
Effect of Restoration Actions on Organic Carbon Pools in the Lagoon—Delta Ciénaga Grande de Santa Marta, Colombian Caribbean

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
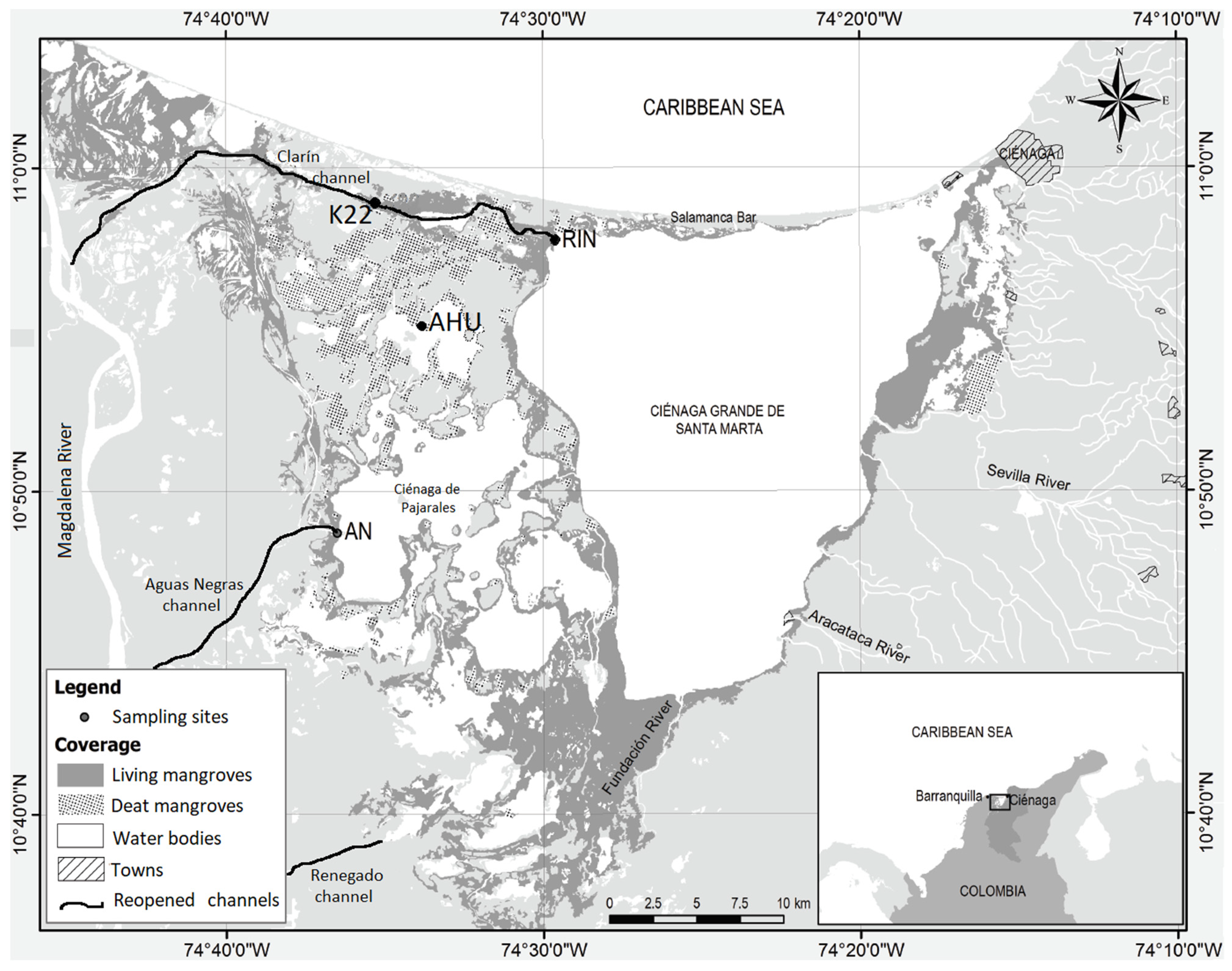
2.1. Study Area
2.2. Ecosystem C Stock
2.2.1. Carbon Stocks in Trees
2.2.2. Root Carbon Stocks
2.2.3. Soil Carbon Stocks
2.3. Statistical Analyses
3. Results
3.1. Carbon Stocks in Trees
3.2. Root Carbon Stocks
3.3. Soil Carbon Stocks
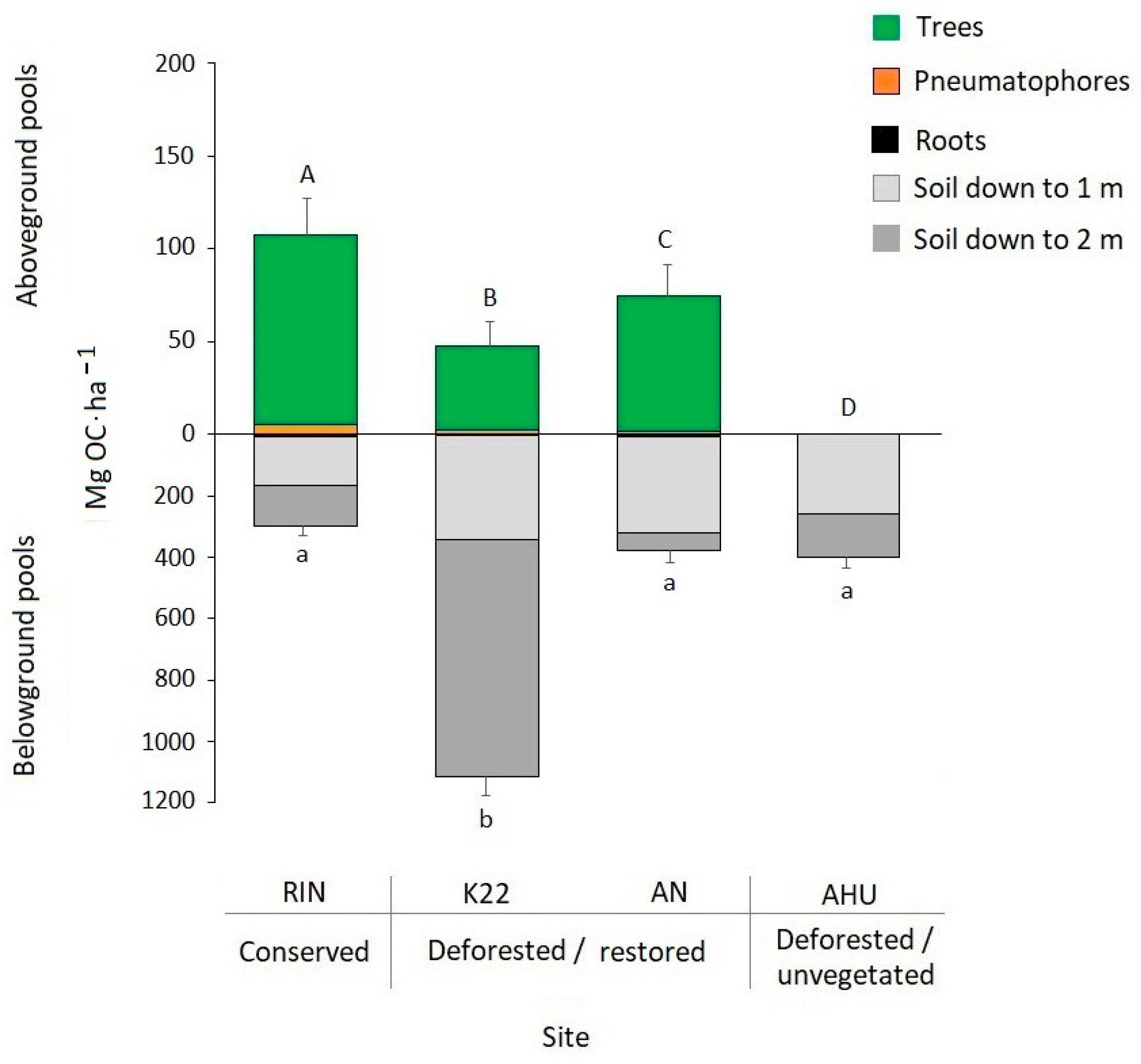
3.4. Mangrove Total Carbon
4. Discussion
4.1. Indirect Estimations of Soil Carbon
4.2. Mangrove Total Carbon
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howard, J.; Hoyt, S.; Isensee, K.; Pidgeon, E.; Telszewski, M. Coastal Blue Carbon: Methods for Assessing Carbon Stocks and Emissions Factors in Mangroves, Tidal Salt Marshes, and Seagrass Meadows; Conservation International; Intergovernmental Oceanographic Commission of UNESCO; International Union for Conservation of Nature: Arlington, VA, USA, 2014. [Google Scholar]
- Lovelock, C.E.; Duarte, C.M. Dimensions of blue carbon and emerging perspectives. Biol. Lett. 2019, 15, 1–5. [Google Scholar] [CrossRef] [PubMed]
- McLeod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Boone, J.K.; Bhomia, R.K. Ecosystem carbon stocks of mangroves across broad environmental gradients in West-Central Africa: Global and regional comparisons. PLoS ONE 2017, 12, e0187749. [Google Scholar] [CrossRef]
- Bindoff, N.L.; Cheung, W.W.L.; Kairo, J.G.; Arístegui, J.; Guinder, V.A.; Hallberg, R.; Hilmi, N.J.M.; Jiao, N.; Karim, M.S.; Levin, L.; et al. Changing Ocean, Marine Ecosystems, and Dependent Communities. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2019; pp. 477–587. [Google Scholar]
- Krauss, K.W.; McKee, K.L.; Lovelock, C.E.; Cahoon, D.R.; Saintilan, N.; Reef, R.; Chen, L. How mangrove forests adjust to rising sea level. New Phytol. 2014, 202, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Núñez, D.A.; Bernal, G.; Pineda, J.E.M. The Relative Role of Mangroves on Wave Erosion Mitigation and Sediment Properties. Estuaries Coasts 2019, 42, 2124–2138. [Google Scholar] [CrossRef]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, biomass, and productivity of mangrove forests: A review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- McKee, K.L. Biophysical controls on accretion and elevation change in Caribbean mangrove ecosystems. Estuar. Coast. Shelf Sci. 2011, 91, 475–483. [Google Scholar] [CrossRef]
- Alongi, D.M. Present state and future of the world’s mangrove forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Worthington, T.; Spalding, M. Mangrove Restoration Potential A global map highlighting a critical opportunity. Geol. Surv. 2018, 36. [Google Scholar]
