
A New, Catchment-Scale Integrated Water Quality Model of Phosphorus, Dissolved Oxygen, Biochemical Oxygen Demand and Phytoplankton: INCA-Phosphorus Ecology (PEco)

 , , , and
, , , and
Abstract
1. Introduction
2. Methods
2.1. Model Development
2.1.1. Terrestrial Phase
2.1.2. Water Phase
2.2. Model Applications
Model Setup
2.3. Sensitivity Analysis
3. Results and Discussion
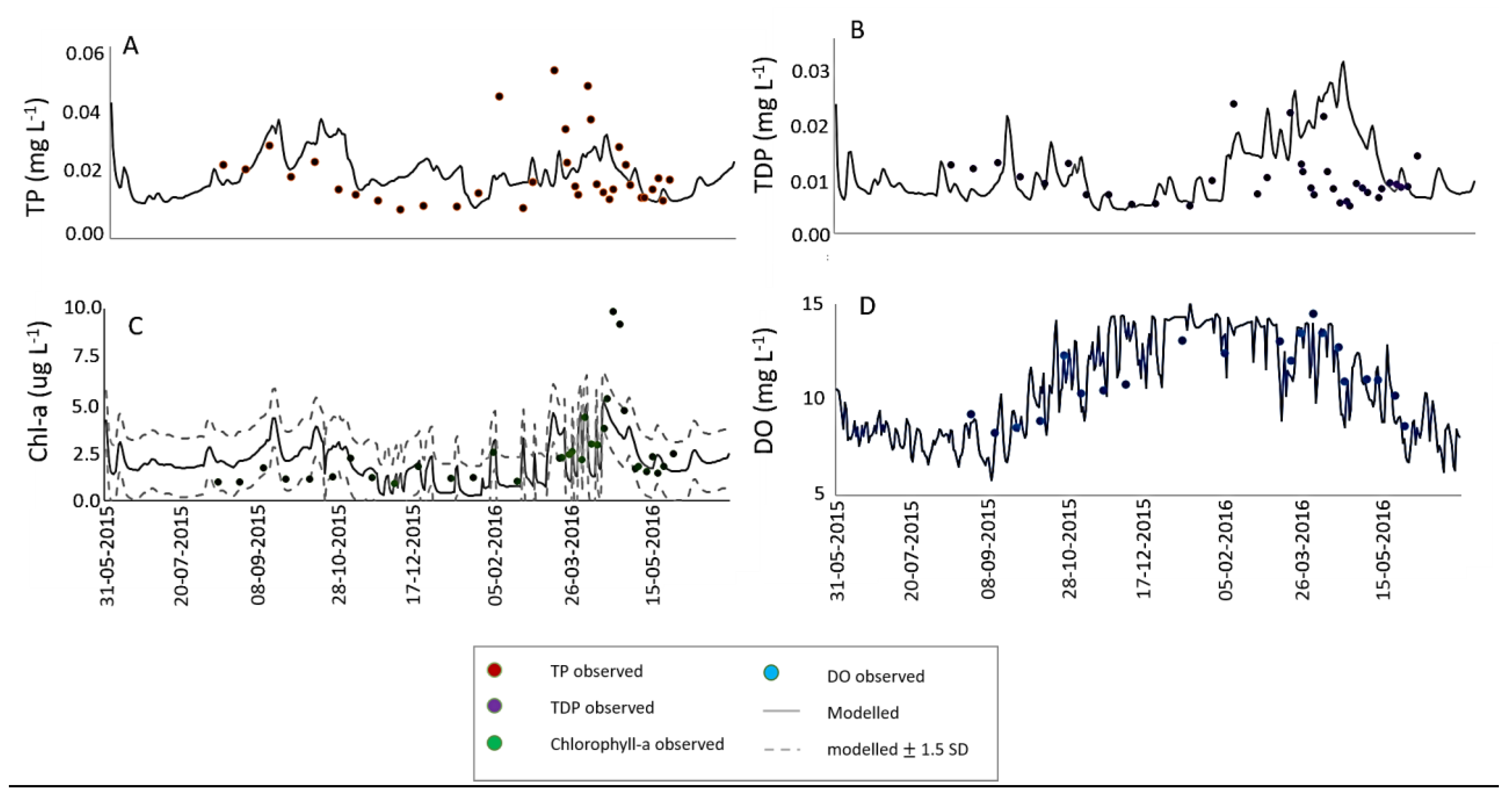
3.1. Calibration
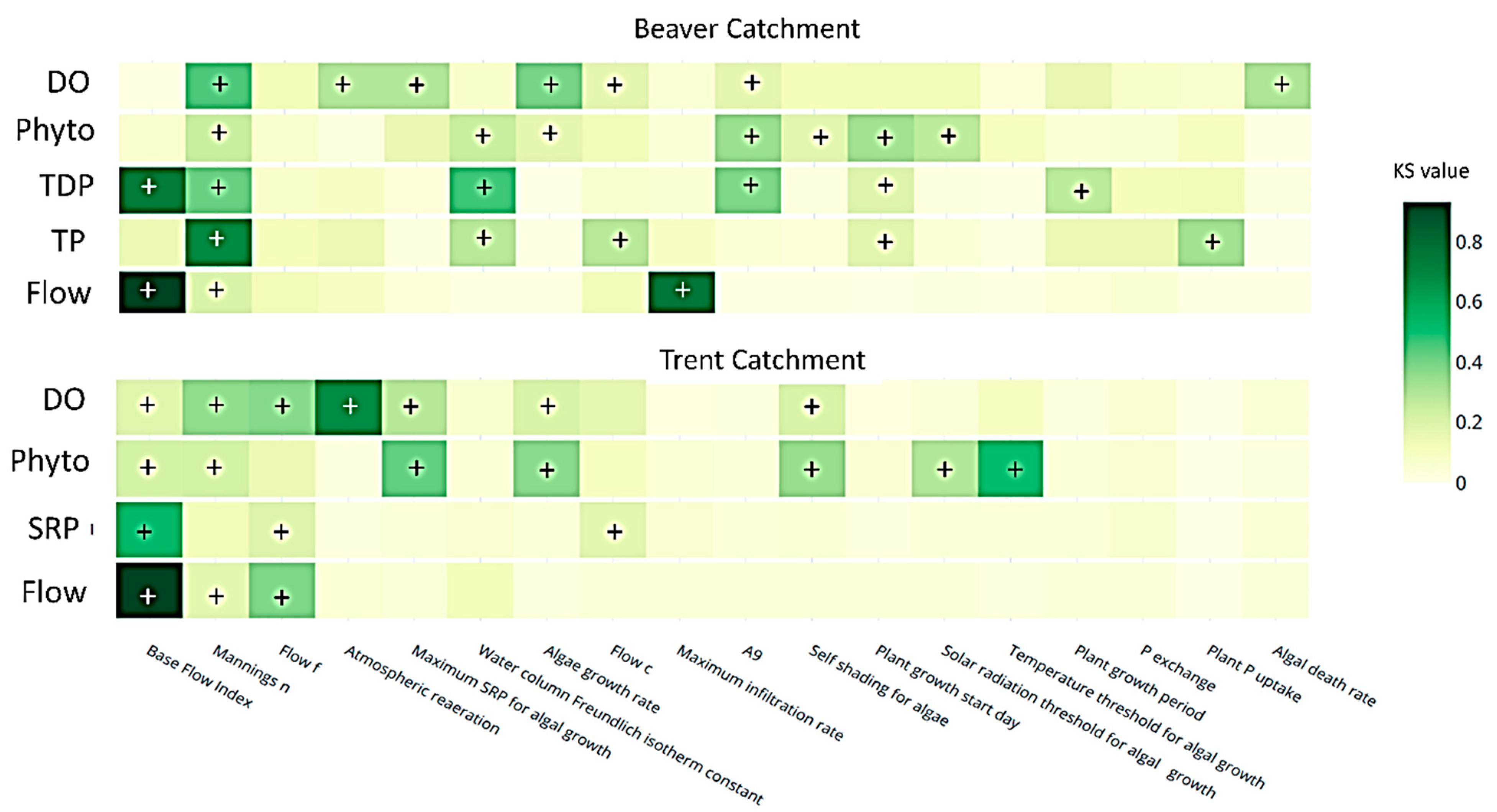
3.2. Sensitivity Analysis
3.2.1. Flow and P
3.2.2. Phytoplankton Concentration
3.2.3. Dissolved Oxygen
3.3. Study Limitations and Further Research
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rabalais, N.N.; Diaz, R.J.; Levin, L.A.; Turner, R.E.; Gilbert, D.; Zhang, J. Dynamics and distribution of natural and human-caused hypoxia. Biogeosciences 2010, 7, 585–619. [Google Scholar] [CrossRef]
- Tian, R. Factors Controlling Hypoxia Occurrence in Estuaries, Chester River, Chesapeake Bay. Water 2020, 12, 1961. [Google Scholar] [CrossRef]
