
Embryotoxicity of Polystyrene Microspheres of Different Sizes to the Marine Medaka Oryzias melastigma (McClelland, 1839)




Abstract
1. Introduction
2. Materials and Methods
2.1. Polystyrene Microplastics
2.2. Fish Maintenance
2.3. Experimental Design
2.4. Calculation of Membrane Pore Size
2.5. Determination of Embryonic Membrane Changes
2.6. Accumulation of Polystyrene Microplastics on and Inside Chorion
2.7. Measurement of Mortality, Heartbeat and Hatching Rate of Fish Embryos
2.8. Determination of Reactive Oxygen Species (ROS) Levels
2.9. Determination of Expression Profiles of Target Genes
2.10. Statistics Analysis
3. Results
3.1. Characterization of Polystyrene Microplastics
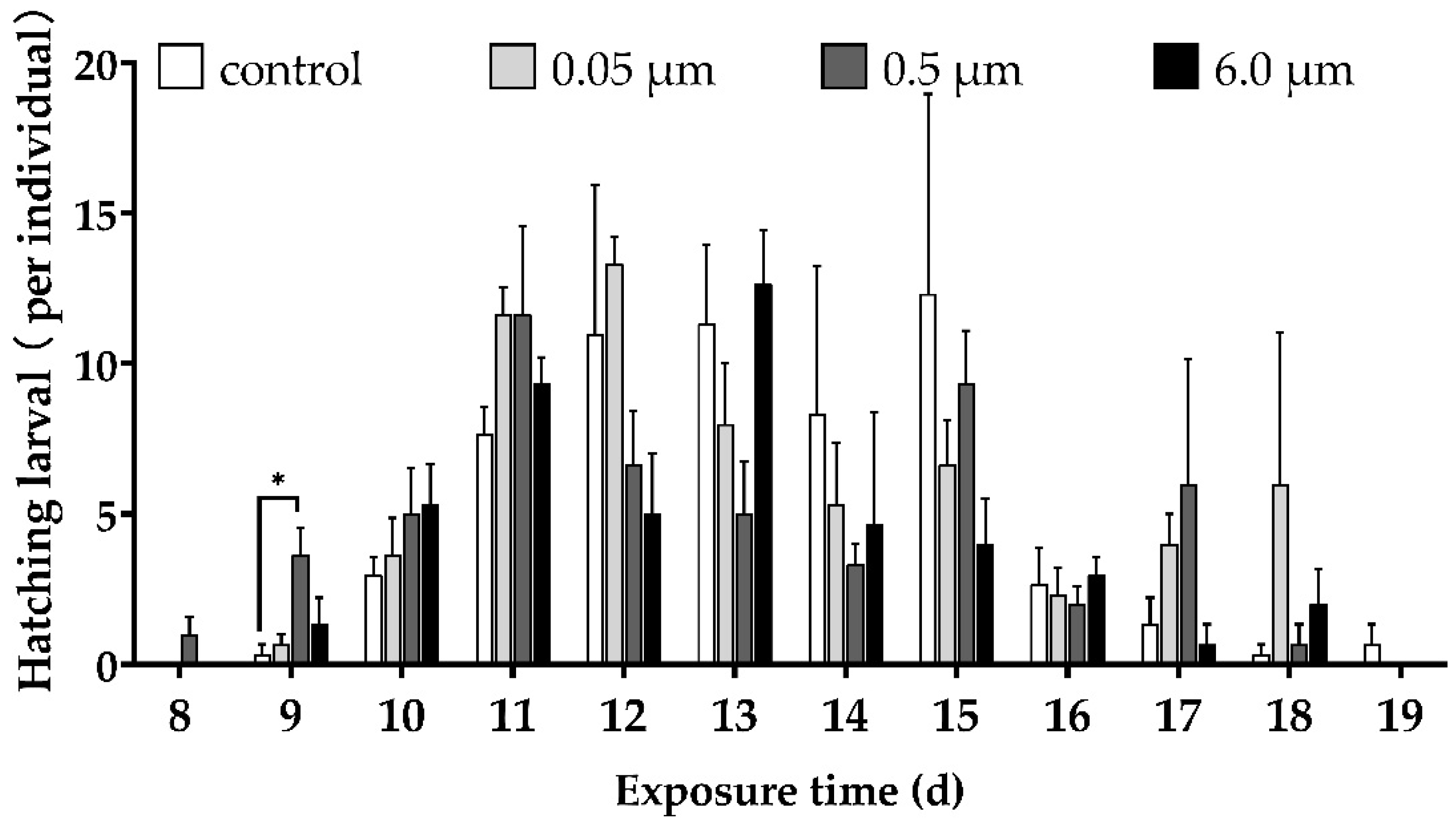
3.2. Hatching Time and Hatching Rate of Embryos
3.3. Embryo Mortality Rate
3.4. Heart Rates
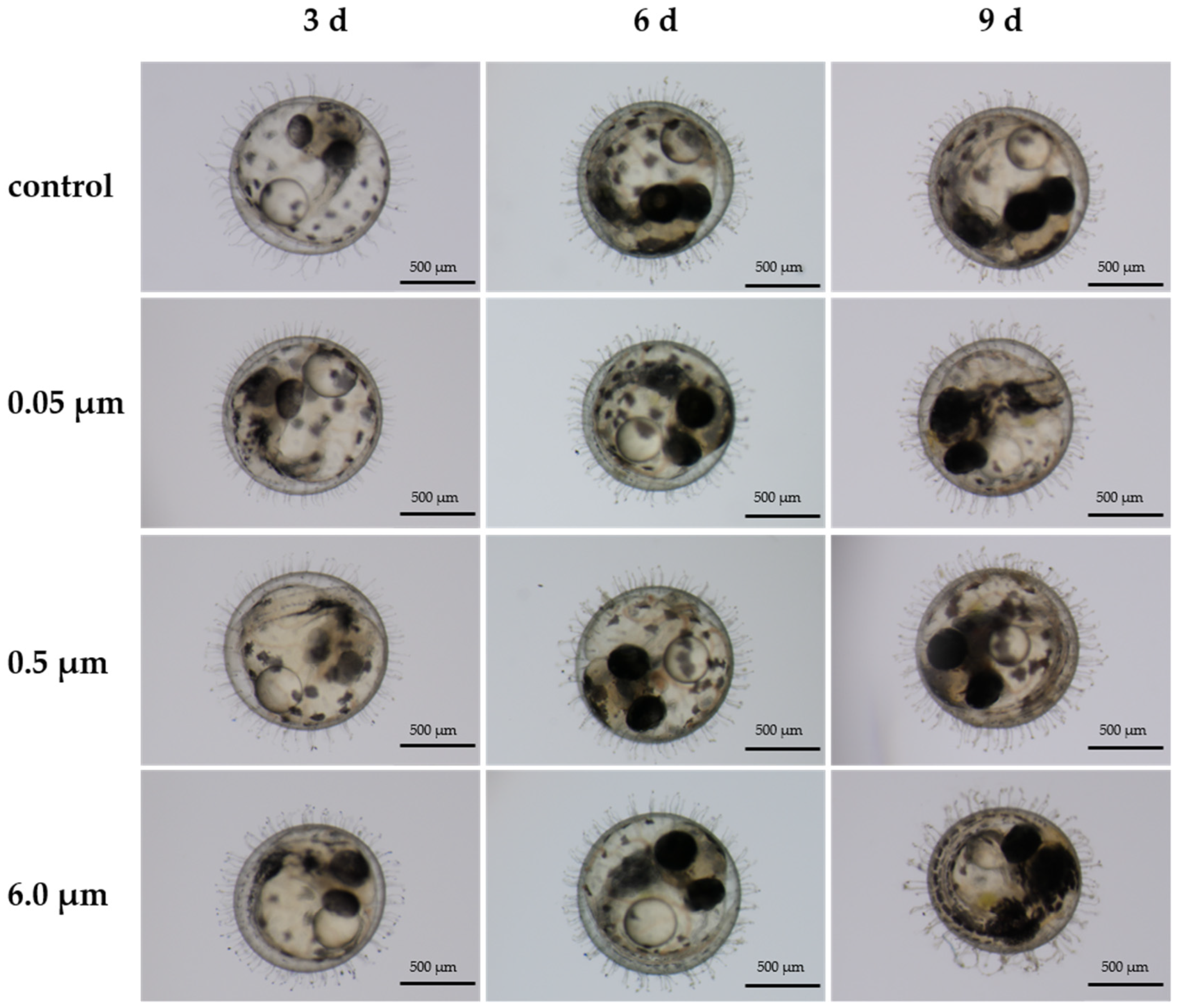
3.5. Embryo Diameter
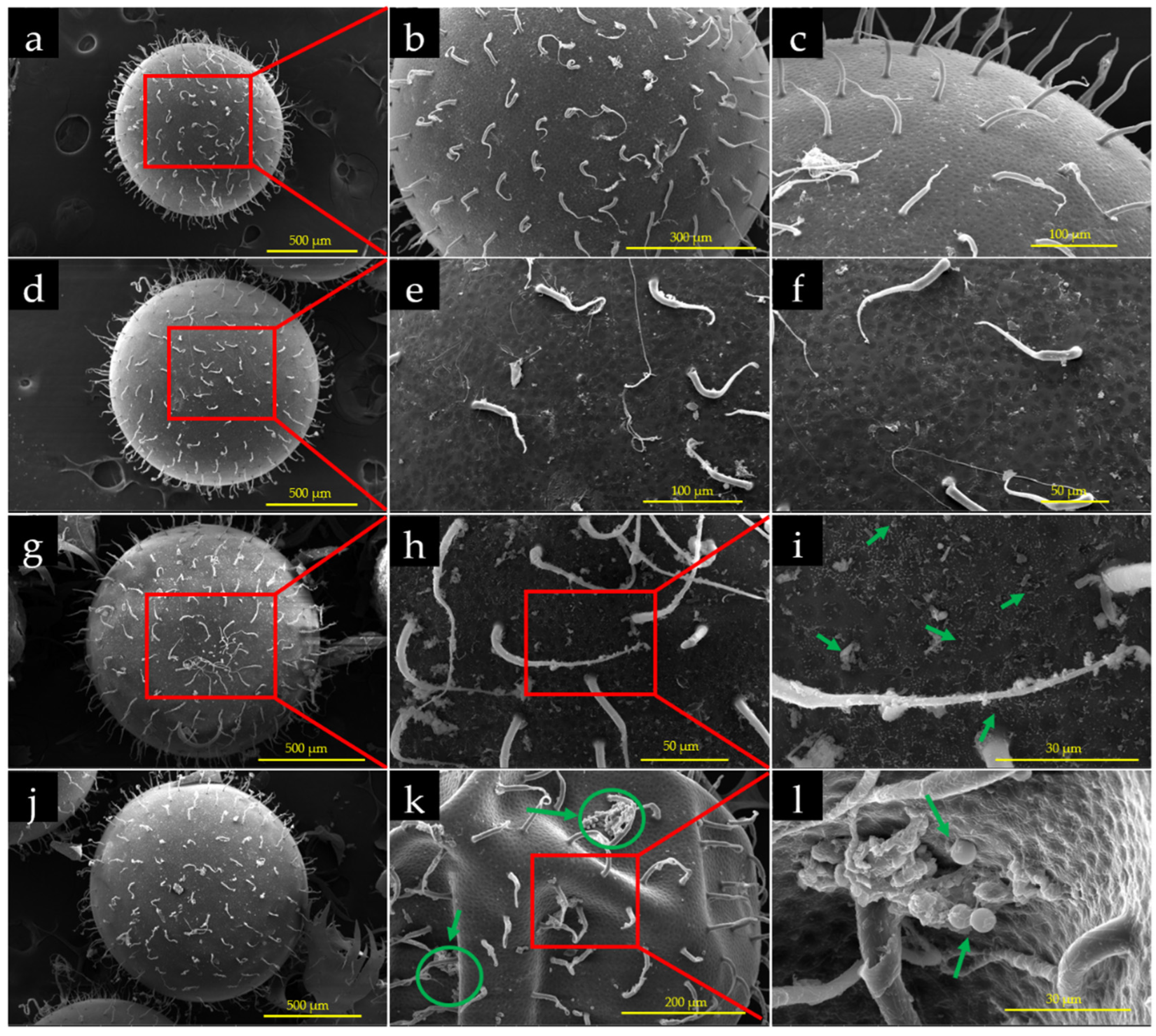
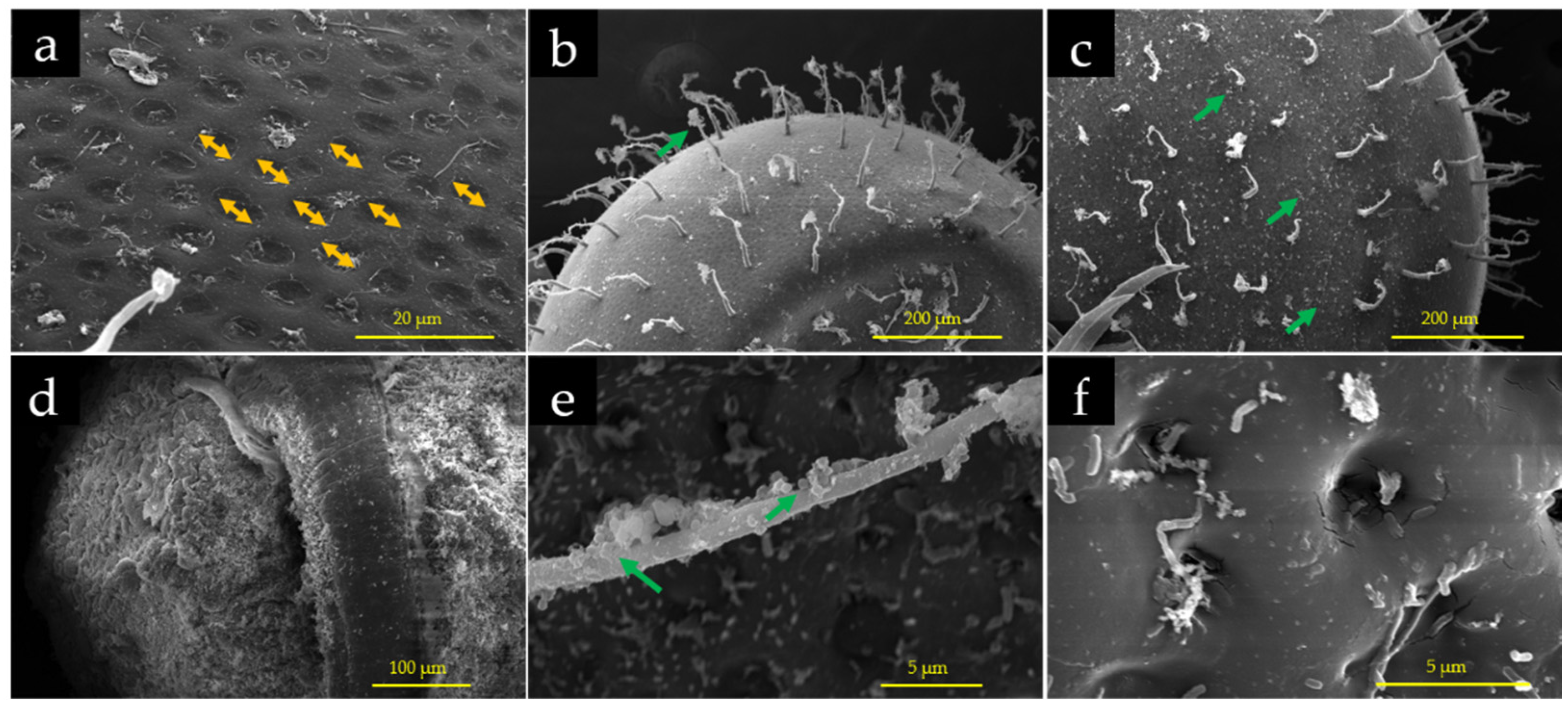
3.6. Effects of PS-MPs on Chorion of the Embryos
3.7. Accumulation of PS-MPs on Chorion of the Embryos
3.8. Accumulation of Fluorescent PS-MPs Inside the Embryos
3.9. Interaction of PS-MPs with Embryos
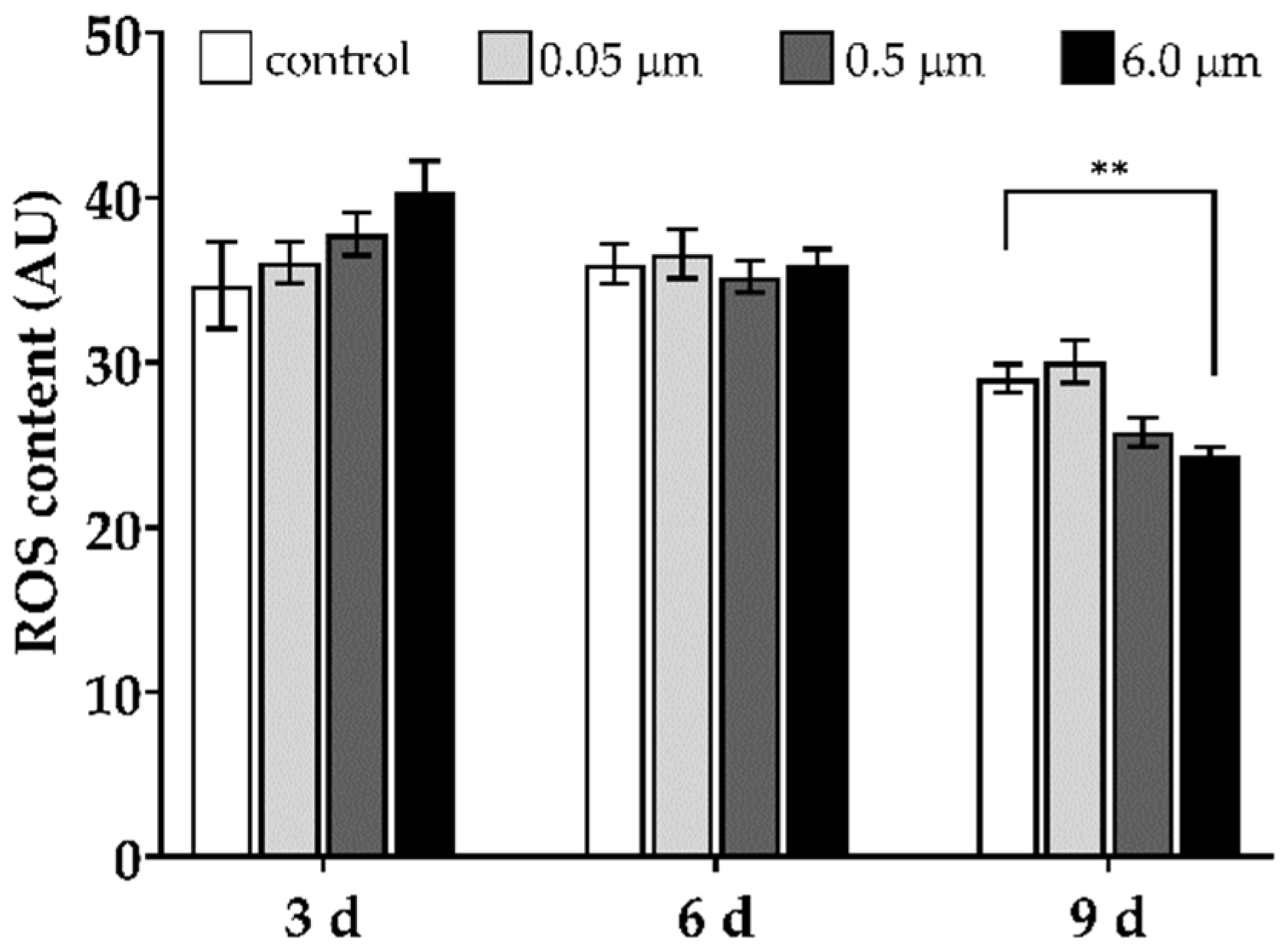
3.10. ROS Production Level
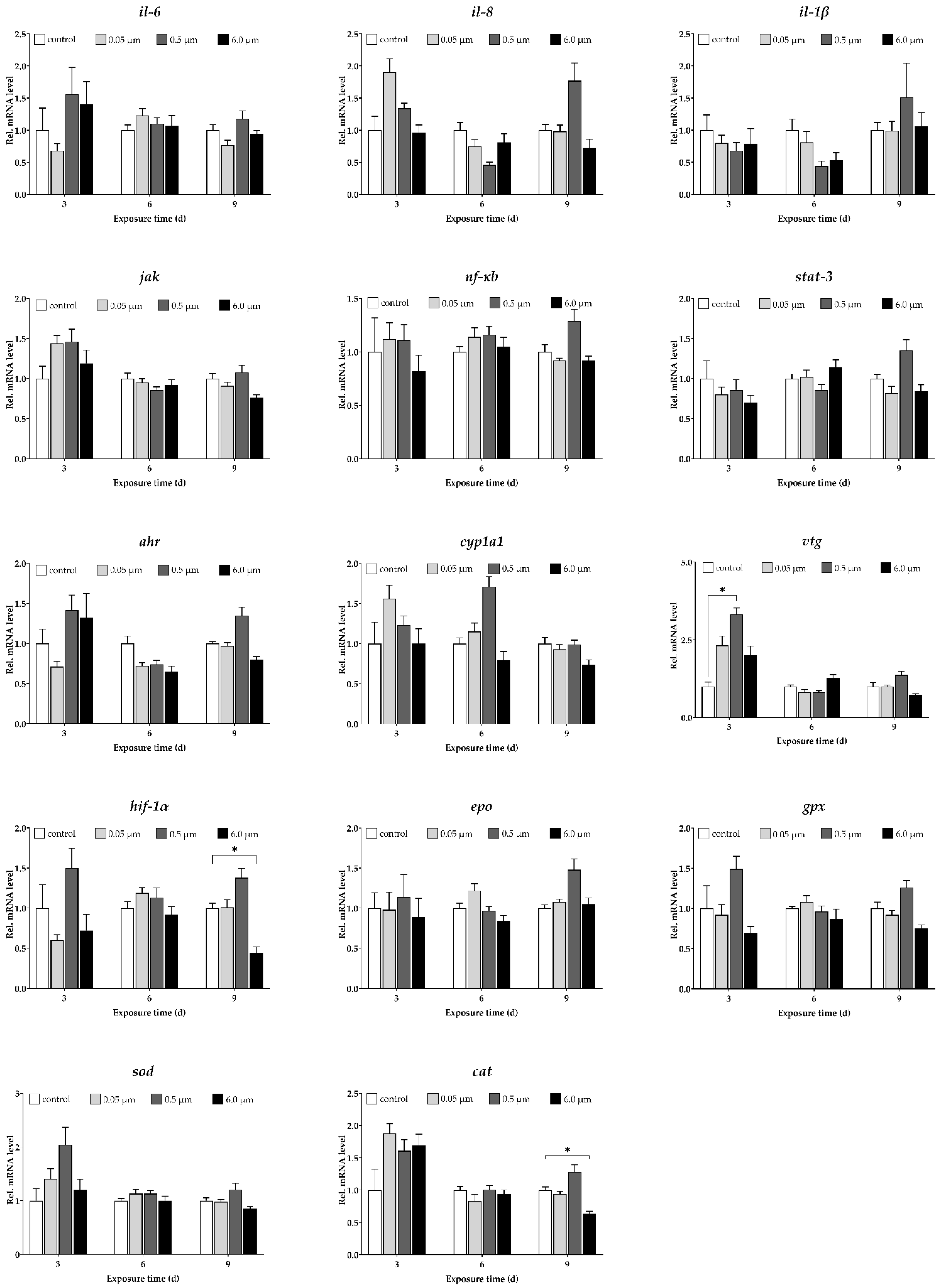
3.11. Expression Profiles of Target Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′ End to 3′ End) |
|---|---|
| 18s | F: GACAAATCGCTCCACCAACT |
| R: CCTGCGGCTTAATTTGACCC | |
| gpx | F: GTGTGCAGAAACGACGTGGCCTGGA |
| R: TCGCCTTCGATGTCGCTGGTGAGGA | |
| il-1β | F: AGGCAGCGACAGCCGCAAAGTTCA |
| R: TGGTGTCCTTGATGCCCAGAGCCA | |
| il-6 | F: GACCTTCTGGCAGGTGACGCTT |
| R: CACCTCTTTCTGGTGCCGTTTGG | |
| il-8 | F: TGCACTGCCGCTGCATCTTGACAG |
| R: TCTGGGTTCAGGCAAACCTCCTGGC | |
| nf-κb | F: CATGGCTACTACGAGGCAGACC |
| R: AACTCCTCCTCCCACACCTTGGAC | |
| jak | F: ATCGCCACCACCTTCCAGCAC |
| R: TGGCAGGATGTCGCTGTCCATG | |
| stat-3 | F: CGCCCTGGAAGAGAAGATTGTGGAT |
| R: CCAGCAACCTGACTTTGTTTGTGAA | |
| epo | F: GACCGGCTTGTGGCTGTT |
| R: CGTGTTGGTGACCGATGGT | |
| vtg | F: CAGGTTGCCAGCTTCACATAC |
| R: CTCAGTCTGTCCAGCTTTGG | |
| hif-1α | F: TGTGATATTTGCAGCATTCCA |
| R: CTACGACAGGAAATCCCATGA | |
| cat | F: GCCAACTACCTGCAGATCCCCGTCA |
| R: AGTTTGGAGCGCCGCCTTGGTTGT | |
| sod | F: TGTACCAGTGCGGGGCCTCACTTCA |
| R: TGCGGTCACATTTCCCAGGTCCCCA | |
| ahr | F: ACTGAACATCCAGGGCAGAC |
| R: ATGTTCTTGGTGCGGATCTC | |
| cyp1a1 | F: TGCTCTGTCTTGGTCAGTGG |
| R: CAGGGTAGGATTTCGGTTCA |
| Treatment | 3d Diameter (mm) | 6d Diameter (mm) | 9d Diameter (mm) |
|---|---|---|---|
| Control | 1.03 ± 0.01 | 1.07 ± 0.01 | 1.06 ± 0.01 |
| 0.05 μm | 1.02 ± 0.01 | 1.06 ± 0.01 | 1.04 ± 0.01 |
| 0.5 μm | 1.01 ± 0.01 | 1.04 ± 0.01 | 1.05 ± 0.01 |
| 6.0 μm | 1.01 ± 0.01 | 1.04 ± 0.01 | 1.05 ± 0.01 |
References
- Frias, J.P.G.L.; Nash, R. Microplastics: Finding a consensus on the definition. Mar. Pollut. Bull. 2019, 138, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Rochman, C.M. Microplastics research-from sink to source. Science 2018, 360, 28–29. [Google Scholar] [CrossRef]
- Thompson, R.C.; Olson, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.G.; McGonigle, D.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef]
- Barros, J.; Seena, S. Plastisphere in freshwaters: An emerging concern. Environ. Pollut. 2021, 290, 118123. [Google Scholar] [CrossRef]
- Ma, H.; Pu, S.; Liu, S.; Bai, Y.; Mandal, S.; Xing, B. Microplastics in aquatic environments: Toxicity to trigger ecological consequences. Environ. Pollut. 2020, 261, 114089. [Google Scholar] [CrossRef]