- Valiela, I.; Bowen, J.L.; York, J. Mangrove Forests: One of the World’s Threatened Major Tropical Environments. Bioscience 2001, 51, 807–815. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.K.; Loveland, T.R.; Masek, J.G.; Duke, N.C. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. The World’s Mangrove Forest 1980–2005; FAO: Rome, Italy, 2007; p. 153. [Google Scholar]
- Hamilton, S.E.; Casey, D. Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Chowdhury, R.R.; Uchida, E.; Chen, L.; Osorio, V.; Yoder, L. Anthropogenic drivers of mangrove loss: Geographic patterns and implications for livelihoods. In Mangrove Ecosystems: A Global Biogeographic Perspective; Springer: Cham, Switzerland, 2017; pp. 275–300. [Google Scholar] [CrossRef]
- Rivera-Monroy, V.H.; Twilley, R.R.; Mancera, J.E.; Alcantara-Eguren, A.; Castañeda-Moya, E.; Casas-Monroy, O.; Reyes, F.; Restrepo, J.; Perdomo Trujillo, L.V.; Campos, E.; et al. Aventuras Y Desventuras En Macondo: Rehabilitación De La Ciénaga Grande De Santa Marta, Colombia. Ecotropicos 2006, 19, 72–93. [Google Scholar]
- Gocke, K.; Pineda, J.E.M.; Vallejo, A. Heterotrophic microbial activity and organic matter degradation in coastal lagoons of Colombia. Rev. Biol. Trop. 2003, 51, 85–98. [Google Scholar]
- Cloern, J.E.; Jassby, A.D. Patterns and scales of phytoplankton variability in estuarine-coastal ecosystems. Estuaries Coasts 2010, 33, 230–241. [Google Scholar] [CrossRef]
- INVEMAR. Monitoreo de las Condiciones Ambientales y los Cambios Estructurales y Funcionales de las Comunidades Vegetales y de los Recursos Pesqueros Durante la Rehabilitación de la Ciénaga Grande De Santa Marta; Informe Técnico Final; INVEMAR: Santa Marta, Colombia, 2018; Volume 17. [Google Scholar]
- Rodríguez-Rodríguez, J.A.; Mancera, J.E.; Trujillo, L.V.P.; Rueda, M.; Ibarra-Gutiérrez, K.P. Ciénaga Grande de Santa Marta: The Largest Lagoon-Delta Ecosystem in the Colombian Caribbean. In The Wetland Book; Finlayson, C., Milton, G., Prentice, R., Davidson, N., Eds.; Springer: Dordrecht, The Netherlands, 2018; pp. 1–16. [Google Scholar]
- Bernal, G. Caracterizacion geomorfologica de la llanura deltaica del Rio Magdalena con enfasis en el sistema lagunar de la Cienaga Grande de Santa Marta, Colombia. Boletín de Investigaciones Marinas y Costeras 1996, 25, 19–48. [Google Scholar] [CrossRef]
- Restrepo, J.D.; Kjerfve, B. Magdalena river: Interannual variability (1975–1995) and revised water discharge and sediment load estimates. J. Hydrol. 2000, 235, 137–149. [Google Scholar] [CrossRef]
- Castaño, A.; Urrego, L.; Bernal, G. Dinámica del manglar en el complejo lagunar de Cispatá (Caribe colombiano) en los últimos 900 años. Rev. Biol. Trop. 2010, 58, 1347–1366. [Google Scholar] [CrossRef][Green Version]