- Wen, Y.; Schoups, G.; van de Giesen, N. Organic pollution of rivers: Combined threats of urbanization, livestock farming and global climate change. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Mallin, M.A.; Johnson, V.L.; Ensign, S.H.; MacPherson, T.A. Factors contributing to hypoxia in rivers, lakes, and streams. Limnol. Oceanogr. 2006, 51, 690–701. [Google Scholar] [CrossRef]
- Jang, G.-S.; An, K.-G. Physicochemical water quality characteristics in relation to land use pattern and point sources in the basin of the Dongjin River and the ecological health assessments using a fish multi-metric model. J. Ecol. Environ. 2016, 40, 6. [Google Scholar] [CrossRef]
- Liu, C.; Li, Z.; Dong, Y.; Chang, X.; Nie, X.; Liu, L.; Xiao, H.; Wang, D.; Peng, H. Response of sedimentary organic matter source to rainfall events using stable carbon and nitrogen isotopes in a typical loess hilly-gully catchment of China. J. Hydrol. 2017, 552, 376–386. [Google Scholar] [CrossRef]
- Nelson, N.G.; Muñoz-Carpena, R.; Phlips, E.J.; Kaplan, D.; Sucsy, P.; Hendrickson, J. Revealing Biotic and Abiotic Controls of Harmful Algal Blooms in a Shallow Subtropical Lake through Statistical Machine Learning. Environ. Sci. Technol. 2018, 52, 3527–3535. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, P.G.; Bussi, G.; Bowes, M.J.; Read, D.S.; Hutchins, M.G.; Elliott, J.A.; Dadson, S.J. Dynamic modelling of multiple phytoplankton groups in rivers with an application to the Thames river system in the UK. Environ. Model. Softw. 2015, 74, 75–91. [Google Scholar] [CrossRef]
- Chapra, S.; Pelletier, G.; Tao, H. Qualk2K: A Modelling Framework for Simulating River and Stream Water Quality (Version 2.11). Documentation and Users Manual; Civil and Environmental Engineering Department, Tufts University: Medford, MA, USA, 2008; Available online: http://www.ecs.umass.edu/cee/reckhow/courses/577/Qual2/Q2KDocv2_11b8%20v211.pdf (accessed on 15 February 2021).
