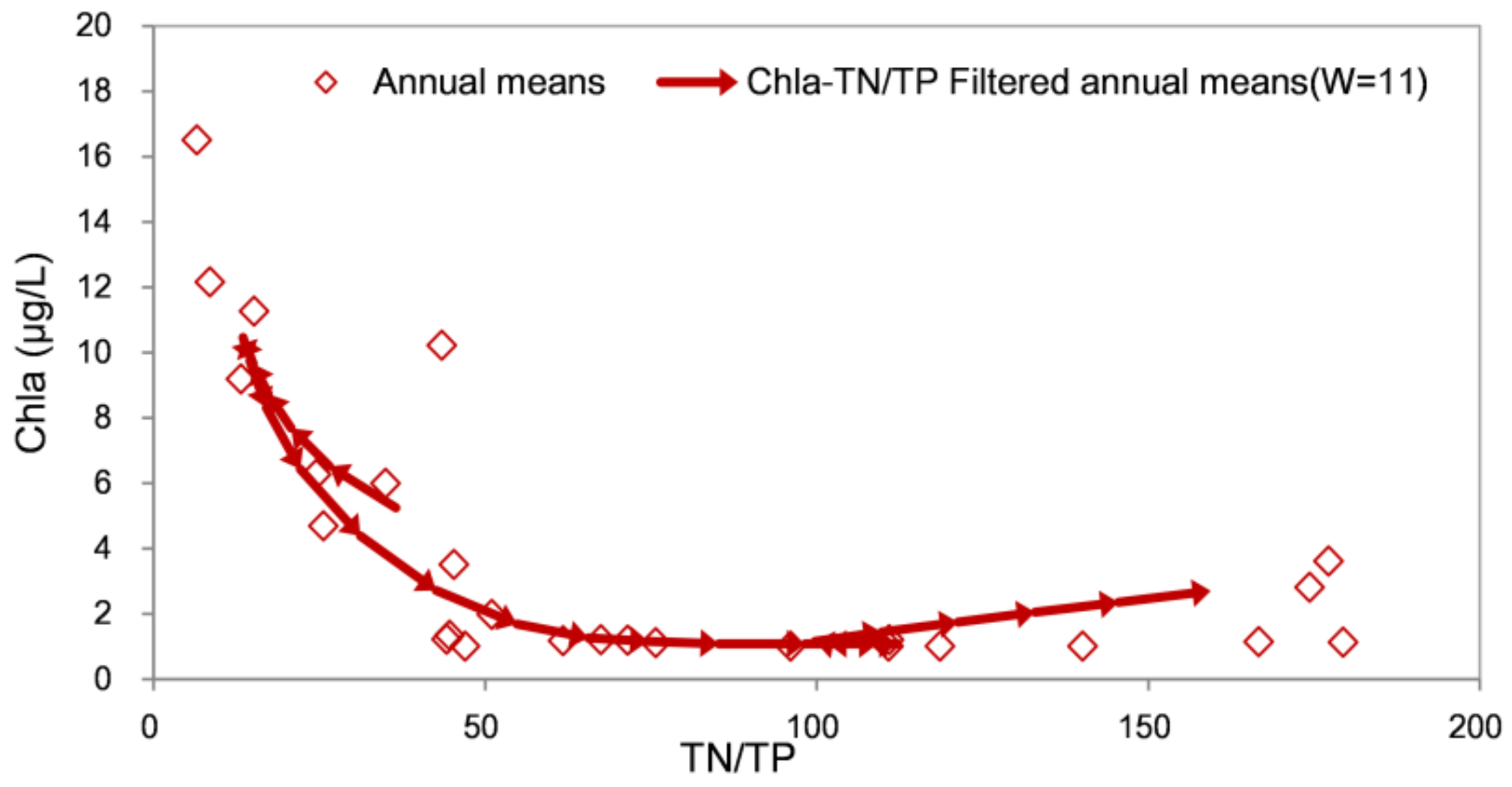
2.3.2. Filtering Trajectory Method (FTM)
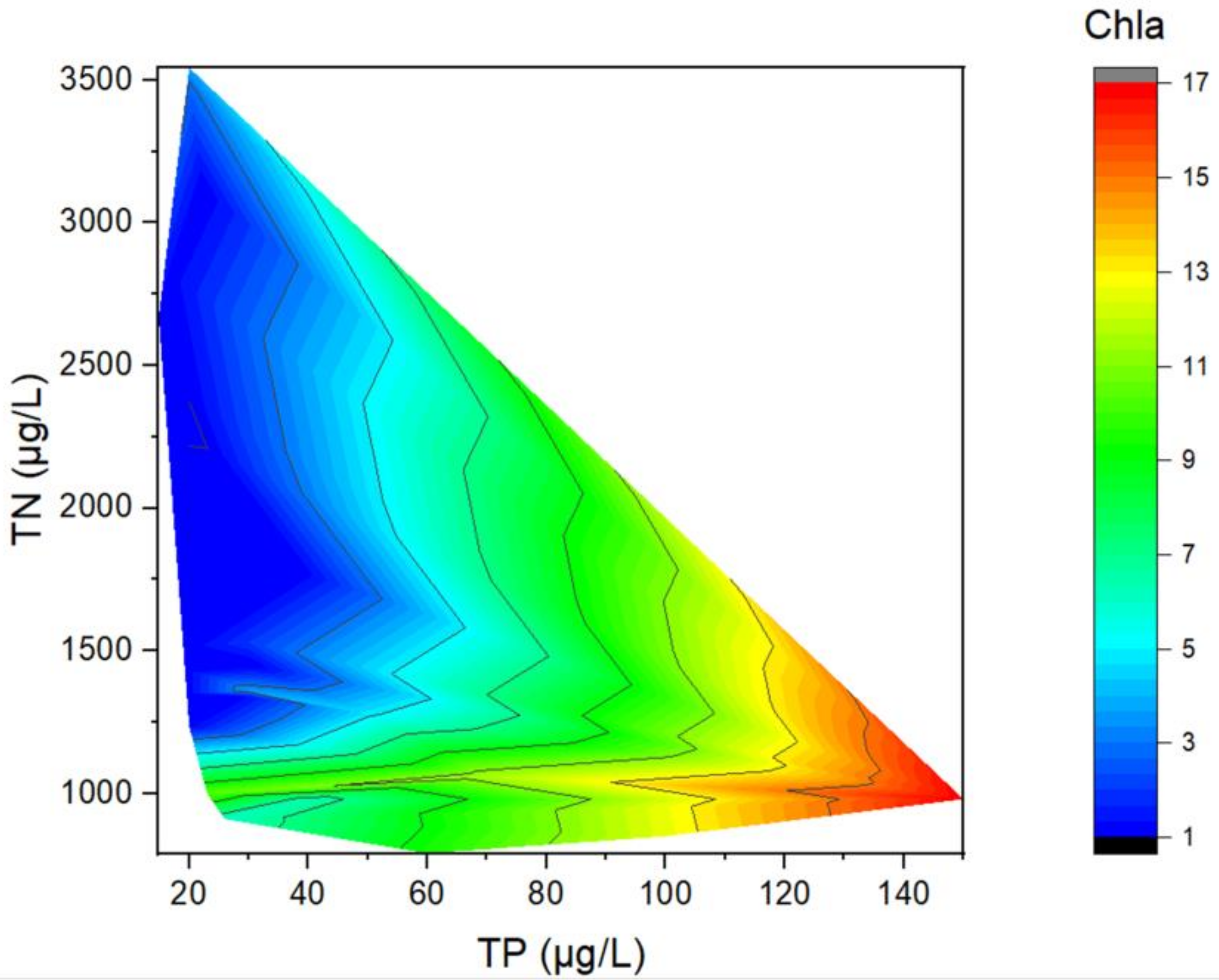
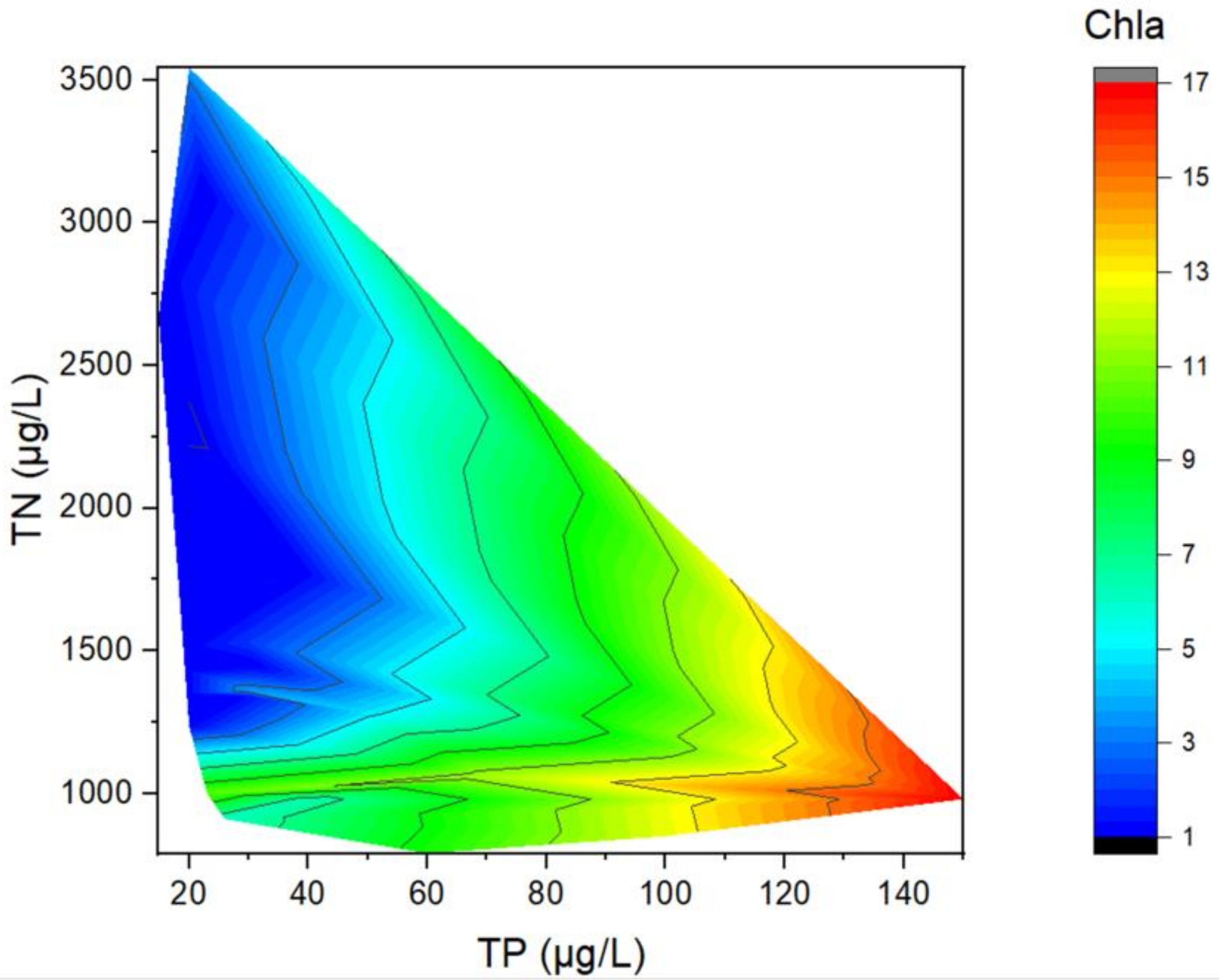
2.3.3. Contour Plot
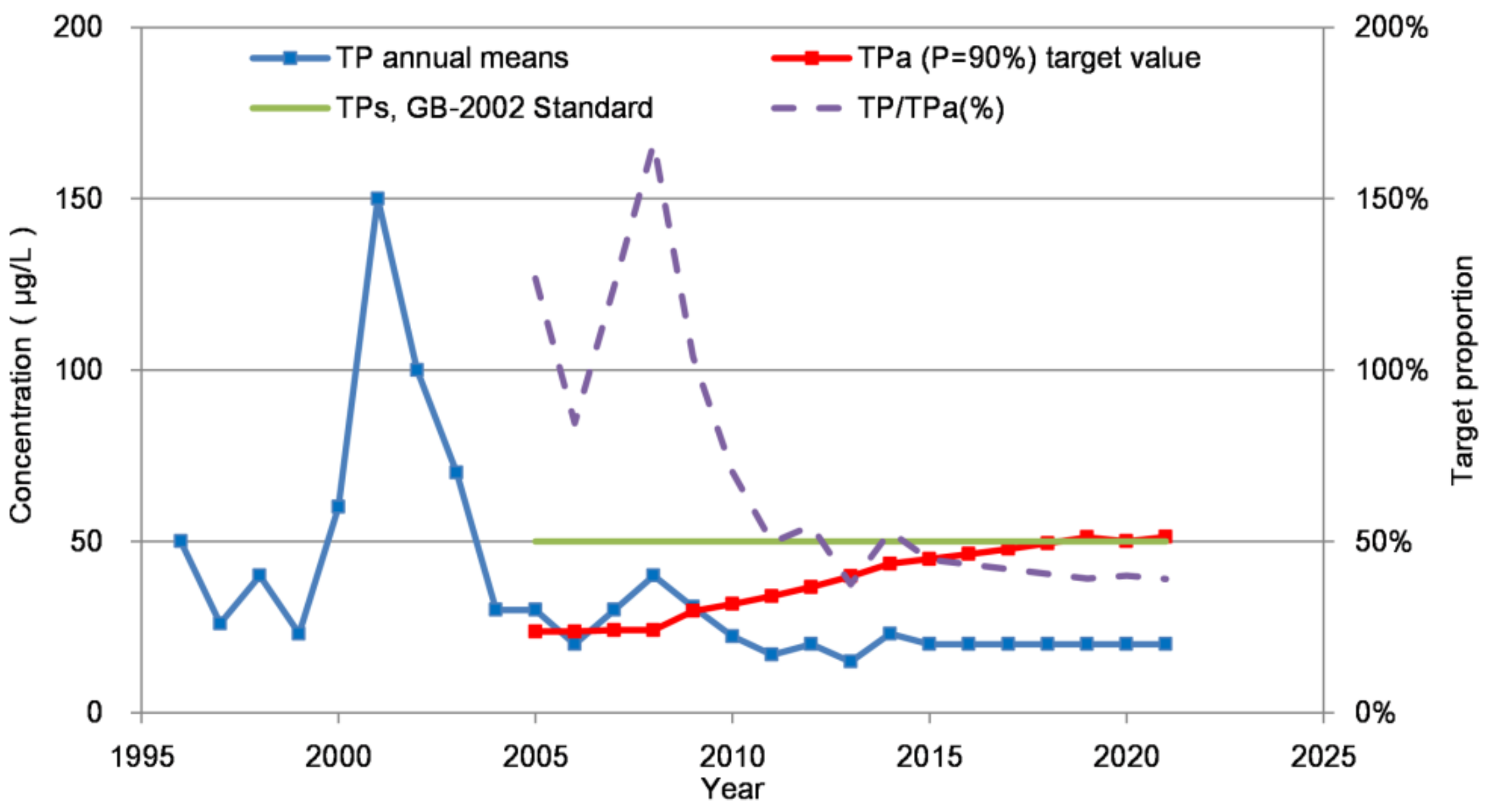
2.3.4. Double Probability Method (DPM)
3. Results
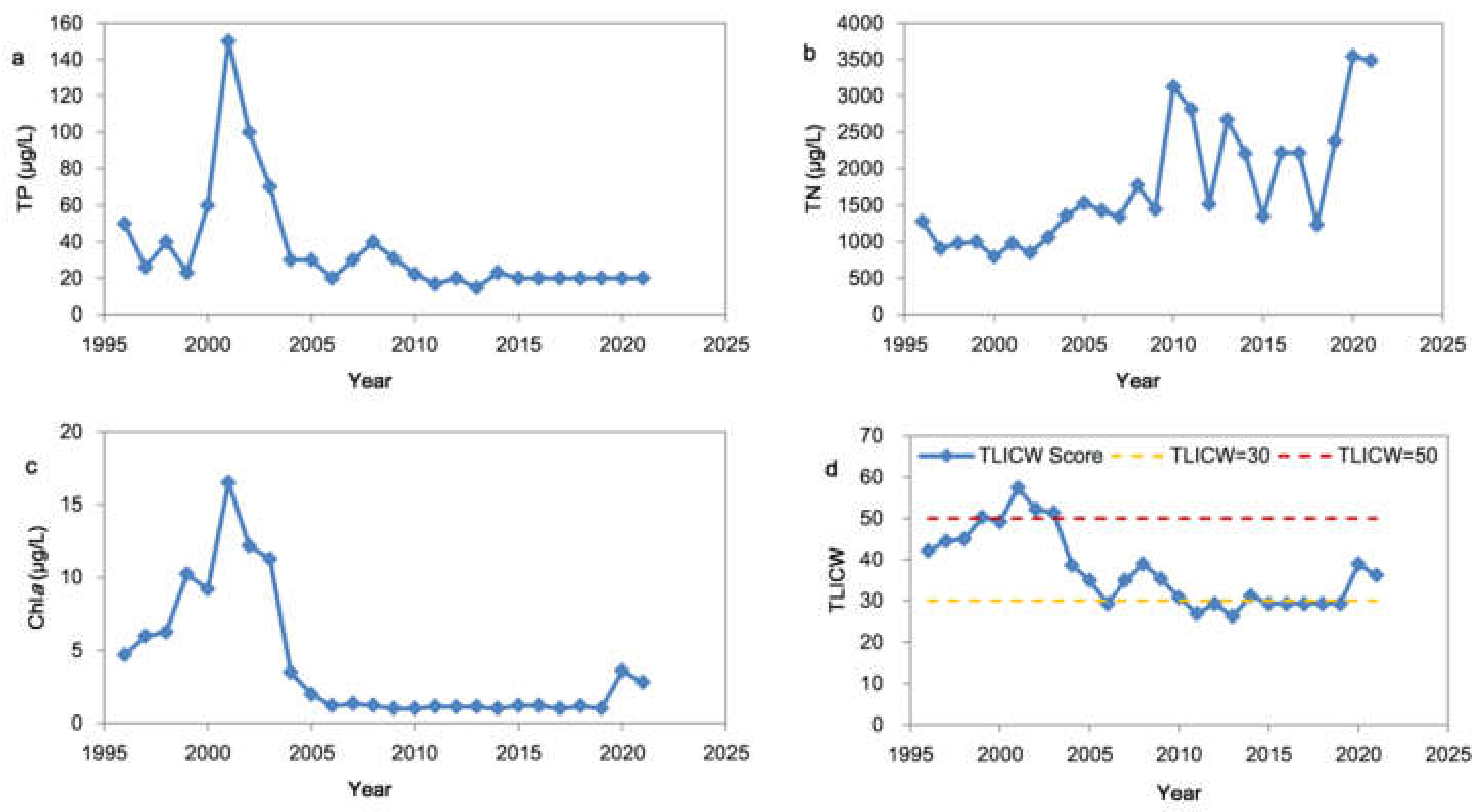
3.1. Interannual Variations of TP, TN, Chla and the Trophic Status
3.2. Relationships between Chla and TP (TN)
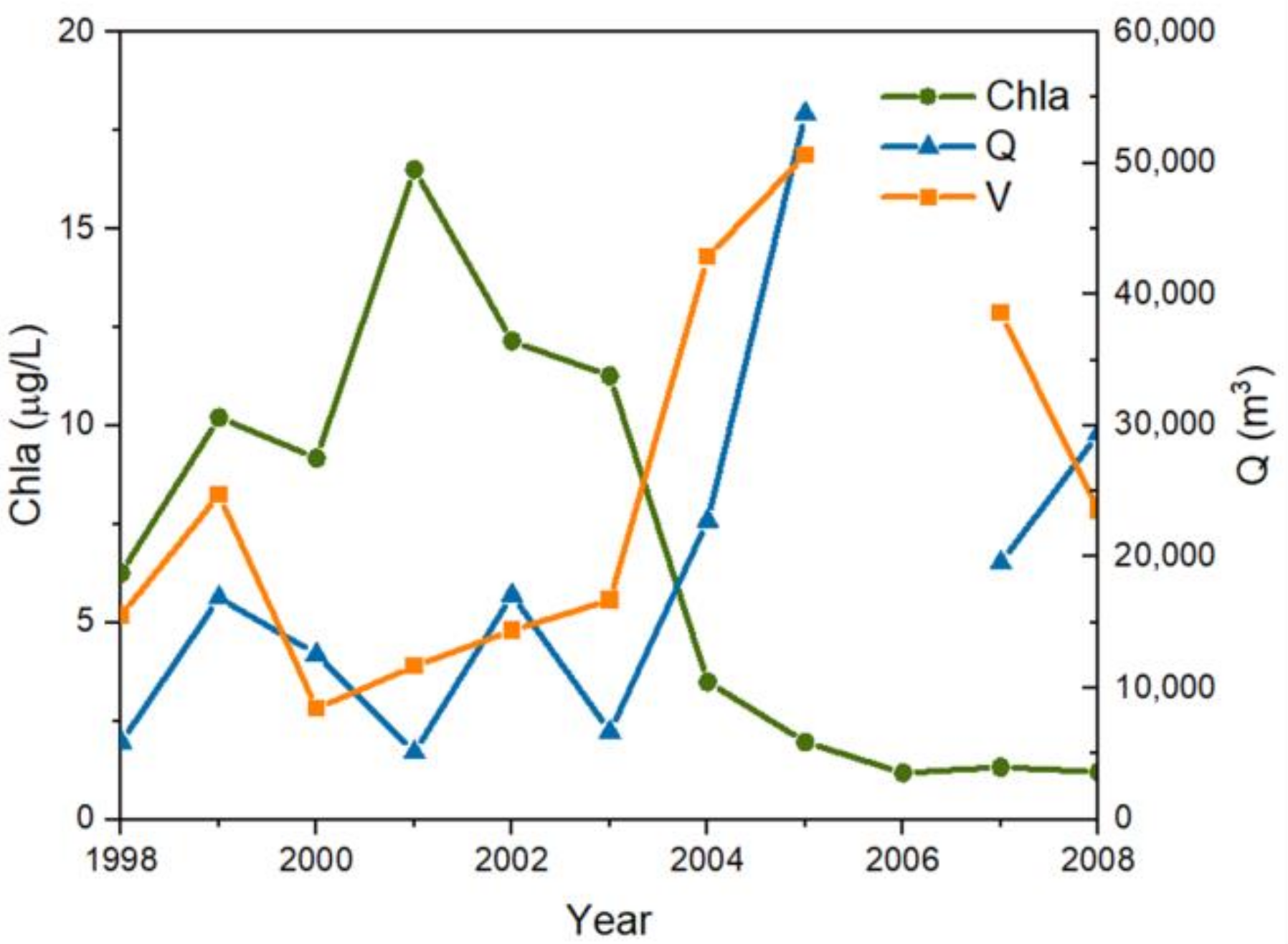
3.3. Chla Response to Hydraulic Factors and Changes in Nutrient Concentrations
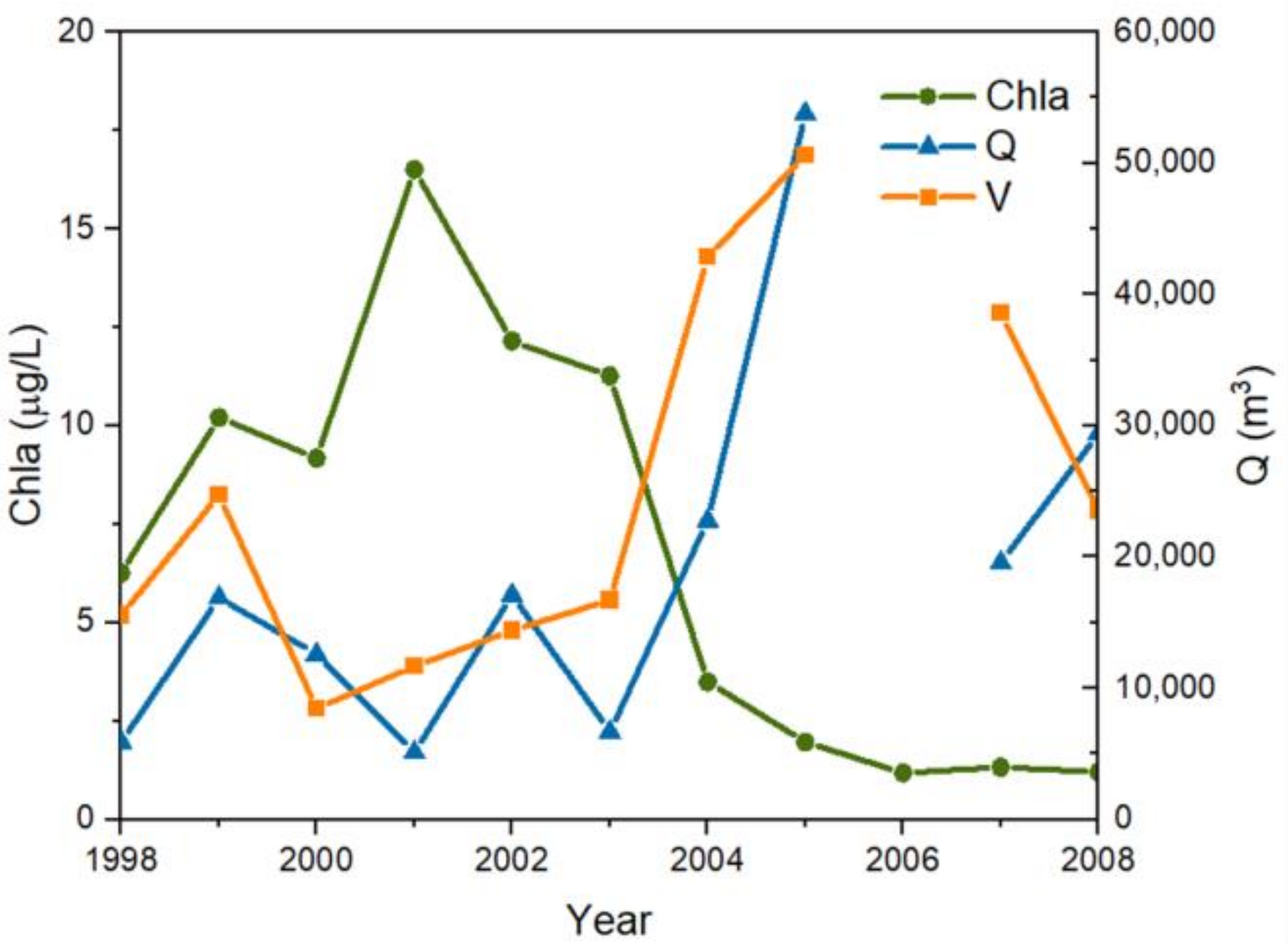
3.3.1. Chla Response to Hydraulic Factors
3.3.2. Chla Response to Changes in Nutrient Concentrations
3.4. Setting the Nutrient Targets
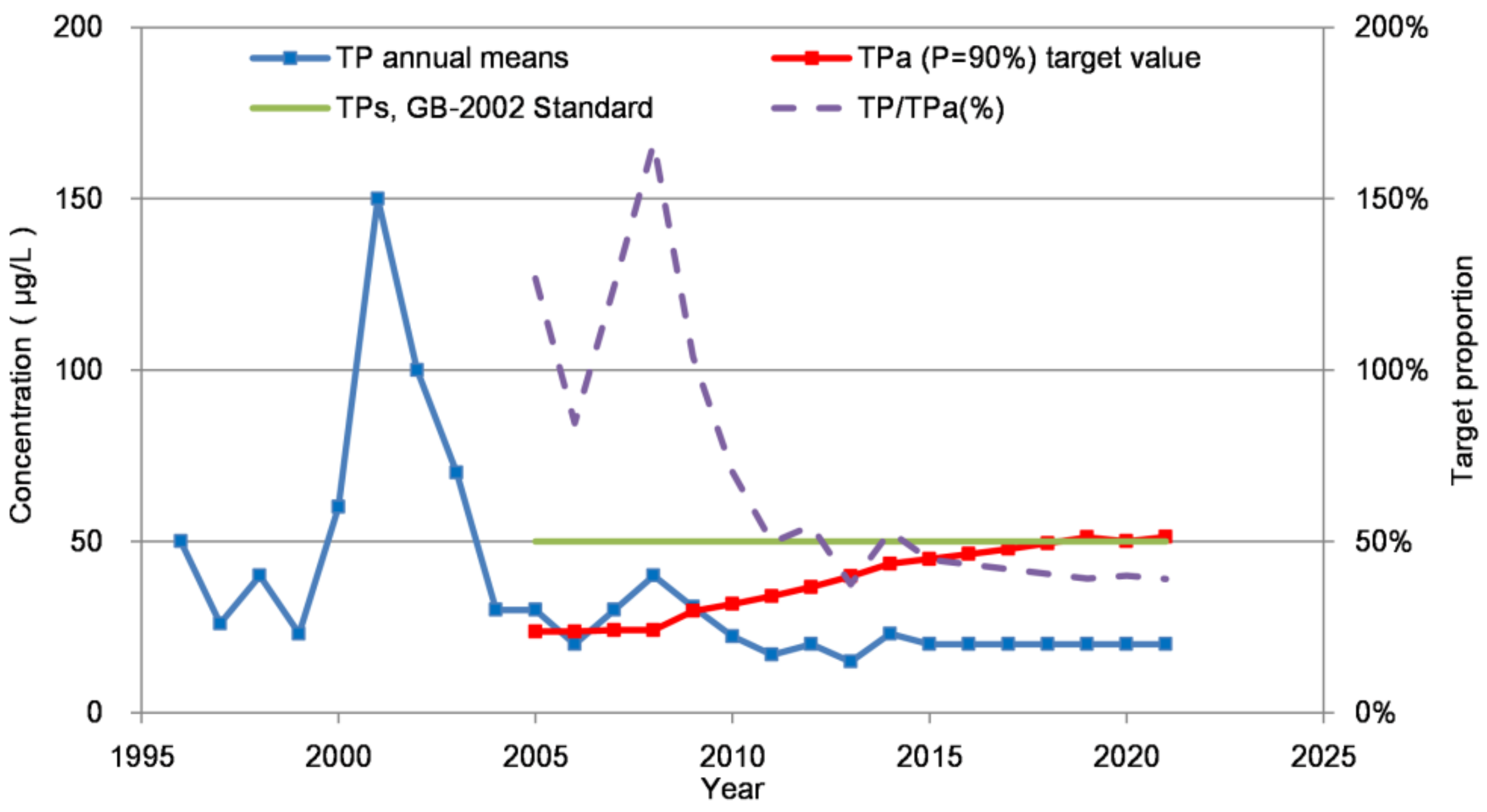
3.4.1. Interannual Variation of TPa
3.4.2. The Thresholds for TP and TN under Different Chla Targets
4. Discussion
4.1. Nitrogen Control Was Unnecessary for Oligotrophication in P-Limited Systems
4.2. Phosphorus Reduction Efforts Exacerbated Nitrogen Pollution
4.3. Nutrient Thresholds and Management Strategies for the Chaihe Reservoir
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, V.H. Responses of estuarine and coastal marine phytoplankton to nitrogen and phosphorus enrichment. Limnol. Oceanogr. 2006, 51, 377–384. [Google Scholar] [CrossRef]
- Schindler, D.W. Recent advances in the understanding and management of eutrophication. Limnol. Oceanogr 2006, 51, 356–363. [Google Scholar]
