
MAPKs and HSPs’ Activation of a Natural Daphnia magna Population in a Man-Perturbed Lake: Implications of Ecological Significance


 ,
, 

Abstract
1. Introduction
2. Materials and Methods
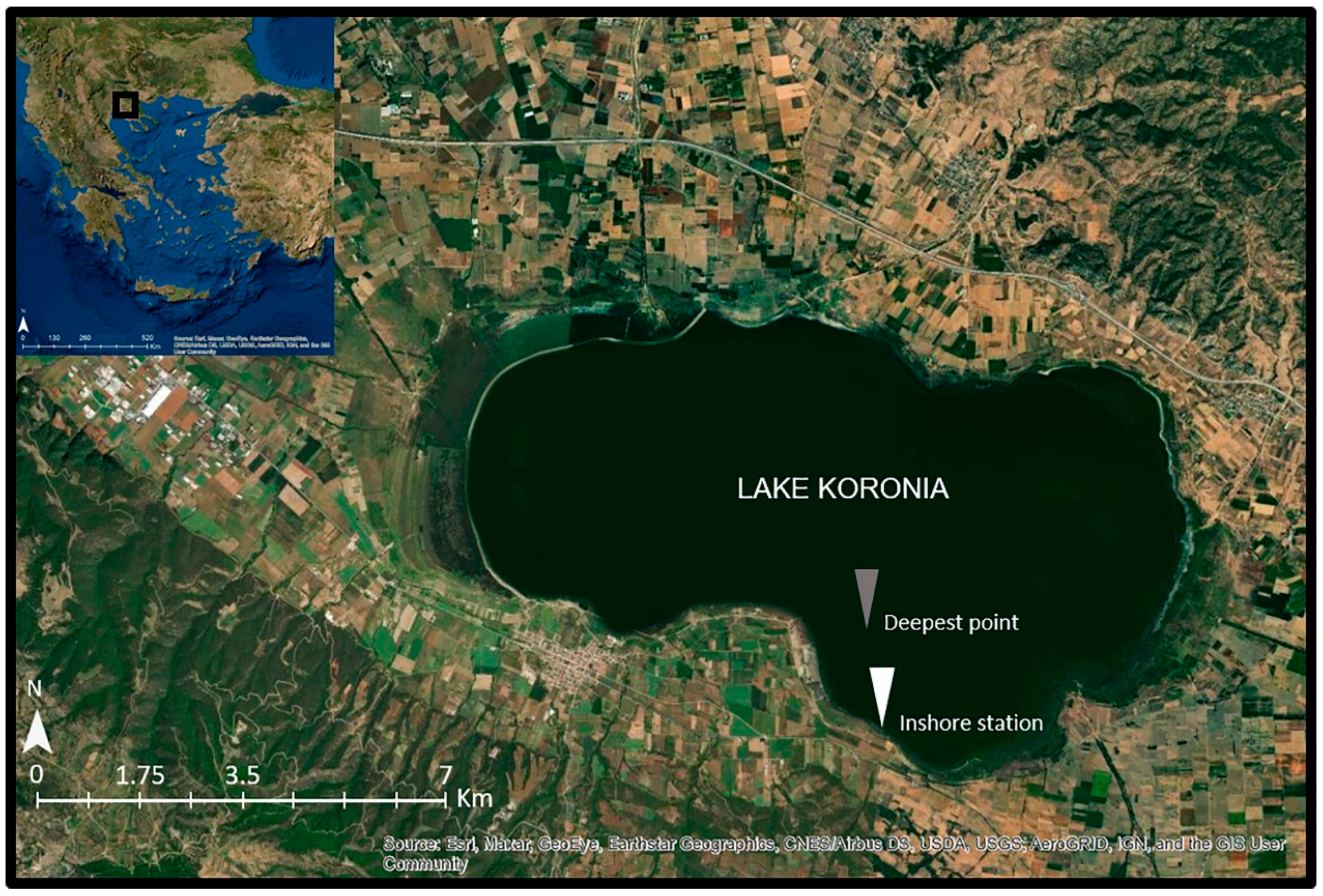
2.1. Study Area
2.2. Sample Selection in Field and Analysis