- Dusaucy, J.; Gateuille, D.; Perrette, Y.; Naffrechoux, E. Microplastic pollution of worldwide lakes. Environ. Pollut. 2021, 284, 117075. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, Y.; Wang, J. Occurrence and ecological impact of microplastics in aquaculture ecosystems. Chemosphere 2021, 274, 129989. [Google Scholar] [CrossRef] [PubMed]
- Campanale, C.; Galafassi, S.; Savino, I.; Massarelli, C.; Ancona, V.; Volta, P.; Uricchio, V.F. Microplastics pollution in the terrestrial environments: Poorly known diffuse sources and implications for plants. Sci. Total Environ. 2022, 805, 150431. [Google Scholar] [CrossRef]
- Zazouli, M.; Nejati, H.; Hashempour, Y.; Dehbandi, R.; Nam, V.T.; Fakhri, Y. Occurrence of microplastics (MPs) in the gastrointestinal tract of fishes: A global systematic review and meta-analysis and meta-regression. Sci. Total Environ. 2022, 815, 152743. [Google Scholar] [CrossRef] [PubMed]
- Reichert, J.; Tirpitz, V.; Anand, R.; Bach, K.; Knopp, J.; Schubert, P.; Wilke, T.; Ziegler, M. Interactive effects of microplastic pollution and heat stress on reef-building corals. Environ. Pollut. 2021, 290, 118010. [Google Scholar] [CrossRef] [PubMed]
- Hamm, T.; Lenz, M. Negative impacts of realistic doses of spherical and irregular microplastics emerged late during a 42 weeks-long exposure experiment with blue mussels. Sci. Total Environ. 2021, 778, 146088. [Google Scholar] [CrossRef]
- Amélineau, F.; Bonnet, D.; Heitz, O.; Mortreux, V.; Harding, A.M.A.; Karnovsky, N.; Walkusz, W.; Fort, J.; Grémillet, D. Microplastic pollution in the Greenland Sea: Background levels and selective contamination of planktivorous diving seabirds. Environ. Pollut. 2016, 219, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Albano, M.; Panarello, G.; Di Paola, D.; Capparucci, F.; Crupi, R.; Gugliandolo, E.; Spanò, N.; Capillo, G.; Savoca, S. The influence of polystyrene microspheres abundance on development and feeding behavior of Artemia salina (Linnaeus, 1758). Appl. Sci. 2021, 11, 3352. [Google Scholar] [CrossRef]
- Albano, M.; Panarello, G.; Di Paola, D.; D’Angelo, G.; Granata, A.; Savoca, S.; Capillo, G. The mauve stinger Pelagia noctiluca (Cnidaria, Scyphozoa) plastics contamination, the Strait of Messina case. Int. J. Environ. Stud. 2021, 78, 977–982. [Google Scholar] [CrossRef]
- Bottari, T.; Savoca, S.; Mancuso, M.; Capillo, G.; GiuseppePanarello, G.; MartinaBonsignore, M.; Crupi, R.; Sanfilippo, M.; D’Urso, L.; Compagnini, G.; et al. Plastics occurrence in the gastrointestinal tract of Zeus faber and Lepidopus caudatus from the Tyrrhenian Sea. Mar. Pollut. Bull. 2019, 146, 408–416. [Google Scholar] [CrossRef]
- Mancuso, M.; Savoca, S.; Bottari, T. First record of microplastics ingestion by European hake MERLUCCIUS MERLUCCIUS from the Tyrrhenian Sicilian coast (Central Mediterranean Sea). J. Fish Biol. 2019, 94, 517–519. [Google Scholar] [CrossRef] [PubMed]
- Capillo, G.; Savoca, S.; Panarello, G.; Mancuso, M.; Branca, C.; Romano, V.; D’Angelo, G.; Bottari, T.; Spanò, N. Quali-quantitative analysis of plastics and synthetic microfibers found in demersal species from southern Tyrrhenian Sea (Central Mediterranean). Mar. Pollut. Bull. 2020, 150, 110596. [Google Scholar] [CrossRef]
- Savoca, S.; Matanović, K.; D’Angelo, G.; Vetri, V.; Anselmo, S.; Bottari, T.; Mancuso, M.; Kužir, S.; Spanò, N.; Capillo, G.; et al. Ingestion of plastic and non-plastic microfibers by farmed gilthead sea bream (Sparus aurata) and common carp (Cyprinus carpio) at different life stages. Sci. Total Environ. 2021, 782, 146851. [Google Scholar] [CrossRef]