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems—A global problem. Environ. Sci. Pollut. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef]
- Kaste, Ø.; Lyche-Solheim, A. Influence of moderate phosphate addition on nitrogen retention in an acidic boreal lake. Can. J. Fish. Aquat. Sci. 2005, 62, 312–321. [Google Scholar] [CrossRef]
- Peterson, B.J.; Hobbie, J.E.; Hershey, A.E.; Lock, M.A.; Ford, T.E.; Vestal, J.R.; McKinley, V.L.; Hullar, M.A.J.; Miller, M.C.; Ventullo, R.M.; et al. Transformation of a Tundra River from Heterotrophy to Autotrophy by Addition of Phosphorus. Science 1985, 229, 1383–1386. [Google Scholar] [CrossRef]
- Davison, W.; George, D.G.; Edwards, N.J.A. Controlled reversal of lake acidification by treatment with phosphate fertilizer. Nature 1995, 377, 504–507. [Google Scholar] [CrossRef]
- Finlay, J.C.; Small, G.E.; Sterner, R.W. Human influences on nitrogen removal in lakes. Science 2013, 342, 247–250. [Google Scholar] [CrossRef]
- Bernhardt, E.S. Ecology. Cleaner lakes are dirtier lakes. Science 2013, 342, 205–206. [Google Scholar] [CrossRef]
- Bergstrom, A.K.; Blomqvist, P.; Jansson, M. Effects of atmospheric nitrogen deposition on nutrient limitation and phytoplankton biomass in unproductive Swedish lakes. Limnol. Oceanogr. 2005, 50, 987–994. [Google Scholar] [CrossRef]