- Botero, L.; Salzwedel, H. Rehabilitation of the Cienaga Grande de Santa Marta, a mangrove-estuarine system in the Caribbean coast of Colombia. Ocean. Coast. Manag. 1999, 42, 243–256. [Google Scholar] [CrossRef]
- Blanco, J.A.; Viloria, E.A.; Narváez, J.C. ENSO and salinity changes in the Ciénaga Grande de Santa Marta coastal lagoon system, Colombian Caribbean. Estuar. Coast. Shelf Sci. 2006, 66, 157–167. [Google Scholar] [CrossRef]
- Álvarez-León, R.; Casas-Monroy, O.; la Hoz, E.C.; Reyes-Forero, S.P.; Troncoso-Olivo, W. La vegetación terrestre, eurihalina y dulceacuícola de la ecorregión Ciénaga Grande de Santa Marta. In Los Manglares de la Ecoregión Ciénaga Grandede Santa Marta: Pasado, Presente y Futuro; Serie de publicaciones especiales No. 11; Garay, J., Restrepo, J., Casas-Monroy, O., Solano, O.D., Newmark, F., Eds.; INVEMAR: Santa Marta, Colombia, 2004; pp. 75–96. [Google Scholar]
- Röderstein, M.; Perdomo, L.; Villamil, C.; Hauffe, T.; Schnetter, M.L. Long-term vegetation changes in a tropical coastal lagoon system after interventions in the hydrological conditions. Aquat. Bot. 2014, 113, 19–31. [Google Scholar] [CrossRef]
- Vilardy, S.; González, J.A. Repensando la Ciénaga: Nuevas Miradas y Estrategias para la Sostenibilidad en la Ciénaga Grande de Santa Marta; Universidad del Magdalena: Santa Marta, Colombia; Universidad Autonoma de Madrid: Madrid, Spain, 2011; Volume 1. [Google Scholar]
- INVEMAR. Monitoreo de las Condiciones Ambientales y los Cambios Estructurales y Funcionales de las Comunidades Vegetales y de los Recursos Pesqueros Durante la Rehabilitación de la Ciénaga Grande de Santa Marta; Informe Técnico Final 2018; INVEMAR: Santa Marta, Colombia, 2019. [Google Scholar]
- Botero, L.; Mancera-Pineda, J.E. Síntesis de los cambios de origen antrópico ocurridos en los últimos 40 años en la Ciénaga de Santa Marta (Colombia). Rev. Acad. Colomb. Cienc. 1996, 20, 465–474. [Google Scholar]
- Poveda, G.; Mesa, O.J. La Corriente de chorro superficial del oeste ‘del Chocó’: Climatología y Variabilidad durante las fases del ENSO. Rev. Acad. Colomb. Cienc. 1999, 23, 517–528. [Google Scholar]
- Giannini, A.; Kushnir, Y.; Cane, M.A. Interannual variability of Caribbean rainfall, ENSO, and the Atlantic Ocean. J. Clim. 2000, 13, 297–311. [Google Scholar] [CrossRef]
- Díaz, L.A.S.; Botero, L.; Cardona, P.; Mancera-Pineda, J.E. Estructura del manglar en el Delta Exterior del río Magdalena-Ciénaga Grande de Santa Marta, una zona tensionada por alteraciones del equilibrio hídrico. Boletín de Investigaciones Marinas y Costeras 1995, 24, 135–164. [Google Scholar]
- Cardona, P.; Botero, L. Soil Characteristics and Vegetation Structure in a Heavily Deteriorated Mangrove Forest in the Caribbean Coast of Colombia. Biotropica 1998, 30, 24–34. [Google Scholar] [CrossRef]