- Savoca, S.; Bottari, T.; Fazio, E.; Bonsignore, M.; Mancuso, M.; Luna, G.M.; Romeo, T.; D’Urso, L.; Capillo, G.; Panarello, G.; et al. Plastics occurrence in juveniles of Engraulis encrasicolus and Sardina pilchardus in the Southern Tyrrhenian Sea. Sci. Total Environ. 2020, 718, 137457. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Su, L.; Li, H.; Liang, M.; Shi, H. Assessing the relationship between the abundance and properties of microplastics in water and in mussels. Sci. Total Environ. 2018, 621, 679–686. [Google Scholar] [CrossRef]
- Zhu, C.; Li, D.; Sun, Y.; Zheng, X.; Peng, X.; Zheng, K.; Hu, B.; Luo, X.; Mai, B. Plastic debris in marine birds from an island located in the South China Sea. Mar. Pollut. Bull. 2019, 149, 110566. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, C.; Burton, H. Origins and biological accumulation of small plastic particles in fur seals from Macquarie Island. AMBIO J. Hum. Environ. 2003, 32, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Lusher, A.; Hernandez-Milian, G.; Berrow, S.; Rogan, E.; O’Connor, I. Microplastics and marine mammals: Studies from Ireland. In Fate and Impact of Microplastics in Marine Ecosystems; Baztan, J., Jorgensen, B., Pahl, S., Thompson, R.C., Vanderlinden, J.-P., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; p. 37. ISBN 978-0-12-812271-6. [Google Scholar]
- Charlene, T.; Jeff, S.; Dayanthi, N. Sub-lethal responses to microplastic ingestion in invertebrates. In Particulate Plastics in Terrestrial and Aquatic Environments, 1st ed.; Bolan, N., Kirkham, M.B., Eds.; CRC Press: Boca Raton, FL, USA, 2020; p. 466. ISBN 978-1-00-305307-1. [Google Scholar] [CrossRef]
- Bergami, E.; Bocci, E.; Vannuccini, M.L.; Monopoli, M.; Salvati, A.; Dawson, K.A.; Corsi, I. Nano-sized polystyrene affects feeding, behavior and physiology of brine shrimp Artemia franciscana larvae. Ecotoxicol. Environ. Saf. 2016, 123, 18–25. [Google Scholar] [CrossRef]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.J.; Le Goïc, N.; Quillien, V.; Mingant, C.; Epelboin, Y.; et al. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, B.; Li, Q.; Liu, N.; Xia, B.; Zhu, L.; Qu, K. Toxicities of polystyrene nano- and microplastics toward marine bacterium Halomonas alkaliphila. Sci. Total Environ. 2018, 642, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Chen, B.; Xia, B.; Shi, X.; Qu, K. Polystyrene microplastics alter the behavior, energy reserve and nutritional composition of marine jacopever (Sebastes schlegelii). J. Hazard. Mater. 2018, 360, 97–105. [Google Scholar] [CrossRef]
- Martellini, T.; Guerranti, C.; Scopetani, C.; Ugolini, A.; Chelazzi, D.; Cincinelli, A. A snapshot of microplastics in the coastal areas of the Mediterranean Sea. TrAC Trends Anal. Chem. 2018, 109, 173–179. [Google Scholar] [CrossRef]
- Kedzierski, M.; Palazot, M.; Soccalingame, L.; Falcou-Préfol, M.; Gorsky, G.; Galgani, F.; Bruzaud, S.; Pedrotti, M.L. Chemical composition of microplastics floating on the surface of the Mediterranean Sea. Mar. Pollut. Bull. 2022, 174, 113284. [Google Scholar] [CrossRef]
- Long, M.; Paul-Pont, I.; Hégaret, H.; Moriceau, B.; Lambert, C.; Huvet, A.; Soudant, P. Interactions between polystyrene microplastics and marine phytoplankton lead to species-specific hetero-aggregation. Environ. Pollut. 2017, 228, 454–463. [Google Scholar] [CrossRef]
- Hidalgo-Ruz, V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the marine environment: A review of the methods used for identification and quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef]
- Browne, M.A.; Galloway, T.S.; Thompson, R.C. Spatial patterns of plastic debris along estuarine shorelines. Environ. Sci. Technol. 2010, 44, 3404–3409. [Google Scholar] [CrossRef]