- Elliott, J.A.; Persson, I.; Thackeray, S.J.; Blenckner, T. Phytoplankton modelling of Lake Erken, Sweden by linking the models PROBE and PROTECH. Ecol. Model. 2007, 202, 421–426. [Google Scholar] [CrossRef]
- Crossman, J.; Futter, M.; Elliott, J.; Whitehead, P.; Jin, L.; Dillon, P. Optimizing land management strategies for maximum improvements in lake dissolved oxygen concentrations. Sci. Total. Environ. 2019, 652, 382–397. [Google Scholar] [CrossRef]
- Moreno-Rodenas, A.M.; Tscheikner-Gratl, F.; Langeveld, J.G.; Clemens, F.H. Uncertainty analysis in a large-scale water quality integrated catchment modelling study. Water Res. 2019, 158, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Tscheikner-Gratl, F.; Bellos, V.; Schellart, A.; Moreno-Rodenas, A.; Muthusamy, M.; Langeveld, J.; Clemens, F.; Benedetti, L.; Rico-Ramirez, M.A.; de Carvalho, R.F.; et al. Recent insights on uncertainties present in integrated catchment water quality modelling. Water Res. 2019, 150, 368–379. [Google Scholar] [CrossRef]
- Getz, W.M.; Marshall, C.R.; Carlson, C.J.; Giuggioli, L.; Ryan, S.J.; Romañach, S.S.; Boettiger, C.; Chamberlain, S.D.; Larsen, L.; D’Odorico, P.; et al. Making ecological models adequate. Ecol. Lett. 2018, 21, 153–166. [Google Scholar] [CrossRef]
- Wade, A.J.; Whitehead, P.G.; Butterfield, D. The Integrated Catchments model of Phosphorus dynamics (INCA-P), a new approach for multiple source assessment in heterogeneous river systems: Model structure and equations. Hydrol. Earth Syst. Sci. 2002, 6, 583–606. [Google Scholar] [CrossRef]
- Jackson-Blake, L.A.; Wade, A.J.; Futter, M.N.; Butterfield, D.; Couture, R.M.; Cox, B.A.; Crossman, J.; Ekholm, P.; Halliday, S.J.; Jin, S.; et al. The INtegrated CAtchment model of phosphorus dynamics (INCA-P); Description and demonstra-tion of new model structure and equations. Environ. Model. Softw. 2016, 83, 356–386. [Google Scholar] [CrossRef]
- Whitehead, P.G.; Wilson, E.J.; Butterfield, D. A semi-distributed Integrated Nitrogen model for multiple source as-sessment in Catchments (INCA): Part I—model structure and process equations. Sci. Total Environ. 1998, 210, 547–558. [Google Scholar] [CrossRef]
- Wang, A.; Solomatine, D.P. Practical Experience of Sensitivity Analysis: Comparing Six Methods, on Three Hydrological Models, with Three Performance Criteria. Water 2019, 11, 1062. [Google Scholar] [CrossRef]
- Cox, B.A. Dynamic Modelling of Dissolved Oxygen: A Case-Study for the River Thames. Ph.D. Thesis, University of Reading, Reading, UK, 2016. [Google Scholar]
- Elliott, J.A.; Reynolds, C.S.; Irish, A.E.; Tett, P. Exploring the potential of the PROTECH model to investigate phyto-plankton community theory. Hydrobiologia 1999, 414, 37–43. [Google Scholar] [CrossRef]
- Crossman, J.; Elliott, J. Bridging the gap between terrestrial, riverine and limnological research: Application of a model chain to a mesotrophic lake in North America. Sci. Total. Environ. 2018, 622–623, 1363–1378. [Google Scholar] [CrossRef] [PubMed]
- Environment Agency Water Quality Archive. Available online: https://environment.data.gov.uk/water-quality/view/download/new (accessed on 8 January 2020).