- Gónima, L.; Mancera-Pineda, J.E.; Botero, L. Aplicación de Imágenes de Satélite al Diagnóstico Ambiental de un Complejo Lagunar Estuarino Tropical: Ciénaga Grande de Santa Marta, Caribe Colombiano; INVEMAR: Santa Marta, Colombia, 1998. [Google Scholar]
- Simard, M.; Rivera-Monroy, V.H.; Mancera-Pineda, J.E.; Castañeda-Moya, E.; Twilley, R.R. A systematic method for 3D mapping of mangrove forests based on Shuttle Radar Topography Mission elevation data, ICEsat/GLAS waveforms and field data: Application to Ciénaga Grande de Santa Marta, Colombia. Remote Sens. Environ. 2008, 112, 2131–2144. [Google Scholar] [CrossRef]
- Botero, L.; Marshall, M. Biodiversity Within the Living, Dying and Dead Mangrove Forests of The Ciénaga Grande De Santa Marta; Mote Marine Laboratory Technical Report Number 366; Mote Marine Laboratory: Tallahassee, FL, USA, 1994. [Google Scholar]
- Mancera, J.E.; Vidal, L.A. Florecimiento De Microalgas Relacionado Con Mortandad Masiva De Peces En El Complejo Lagunar Cienaga Grande De Santa Marta, Caribe Colombiano. Bull. Mar. Coast. Res. 1994, 23. [Google Scholar] [CrossRef]
- Ibarra, K.P.; Gómez, M.C.; Viloria, E.A.; Arteaga, E.; Cuadrado, I.; Martínez, M.F.; Nieto, Y.; Rodríguez, J.A.; Licero, L.V.; Perdomo, L.V.; et al. Monitoreo de las Condiciones Ambientales y los Cambios Estructurales y Funcionales de las Comunidades Vegetales y de los Recursos Pesqueros Durante la Rehabilitación de la Ciénaga Grande de Santa Marta; INVEMAR: Santa Marta, Colombia, 2014. [Google Scholar]
- CORPAMAG. Agua Dulce para la Ciénaga Grande de Santa Marta. 2020. Available online: https://www.corpamag.gov.co/index.php/es/homepage/79-contenido-espanol/noticias/786-agua-dulce-para-la-cienaga (accessed on 10 March 2021).
- Cardona, P. Característica de los Suelos del Manglar del Costado Noraccidental de la Ciénaga Grande de Santa Marta (Complejo de Pajares e Isla de Salamanca) y su Relacióncon Algunos Atributos de la Vegetación. Master’s Thesis, Universidad de Antióquia, Antióquia, Colombia, 1991. [Google Scholar]
- Casas-Monroy, O. Patrones de Regeneración Natural del Manglar de la Region de la Ciénaga Grande de Santa Marta, Caribe Colombiano. Ph.D. Thesis, Universidad Jorge Tadeo Lozano, Bogota, Colombia, 1999. [Google Scholar]
- Betancourt-Portela, J.M.; Parra, J.P.; Villamil, C. Emisión de metano y óxido nitroso de los sedimentos de manglar de la ciénaga grande de santa marta, Caribe Colombiano. Boletín de Investigaciones Marinas y Costeras 2013, 42, 131–152. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Donato, D.C. Protocols for the Measurement, Monitoring and Reporting of Structure, Biomass and Carbon Stocks in Mangrove Forests; Working Paper; CIFOR: Bogor, Indonesia, 2012; p. 86. [Google Scholar] [CrossRef]
- Adame, M.F.; Cherian, S.; Reef, R.; Stewart-Koster, B. Mangrove root biomass and the uncertainty of belowground carbon estimations. For. Ecol. Manag. 2017, 403, 52–60. [Google Scholar] [CrossRef]