- Barnes, D.K.A.; Galgani, F.; Thompson, R.C.; Barlaz, M. Accumulation and fragmentation of plastic debris in global environments. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Santos, T.; Duarte, A.C. A critical overview of the analytical approaches to the occurrence, the fate and the behavior of microplastics in the environment. TrAC Trends Anal. Chem. 2015, 65, 47–53. [Google Scholar] [CrossRef]
- Jeong, C.B.; Won, E.J.; Kang, H.M.; Lee, M.C.; Hwang, D.S.; Hwang, U.K.; Zhou, B.; Souissi, S.; Lee, S.J.; Lee, J.S. Microplastic size-dependent toxicity, oxidative stress induction, and p-JNK and p-P38 activation in the monogonont rotifer (Brachionus koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef]
- Jin, Y.; Xia, J.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z. Polystyrene microplastics induce microbiota dysbiosis and inflammation in the gut of adult zebrafish. Environ. Pollut. 2018, 235, 322–329. [Google Scholar] [CrossRef]
- Brandts, I.; Teles, M.; Gonçalves, A.P.; Barreto, A.; Franco-Martinez, L.; Tvarijonaviciute, A.; Martins, M.A.; Soares, A.M.V.M.; Tort, L.; Oliveira, M. Effects of nanoplastics on Mytilus galloprovincialis after individual and combined exposure with carbamazepine. Sci. Total Environ. 2018, 643, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Galloway, T.S. The impact of polystyrene microplastics on feeding, function and fecundity in the marine copepod Calanus helgolandicus. Environ. Sci. Technol. 2015, 49, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Suman, T.Y.; Jia, P.P.; Li, W.G.; Junaid, M.; Xin, G.Y.; Wang, Y.; Pei, D.S. Acute and chronic effects of polystyrene microplastics on brine shrimp: First evidence highlighting the molecular mechanism through transcriptome analysis. J. Hazard. Mater. 2020, 400, 123220. [Google Scholar] [CrossRef] [PubMed]
- Alimba, C.G.; Faggio, C. Microplastics in the marine environment: Current trends in environmental pollution and mechanisms of toxicological profile. Environ. Toxicol. Pharmacol. 2019, 68, 61–74. [Google Scholar] [CrossRef]
- Plastics Europe. Plastics—The Facts 2021: An Analysis of European Plastics Production, Demand and Waste Data. Available online: https://plasticseurope.org/knowledge-hub/plastics-the-facts-2021/ (accessed on 2 February 2022).
- Chen, J.C.; Chen, M.Y.; Fang, C.; Zheng, R.H.; Jiang, Y.L.; Zhang, Y.S.; Wang, K.J.; Bailey, C.; Segner, H.; Bo, J. Microplastics negatively impact embryogenesis and modulate the immune response of the marine medaka Oryzias melastigma. Mar. Pollut. Bull. 2020, 158, 111349. [Google Scholar] [CrossRef]
- Rochman, C.M.; Manzano, C.; Hentschel, B.T.; Simonich, S.L.M.; Hoh, E. Polystyrene plastic: A source and sink for polycyclic aromatic hydrocarbons in the marine environment. Environ. Sci. Technol. 2013, 47, 13976–13984. [Google Scholar] [CrossRef]
- Naruse, K. Classification and phylogeny of fishes of the genus Oryzias and its relatives. Fish Biol. J. Medaka 1996, 8, 1–9. [Google Scholar]
- Kim, B.M.; Kim, J.; Choi, I.Y.; Raisuddin, S.; Au, D.W.T.; Leung, K.M.Y.; Wu, R.S.S.; Rhee, J.S.; Lee, J.S. Omics of the marine medaka (Oryzias melastigma) and its relevance to marine environmental research. Mar. Environ. Res. 2016, 113, 141–152. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, B.Y.; Han, J.; Jeong, C.B.; Hwang, D.S.; Lee, M.C.; Kang, H.M.; Kim, D.H.; Lee, D.; Kim, J.; et al. The genome of the marine medaka Oryzias melastigma. Mol. Ecol. Resour. 2018, 18, 656–665. [Google Scholar] [CrossRef]