- Bussi, G.; Whitehead, P.G. Impacts of droughts on low flows and water quality near power stations. Hydrol. Sci. J. 2020, 65, 898–913. [Google Scholar] [CrossRef]
- Rivett, M.O.; Ellis, P.A.; Mackay, R. Urban groundwater baseflow influence upon inorganic river-water quality: The River Tame headwaters catchment in the City of Birmingham, UK. J. Hydrol. 2011, 400, 206–222. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmerman, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Kö ppen-Geiger climate classification maps at 1km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed]
- Futter, M.N.; Erlandsson, M.A.; Butterfield, D.; Whitehead, P.G.; Oni, S.K.; Wade, A.J. PERSiST: A flexible rainfall-runoff modelling toolkit for use with the INCA family of models. Hydrol. Earth Syst. Sci. 2014, 18, 855–873. [Google Scholar] [CrossRef]
- Thornton, M.M.; Shrestha, Y.W.; Thornton, P.E.; Kao, S.; Wilson, B.E. Daymet: Daily Surface Weather Data on a 1-km Grid for North America, Version 4; ORNL DAAC: Oak Ridge, TN, USA, 2020. [CrossRef]
- Met Office. Met Office Integrated Data Archive System (MIDAS) Land and Marine Surface Stations Data (1853-Current). Center for Environmental Data Analyais. 2012. Available online: https://catalogue.ceda.ac.uk/uuid/dbd451271eb04662beade68da43546e1 (accessed on 8 January 2020).
- OMAFRA. Ontario Ministry of Agriculture, Food and Rural Affairs: Agronomy Guide for Field Crops. 2009. Available online: http://www.omafra.gov.on.ca/english/crops/pub811/p811toc.html (accessed on 30 January 2021).
- Statistics Canada. Farm and Operator Data. Census of Agriculture. 2011. Available online: https://www.statcan.gc.ca/eng/ca2011/index (accessed on 30 January 2021).
- Stephens, S.L.S. Optimising Agricultural and Urban Pollution Remediation Measures Using Watershed Modelling: Review, Calibration, Validation and Applications of the CANWET Model in the Lake Simcoe Watershed. Master’s Thesis, Trent University, Trent, ON, Canada, 2007. [Google Scholar]
- Louis Berger Group Inc. Estimation of the Phosphorus Loadings to Lake Simcoe; The Louis Berger Group Inc.: Washington, DC, USA, 2020. [Google Scholar]
- Ramwakellan, J.; Gharabaghi, B.; Winder, J.G. Application of weather radar in estimation of bulk atmospheric deposition of total phosphorus over Lake Simcoe. J. Can. Water Resour. 2009, 34, 37–60. [Google Scholar] [CrossRef]
- Fournier, R.E.; Morrison, I.K.; Hopkin, A.A. Short range variability of soil chemistry in three acid soils in Ontario, Canada. Commun. Soil Sci. Plant Anal. 1994, 25, 3069–3082. [Google Scholar] [CrossRef]
- Peltouvouri, T. Phosphorus in Agricultural Soils of Finland—Characterisation of Reserves and Retention in Mineral Soil Profiles, Pro Terra No. 26. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2006. [Google Scholar]
- Vaananen, R. Phosphorus Retention in Forest Soils and the Functioning of Buffer Zones Used in Forestry. Ph.D. Thesis, Department of Forest Ecology, University of Helsinki, Helsinki, Finland, 2008; p. 42. [Google Scholar] [CrossRef]
- Koski–Vähälä, J. Role of Resuspension and Silicate in Internal Phosphorus Loading. Master’s Thesis, Department of Limnology and Environmental Protection, Department of Applied Chemistry and Microbiology, University of Helsinki, Helsinki, Finland, 2001. [Google Scholar]
- Provincial Groundwater Monitoring Network. Provincial Groundwater Monitoring Network Program: Groundwater Level Data, Groundwater Chemistry Data, and Precipitation Data, Ministry of Environment. 2012. Available online: https://www.javacoeapp.lrc.gov.on.ca/geonetwork/srv/en/metadata (accessed on 8 November 2019).
- DEFRA. Soil Nutrient Balances. UK Provisional Estimates for 2012. Available online: https://www.farminguk.com/content/knowledge/Soil%20nutrient%20balances%20UK%20provisional%20estimates%20for%202012(335).pdf (accessed on 10 January 2020).