- Cintrón, G.; Schaeffer-Novelli, Y. Introducción a la Ecología del Manglar; UNESCO: Montevideo, Uruguay, 1983. [Google Scholar]
- Yepes, A.; Zapata, M.; Bolivar, J.; Monsalve, A.; Espinosa, S.M.; Sierra-Correa, P.C.; Sierra, A. Ecuaciones alométricas de biomasa aérea para la estimación de los contenidos de carbono en manglares del Caribe Colombiano. Revista Biología Tropical 2016, 64, 913–926. [Google Scholar] [CrossRef]
- Medina, J.H. Estructura, Asignación de Biomasa y Producción Primaria Neta en Bosques de Manglar de un Complejo Karstico de Origen Carbonatado en una Isla Oceánica; Universidad Nacional de Colombia: Bogota, Colombia, 2016. [Google Scholar]
- Angeles, G.; López–Portillo, J.; Ortega-Escalona, F. Functional anatomy of the secondary xylem of roots of the mangrove Laguncularia racemosa (L.) Gaertn. (Combretaceae). Trees Struct. Funct. 2002, 16, 338–345. [Google Scholar] [CrossRef]
- McKee, K.L.; Faulkner, P.L. Restoration of biogeochemical function in mangrove forests. Restor. Ecol. 2000, 8, 247–259. [Google Scholar] [CrossRef]
- Lukac, M. Fine Root Turnover. In Measuring Roots: An Updated Approach; Mancuso, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 363–373. [Google Scholar]
- Castañeda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H.; Marx, B.D.; Coronado-Molina, C.; Ewe, S.M.L. Patterns of Root Dynamics in Mangrove Forests Along Environmental Gradients in the Florida Coastal Everglades, USA. Ecosystems 2011, 14, 1178–1195. [Google Scholar] [CrossRef]
- Davies, B.E. Loss-on-Ignition as an Estimate of Soil Organic Matter. Soil Sci. Soc. Am. J. 1974, 38, 150–151. [Google Scholar] [CrossRef]
- Torres-Duque, J. Complejidad Estructural Aérea de Bosques de Manglar y su Relación con Contenido de Carbono Azul en Suelos; Universidad Nacional de Colombia: Bogota, Colombia, 2020. [Google Scholar]
- Bhomia, R.K.; Kauffman, J.B.; McFadden, T.N. Ecosystem carbon stocks of mangrove forests along the Pacific and Caribbean coasts of Honduras. Wetl. Ecol. Manag. 2016, 24, 187–201. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon cycling in the world’s mangrove ecosystems revisited: Significance of non-steady state diagenesis and subsurface linkages between the forest floor and the coastal ocean. Forests 2020, 11, 977. [Google Scholar] [CrossRef]
- Bolívar, J.; Yepes, A.; Sierra-Correa, P.C.; Urrego, L.; Moreno, F.; Polanía, J.; Monsalve, A.; Espinosa, S.M.; Herrera, D.; Posada, J.; et al. Contenido de carbono en manglares del Caribe colombiano: Bahía de Cispatá, La Balsa-Tinajones y sectores aledaños del delta estuarino del río Sinú. In Proceedings of the Seminario Nacional de Ciencias y Tecnologías del Mar Libro Memorias—Libro de Memorias, Santa Maria, Colombia, 18–22 October 2015; SENALMAR: Santa Maria, Colombia, 2015; p. 220. Available online: http://www.parquesnacionales.gov.co/portal/wp-content/uploads/2013/08/Memorias-VXI-SENALMAR-COLACMAR-2015.pdf (accessed on 10 March 2021).
