
Who’s Next? Non-Indigenous Cnidarian and Ctenophoran Species Approaching to the Italian Waters



Abstract
:1. Introduction
2. Materials and Methods
- –
- establishment success in the Mediterranean Sea according to Zenetos et al. [15]: established (widely recorded at some sites), invasive (able to disseminate from their area of initial introduction), casual (few records), cryptogenic (species with no definite evidence of their native or introduced status), questionable (species with taxonomic status unresolved), unknown;
- –
- first record in the Mediterranean Sea (date, locality, reference: the date and location of the first observation) of each species in the Mediterranean Sea were extracted from the literature; if possible, the actual date of first record was reported, along with its publication date;
- –
- native distribution range: following the global marine biogeographic realms proposed by Spalding et al. [42];
- –
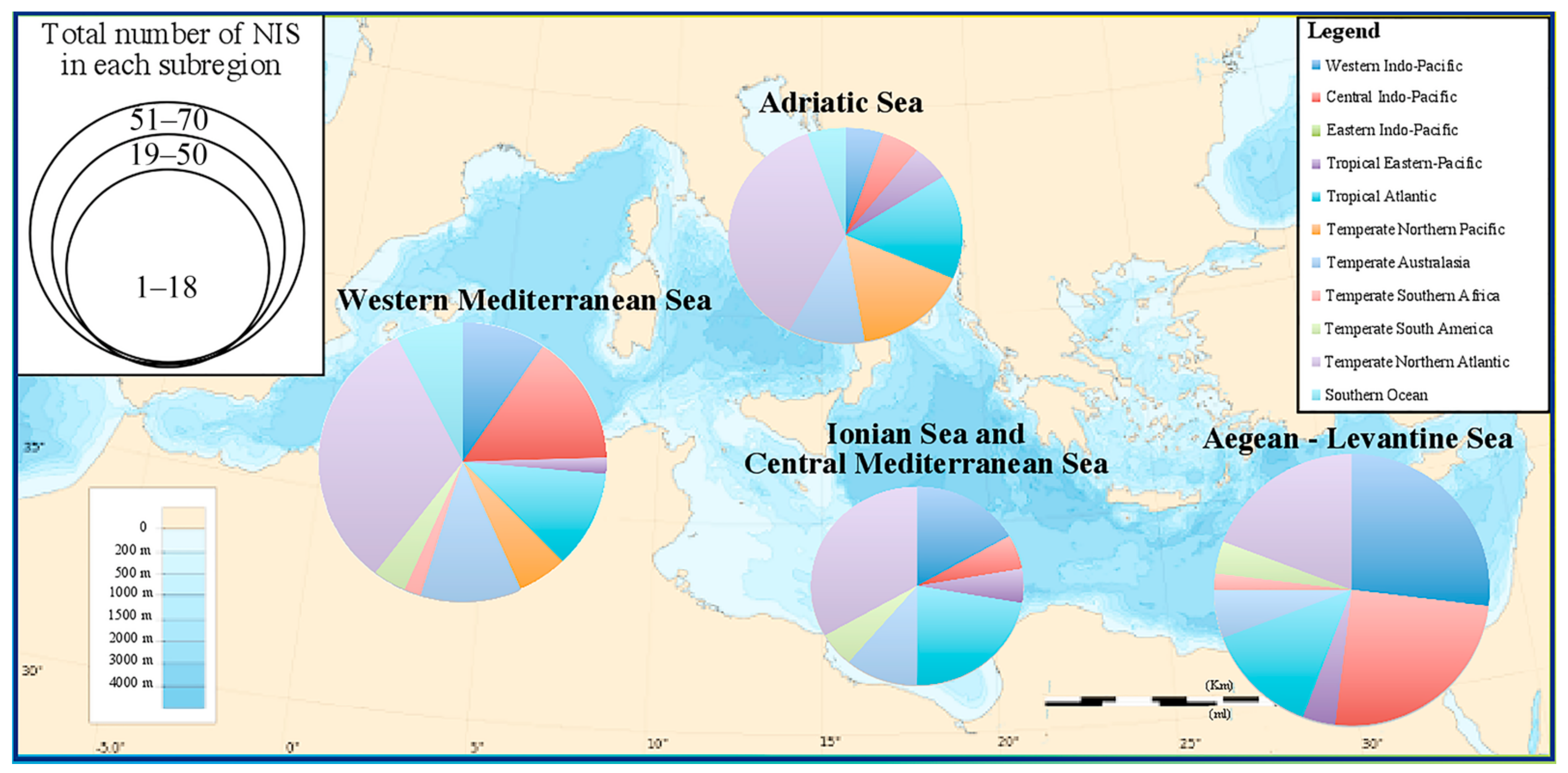
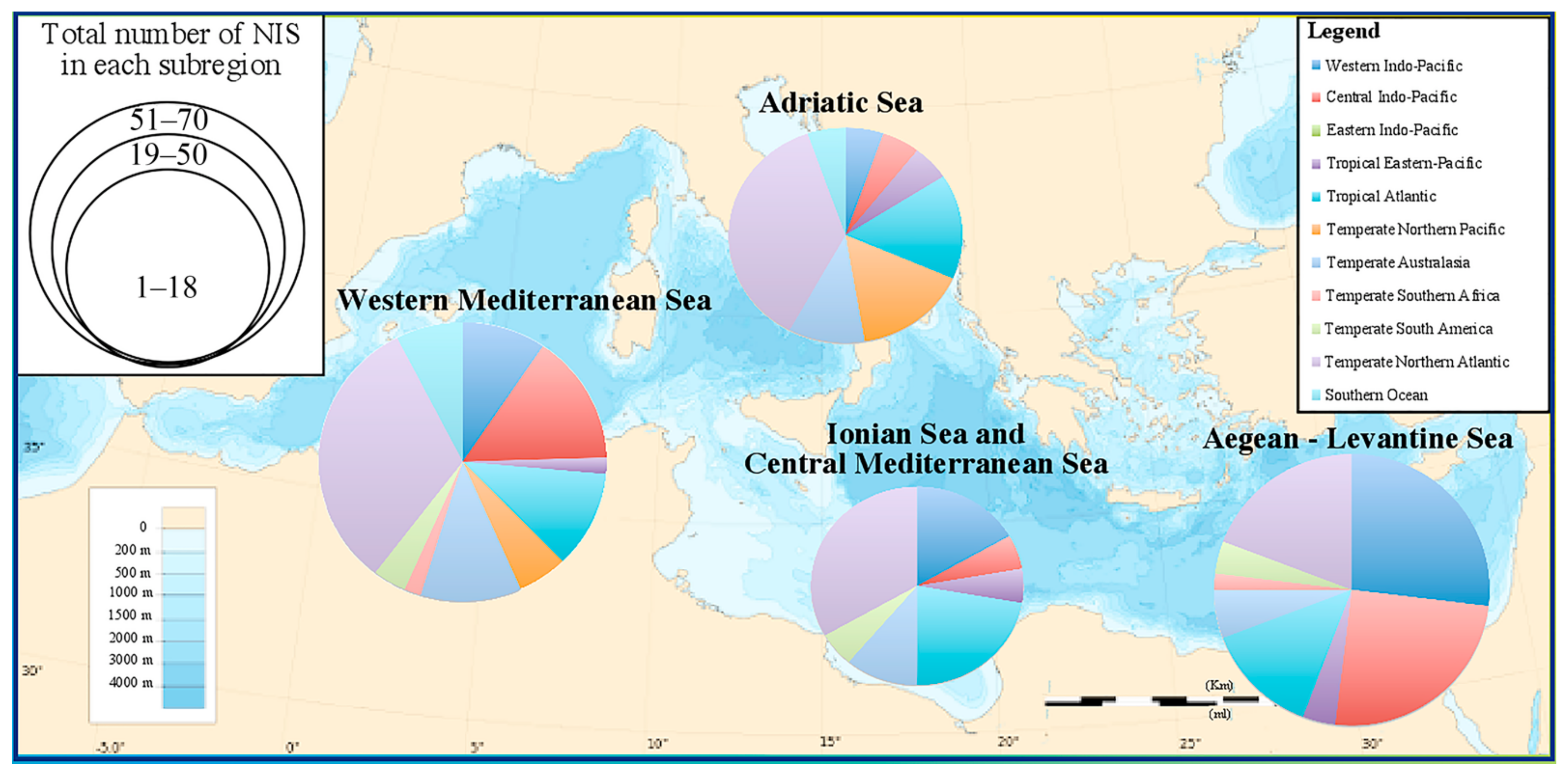
- distribution per Mediterranean subregion (following MSFD, Marine Strategy Framework Directive, 2017) [43]: Aegean-Levantine Sea, Ionian Sea and the Central Mediterranean Sea, Adriatic Sea, Western Mediterranean Sea;
- –
- extra-Mediterranean distribution;
- –
- the taxonomic position of each species (Class, Order, Family);
- –
- the primary pathway(s) of introduction in Europe based on CBD [44]: indicating the frequency if low (L), medium (M) or high (H);
- –
- possible notes;
- –
- code of the main pathway(s) of introduction: A (aquaculture), G (range expansion through the Strait of Gibraltar), Lp (Lessepsian), S (shipping), U (unknown).