- Dong, S.; Kang, M.; Wu, X.; Ye, T. Development of a promising fish model (Oryzias melastigma) for assessing multiple responses to stresses in the marine environment. BioMed Res. Int. 2014, 2014, e563131. [Google Scholar] [CrossRef]
- Steer, M.; Cole, M.; Thompson, R.C.; Lindeque, P.K. Microplastic ingestion in fish larvae in the western english channel. Environ. Pollut. 2017, 226, 250–259. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Lu, L.; Zheng, M.; Zhang, X.; Tian, H.; Wang, W.; Ru, S. Polystyrene microplastics cause tissue damages, sex-specific reproductive disruption and transgenerational effects in marine medaka (Oryzias melastigma). Environ. Pollut. 2019, 254, 113024. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Huang, W.; Liu, J.; Ye, Z.; Dou, S. Toxicity of short-term copper exposure to early life stages of red sea bream, Pagrus major. Environ. Toxicol. Chem. 2010, 29, 2044–2052. [Google Scholar] [CrossRef]
- Mohammed, A. Why are early life stages of aquatic organisms more sensitive to toxicants than adults? In New Insights into Toxicity and Drug Testing, 1st ed.; Gowder, S., Ed.; IntechOpen: London, UK, 2013; p. 254. ISBN 978-953-51-0946-4. [Google Scholar] [CrossRef]
- Xia, B.; Sui, Q.; Du, Y.; Wang, L.; Jing, J.; Zhu, L.; Zhao, X.; Sun, X.; Booth, A.M.; Chen, B.; et al. Secondary PVC microplastics are more toxic than primary PVC microplastics to Oryzias melastigma embryos. J. Hazard. Mater. 2022, 424, 127421. [Google Scholar] [CrossRef]
- Li, Y.J.; Yang, G.X.; Wang, J.; Lu, X.; Li, X.; Zheng, Y.Q.; Zhang, Z.Z.; Ru, S.G. Microplastics increase the accumulation of phenanthrene in the ovaries of marine medaka (Oryzias melastigma) and its transgenerational toxicity. J. Hazard. Mater. 2022, 424 Pt D, 127754. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Chen, M.; He, S.; Fang, C.; Chen, M.; Li, D.; Wu, D.; Chernick, M.; Hinton, D.E.; Bo, J.; et al. Microplastics decrease the toxicity of triphenyl phosphate (TPhP) in the marine medaka (Oryzias melastigma) larvae. Sci. Total Environ. 2021, 763, 143040. [Google Scholar] [CrossRef]
- Masato, K.; Kenji, M.; Kiyoshi, N.; Minoru, T. Looking at the medaka embryos. In Medaka: Biology, Management, and Experimental Protocols, 1st ed.; Masato, K., Kenji, M., Kiyoshi, N., Eds.; Wiley-Blackwell: Ames, IA, USA, 2009; Volume 2, pp. 165–180. ISBN 9781119575290. [Google Scholar]
- Li, H.; Chen, H.; Wang, J.; Li, J.; Liu, S.; Tu, J.; Chen, Y.; Zong, Y.; Zhang, P.; Wang, Z.; et al. Influence of microplastics on the growth and the intestinal microbiota composition of brine shrimp. Front. Microbiol. 2021, 12, 2671. [Google Scholar] [CrossRef] [PubMed]
- Phuong, N.N.; Zalouk-Vergnoux, A.; Poirier, L.; Kamari, A.; Châtel, A.; Mouneyrac, C.; Lagarde, F. Is there any consistency between the microplastics found in the field and those used in laboratory experiments? Environ. Pollut. 2016, 211, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Araújo, A.P.; Gomes, A.R.; Malafaia, G. Hepatotoxicity of pristine polyethylene microplastics in neotropical physalaemus Cuvieri tadpoles (Fitzinger, 1826). J. Hazard. Mater. 2020, 386, 121992. [Google Scholar] [CrossRef]
- Doyle, M.J.; Watson, W.; Bowlin, N.M.; Sheavly, S.B. Plastic particles in coastal pelagic ecosystems of the Northeast Pacific Ocean. Mar. Environ. Res. 2011, 71, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Shim, W.J.; Kwon, O.Y.; Kang, J.H. Size-dependent effects of micro polystyrene particles in the marine copepod Tigriopus japonicus. Environ. Sci. Technol. 2013, 47, 11278–11283. [Google Scholar] [CrossRef] [PubMed]