- Wang, H.; Wang, T.; Yang, S.; Liu, X.; Kou, L.; Huang, T.; Wen, G. Nitrogen Removal in Oligotrophic Reservoir Water by a Mixed Aerobic Denitrifying Consortium: Influencing Factors and Immobilization Effects. Int. J. Environ. Res. Public Health 2019, 16, 583. [Google Scholar] [CrossRef]
- Müller, B.; Meyer, J.S.; Gächter, R. Denitrification and Nitrogen Burial in Swiss Lakes. Environ. Sci. Technol. 2022, 56, 2794–2802. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling Eutrophication: Nitrogen and Phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Scott, J.T.; McCarthy, M.J.; Newell, S.E.; Gardner, W.S.; Havens, K.E.; Hoffman, D.K.; Wilhelm, S.W.; Wurtsbaugh, W.A. It Takes Two to Tango: When and Where Dual Nutrient (N & P) Reductions Are Needed to Protect Lakes and Downstream Ecosystems. Environ. Sci. Technol. 2016, 50, 10805–10813. [Google Scholar] [CrossRef] [PubMed]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Han, X.; Fu, g.; Liu, q.; Zhang, S.; Zhao, J. Changed weight trophic level index method and its application to Longhe tributary of the Three Gorges reservoir area. J. Environ. Eng. Technol. 2018, 8, 627–634. [Google Scholar] [CrossRef]
- Zhao, J.; Fu, G. Assessment of lake eutrophication recovery: The filtering trajectory method (FTM) and its application to Dianchi Lake, China. Environ. Monit. Assess. 2019, 191, 360. [Google Scholar] [CrossRef]
- Schindler, D.W. Evolution of Phosphorus Limitation in Lakes. Science 1977, 195, 260–262. [Google Scholar] [CrossRef]