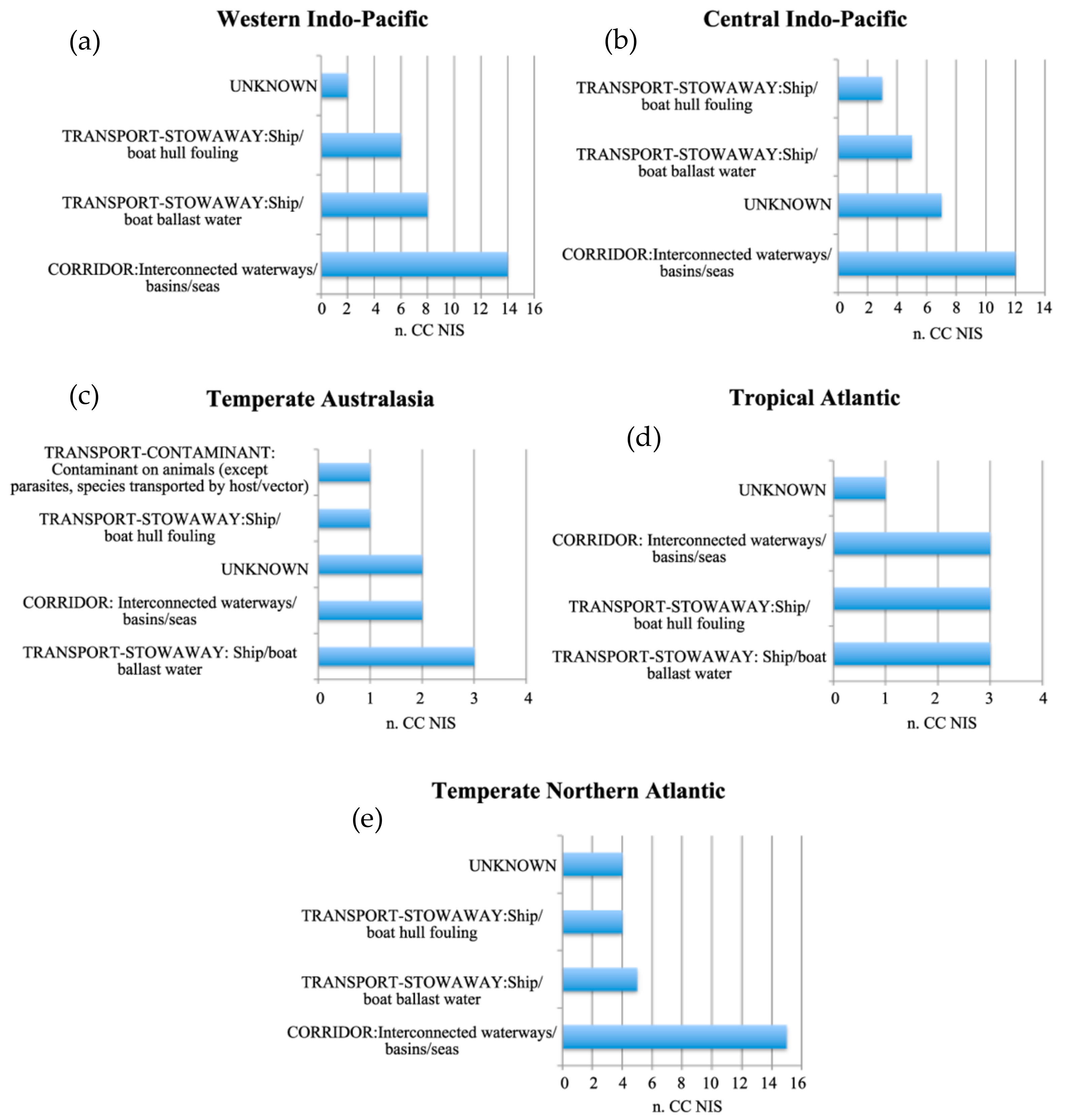
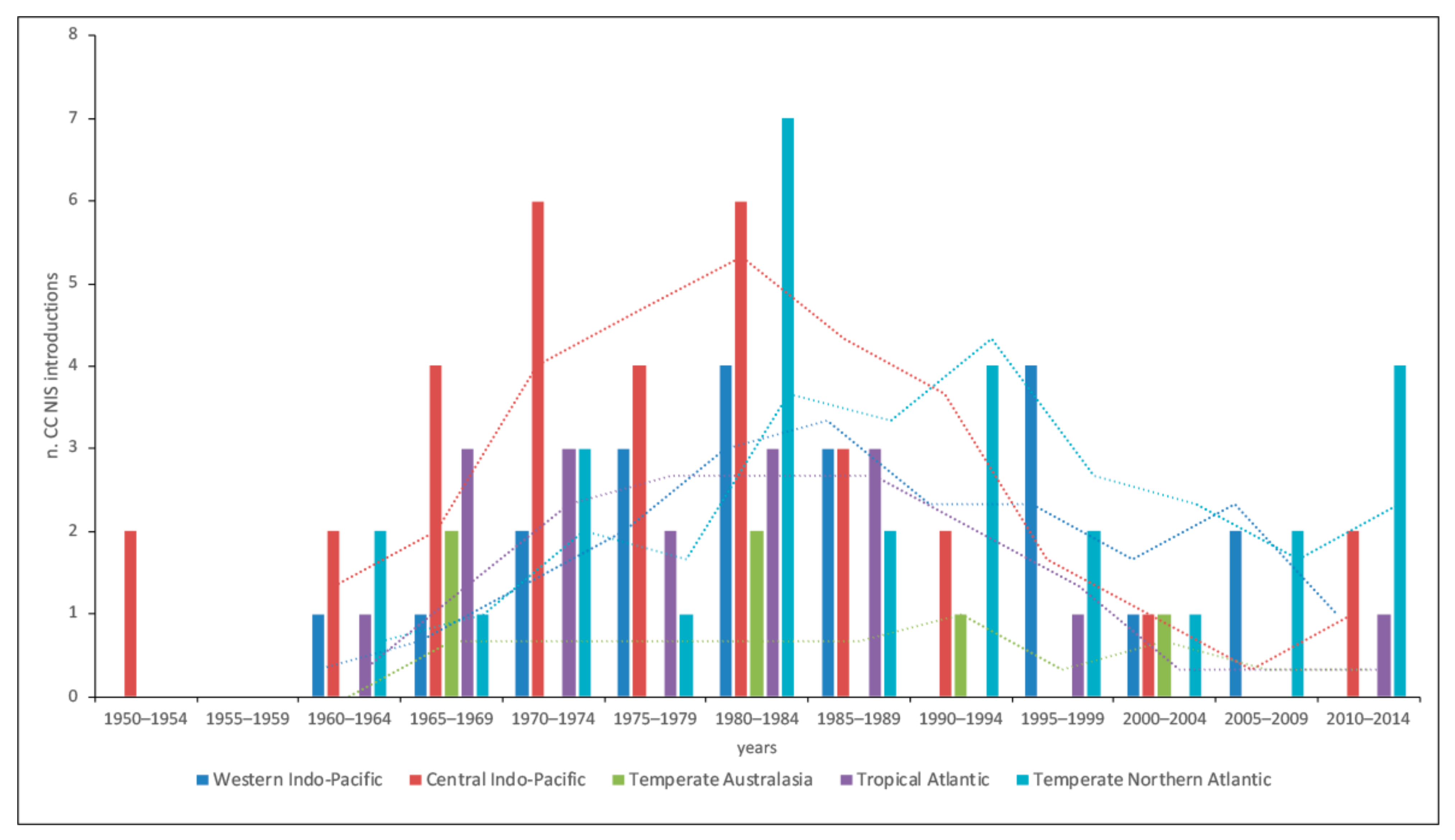
3. Results
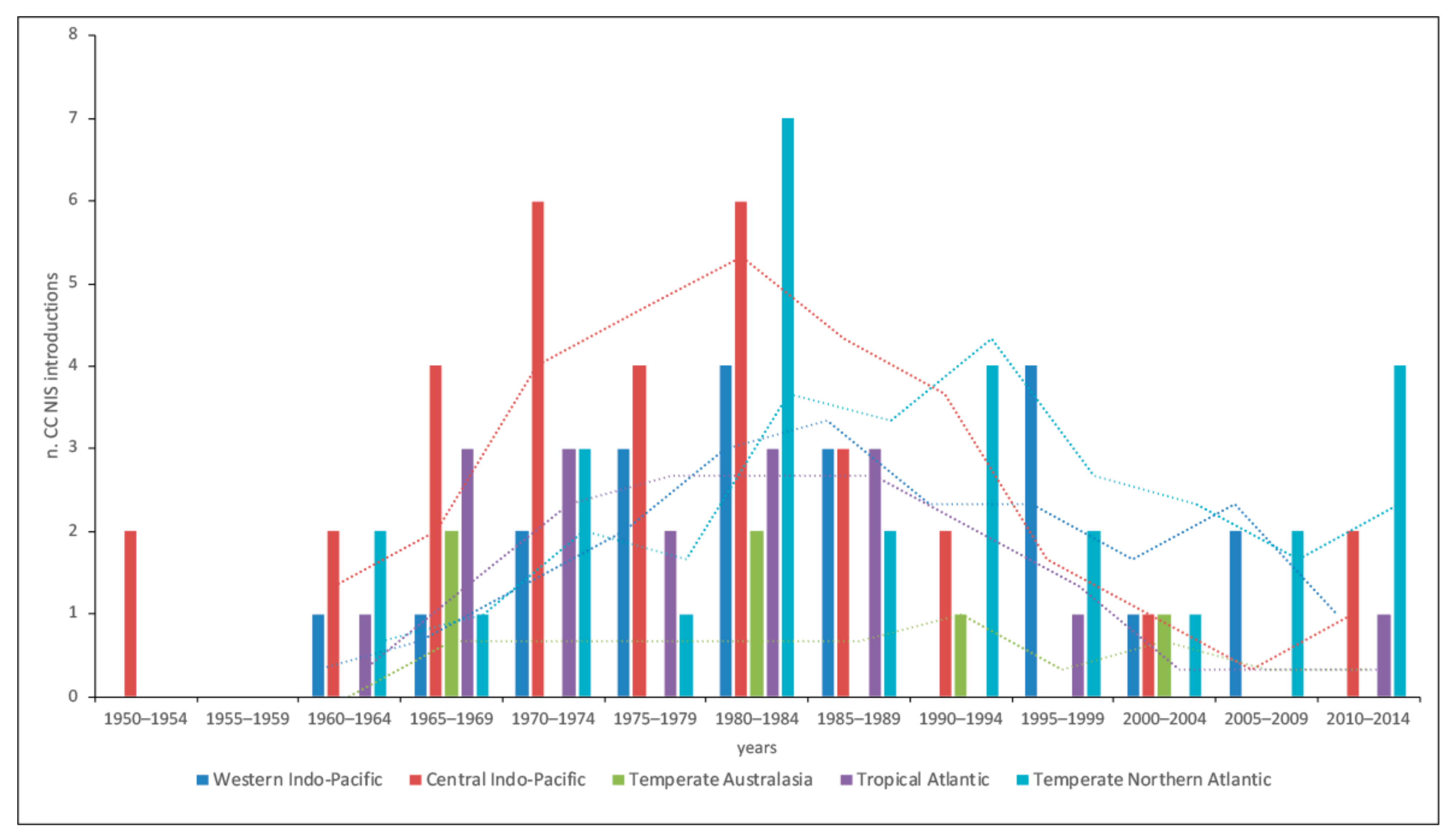
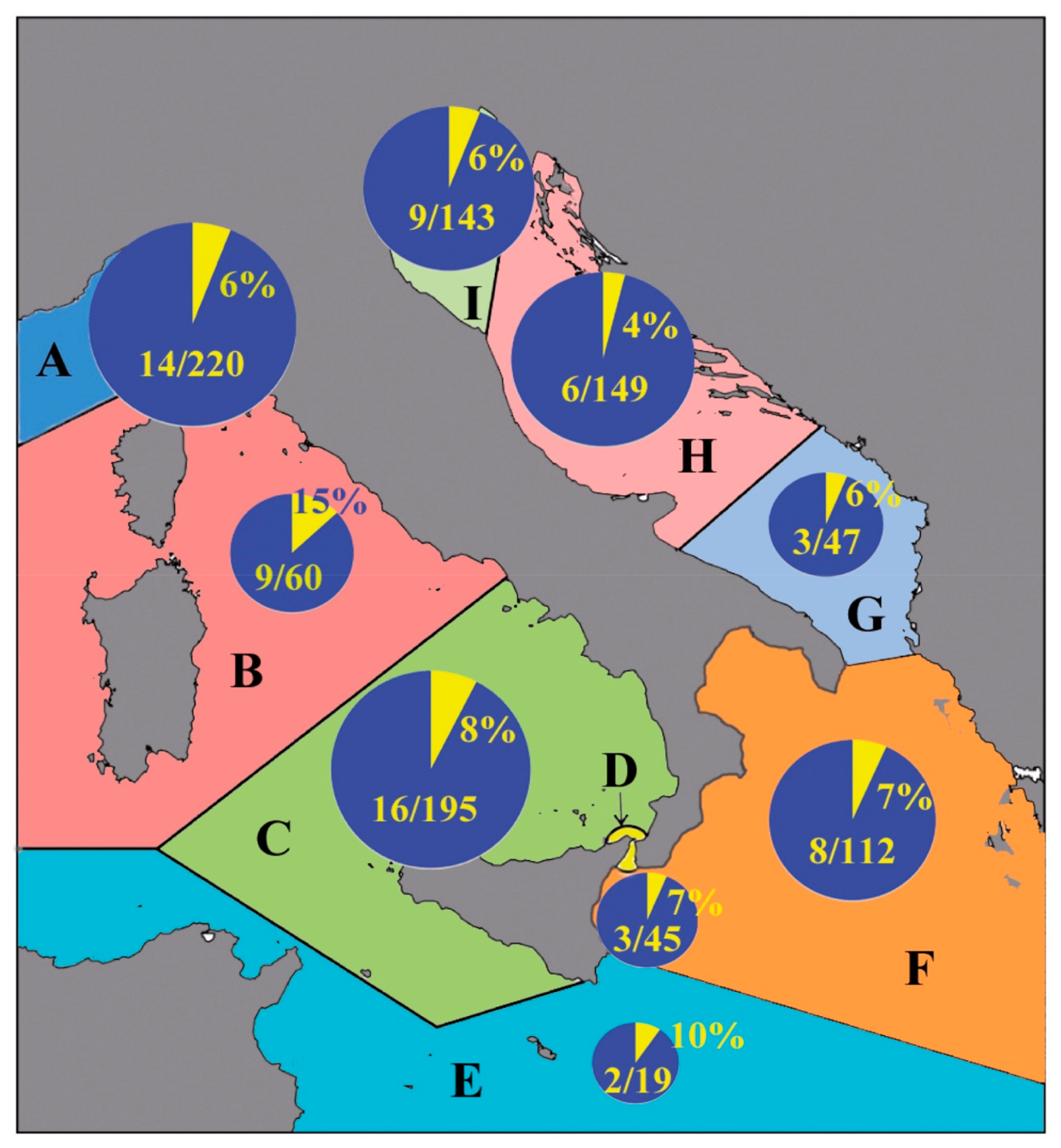
3.1. Cnidaria and Ctenophora Potential Invaders of the Italian Waters
3.2. Oculina patagonica, a Positive Alien Species?
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tsiamis, K.; Zenetos, A.; Deriu, I.; Gervasini, E.; Cardoso, A.C. The native distribution of the European marine non-indigenous species. Aquat. Invasions 2018, 13, 187–198. [Google Scholar] [CrossRef]
- Servello, G.; Andaloro, F.; Azzurro, E.; Castriota, L.; Catra, M.; Chiarore, A.; Crocetta, F.; D’alessandro, M.; Denitto, F.; Froglia, C.; et al. Marine Alien Species in Italy: A Contribution to the Implementation of Descriptor D2 of The Marine Strategy Framework Directive. Mediterr. Mar. Sci. 2019, 20, 1–48. [Google Scholar] [CrossRef] [Green Version]
- Wallentinus, I.; Nyberg, C.D. Introduced marine organisms as habitat modifiers. Mar. Pollut. Bull. 2007, 55, 323–332. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Cardoso, A.C. Impacts of marine invasive alien species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Ojaveer, H.; Galil, B.G.; Carlton, J.T.; Alleway, H.; Goulletquer, P.; Lehtiniemi, M.; Marchini, A.; Miller, W.; Occhipinti-Ambrogi, A.; Peharda, M.; et al. Historical baselines in marine bioinvasions: Implications for policy and management. PLoS ONE 2018, 13, e0202383. [Google Scholar] [CrossRef] [Green Version]
- Nellemann, C.; Hain, S.; Alder, J. Rapid Response Assessment in Dead Water—Merging of Climate Change with Pollution, Over-Harvest, and Infestations in the World’s Fishing Grounds; United Nations Environment Programme, GRID-Arendal: Arendal, Norway, 2008. [Google Scholar]
- Costello, M.J.; Coll, M.; Danovaro, R.; Halpin, P.; Ojaveer, H.; Miloslavich, P. A census of marine biodiversity knowledge, resources, and future challenges. PLoS ONE 2010, 5, e12110. [Google Scholar] [CrossRef] [Green Version]
- Coll, M.; Piroddi, M.; Albouy, C.; Ben Rais Lasram, F.; Cheung, W.W.L.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean Sea under siege: Spatial overlap between marine biodiversity, cumulative threats and marine reserves. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Schlaepfer, M.A. Do non-native species contribute to biodiversity? PLoS Biol. 2018, 16, e2005568. [Google Scholar] [CrossRef] [Green Version]
- Boudouresque, C.F.; Verlaque, M. Biological pollution in the Mediterranean Sea: Invasive versus introduced macrophytes. Mar. Pollut. Bull. 2002, 44, 32–38. [Google Scholar] [CrossRef]
- Piazzi, L.; Meinesz, A.; Verlaque, M.; Akçali, B.; Antolić, B.; Argyrou, M.; Balata, D.; Ballesteros, E.; Calvo, S.; Cinelli, F.; et al. Invasion of Caulerpa racemosa var. cylindracea (Caulerpales, Chlorophyta) in the Mediterranean Sea: An assessment of the spread. Cryptogam. Algol. 2005, 26, 189–202. [Google Scholar]