- Bo, J.; Cai, L.; Xu, J.H.; Wang, K.J.; Au, D.W.T. The marine medaka Oryzias melastigma—A potential marine fish model for innate immune study. Mar. Pollut. Bull. 2011, 63, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Bo, J.; Giesy, J.P.; Ye, R.; Wang, K.J.; Lee, J.S.; Au, D.W.T. Identification of differentially expressed genes and quantitative expression of complement genes in the liver of marine medaka Oryzias melastigma challenged with Vibrio parahaemolyticus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2012, 7, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Kong, R.Y.C.; Giesy, J.P.; Wu, R.S.S.; Chen, E.X.H.; Chiang, M.W.L.; Lim, P.L.; Yuen, B.B.H.; Yip, B.W.P.; Mok, H.O.L.; Au, D.W.T. Development of a marine fish model for studying in vivo molecular responses in ecotoxicology. Aquat. Toxicol. 2008, 86, 131–141. [Google Scholar] [CrossRef]
- Chen, X.; Li, L.; Cheng, J.; Chan, L.L.; Wang, D.Z.; Wang, K.J.; Baker, M.E.; Hardiman, G.; Schlenk, D.; Cheng, S.H. Molecular staging of marine medaka: A model organism for marine ecotoxicity study. Mar. Pollut. Bull. 2011, 63, 309–317. [Google Scholar] [CrossRef]
- Jung, G.; Hug, M.; Halter, C.; Friesenhengst, A.; Walzer, J.; Czerny, T. Diffusion of small molecules into medaka embryos improved by electroporation. BMC Biotechnol. 2013, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- Fausto, A.M.; Picchietti, S.; Taddei, A.R.; Zeni, C.; Scapigliati, G.; Mazzini, M.; Abelli, L. Formation of the egg envelope of a teleost, Dicentrarchus Labrax (L.): Immunochemical and cytochemical detection of multiple components. Anat. Embryol. 2004, 208, 43–53. [Google Scholar] [CrossRef]
- Fischer, S.; Klüver, N.; Burkhardt-Medicke, K.; Pietsch, M.; Schmidt, A.; Wellner, P.; Schirmer, K.; Luckenbach, T. Abcb4 acts as multixenobiotix transporter and active barrier against chemical uptake in zebrafish (Danio rerio) embryos. BMC Biol. 2013, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, M.; Chen, X.; Yang, C.; Wu, L. Combined toxicity of microplastics and cadmium on the zebrafish embryos (Danio rerio). Sci. Total Environ. 2020, 743, 140638. [Google Scholar] [CrossRef] [PubMed]
- Hicken, C.E.; Linbo, T.L.; Baldwin, D.H.; Willis, M.L.; Myers, M.S.; Holland, L.; Larsen, M.; Stekoll, M.S.; Rice, S.D.; Collier, T.K.; et al. Sublethal exposure to crude oil during embryonic development alters cardiac morphology and reduces aerobic capacity in adult fish. Proc. Natl. Acad. Sci. USA 2011, 108, 7086–7090. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, M.; Lu, L.; Li, X.; Zhang, Z.; Ru, S. Adaptation of life-history traits and trade-offs in marine medaka (Oryzias melastigma) after whole life-cycle exposure to polystyrene microplastics. J. Hazard. Mater. 2021, 414, 125537. [Google Scholar] [CrossRef]
- Capó, X.; Rubio, M.; Solomando, A.; Alomar, C.; Compa, M.; Sureda, A.; Deudero, S. Microplastic intake and enzymatic responses in Mytilus galloprovincialis reared at the vicinities of an aquaculture station. Chemosphere 2021, 280, 130575. [Google Scholar] [CrossRef]
- Qiao, R.; Sheng, C.; Lu, Y.; Zhang, Y.; Ren, H.; Lemos, B. Microplastics induce intestinal inflammation, oxidative stress, and disorders of metabolome and microbiome in zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Solomando, A.; Capó, X.; Alomar, C.; Compa, M.; Valencia, M.J.; Sureda, A.; Deudero, S. Assessment of the effect of long-term exposure to microplastics and depuration period in Sparus aurata Linnaeus, 1758: Liver and blood biomarkers. Sci. Total Environ. 2021, 786, 147479. [Google Scholar] [CrossRef]