- Schindler, D.W. The dilemma of controlling cultural eutrophication of lakes. Proc. Biol. Sci 2012, 279, 4322–4333. [Google Scholar] [CrossRef]
- Jeppesen, E.; Sondergaard, M.; Jensen, J.P.; Havens, K.E.; Anneville, O.; Carvalho, L.; Coveney, M.F.; Deneke, R.; Dokulil, M.T.; Foy, B.O.B.; et al. Lake responses to reduced nutrient loading—An analysis of contemporary long-term data from 35 case studies. Freshw. Biol. 2005, 50, 1747–1771. [Google Scholar] [CrossRef]
- Welch, E.B. Should nitrogen be reduced to manage eutrophication if it is growth limiting? Evidence from Moses Lake. Lake Reserv. Manag. 2009, 25, 401–409. [Google Scholar] [CrossRef]
- Reid, N.J.; Hamilton, S.K. Controls on algal abundance in a eutrophic river with varying degrees of impoundment (Kalamazoo River, Michigan, USA). Lake Reserv. Manag. 2007, 23, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Padovesi-Fonseca, C.; Philomeno, M.; Andreoni-Batista, C. Limnological features after a flushing event in Paranoá Reservoir, central Brazil. Acta Limnol. Bras. 2009, 21, 277–285. [Google Scholar]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Dong, L.; He, W.; Wang, Q.; Mooij, W.M.; Xu, F. Estimation of the long-term nutrient budget and thresholds of regime shift for a large shallow lake in China. Ecol. Indic. 2015, 52, 231–244. [Google Scholar] [CrossRef]
- Klaus, M.; Bergstrom, A.-K.; Jonsson, A.; Deininger, A.; Geibrink, E.; Karlsson, J. Weak response of greenhouse gas emissions to whole lake N enrichment. Limnol. Oceanogr. 2018, 63, S340–S353. [Google Scholar] [CrossRef]