- Piazzi, L.; Balata, D. Invasion of alien macroalgae in different Mediterranean habitats. Biol. Invasions 2009, 11, 193–204. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; Tuya, F.; Silliman, B.R. Evidence for impact of nonindigenous macroalgae: A meta-analysis of experimental and field studies. J. Phycol. 2009, 45, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Zenetos, A.; Çinar, M.E.; Pancucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; García Raso, J.E.; Çinar, M.E.; Almogi-Labin, A.; Ates, A.S.; Azzurro, E.; et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef] [Green Version]
- Katsanevakis, S.; Zenetos, A.; Belchior, C.; Cardoso, A.C. Invading European seas: Assessing pathways of introduction of marine aliens. Ocean Coast. Manag. 2013, 76, 64–74. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A.; Minchin, D.; Narščius, A.; Ojaveer, H.; Olenin, S. International arrivals: Widespread bioinvasions in European Seas. Ethol. Ecol. Evol. 2014, 26, 152–171. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.L.; Katsanevakis, S.; Zenetos, A.; Cardoso, A.C. Gateways to alien invasions in the European Seas. Aquat. Invasions 2014, 9, 133–144. [Google Scholar] [CrossRef]
- Cardeccia, A.; Marchini, A.; Occhipinti-Ambrogi, A.; Galil, B.; Gollasch, S.; Minchin, D.; Narščius, A.; Olenin, S.; Ojaveer, H. Assessing biological invasions in European Seas: Biological traits of the most widespread non-indigenous species. Estuar. Coast. Shelf Sci. 2016, 201, 17–28. [Google Scholar] [CrossRef]
- Ojaveer, H.; Olenin, S.; Narščius, A.; Florin, A.B.; Ezhova, E.; Gollasch, S.; Jensen, K.R.; Lehtiniemi, M.; Minchin, D.; Normant-Saremba, M.; et al. Dynamics of biological invasions and pathways over time: A case study of a temperate coastal sea. Biol. Invasions 2016, 19, 799–813. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A.; Ojaveer, H. The enlargement of the Suez Canal—Erythraean introductions and management challenges. Manag. Biol. Invasions 2017, 8, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A. Progress in Mediterranean bioinvasions two years after the Suez Canal enlargement. Acta Adriat. 2017, 58, 347–358. [Google Scholar] [CrossRef]
- Zenetos, A.; Çinar, M.E.; Crocetta, F.; Golani, D.; Rosso, A.; Servello, G.; Shenkar, N.; Turon, X.; Verlaque, M. Uncertainties and validation of alien species catalogues: The Mediterranean as an example. Estuar. Coast. Shelf Sci. 2017, 191, 171–187. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Zenetos, A.; Corsini-Foka, M.; Crocetta, F.; Gerovasileiou, V.; Karachle, P.K.; Simboura, N.; Tsiamis, K.; Pancucci, A. Deep cleaning of alien and cryptogenic species records in the Greek Seas (2018 update). Manag. Biol. Invasions 2018, 9, 209–226. [Google Scholar] [CrossRef] [Green Version]
- Katsanevakis, S.; Poursanidis, D.; Hoffman, R.; Rizgalla, J.; Rothman, S.B.-S.; Levitt-Barmats, Y.; Hadjioannou, L.; Trkov, D.; Garmendia, M.K.; Rizzo, M.; et al. Unpublished Mediterranean records of marine alien and cryptogenic species. BioInvasions Rec. 2020, 9, 165–182. [Google Scholar] [CrossRef]