- Sureda, A.; Box, A.; Enseñat, M.; Alou, E.; Tauler, P.; Deudero, S.; Pons, A. Enzymatic antioxidant responses of a labrid fish (Coris julis) liver to environmental caulerpenyne. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 144, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Teles, M.; Fierro-Castro, C.; Na-Phatthalung, P.; Tvarijonaviciute, A.; Trindade, T.; Soares, A.M.V.M.; Tort, L.; Oliveira, M. Assessment of gold nanoparticle effects in a marine teleost (Sparus aurata) using molecular and biochemical biomarkers. Aquat. Toxicol. 2016, 177, 125–135. [Google Scholar] [CrossRef]
- Morcillo, P.; Cordero, H.; Meseguer, J.; Esteban, Á.M.; Cuesta, A. In vitro immunotoxicological effects of heavy metals on European sea bass (Dicentrarchus labrax L.) head-kidney leucocytes. Fish Shellfish Immunol. 2015, 47, 245–254. [Google Scholar] [CrossRef]
- Lu, K.; Qiao, R.; An, H.; Zhang, Y. Influence of microplastics on the accumulation and chronic toxic effects of cadmium in zebrafish (Danio rerio). Chemosphere 2018, 202, 514–520. [Google Scholar] [CrossRef]
- Umamaheswari, S.; Priyadarshinee, S.; Bhattacharjee, M.; Kadirvelu, K.; Ramesh, M. Exposure to polystyrene microplastics induced gene modulated biological responses in zebrafish (Danio rerio). Chemosphere 2020, 281, 128592. [Google Scholar] [CrossRef]
- Yang, H.; Xiong, H.; Mi, K.; Xue, W.; Wei, W.; Zhang, Y. Toxicity comparison of nano-sized and micron-sized microplastics to Goldfish Carassius auratus larvae. J. Hazard. Mater. 2020, 388, 122058. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Xu, W.; Na, J.; Lv, Z.; Zhang, Y. How long-term exposure of environmentally relevant antibiotics may stimulate the growth of Prorocentrum lima: A probable positive factor for red tides. Environ. Pollut. 2019, 255 Pt 1, 113149. [Google Scholar] [CrossRef]
- Karami, A.; Groman, D.B.; Wilson, S.P.; Ismail, P.; Neela, V.K. Biomarker responses in zebrafish (Danio rerio) larvae exposed to pristine low-density polyethylene fragments. Environ. Pollut. 2017, 223, 466–475. [Google Scholar] [CrossRef]
- Incardona, J.P.; Day, H.L.; Collier, T.K.; Scholz, N.L. Developmental toxicity of 4-ring polycyclic aromatic hydrocarbons in zebrafish is differentially dependent on AH receptor isoforms and hepatic cytochrome P4501A metabolism. Toxicol. Appl. Pharmacol. 2006, 217, 308–321. [Google Scholar] [CrossRef]
- Lee, W.S.; Cho, H.J.; Kim, E.; Huh, Y.H.; Kim, H.J.; Kim, B.; Kang, T.; Lee, J.S.; Jeong, J. Bioaccumulation of polystyrene nanoplastics and their effect on the toxicity of Au ions in zebrafish embryos. Nanoscale 2019, 11, 3173–3185. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, K.; Hamazaki, T.S.; Yasumasu, S.; Masuda, K.; Iuchi, I. Molecular and cellular basis of formation, hardening, and breakdown of the egg envelope in fish. Int. Rev. Cytol. 1992, 136, 51–92. [Google Scholar] [CrossRef]
- Herbert, N.A.; Steffensen, J.F. The response of Atlantic cod, Gadus morhua, to progressive hypoxia: Fish swimming speed and physiological stress. Mar. Biol. 2005, 147, 1403–1412. [Google Scholar] [CrossRef]
- Ton, C.; Stamatiou, D.; Liew, C.C. Gene expression profile of zebrafish exposed to hypoxia during development. Physiol. Genom. 2003, 13, 97–106. [Google Scholar] [CrossRef]
- Catron, T.; Mendiola, M.A.; Smith, S.M.; Born, J.; Walker, M.K. Hypoxia regulates avian cardiac Arnt and HIF-1α mRNA expression. Biochem. Biophys. Res. Commun. 2001, 282, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Royer, C.; Lachuer, J.; Crouzoulon, G.; Roux, J.C.; Peyronnet, J.; Mamet, J.; Pequignot, J.M.; Dalmaz, Y. Effects of gestational hypoxia on mRNA levels of Glut3 and Glut4 transporters, Hypoxia inducible factor-1 and thyroid hormone receptors in developing rat brain. Brain Res. 2000, 856, 119–128. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Corà, S.; Bernardini, G.; Gornati, R.; Saroglia, M. Acute and chronic hypoxia affects HIF-1α mRNA levels in sea bass (Dicentrarchus labrax). Aquaculture 2008, 279, 150–159. [Google Scholar] [CrossRef]