- Palacios, M.L.; Cantera, J.R.; Peña, E.J. Carbon stocks in mangrove forests of the Colombian Pacific. Estuar. Coast. Shelf Sci. 2019, 227, 106299. [Google Scholar] [CrossRef]
- Monsalve, A.; Ramírez, G. Caracterización de la Estructura y Contenido de Carbono de los Bosques de Manglar en el Área de Jurisdicción del Consejo Comunitario La Plata, Bahía Málaga, Valle del Cauca; Centro de Investigación en Ecosistemas y Cambio Global, Carbono & Bosques: Medellín, Colombia, 2015. [Google Scholar]
- Senger, D.F.; Hortua, D.A.S.; Engel, S.; Schnurawa, M.; Moosdorf, N.; Gillis, L.G. Impacts of wetland dieback on carbon dynamics: A comparison between intact and degraded mangroves. Sci. Total Environ. 2021, 753, 141817. [Google Scholar] [CrossRef] [PubMed]
- Adame, M.F.; Santini, N.S.; Tovilla, C.; Vázquez-Lule, A.; Castro, L.; Guevara, M. Carbon stocks and soil sequestration rates of tropical riverine wetlands. Biogeosciences 2015, 12, 3805–3818. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Trejo, H.H.; Garcia, M.d.J.; Heider, C.; Contreras, W.M. Carbon stocks of mangroves and losses arising from their conversion to cattle pastures in the Pantanos de Centla, Mexico. Wetl. Ecol. Manag. 2016, 24, 203–216. [Google Scholar] [CrossRef]
- Adame, M.F.; Kauffman, J.B.; Medina, I.; Gamboa, J.N.; Torres, O.; Caamal, J.P.; Reza, M.; Herrera-Silveira, J.A. Carbon Stocks of Tropical Coastal Wetlands within the Karstic Landscape of the Mexican Caribbean. PLoS ONE 2013, 8, e56569. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, J.B.; Heider, C.; Norfolk, J.; Payton, F. Carbon stocks of intact mangroves and carbon emissions arising from their conversion in the Dominican Republic. Ecol. Appl. 2014, 24, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Libreros, J.F.; Ortiz-Acevedo, L.F.; Urrego, L.E. Reservorios de biomasa aérea y de carbono en los manglares del golfo de Urabá (Caribe colombiano). Actualidades Biológicas 2015, 37, 131–141. [Google Scholar] [CrossRef]
- INVEMAR. Monitoreo de las Condiciones Ambientales y los Cambios Estructurales y Funcionales de las Comunidades Vegetales y de los Recursos Pesqueros Durante la Rehabilitación de la Ciénaga Grande de Santa Marta; Informe Técnico Final 2015; INVEMAR: Santa Marta, Colombia, 2016; Volume 14. [Google Scholar]
- Lovelock, C.E.; Fourqurean, J.W.; Morris, J.T. Modeled CO2 emissions from coastal wetland transitions to other land uses: Tidal marshes, mangrove forests, and seagrass beds. Front. Mar. Sci. 2017, 4, 1–11. [Google Scholar] [CrossRef]