- Galil, B.S.; Boero, F.; Campbell, M.L.; Carlton, J.T.; Cook, E.; Fraschetti, S.; Gollasch, S.; Hewitt, C.L.; Jelmert, A.; Macpherson, E.; et al. ‘Double trouble’: The expansion of the Suez Canal and marine bioinvasions in the Mediterranean Sea. Biol. Invasions 2015, 17, 973–976. [Google Scholar] [CrossRef] [Green Version]
- Masters, G.; Norgrove, L. Climate change and invasive alien species. CABI Work. Pap. 2010, 1, 30. [Google Scholar]
- Occhipinti-Ambrogi, A.; Galil, B.S. Marine alien species as an aspect of global change. Adv. Oceanogr. Limnol. Adv. Oceanogr. Limnol. 2010, 1, 199–218. [Google Scholar] [CrossRef]
- Gravili, C. Alien jellyfish in expansion: The contribution of taxonomy to ecology. In Jellyfish: Ecology, Distribution Patterns and Human Interactions; Mariottini, G.L., Ed.; Nova Publishers: New York, NY, USA, 2017; pp. 27–49. [Google Scholar]
- Ponti, M.; Perlini, R.A.; Ventra, V.; Grech, D.; Abbiati, M.; Cerrano, C. Ecological shifts in Mediterranean coralligenous assemblages related to gorgonian forest loss. PLoS ONE 2014, 9, e102782. [Google Scholar] [CrossRef]
- Boero, F. The future of the Mediterranean Sea ecosystem: Towards a different tomorrow. Rend. Fis. Acc. Lincei 2015, 26, 3–12. [Google Scholar] [CrossRef]
- Dukes, J.S.; Mooney, H.A. Does global change increase the success of biological invaders? Trends Ecol. Evol. 1999, 14, 135–139. [Google Scholar] [CrossRef]
- Cox, G.W. Alien Species and Evolution: The Evolutionary Ecology of Exotic Plants, Animals, Microbes, and Interacting Native Species; Island Press: Washington, DC, USA, 2004. [Google Scholar]
- Mannino, A.; Balistreri, P.; Deidun, A. The marine biodiversity of the Mediterranean Sea in a changing climate: The impact of biological invasions. In Mediterranean Identities—Environment, Society, Culture; Fuerst-Bjeliš, B., Ed.; IntechOpen: Zagabria, Croatia, 2017; pp. 101–127. [Google Scholar]
- Galil, B.S. Seeing Red: Alien species along the Mediterranean coast of Israel. Aquat. Invasions 2007, 2, 281–312. [Google Scholar] [CrossRef]
- Zenetos, A.; Pancucci-Papadopoulou, M.A.; Zogaris, S.; Papastergiadou, E.; Vardakas, L.; Aligizaki, K.; Economou, A. Aquatic alien species in Greece: Tracking sources, patterns and effects on the ecosystem. J. Biol. Res. Thessalon. 2009, 12, 135–172. [Google Scholar]
- Occhipinti-Ambrogi, A.; Marchini, A.; Cantone, G.; Castelli, A.; Chimenz, C.; Cormaci, M.; Froglia, C.; Furnari, G.; Gambi, M.C.; Giaccone, G.; et al. Alien species along the Italian coasts: An overview. Biol. Invasions 2011, 13, 215–237. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Çinar, M.E.; García Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr. Mar. Sci. 2010, 112, 381–493. [Google Scholar] [CrossRef] [Green Version]
- Appeltans, W.; Bouchet, P.; Boxshall, G.A.; De Broyer, C.; de Voogd, N.J.; Gordon, D.P.; Hoeksema, B.W.; Horton, T.; Kennedy, M.; Mees, J.; et al. (Eds.) World Register of Marine Species. 2012. Available online: http://www.marinespecies.org (accessed on 17 October 2020).