- He, L.; Cao, T.; Hu, X.; Zhang, H.; Zhang, X.; Ni, L.; Xie, P. Effects of NH4+and K+enrichments on carbon and nitrogen metabolism, life history and asexual reproduction ofVallisneria natansL. in aquarium experiments. J. Freshw. Ecol. 2014, 30, 391–406. [Google Scholar] [CrossRef]
- Gobler, C.J.; Burkholder, J.M.; Davis, T.W.; Harke, M.J.; Johengen, T.; Stow, C.A.; Van de Waal, D.B. The dual role of nitrogen supply in controlling the growth and toxicity of cyanobacterial blooms. Harmful Algae 2016, 54, 87–97. [Google Scholar] [CrossRef]
- Paerl, H.W.; Havens, K.E.; Xu, H.; Zhu, G.; McCarthy, M.J.; Newell, S.E.; Scott, J.T.; Hall, N.S.; Otten, T.G.; Qin, B. Mitigating eutrophication and toxic cyanobacterial blooms in large lakes: The evolution of a dual nutrient (N and P) reduction paradigm. Hydrobiologia 2020, 847, 4359–4375. [Google Scholar] [CrossRef]
- Sterner, R.W. On the Phosphorus Limitation Paradigm for Lakes. Int. Rev. Hydrobiol. 2008, 93, 433–445. [Google Scholar] [CrossRef]
- Liang, Z.; Soranno, P.A.; Wagner, T. The role of phosphorus and nitrogen on chlorophyll a: Evidence from hundreds of lakes. Water Res. 2020, 185, 116236. [Google Scholar] [CrossRef]
- van Gerven, L.P.A.; Kuiper, J.J.; Mooij, W.M.; Janse, J.H.; Paerl, H.W.; de Klein, J.J.M. Nitrogen fixation does not axiomatically lead to phosphorus limitation in aquatic ecosystems. Oikos 2019, 128, 563–570. [Google Scholar] [CrossRef]