- Komiyama, A.; Poungparn, S.; Kato, S. Common allometric equations for estimating the tree weight of mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- World Bank. State and Trends of Carbon Pricing 2019; World Bank: Washington, DC, USA, 2019. [Google Scholar]
- De la Peña, A.; Rojas, C.; de la Peña, M. Valoración Económica del Manglar por el Almacenamiento de Carbono, Ciénaga Grande de Santa Marta. CLIO América 2010, 4, 133–150. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Predominant Species | n | Average DBH (cm) | Maximum DBH (cm) | Density (ind. ha−1) | Basal Area * (m2 ha−1) | Tree Biomass (Mg ha−1) | Tree C ** (Mg C ha−1) |
|---|---|---|---|---|---|---|---|---|
| RIN | A. germinans | 75 | 16.1 ± 1.2 | 40.1 | 937.5 ± 62.5 | 27.1 ± 0.5 | 212.6 ± 3.8 | 102.1 ± 1.8 a |
| K22 | A. germinans L. racemosa | 186 | 11.9 ± 0.4 | 29.9 | 1162.5 ± 41.5 | 15.8 ± 1.7 | 94.3 ± 12.1 | 45.2 ± 5.8 b |
| AN | A. germinans L. racemosa | 361 | 10.8 ± 0.2 | 35.0 | 2256.3 ± 386.9 | 25.6 ± 0.9 | 153.2 ± 11.8 | 73.5 ± 5.6 c |
| AHU | Without vegetation | - | - | - | - | 0 | 0 | 0 d |
| Sites | Pneumatophores * | Belowground Roots ** | ||
|---|---|---|---|---|
| Biomass (Mg ha−1) | Carbon (Mg C ha−1) | Biomass (Mg ha−1) | Carbon (Mg C ha−1) | |
| RIN | 10.0 ± 2.2 | 4.8 ± 1.1 a | 20.4 ± 7.2 | 7.5 ± 2.6 a |
| K22 | 4.4 ± 1.3 | 2.0 ± 0.9 b | 8.4 ± 1.7 | 3.2 ± 0.9 a |
| AN | 2.3 ± 0.5 | 1.1 ± 0.3 b | 17.3 ± 2.5 | 6.7 ± 1.3 a |
| AHU | Without vegetation | Without vegetation | ||
| Sitio | SOC1m * (Mg ha−1) | SOC2m ** (Mg ha−1) | DT (m) |
|---|---|---|---|
| RIN | 156.4 ± 15.6 a | 288.5 ± 14.6 a | 4.4 ± 0.6 |
| K22 | 339.2 ± 19.0 b | 1110.3 ± 32.1 b | 3.1 ± 0.4 |
| AN | 312.4 ± 12.8 bc | 373.0 ± 13.4 a | 2.2 ± 0.2 |
| AHU | 257.9 ± 2.0 c | 398.2 ± 19.0 a | 4.0 ± 0.2 |
| Site | n | Coefficient of Determination r2 | Model | Source |
|---|---|---|---|---|
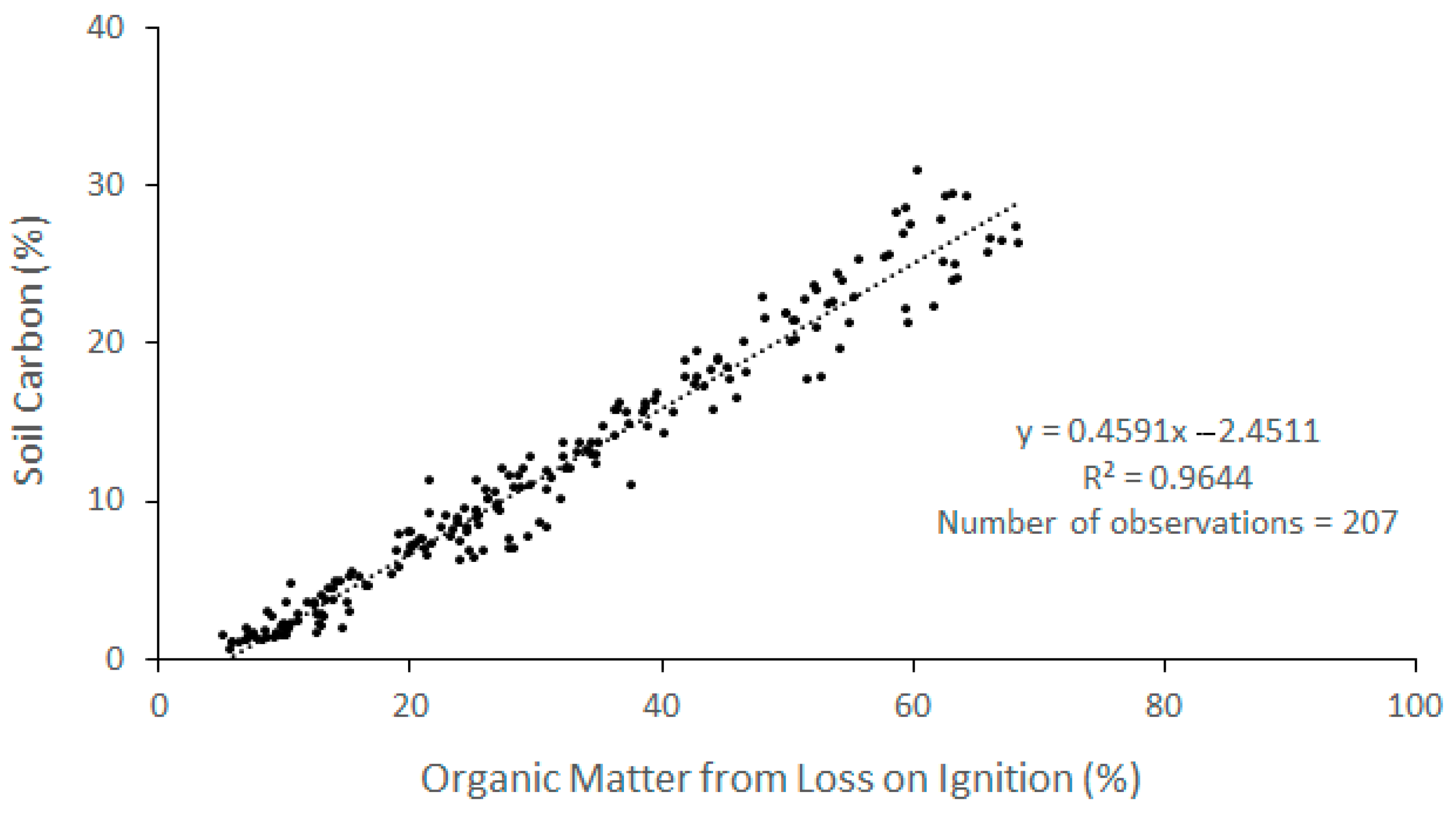
| CGSM 1 | 207 | 0.9644 | % SC = 0.4591 · % OM (LOI) − 2.4511 | This study |
| Republic of Palau 2 | * | 0.5916 | % SC = 0.415 · % OM (LOI) + 2.8857 | [46] |
| Site | n | Coefficient of Determination r2 | Model | Source |
|---|---|---|---|---|
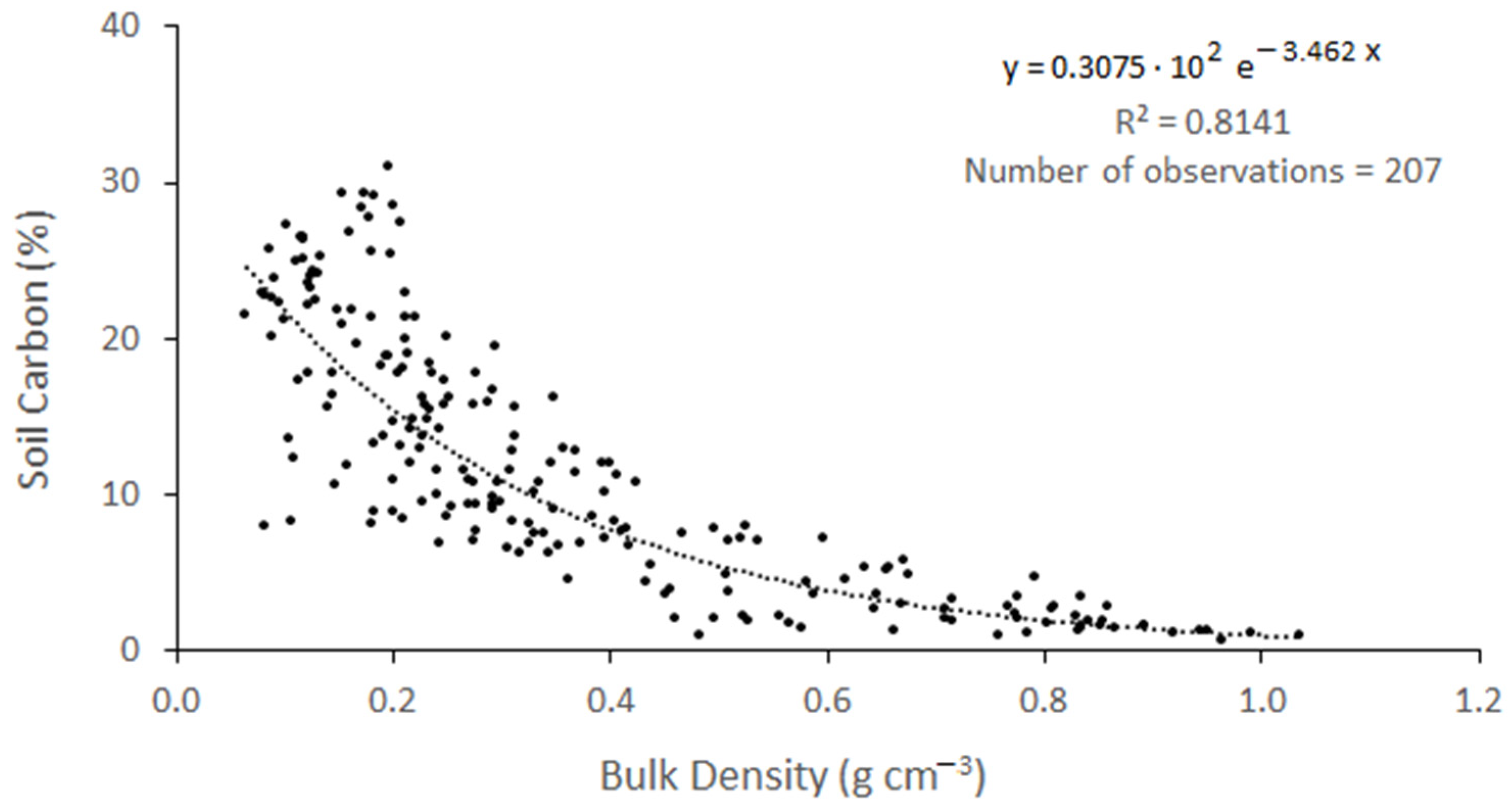
| CGSM 1 | 207 | −0.80 | % C = 0.3075·102 e−3.462(BD) | This study |
| Honduras Pacific and Caribbean coast 2 | 518 | −0.83 | % C = 1.01 − 16.12 ln (BD) | [57] |
| Site | Ecosystem Type/Forest | Predominant Species | Mg C ha−1 | Source |
|---|---|---|---|---|
| CGSM—RIN | Basin, conserved | A. germinans | 403.0 ± 77.1 1 | Present study |
| CGSM—K22 | Basin, around 22 years of recovery | A. germinans L. racemosa | 1160.8 ± 130.9 1 | Present study |
| CGSM—AN | Basin, around 20 years of recovery | A. germinans L. racemosa | 454.5 ± 81.9 1 | Present study |
| CGSM—AHU | Basin, without vegetation recovery | - | 398.2 ± 76.7 1 | Present study |