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, C.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- MSFD. Technical Document on the Delineation of MSFD Article 4 Marine Regions and Subregions; Jensen, H.M., Panagiotidis, P., Eds.; 16/05/2017. Version 1.0; European Environment Agency: København, Denmark, 2017; 21p. [Google Scholar]
- CBD. Pathways of Introduction of Invasive Species, Their Prioritization and Management; UNEP/CBD/SBSTTA/18/9/Add.1; CBD: Montreal, QC, Canada, June 2014; 18p. [Google Scholar]
- Boero, F.; Putti, M.; Trainito, E.; Prontera, E.; Piraino, S.; Shiganova, T.A. First records of Mnemiopsis leidyi (Ctenophora) from the Ligurian, Thyrrhenian and Ionian Seas (Western Mediterranean) and first record of Phyllorhiza punctata (Cnidaria) from the Western Mediterranean. Aquat. Invasions 2009, 4, 675–680. [Google Scholar] [CrossRef]
- Crocetta, F.; Agius, D.; Balistreri, P.; Bariche, M.; Bayhan, Y.K.; Çakir, M.; Ciriaco, S.; Corsini-Foka, M.; Deidun, A.; El Zrelli, R.; et al. New Mediterranean Biodiversity Records (October 2015). Mediterr. Mar. Sci. 2015, 16, 682–702. [Google Scholar] [CrossRef] [Green Version]
- Boero, F.; Brotz, L.; Gibbons, M.J.; Piraino, S.; Zampardi, S. Impacts and effects of ocean warming on jellyfish. In Explaining Ocean Warming: Causes, Scale, Effects and Consequences; Lagìffoley, D., Baxter, J.M., Eds.; IUCN: Gland, Switzerland, 2016; pp. 213–237. [Google Scholar]
- Balistreri, P.; Spiga, A.; Deidun, A.; Gueroun, S.K.; Yahia, M.N.D. Further spread of the venomous jellyfish Rhopilema nomadica Galil, Spannier & Ferguson, 1990 (Rhizostomeae, Rhizostomatidae) in the western Mediterranean. BioInvasions Rec. 2017, 6, 19–24. [Google Scholar] [CrossRef]
- Malej, A.; Tirelli, V.; Lučić, D.; Paliaga, P.; Vodopivec, M.; Goruppi, A.; Sara, A.; Benzi, M.; Bettoso, N.; Camatti, E.; et al. Mnemiopsis leidyi in the northern Adriatic: Here to stay? J. Sea Res. 2017, 124, 10–16. [Google Scholar] [CrossRef]
- Morri, C.; Puce, S.; Bianchi, C.N.; Bitar, G.; Zibrowius, H.; Bavestrello, G. Hydroids (Cnidaria: Hydrozoa) from the Levant Sea (mainly Lebanon), with emphasis on alien species. J. Mar. Biol. Assoc. U.K. 2009, 89, 49–62. [Google Scholar] [CrossRef]
- Schmidt, H.E.; Benović, A. Notes on the Hydromedusae (Cnidaria) from the Adriatic Sea. J. Mar. Biol. Assoc. U.K. 1977, 57, 635–640. [Google Scholar] [CrossRef]
- Dallot, S.; Goy, J.; Carré, C. Peuplements de carnivores planctoniques gélatineux et structures productives en Méditerranée occidentale. Oceanol. Acta 1988, SP, 193–209. [Google Scholar]
- Goy, J.; Lakkis, S.; Zeidane, R. Les Méduses de la Méditerranée orientale. Rapp. Comm. int. Mer Méditerr. 1988, 31, 299. [Google Scholar]
- Goy, J.; Lakkis, S.; Zeidane, R. Les Méduses de la Méditerranée orientale. In A Propos des Migrations Lessepsiennes; Godeaux, J., Ed.; Bulletin de l’Institut Océanographique, No. Spéc.; Musée Océanographique: Monaco-Ville, Monaco, 1990; Volume 7, pp. 79–88. [Google Scholar]
- Goy, J.; Lakkis, S.; Zeidane, R. Les méduses (Cnidaria) des eaux Libanaises. The medusae Cnidaria of Lebanese waters. Ann. Inst. Océanogr. Monaco 1991, 67, 99–128. [Google Scholar]
- Carré, D.; Carré, C. Complex reproductive cycle in Eucheilota paradoxica (Hydrozoa: Leptomedusae): Medusae, polyps and frustules produced from medusa stage. Mar. Biol. Berl. 1990, 104, 303–310. [Google Scholar] [CrossRef]
- Buecher, E.; Gibbons, M.J. Temporal persistence in the vertical structure of the assemblage of planktonic medusae in the NW Mediterranean Sea. Mar. Ecol. Prog. Ser. 1999, 189, 105–115. [Google Scholar] [CrossRef]
- Touzri, C.; Yahia, O.K.D.; Hamdi, H.; Goy, J.; Yahia, M.N.D. Spatio-temporal distribution of Medusae (Cnidaria) in the Bay of Bizerte (South Western Mediterranean Sea). Cah. Biol. Mar. 2010, 51, 167–176. [Google Scholar]
- Ounifi-Ben Amor, K.; Rifi, M.; Ghanem, R.; Draief, I.; Zaouali, J.; Ben Soussi, J. Update of alien fauna and new records from Tunisian marine waters. Mediterr. Mar. Sci. 2016, 17, 124–143. [Google Scholar] [CrossRef]
- Kherchouche, A.; Hafferssas, A. Species composition and distribution of Medusae (Cnidaria: Medusozoa) in the Algerian coast between 2°e and 7°e (SW Mediterranean Sea). Mediterr. Mar. Sci. 2020, 21, 52–61. [Google Scholar] [CrossRef]
- Lakkis, S.; Zeidane, R. Les hydroméduses des eaux néritiques libanaises: Composition et distribution. Rapp. Procès-Verbaux Réun. Comm. Int. Explor. Sci. Mer Méditerr. 1985, 29, 179–180. [Google Scholar]
- Benović, A.; Lucić, D.; Onofri, V.; Batistić, M.; Njire, J. Bathymetric distribution of medusae in the open waters of the middle and south Adriatic Sea during spring 2002. J. Plankton Res. 2005, 27, 79–89. [Google Scholar] [CrossRef]
- Casanova, J.-P. Campagnes du “Meteor” dans l’Atlantique, N.-E. Siphonophores, Méduses et Thécosomes. Distribution verticale et comparaisons faunistiques avec la Méditerranée. Meteor Forsch.-Erbgeb. 1980, 32, 15–32. [Google Scholar]
- Gili, J.M. Estudio bionómico y ecológico de los cnidarios bentónicos de las islas Medes (Girona). Oecol. Aquat. 1981, 5, 105–123. [Google Scholar]
- Gili, J.M. Estudio Sistematico y Faunistico de los Cnidarios de la Costa Catalana. Ph.D. Thesis, University of Barcelona, Barcelona, Spain, 1986; pp. 1–565. [Google Scholar]
- Gili, J.M.; Pagès, F.; Vives, F. Distribution and ecology of a population of planktonic cnidarians in the western Mediterranean. In Modern Trends in the Systematics, Ecology and Evolution of Hydroids and Hydromedusae; Bouillon, J., Boero, F., Cicogna, F., Cornelius, P.F.S., Eds.; Clarendon Press: Oxford, UK, 1987; pp. 157–170. [Google Scholar]
- Gili, J.M.; Bouillon, J.; Pagès, F.; Palanques, A.; Puig, P.; Heussner, S. Origin and biogeography of the deep-water Mediterranean Hydromedusae including the description of two new species collected in submarine canyons of Northwestern Mediterranean. Sci. Mar. 1998, 62, 113–134. [Google Scholar] [CrossRef] [Green Version]
- Gili, J.M.; Pagès, F.; Bouillon, J.; Palanques, A.; Puig, P.; Heussner, S.; Calafat, A.; Canals, M.; Monaco, A. A multidisciplinary approach to the understanding of hydromedusan populations inhabiting Mediterranean submaryne canyons. Deep Sea Res. Part I 2000, 47, 1513–1533. [Google Scholar] [CrossRef]
- Touzri, C.; Hamdi, H.; Goy, J.; Yahia, M.N.D. Diversity and distribution of gelatinous zooplankton in the Southwestern Mediterranean Sea. Mar. Ecol. 2012, 33, 393–406. [Google Scholar] [CrossRef]
- Bitar, G.; Bitar-Kouli, S. Impact de la pollution sur la répartition des peuplements de substrat dur à Beyrouth (Liban—Mediterranée Orientale). Rapp. Comm. Int. Mer Méditerr. 1995, 34, 19. [Google Scholar]
- Zibrowius, H.; Bitar, G. Invertebres marins exotiques sur la côte du Liban. Leban. Sci. J. 2003, 4, 67–74. [Google Scholar]
- Çinar, M.E.; Bilecenoglu, M.; Öztürk, B.; Can, A. New records of alien species on the Levantine coast of Turkey. Aquat. Invasions 2006, 1, 84–90. [Google Scholar] [CrossRef]
- Uysal, I.; Turan, C. Impacts and risk of venomous and sting marine alien species in Turkish marine waters. Biharean Biol. 2019, 14, 41–48. [Google Scholar]
- Zakaria, H.Y. Pelagic coelenterates in the waters of the western part of the Egyptian Mediterranean coast during summer and winter. Oceanologia 2004, 46, 253–268. [Google Scholar]