- CIESIN. University, Center for International Earth Science Information Network—CIESIN—Columbia. Documentation for the Gridded Population of the World, Version 4 (GPWv4); NASA Socioeconomic Data and Applications Center (SEDAC): Palisades, NY, USA, 2016. [Google Scholar] [CrossRef]
- Harman, M.; Krinke, J.; Ren, J.; Yoo, S. Search Based Data Sensitivity Analysis Applied to Requirement Engineering. In Proceedings of the 11th Annual conference on Genetic and evolutionary computation, Montreal, QC, Canada, 8–12 July 2009; pp. 1681–1688. [Google Scholar]
- Niida, A.; Hasegawa, T.; Miyano, S. Sensitivity analysis of agent-based simulation utilizing massively parallel computation and interactive data visualization. PLoS ONE 2019, 14, e0210678. [Google Scholar] [CrossRef]
- Arvola, L. Spring phytoplankton of 54 small lakes in southern Finland. Hydrobiologia 1986, 137, 125–134. [Google Scholar] [CrossRef]
- Akomeah, E.; Chun, K.P.; Lindenschmidt, K. Dynamic water quality modelling and uncertainty analysis of phyto-plankton and nutrient cycles for the upper South Saskatchewan River. Environ. Sci. Pollut. Res. 2015, 22, 18239–18251. [Google Scholar] [CrossRef] [PubMed]
- Peters, F.; Dietmar, S.; Lorke, A.; Livingstone, D.M. Earlier onset of the spring phytoplankton bloom in lakes of the t emperate zone in a warmer climate. Glob. Chang. Biol. 2007, 13, 1898–1909. [Google Scholar] [CrossRef]
- O’Donnell, J.; Douglas, T.; Barker, A.; Guo, L. Changing Biogeochemical Cycles of Organic Carbon, Nitrogen, Phosphorus, and Trace Elements in Arctic Rivers. In Arctic Hydrology, Permafrost and Ecosystems; Springer: Cham, Switzerland, 2021; pp. 315–348. [Google Scholar]
- Crossman, J.; Futter, M.N.; Whitehead, E.; Stainsby, E.; Baulch, H.M.; Jin, L.; Oni, S.K.; Wilby, R.L.; Dillon, P.J. Flow pathways and nutrient transport mechanisms drive hydrochemical sensitivity to climate change across catchments with different geology and topography. Hydrol. Earth Syst. Sci. 2014, 18, 5125–5148. [Google Scholar] [CrossRef]
- Edwards, K.F.; Thomas, M.K.; Klausmeier, A.; Litchman, E. Phytoplankton growth and the interaction of light and temperature: A synethesissynthesis at the species and community level. Limnol. Oceanogr. 2016, 61, 1232–1244. [Google Scholar] [CrossRef]
- Skidmore, R.E.; Maberly, S.C.; Whitton, B.A. Patterns of spatial and temporal variation in phytoplankton chloro-phyll a in the River Trent and its tributaries. Sci. Total Environ. 1998, 210–211, 357–365. [Google Scholar] [CrossRef]
- Pinder, A.; Marker, A.F.H.; Pinder, A.C.; Ingram, J.K.G.; Leach, D.V.; Collet, G.D. Concentrations of suspended chlorophyll a in the Humber Rivers. Sci. Total Environ. 1997, 194–195, 373–378. [Google Scholar] [CrossRef]
- Shigesada, N.; Akira, O. Analysis of the self-shading effect on algal vertical distribution in natural waters. J. Math. Biol. 1981, 12, 311–326. [Google Scholar] [CrossRef]
- Trybula, E.M.; Cibin, R.; Burks, J.L.; Chaubey, I.; Brouder, S.M.; Volenec, J.J. Perennial rhizomatous grasses as bioenergy feedstock in SWAT: Parameter development and model improvement. Gcb Bioenergy 2015, 7, 1185–1202. [Google Scholar] [CrossRef]