- Zhou, J.; Han, X.X.; Brookes, J.D.; Qin, B.Q. High probability of nitrogen and phosphorus co-limitation occurring in eutrophic lakes. Environ. Pollut. 2022, 292, 118276. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, J.; Ding, L.; Li, Y.; Liu, H.; Zhao, Y.; Fu, G. Eutrophication evolution trajectory influenced by human activities and climate in the shallow Lake Gehu, China. Ecol. Indic. 2022, 138, 108821. [Google Scholar] [CrossRef]
- Wang, G.; Li, J.; Sun, W.; Xue, B.; Yinglan, A.; Liu, T. Non-point source pollution risks in a drinking water protection zone based on remote sensing data embedded within a nutrient budget model. Water Res. 2019, 157, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Xu, W.; Li, Q.; Han, M.; Tang, A.; Zhang, Y.; Luo, X.; Shen, J.; Wang, W.; Li, K.; et al. Changes of nitrogen deposition in China from 1980 to 2018. Environ. Int. 2020, 144, 106022. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, C.; Ji, L.; Liu, Y.; Xiao, J.; Cao, X.; Zhou, Y. Cause and effect of N/P ratio decline with eutrophication aggravation in shallow lakes. Sci. Total Environ. 2018, 627, 1294–1302. [Google Scholar] [CrossRef]
- Scott, J.T.; McCarthy, M.J.; Paerl, H.W. Nitrogen transformations differentially affect nutrient-limited primary production in lakes of varying trophic state. Limnol. Oceanogr. Lett. 2019, 4, 96–104. [Google Scholar] [CrossRef]