- García Corrales, P.; Aguirre Inchaurbe, A.; González Mora, D. Contribución al conocimiento de los hidrozoos de las costas españolas. Parte I: Halecidos, Campanularidos y Plumularidos. Boln. Inst. Esp. Oceanogr. 1978, 4, 5–73. [Google Scholar]
- Roca, I.; Moreno, I. Distribución de los cnidarios bentónicos litorales en tres localidades de la margen W de la Bahía de Palma de Mallorca. Boll. Soc. Hist. Nat. Balear. 1985, 29, 19–30. [Google Scholar]
- Roca Martinez, I. Estudio de los Cnidarios Bentónicos de las Aguas Costeras de Mallorca. Bachelor’s Thesis, Universitat de les Illes Balears, Facultat de Ciències, Palma de Mallorca, Spain, 1986; pp. 1–32. [Google Scholar]
- El Beshbeeshy, M. Descriptions of some thecate hydroids (Cnidaria-Hydrozoa) from the Egyptian Mediterranean waters. Part 4. Families: Plumulariidae, Halopteridae & Kirchenpaueriidae. Bull. Nat. Inst. Oceanogr. Fish. 1995, 21, 397–412. [Google Scholar]
- Billard, A. Hydroïdes des côtes de Syrie (Mission Gruvel). In Les États de Syrie. Richesse Marines et Fluviales. Exploitation Actuelle—Avenir; Gruvel, A., Ed.; Bibltque Faune Colon Françaises; 1931; Volume 3, pp. 389–395. [Google Scholar]
- Picard, J. Origines et affinités de la faune d’hydropolypes (Gymnoblastes et Calyptoblastes) et d’hydroméduses (Anthoméduses et Leptoméduses) de la Méditerranée. Rapp. Procès-Verbaux Réun. Comm. Int. Explor. Sci. Mer Méditerr. 1958, 14, 187–199. [Google Scholar]
- Marinopoulos, J. Biological survey of the eastern Mediterranean Sea: Hydroids (preliminary study). Rapp. Procès-Verbaux Réun. Comm. Int. Explor. Sci. Mer Méditerr. 1979, 2526, 119–120. [Google Scholar]
- El Beshbeeshy, M.E. Descriptions of some thecate hydroids (Cnidaria-Hydrozoa) from the Egyptian Mediterranean waters. Part 3 family: Sertulariidae. Bull. Natl. Inst. Oceanogr. Fish. 1994, 20, 145–159. [Google Scholar]
- González-Duarte, M.M.; Megina, C.; Bethencourt, M. Sertularia marginata (Cnidaria: Hydrozoa) in the Mediterranea: An alien species in expansion? Mediterr. Mar. Sci. 2013, 14, 384–389. [Google Scholar] [CrossRef] [Green Version]
- Lakkis, S. Contribution à l’étude du zooplancton des eaux libanaises. Mar. Biol. Berl. 1971, 11, 138–148. [Google Scholar]
- Redier, L. Hydraires et bryozoaires de Méditerranée. II—Banyuls-sur-Mer. Cah. Nat. 1962, 18, 33–38. [Google Scholar]
- Medel, M.D.; Vervoort, W. Plumularian hydroids (Cnidaria: Hydrozoa) from the Strait of Gibraltar and nearly areas. Zool. Verh. Leiden 1995, 300, 1–72. [Google Scholar]
- Galil, B.S.; Gershwin, L.-A.; Zorea, M.; Rahav, A.; Rothman, S.B.-S.; Fine, M.; Lubinevsky, H.; Douek, J.; Paz, G.; Rinkevich, B. Cotylorhiza erythraea Stiasny, 1920 (Scyphozoa: Rhizostomeae: Cepheidae), yet another erythraean jellyfish from the Mediterranean coast of Israel. Mar. Biodiv. 2017, 47, 229–235. [Google Scholar] [CrossRef]
- Galil, B.S.; Gershwin, L.-A.; Douek, J.; Rinkevich, B. Marivagia stellata gen. et sp. nov. (Scyphozoa: Rhizostomeae: Cepheidae), another alien jellyfish from the Mediterranean coast of Israel. Aquat. Invasions 2010, 5, 331–340. [Google Scholar] [CrossRef]
- Zaatari, M. Odd Marine Changes Strike Coast of Sidon. The Daily Star Lebanon. 17 October 2010. Available online: https://www.dailystar.com.lb/News/Lebanon-News/2010/Oct-07/59848-odd-marine-changes-strike-coast-of-sidon.ashx (accessed on 17 October 2020).
- Bitar, G.; Badreddine, A. First record of Marivagia stellata Galil and Gershwin, 2010 (Scyphozoa: Rhizostomeae: Cepheidae) from the Lebanese waters in the eastern Mediterranean Sea. J. Black Sea/Mediterr. Environ. 2019, 25, 172–177. [Google Scholar]
- Mamish, S.; Durgham, H.; Al-Masri, M.S. First record of the new alien sea jelly species Marivagia stellata Galil and Gershwin, 2010 off the Syrian coast. Mar. Biodivers. Rec. 2016, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; Ocaña Vicente, O. First record of the Central Indo-Pacific reef coral Oulastrea crispata in the Mediterranean Sea. Mediterr. Mar. Sci. 2014, 15, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.N. Proposta di suddivisione dei mari italiani in settori biogeografici. Notiziario SIBM 2004, 46, 57–59. [Google Scholar]
- Zibrowius, H. Oculina patagonica, scléractiniaire hermatypique introduit en Méditerranée. Helgol. Wiss. Meeresun. 1974, 26, 153–173. [Google Scholar] [CrossRef] [Green Version]
- Fine, M.; Zibrowius, H.; Loya, Y. Oculina patagonica: A non-lessepsian scleractinian coral invading the Mediterranean Sea. Mar. Biol. 2001, 138, 1195–1203. [Google Scholar] [CrossRef]
- Salomidi, M.; Katsanevakis, S.; Issaris, Y.; Tsiamis, K. Anthropogenic disturbance of coastal habitats promotes the spread of the introduced scleractinian coral Oculina patagonica in the Mediterranean Sea. Biol. Invasions 2013, 15, 1961–1971. [Google Scholar] [CrossRef]
- Serrano, E.; Coma, R.; Ribes, M. Pattern of Oculina patagonica occurrence along the Iberian peninsula coastline: A first step to understand the factors affecting its invasion dynamics. Rapp. Comm. Int. Mer Méditerr. 2013, 40, 603. [Google Scholar]
- Rubio-Portillo, E.; Vazquez, M.; Valle, C. Growth and bleaching of the coral Oculina patagonica under different environmental conditions in the western Mediterranean Sea. Mar. Biol. 2014, 161, 1333–2343. [Google Scholar] [CrossRef]
- Armoza-Zvuloni, R.; Segal, R.; Kramarsky-Winter, E.; Loya, Y. Repeated bleaching events may result in high tolerance and notable gametogenesis in stony corals: Oculina patagonica as a model. Mar. Ecol. Prog. Ser. 2011, 426, 149–159. [Google Scholar] [CrossRef]
- Coma, R.; Serrano, E.; Linares, C.; Ribes, M.; Díaz, D.; Ballesteros, E. Sea urchins predation facilitates coral invasion in a marine reserve. PLoS ONE 2011, 6, e22017. [Google Scholar] [CrossRef] [Green Version]
- Airi, V.; Gizzi, F.; Marchini, C.; Goffredo, S. Sexual reproduction of Mediterranean scleractinian corals. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 269–278. [Google Scholar] [CrossRef]
- Quintanilla, E.; Gili, J.M.; López-González, P.J.; Tsounis, G.; Madurell, T.; Fiorillo, I.; Rossi, S. Sexual reproductive cycle of the epibiotic soft coral, Alcyonium coralloides (Octocorallia, Alcyonacea), growing on the Mediterranean gorgonian Paramuricea clavata. Aquat. Biol. 2013, 18, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Isla, E.; Bosch-Belmar, M.; Galli, G.; Gori, A.; Gristina, M.; Ingrosso, G.; Milisenda, G.; Piraino, S.; Rizzo, L.; et al. Changes of energy fluxes in the marine animal forest of the Anthropocene: Factors shaping the future seascape. ICES J. Mar. Sci. 2019, 76, 2008–2019. [Google Scholar] [CrossRef]
- Serrano, E.; Ribes, M.; Coma, R. Demographics of the zooxanthellate coral Oculina patagonica along the Mediterranean Iberian coast in relation to environmental parameters. Sci. Total Environ. 2018, 634, 1580–1592. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Azzola, A.; Parravicini, V.; Peirano, A.; Morri, C.; Montefalcone, M. Abrupt change in a subtidal rocky reef community coincided with a rapid acceleration of sea water warming. Diversity 2019, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Dragičević, B.; Anadoli, O.; Angel, D.; Benabdi, M.; Bitar, G.; Castriota, L.; Crocetta, F.; Deidun, A.; Jakov, D.; Edelist, D.; et al. New Mediterranean Biodiversity Records (December 2019). Mediterr. Mar. Sci. 2019, 20, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Ramil, F.; Vervoort, W. Report on the Hydroida collected by the “BALGIM” expedition in and around the Strait of Gibraltar. Zool. Verh. Leiden 1992, 277, 1–262. [Google Scholar]