- Bowes, M.J.; Loewenthal, M.; Read, D.S.; Hutchins, M.G.; Prudhomme, C.; Armstrong, L.K.; Harman, S.A.; Wickham, H.D.; Gozzard, E.; Carvalho, L. Identifying multiple stressor controls on phytoplankton dynamics in the River Thames (UK) using high-frequency water quality data. Sci. Total Environ. 2016, 569–570, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.T. Dynamics of a phytoplankton community in an ice-covered lake1. Limnol. Oceanogr. 1964, 9, 163–178. [Google Scholar] [CrossRef]
- Radwan, M.; Willems, P.; El-Sadek, A.; Berlamont, J. Modelling of dissolved oxygen and biochemical oxygen de-mand in river water using a detailed and a simplified model. Int. J. River Basin Manag. 2003, 1, 97–103. [Google Scholar] [CrossRef]
- Scheffer, M.; Rinaldi, S.; Huisman, J.; Weissing, F.J. Why plankton communities have no equilibrium: Solutions to the paradox. Hydrobiol. 2003, 491, 9–18. [Google Scholar] [CrossRef]
- Kruk, C.; Peeters, E.T.H.M.; Van Nes, E.H.; Huszar, V.L.M.; Costa, L.S.; Scheffer, M. Phytoplankton community com-position can be predicted best in terms of morphological groups. Limnol. Oceanogr. 2011, 56, 110–118. [Google Scholar] [CrossRef]
- Striebel, M.; Schabhuttl, S.; Hodapp, D.; Hinsamer, P.; Hillebrand, H. Phytoplankton responses to temperature in-creases are constrained by abiotic conditions and community composition. Oecologia 2016, 182, 815–827. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Application Characteristics | Beaver River, Canada | Trent River, UK |
|---|---|---|
| Outflow Location | 44°25′56.32″ N 79°10′0.89″ W | 53°14′43.27″ N 0°46′37.26″ W |
| Köppen-Geiger Climate Zone | DfB | CfB |
| Catchment Area (km2) | 327 | 8500 |
| 2015 Average annual flow (m3/s) | 2.9 | 88.9 |
| Sub catchments modelled | 26 | 20 |
| Agricultural land use (%) | 65 | 35 |
| Urban area (%) | 5 | 15 |
| Groundwater contribution (%) | 65% | 63% |
| Average summer temp (°C) | 18.9 | 16.4 |
| Average winter temp (°C) | −5.9 | 4.5 |
| Total annual rainfall (mm) | 777 | 747 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crossman, J.; Bussi, G.; Whitehead, P.G.; Butterfield, D.; Lannergård, E.; Futter, M.N. A New, Catchment-Scale Integrated Water Quality Model of Phosphorus, Dissolved Oxygen, Biochemical Oxygen Demand and Phytoplankton: INCA-Phosphorus Ecology (PEco). Water 2021, 13, 723. https://doi.org/10.3390/w13050723
Crossman J, Bussi G, Whitehead PG, Butterfield D, Lannergård E, Futter MN. A New, Catchment-Scale Integrated Water Quality Model of Phosphorus, Dissolved Oxygen, Biochemical Oxygen Demand and Phytoplankton: INCA-Phosphorus Ecology (PEco). Water. 2021; 13(5):723. https://doi.org/10.3390/w13050723
Chicago/Turabian StyleCrossman, Jill, Gianbattista Bussi, Paul G. Whitehead, Daniel Butterfield, Emma Lannergård, and Martyn N. Futter. 2021. "A New, Catchment-Scale Integrated Water Quality Model of Phosphorus, Dissolved Oxygen, Biochemical Oxygen Demand and Phytoplankton: INCA-Phosphorus Ecology (PEco)" Water 13, no. 5: 723. https://doi.org/10.3390/w13050723
APA StyleCrossman, J., Bussi, G., Whitehead, P. G., Butterfield, D., Lannergård, E., & Futter, M. N. (2021). A New, Catchment-Scale Integrated Water Quality Model of Phosphorus, Dissolved Oxygen, Biochemical Oxygen Demand and Phytoplankton: INCA-Phosphorus Ecology (PEco). Water, 13(5), 723. https://doi.org/10.3390/w13050723