- Sollie, S.; Janse, J.H.; Mooij, W.M.; Coops, H.; Verhoeven, J.T.A. The Contribution of Marsh Zones to Water Quality in Dutch Shallow Lakes: A Modeling Study. Environ. Manag. 2008, 42, 1002–1016. [Google Scholar] [CrossRef]
- Zhao, J.; Gao, Q.; Liu, Q.; Fu, G. Lake eutrophication recovery trajectories: Some recent findings and challenges ahead. Ecol. Indic. 2020, 110, 105878. [Google Scholar] [CrossRef]
- Fu, Q.; Xiong, Y.; Lu, X. Nitrogen and Phosphorus Pollution Characteristic and Control Measures in Chaihe Reservoir. J. Basic Sci. Eng. 2010, 18, 66–74. [Google Scholar] [CrossRef]
- Rabalais, N.N. Nitrogen in aquatic ecosystems. AMBIO 2002, 31, 102–112. [Google Scholar] [CrossRef] [PubMed]
- de Vries, W. Impacts of nitrogen emissions on ecosystems and human health: A mini review. Curr. Opin. Environ. Sci. Health 2021, 21, 100249. [Google Scholar] [CrossRef]
- Guo, K.; Zhao, W.; Yan, H.-Q. Study on the Spatio-temporal Pattern of Plankton Community Structure and its Productivity in Chaihe Reservoir. J. Hydroecology 2013, 34, 22–26. [Google Scholar] [CrossRef]
- Smith, V.H. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science 1983, 221, 669–671. [Google Scholar] [CrossRef]
- Wilander, A.; Persson, G. Recovery from eutrophication: Experiences of reduced phosphorus input to the four largest lakes of Sweden. AMBIO 2001, 30, 475–485. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TLICW | Trophic Classes |
|---|---|