| Colombia—Caribbean Cispatá—La Balsa—Tinajones | Basin and fringe | A. germinans R. mangle L. racemosa | 521.3 2 | [59] |
| Colombia—Pacific Bahía Málaga and Bahía de Buenaventura | Riverine and fringe | Rhizophora spp. Mora oleífera Pelliciera rhizophorae | 500–710 3 * | [60] |
| Colombia—Pacific Bahía Málaga | Riverine and fringe | Rhizophora spp. P. rhizophorae M. oleífera | 220.2 4 | [61] |
| Bonaire—Caribbean | Coastal lagoon Degraded—Intact | R. mangle A. germinans L. racemosa | 60–140 5 | [62] |
| Honduras, Pacific coast, Caribbean cost and Bay Islands | Low: <3 m medium 3−10 m Tall: >10 m | R. mangle L. racemosa | 306–1632 6 | [57] |
| México—Pacific | Riverine | R. mangle A. germinans L. racemosa | 536–1114 7 | [63] |
| México—Caribbean Pantanos de Centla | Riverine and fringe | R. mangle A. germinans L. racemosa C. erectus | 342–2098 8 | [64] |
| México—Caribbean Península de Yucatán | Dwarf: <1.5 m Medium: 3−5 m Tall >5 m | R. mangle | 297–1325 9 | [65] |
| Dominican Republic, Monte Cristi | Small: <3 m Medium: 3–10 m Tall: >10 m | R. mangle | 706–1131 10 | [66] |
| Site | History | Potentials Emissions * (Mg CO2-equivalents. ha−1) | Carbon Value (USD ha−1) |
|---|---|---|---|
| RIN | Conserved forest | 1451.0 | $7371 |
| K22 | Naturally recovered site after the Clarín channel opening (1996) | 4179.2 | $21,230 |
| AN | Naturally recovered site after the Aguas Negras channel opening (1998) | 1636.3 | $8312 |
| AHU | Degraded—Without vegetation | 1433.7 | $7283 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perdomo-Trujillo, L.V.; Mancera-Pineda, J.E.; Medina-Calderón, J.H.; Sánchez-Núñez, D.A.; Schnetter, M.-L. Effect of Restoration Actions on Organic Carbon Pools in the Lagoon—Delta Ciénaga Grande de Santa Marta, Colombian Caribbean. Water 2021, 13, 1297. https://doi.org/10.3390/w13091297
Perdomo-Trujillo LV, Mancera-Pineda JE, Medina-Calderón JH, Sánchez-Núñez DA, Schnetter M-L. Effect of Restoration Actions on Organic Carbon Pools in the Lagoon—Delta Ciénaga Grande de Santa Marta, Colombian Caribbean. Water. 2021; 13(9):1297. https://doi.org/10.3390/w13091297
Chicago/Turabian StylePerdomo-Trujillo, Laura Victoria, Jose Ernesto Mancera-Pineda, Jairo Humberto Medina-Calderón, David Alejandro Sánchez-Núñez, and Marie-Luise Schnetter. 2021. "Effect of Restoration Actions on Organic Carbon Pools in the Lagoon—Delta Ciénaga Grande de Santa Marta, Colombian Caribbean" Water 13, no. 9: 1297. https://doi.org/10.3390/w13091297
APA StylePerdomo-Trujillo, L. V., Mancera-Pineda, J. E., Medina-Calderón, J. H., Sánchez-Núñez, D. A., & Schnetter, M.-L. (2021). Effect of Restoration Actions on Organic Carbon Pools in the Lagoon—Delta Ciénaga Grande de Santa Marta, Colombian Caribbean. Water, 13(9), 1297. https://doi.org/10.3390/w13091297