- Bouillon, J.; Medel, M.D.; Pagès, F.; Gili, J.-M.; Boero, F.; Gravili, C. Fauna of the Mediterranean Hydrozoa. Sci. Mar. 2004, 68 (Suppl. 2), 5–438. [Google Scholar]
- Galil, B.S. Alien species in the Mediterranean Sea—Which, when, where, why? Hydrobiologia 2008, 606, 105–116. [Google Scholar] [CrossRef]
- UNEP/MAP/MED POL. Mariculture in the Mediterranean. MAP Tech. Rep. Ser. 2004, 140, 80. [Google Scholar]
- Azzurro, E.; Maynou, F.; Belmaker, J.; Golani, D.; Crooks, J.A. Lag times in Lessepsian fish invasion. Biol. Invasions 2016, 18, 2761–2772. [Google Scholar] [CrossRef]
- Zenetos, A.; Gratsia, E.; Cardoso, A.; Tsiamis, K. Time lags in reporting of biological invasions: The case of Mediterranean Sea. Mediterr. Mar. Sci. 2019, 20, 469–475. [Google Scholar] [CrossRef]
- EU. Council Regulation (EC) No 708/2007 of 11 June 2007 concerning use of alien and locally absent species in aquaculture. OJEU Off. J. Eur. Union 2007, 168, 1–17. [Google Scholar]
- Savini, D.; Occhipinti-Ambrogi, A.; Marchini, A.; Tricarico, E.; Gherardi, F.; Olenin, S.; Gollasch, S. The top 27 animal alien species introduced into Europe for aquaculture and related activities. J. Appl. Ichthyol. 2010, 26, 1–7. [Google Scholar] [CrossRef]
- MEPC. Guidelines for the Control and Management of Ships’ Biofouling to Minimize the Transfer of Invasive Aquatic Species; Annex 26. Resolution MEPC.207(62); Marine Environment Protection Committee: London, UK, 15 July 2011; 25p. [Google Scholar]
- Mineur, F.; Johnson, M.P.; Maggs, C.A. Macroalgal introductions by hull fouling on recreational vessels: Seaweeds and sailors. Environ. Manag. 2008, 42, 667–676. [Google Scholar] [CrossRef]
- Murray, C.C.; Pakhomov, E.A.; Therriault, T.W. Recreational boating: A large unregulated vector transporting marine invasive species. Divers. Distrib. 2011, 17, 1161–1172. [Google Scholar] [CrossRef]
- Gravili, C.; Di Camillo, C.G.; Piraino, S.; Boero, F. Hydrozoan species richness in the Mediterranean Sea: Past and present. Mar. Ecol. 2013, 34 (Suppl. 1), 41–62. [Google Scholar] [CrossRef]
- Foster, V.; Giesler, R.J.; Wilson, A.M.W.; Nall, C.R.; Cook, E.J. Identifying the physical features of marina infrastructure associated with the presence of non-native species in the UK. Mar. Biol. 2016, 163, 173. [Google Scholar] [CrossRef] [Green Version]
- Buonocore, E.; Franzese, P.P.; Russo, G.F. Marine ecosystem services: Understanding the value of nature. Water 2021. In press. [Google Scholar]
- González-Duarte, M.M.; Megina, C.; López-González, P.J.; Galil, B. Cnidarian alien species in expansion. In The Cnidaria, Past, Present and Future. The world of Medusa and her sisters; Goffredo, S., Dubinsky, Z., Eds.; Springer: Cham, Switzerland, 2016; pp. 139–160. [Google Scholar]
- Moura, C.J.; Cuhna, M.R.; Porteiro, F.M.; Rogers, A.D. A molecular phylogenetic appraisal of the systematics of the Aglaopheniidae (Cnidaria: Hydrozoa, Leptothecata) from the north-east Atlantic and west Mediterranean. Zool. J. Linnean Soc. 2012, 164, 717–727. [Google Scholar] [CrossRef] [Green Version]
- Byers, J.E.; Noonburg, E.G. Scale dependent effects of biotic resistance to biological invasion. Ecology 2003, 84, 1428–1433. [Google Scholar] [CrossRef]
- Corriero, G.; Pierri, C.; Accoroni, S.; Alabiso, G.; Bavestrello, G.; Barbone, E.; Bastianini, M.; Bazzoni, A.M.; Bernardi Aubry, F.; Boero, F.; et al. Ecosystem vulnerability to alien and invasive species: A case study on marine habitats along the Italian coast. Aquat. Conserv. Mar. Freshw. Ecosyst. 2015, 26, 392–409. [Google Scholar] [CrossRef]
- Levine, J.M. Species diversity and biological invasions: Relating local process to community pattern. Science 2000, 288, 852–854. [Google Scholar] [CrossRef] [Green Version]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Thieltges, D.W.; Strasser, M.; Reise, K. How bad are invaders in coastal waters? The case of the American slipper limpet Crepidula fornicata in western Europe. Biol. Invasions 2006, 8, 1673–1680. [Google Scholar] [CrossRef]
- Schlaepfer, M.A.; Sax, D.F.; Olden, J.D. The potential conservation value of non-native species. Conserv. Biol. 2011, 25, 428–437. [Google Scholar] [CrossRef] [PubMed]
- McLaughlan, C.; Gallardo, B.; Aldridge, D.C. How complete is our knowledge of the ecosystem services impacts of Europe’s top 10 invasive species? Acta Oecol. 2013, 54, 119–130. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Byers, J.E.; Schiel, D.R.; Bruno, J.F.; Olden, J.D.; Wernberg, T.; Silliman, B.R. Impacts of marine invaders on biodiversity depend on trophic position and functional similarity. Mar. Ecol. Prog. Ser. 2014, 495, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. BioScience 2000, 50, 53–65. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Byrnes, J.E. Species diversity, invasion success, and ecosystem functioning: Disentangling the influence of resource competition, facilitation, and extrinsic factors. Mar. Ecol. Prog. Ser. 2006, 311, 251–262. [Google Scholar] [CrossRef]
- Galil, B.S. Loss or gain? Invasive aliens and biodiversity in the Mediterranean Sea. Mar. Pollut. Bull. 2007, 55, 314–322. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Olden, J.D.; Wernberg, T.; Griffin, J.N.; Silliman, B.R. A broad framework to organize and compare ecological invasion impacts. Environ. Res. 2011, 111, 899–908. [Google Scholar] [CrossRef]
- Perrings, C.; Williamson, M.; Barbier, E.B.; Delfino, D.; Dalmazzone, S.; Shogren, J.; Simmons, P.; Watkinson, A. Biological invasion risks and the public good: An economic perspective. Conserv. Ecol. 2002, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Dick, J.T.A.; Alexander, M.E.; Jeschke, J.M.; Ricciardi, A.; MacIsaac, H.J.; Robinson, T.B.; Kumschick, S.; Weyl, O.L.F.; Dunn, A.M.; Hatcher, M.J.; et al. Advancing impact prediction and hypothesis testing in invasion ecology using a comparative functional response approach. Biol. Invasions 2014, 16, 735–753. [Google Scholar] [CrossRef] [Green Version]
- Ojaveer, H.; Galil, B.S.; Campbell, M.L.; Carlton, J.T.; Canning-Clode, J.; Cook, E.J.; Davidson, A.D.; Hewitt, C.L.; Jelmert, A.; Marchini, A.; et al. Classification of non-indigenous species based on their impacts: Considerations for application in marine management. PLoS Biol. 2015, 13, e1002130. [Google Scholar] [CrossRef] [Green Version]
- Giakoumi, S.; Katsanevakis, S.; Albano, P.G.; Azzurro, E.; Cardoso, A.C.; Cebrian, E.; Deidun, A.; Edelist, D.; Francour, P.; Jimenez, C.; et al. Management priorities for marine invasive species. Sci. Total Environ. 2019, 688, 976–982. [Google Scholar] [CrossRef]