| ≤ 30 | Oligotrophic |
| 30 <TLICW ≤ 50 | Mesotrophic |
| 50<TLICW ≤ 60 | Light-eutrophic |
| 60<TLICW ≤ 70 | Mesoeutrophic |
| 70<TLICW ≤ 100 | Hypereutrophic |
| The Goal of Chla (μg/L) | 10 | 8 | 6 | 4 | Data |
|---|---|---|---|---|---|
| TPa in the early period (2004–2009) | 25 | 20 | 15 | 10 | For all data |
| TPa in the recent period (2017–2021) | 50 | 40 | 30 | 20 | For all data |
| TPa in the recent period (2017–2021) | 89 | 71 | 53 | 35 | Data after 2004 |
| TNa = 10 TPa (2017–2021) | 890 | 710 | 530 | 350 | Data after 2004 |
| TNb = 30 TPa (2017–2021) | 2670 | 2130 | 1590 | 1050 | Data after 2004 |
| TN threshold | >2700 | >2100 | >1600 | >1000 | Data after 2004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Ding, L.; Liu, Y.; Liu, H.; Xuan, B.; Zhao, J.; Li, Y. Reoligotrophication of a High-Nitrogen Reservoir with Phosphorus Removal and Implications for Management. Water 2022, 14, 2629. https://doi.org/10.3390/w14172629
Zhang X, Ding L, Liu Y, Liu H, Xuan B, Zhao J, Li Y. Reoligotrophication of a High-Nitrogen Reservoir with Phosphorus Removal and Implications for Management. Water. 2022; 14(17):2629. https://doi.org/10.3390/w14172629
Chicago/Turabian StyleZhang, Xiaoyu, Lin Ding, Yue Liu, Haixia Liu, Boyu Xuan, Jian Zhao, and Yuan Li. 2022. "Reoligotrophication of a High-Nitrogen Reservoir with Phosphorus Removal and Implications for Management" Water 14, no. 17: 2629. https://doi.org/10.3390/w14172629