2.2.1. Water Physicochemical Parameters, Phytoplankton and Zooplankton Analysis
2.2.2. Daphnia magna Western Blot Analysis
2.3. Statistical Analysis
3. Results
3.1. Environmental Parameters
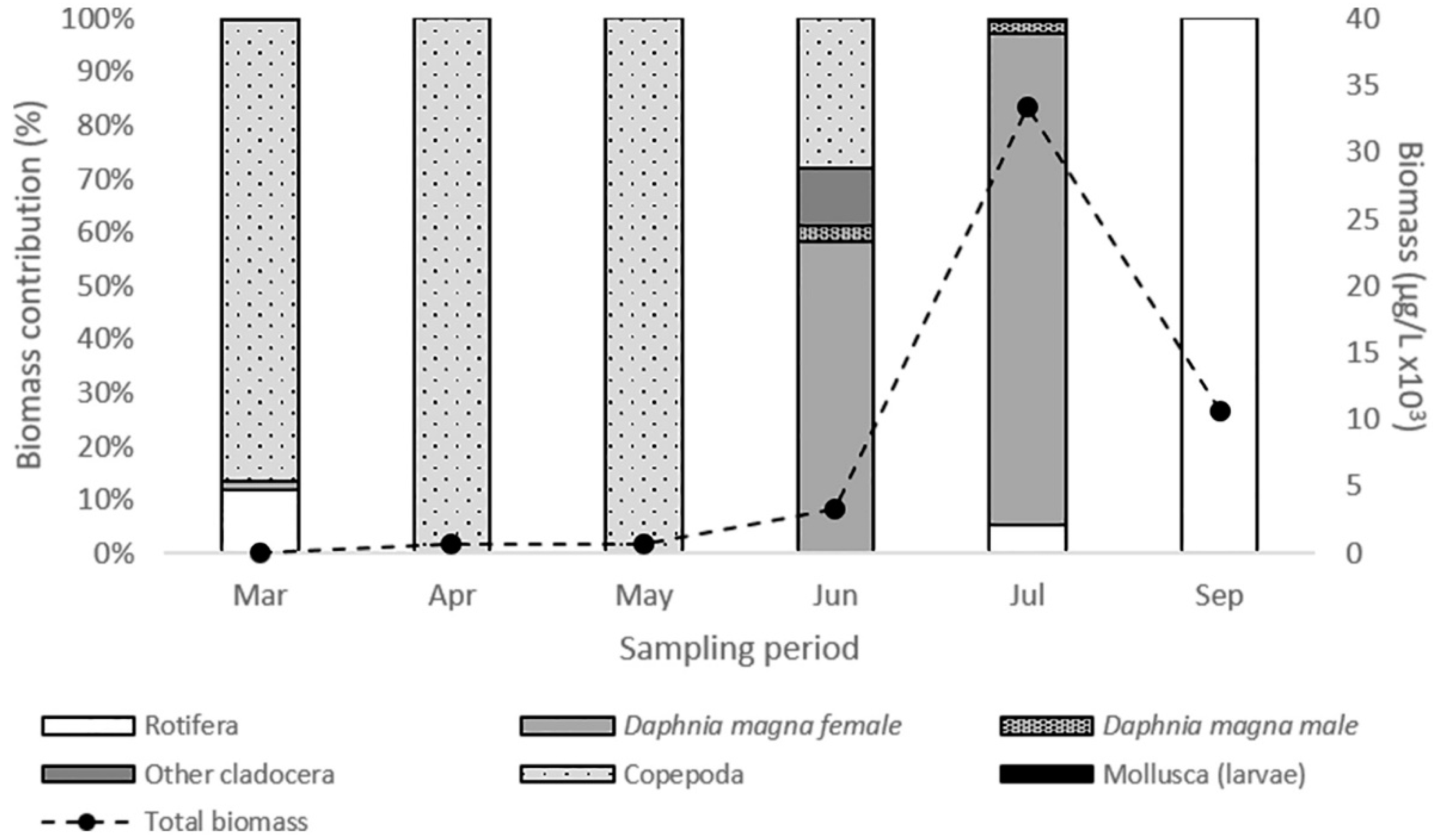
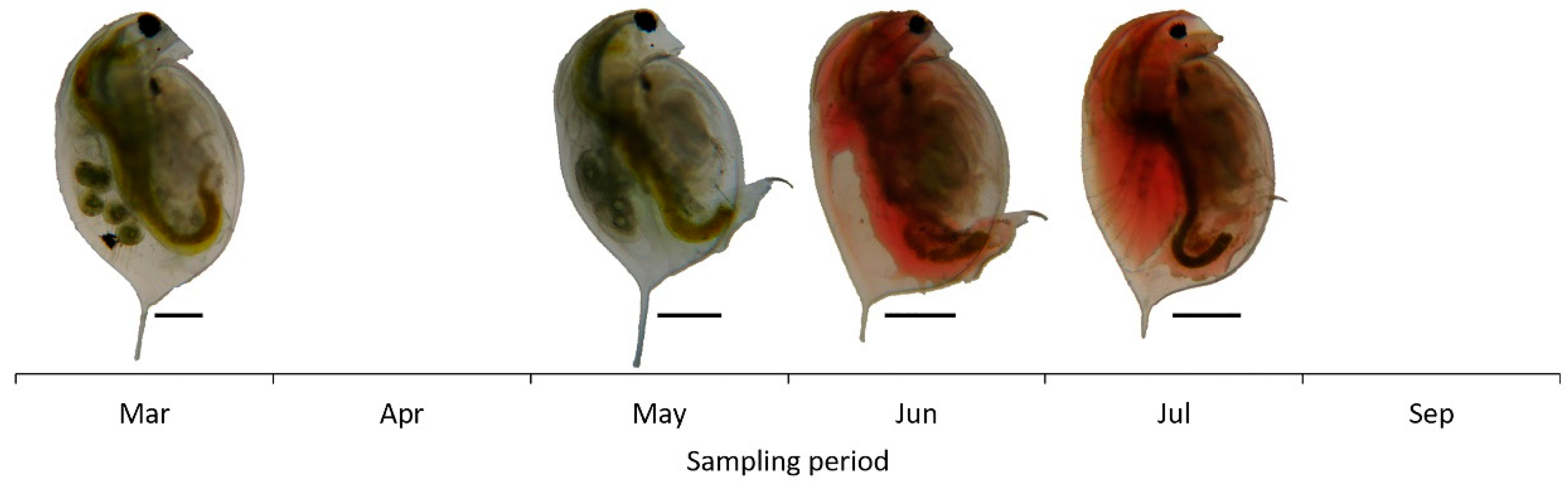
3.2. Daphnia magna and Zooplankton Community Structure