- Giangrande, A.; Pierri, C.; Del Pasqua, M.; Gravili, C.; Gambi, M.C.; Gravina, M.F. The Mediterranean in check: Biological invasions in a changing sea. Mar. Ecol. 2020, 41, e12583. [Google Scholar] [CrossRef]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invisibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Zaiko, A.; Olenin, S.; Daunys, D.; Nalepa, T. Vulnerability of benthic habitats to the aquatic invasive species. Biol. Invasions 2007, 9, 703–714. [Google Scholar] [CrossRef]
- Paolucci, E.M.; MacIsaac, K.J.; Ricciardi, A. Origin matters: Alien consumers inflict greater damage on prey populations than do native consumers. Divers. Distrib. 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Simberloff, D. Biological invasions: What’s worth fighting and what can be won? Ecol. Eng. 2014, 65, 112–121. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Lorenzoni, M. The role of climate changes in the spread of freshwater fishes: Implications for alien cool and warm-water species in a Mediterranean basin. Water 2021, 13, 347. [Google Scholar] [CrossRef]
- Ceccherelli, G.; Campo, D. Different effects of Caulerpa racemosa on two co-occurring seagrasses in the Mediterranean. Bot. Mar. 2002, 45, 71–76. [Google Scholar] [CrossRef]
- Grosholz, E. Ecological and evolutionary consequences of coastal invasions. Trends Ecol. Evol. 2002, 17, 22–27. [Google Scholar] [CrossRef]
- Arenas, F.; Sanchez, I.; Hawkins, S.; Jenkins, S.R. The invasibility of marine algal assemblages: Role of functional diversity and identity. Ecology 2006, 87, 2851–2861. [Google Scholar] [CrossRef]
- Ricciardi, A.; Cohen, J. The invasiveness of an introduced species does not predict its impact. Biol. Invasions 2007, 9, 309–315. [Google Scholar] [CrossRef]
- Kumschick, S.; Gaertner, M.; Vilà, M.; Essl, F.; Jeschke, J.M.; Pyšek, P.; Ricciardi, A.; Bacher, S.; Blackburn, T.M.; Dick, J.T.A.; et al. Ecological impacts of alien species: Quantification, scope, caveats, and recommendations. Bioscience 2015, 65, 55–63. [Google Scholar] [CrossRef]
- Zwerschke, N.; Hollyman, P.R.; Wild, R.; Strigner, R.; Turner, J.R.; King, J.W. Limited impact of an invasive oyster on intertidal assemblage structure and biodiversity: The importance of environmental context and functional equivalency with native species. Mar. Biol. 2018, 165, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanno, G.; Orlando-Bonaca, M. Non-indigenous marine species in the Mediterranean Sea—Myth and reality. Environ. Sci. Policy 2019, 96, 123–131. [Google Scholar] [CrossRef]
- Sagoff, M. Do non-native species threaten the natural environment? J. Agric. Environ. Ethics 2005, 18, 215–236. [Google Scholar] [CrossRef]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.A.; Chew, M.K.; Hobbs, R.J.; Lugo, A.E.; Ewel, J.J.; Vermeij, G.J.; Brown, J.H.; Rosenzweig, M.L.; Gardener, M.R.; Carroll, S.P.; et al. Don’t judge species on their origins. Nature 2011, 474, 153–154. [Google Scholar] [CrossRef]
- Galil, B.S. Taking stock: Inventory of alien species in the Mediterranean Sea. Biol. Invasions 2009, 11, 359–372. [Google Scholar] [CrossRef]
- Galil, B.S. The alien crustaceans in the Mediterranean Sea: An historical review. In In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: Cham, Netherlands, 2011; pp. 377–401. [Google Scholar]
- Sala, E.; Kizilkaya, Z.; Yildirim, D.; Ballesteros, E. Alien marine fishes deplete algal biomass in the eastern Mediterranean. PLoS ONE 2011, 6, e17356. [Google Scholar] [CrossRef] [Green Version]
- Killi, N.; Tarkan, A.S.; Kozic, S.; Copp, G.H.; Davison, P.I.; Vilizzi, L. Risk screening of the potential invasiveness of non-native jellyfishes in the Mediterranean Sea. Mar. Pollut. Bull. 2020, 150, 110728. [Google Scholar] [CrossRef]
- Ekebom, J. The long and winding road of the ecosystem approach into marine environmental policies. Aquat. Conserv. Mar. Freshw. Ecosyst 2013, 23, 1–6. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Origin | Main Way of Introduction | Distribution in Mediterranean Sea and Records | Last Record
(Locality and Year) | Probable Invasiveness |
|---|---|---|---|---|---|
| Hydrozoa | |||||
| Dynamena quadridentata (Ellis & Solander, 1786) | Tropical Atlantic | Suez Canal (Lessepsian species) | Levant Sea [50] | Lebanon coast (2004) | ++ |
| Eucheilota paradoxica Mayer, 1900 | Tropical Atlantic | Unknown | Croatian coast [51], Alborán Sea [52], Levant Sea [53,54,55], French coast [56,57], Tunisian coast [58,59], Algerian coast [60] | Tunisian coast (2015) | ++ |
| Eucheilota ventricularis McCrady, 1859 | North temperate Atlantic | Suez Canal (Lessepsian species) | Levant Sea [53,54,55,61] | Lebanon coast (1989) | + |
| Haliscera bigelowi Kramp, 1947 | Tropical eastern Pacific | Unknown | Croatian coast [51,62], Alborán Sea [63], Spanish coast [64,65,66,67,68], Tunisian coast [59,69] | Tunisian coast (2015) | ++ |
| Macrorhynchia philippina Kirchenpauer, 1872 | Central Indo- Pacific | Suez Canal (Lessepsian species) | Lebanon coast [50,70,71], Levantine coast of Turkey [72], Suez Canal [22], Aegean Sea [73] | Suez Canal (2014) | +++ |
| Moerisia carine Bouillon, 1978 | Central Indo- Pacific | Suez Canal (Lessepsian species) | Levant Sea [53,54,55] | Lebanon coast (1989) | + |
| Olindias singularis Browne, 1905 | Central Indo- Pacific | Suez Canal (Lessepsian species) | Egyptian Mediterranean coast [74] | Egyptian Mediterranean coast (extending from Alexandria in the east to Sidi Barani in the west) (2001) | ++ |
| Plumularia pulchella Bale, 1882 | Temperate Australasia | Unknown | Alborán Sea [65], Spanish coast [75,76,77], Egyptian Mediterranean waters [78] | Egyptian Mediterranean waters (1977) | + |
| Sertularia marginata (Kirchenpauer, 1864) | Western Indo- Pacific | Suez Canal (Lessepsian species) | Levant Sea [50,79,80,81,82], Alborán Sea [83] | Alborán Sea (2012) | +++ |
| Sertularia tongensis (Stechow, 1919) as S. thecocarpa | Western Indo- Pacific | Suez Canal (Lessepsian species) | Levant Sea [50] | Lebanon coast (2004) | ++ |
| Tetrorchis erythrogaster Bigelow, 1909 | Tropical eastern Pacific | Unknown | Beyrouth [53,54,55,61,84] | Lebanon coast (1982) | + |
| Gymnangium montagui (Billard, 1912) | Temperate North Atlantic | Espansion natural range | French coast [85], Alborán Sea [86] | Strait of Gibraltar and nearly areas (1993) | ++ |
| Scyphozoa | |||||
| Cotylorhiza erythraea Stiasny, 1920 | Western Indo- Pacific | Suez Canal (Lessepsian species) | Levant Sea [87] | Israel coast (2015) | +++ |
| Marivagia stellata Galil & Gershwin, 2010 | Western Indo- Pacific | Shipping (ballast waters) (Lessepsian species) | Levant Sea [88,89,90,91] | Syrian coast (2015), Lebanese waters (2015) | +++ |
| Anthozoa | |||||
| Oulastrea crispata (Lamarck, 1816) | Central Indo- Pacific | Unknown | Corsica [92] | Corsica (2014) | ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gravili, C.; Rossi, S. Who’s Next? Non-Indigenous Cnidarian and Ctenophoran Species Approaching to the Italian Waters. Water 2021, 13, 1062. https://doi.org/10.3390/w13081062
Gravili C, Rossi S. Who’s Next? Non-Indigenous Cnidarian and Ctenophoran Species Approaching to the Italian Waters. Water. 2021; 13(8):1062. https://doi.org/10.3390/w13081062
Chicago/Turabian StyleGravili, Cinzia, and Sergio Rossi. 2021. "Who’s Next? Non-Indigenous Cnidarian and Ctenophoran Species Approaching to the Italian Waters" Water 13, no. 8: 1062. https://doi.org/10.3390/w13081062