- Rahman, M.S.; Thomas, P. Molecular cloning, characterization and expression of two hypoxia-inducible factor alpha subunits, HIF-1alpha and HIF-2alpha, in a hypoxia-tolerant marine teleost, Atlantic croaker (Micropogonias undulatus). Gene 2007, 396, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Thomas, P. Characterization of three IGFBP mRNAs in Atlantic croaker and their regulation during hypoxic stress: Potential mechanisms of their upregulation by hypoxia. Am. J Physiol. Endocrinol. Metab. 2011, 301, E637–E648. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.M.; Chen, E.X.; Kong, R.Y.; Ng, P.K.; Mok, H.O.; Au, D.W. Hypoxia induces telomerase reverse transcriptase (TERT) gene expression in non-tumor fish tissues in vivo: The marine medaka (Oryzias melastigma) model. BMC Mol. Biol. 2006, 7, 27. [Google Scholar] [CrossRef][Green Version]
- Sugawara, T. Screening systems for endocrine disruptors. In Reproductive and Developmental Toxicology, 1st ed.; Gupta, R.C., Ed.; Elsevier Inc.: London, UK, 2011; pp. 893–902. [Google Scholar] [CrossRef]
- Yang, C.Z.; Yaniger, S.I.; Jordan, V.C.; Klein, D.J.; Bittner, G.D. Most plastic products release estrogenic chemicals: A potential health problem that can be solved. Environ. Health Perspect. 2011, 119, 989. [Google Scholar] [CrossRef] [PubMed]
- Guart, A.; Bono-Blay, F.; Borrell, A.; Lacorte, S. Migration of plasticizers phthalates, bisphenol a and alkylphenols from plastic containers and evaluation of risk. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2011, 28, 676–685. [Google Scholar] [CrossRef]
- Rochman, C.M.; Hoh, E.; Kurobe, T.; Teh, S.J. Ingested plastic transfers hazardous chemicals to fish and induces hepatic stress. Sci. Rep. 2013, 3, 3263. [Google Scholar] [CrossRef] [PubMed]
- Mak, C.W.; Ching-Fong Yeung, K.; Chan, K.M. Acute toxic effects of polyethylene microplastic on adult zebrafish. Ecotoxicol. Environ. Saf. 2019, 182, 109442. [Google Scholar] [CrossRef] [PubMed]





| Treatment | Hatching Time (d) | Hatching Rate (%) |
|---|---|---|
| Control | 13.48 ± 0.16 | 66.50 ± 7.93 |
| 0.05 μm | 13.34 ± 0.18 | 61.67 ± 4.06 |
| 0.5 μm | 12.87 ± 0.18 ** | 54.33 ± 1.33 * |
| 6.0 μm | 12.77 ± 0.18 ** | 48.00 ± 3.51 * |
| Treatment | Mortality (%) |
|---|---|
| Control | 29.50 ± 6.98 |
| 0.05 μm | 37.00 ± 3.21 |
| 0.5 μm | 40.33 ± 2.33 |
| 6.0 μm | 45.67 ± 6.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Fang, C.; Zheng, R.; Bo, J. Embryotoxicity of Polystyrene Microspheres of Different Sizes to the Marine Medaka Oryzias melastigma (McClelland, 1839). Water 2022, 14, 1831. https://doi.org/10.3390/w14121831
Chen J, Fang C, Zheng R, Bo J. Embryotoxicity of Polystyrene Microspheres of Different Sizes to the Marine Medaka Oryzias melastigma (McClelland, 1839). Water. 2022; 14(12):1831. https://doi.org/10.3390/w14121831
Chicago/Turabian StyleChen, Jincan, Chao Fang, Ronghui Zheng, and Jun Bo. 2022. "Embryotoxicity of Polystyrene Microspheres of Different Sizes to the Marine Medaka Oryzias melastigma (McClelland, 1839)" Water 14, no. 12: 1831. https://doi.org/10.3390/w14121831
APA StyleChen, J., Fang, C., Zheng, R., & Bo, J. (2022). Embryotoxicity of Polystyrene Microspheres of Different Sizes to the Marine Medaka Oryzias melastigma (McClelland, 1839). Water, 14(12), 1831. https://doi.org/10.3390/w14121831