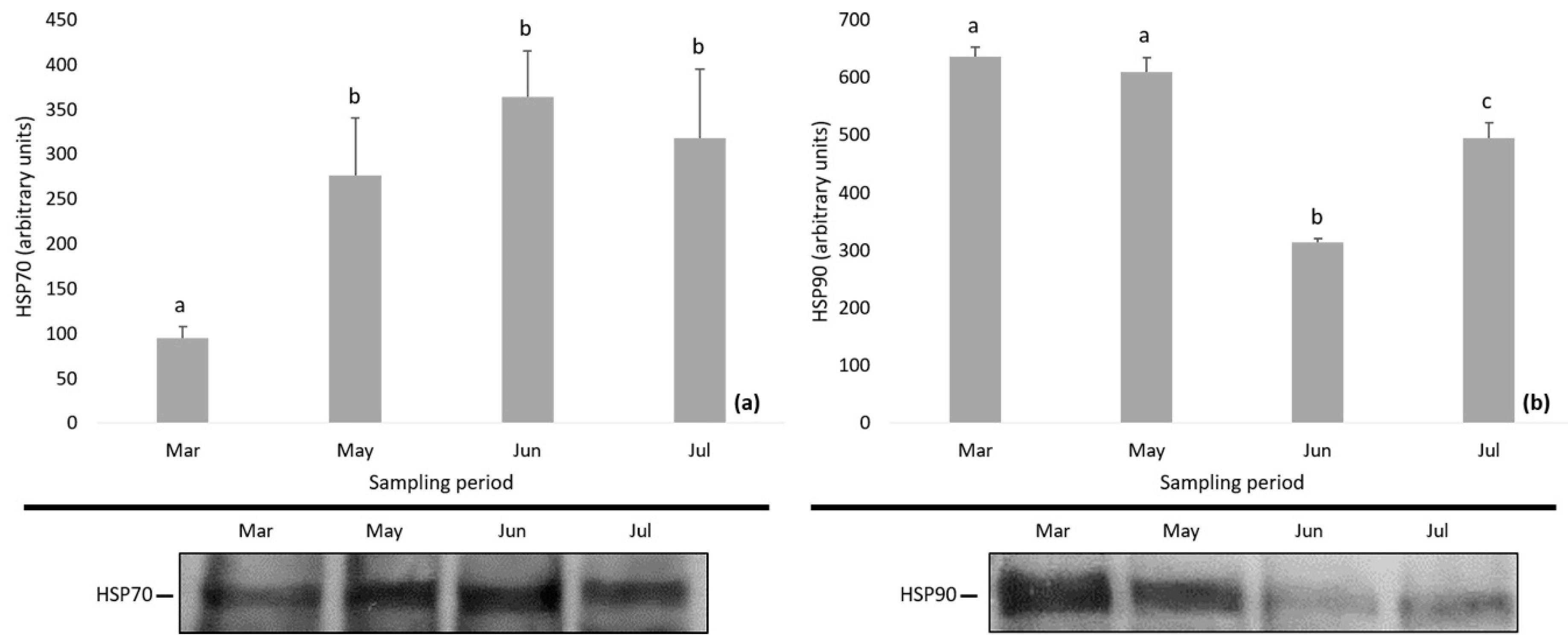
3.3. Protein Expression
3.4. Interrelationships: Molecural Biomarkers and Environmental Parameters
4. Discussion
4.1. Daphnia magna in the Zooplankton Community
4.2. Changes in Daphnia magna Protein Expression
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smolders, R.; Baillieul, M.; Blust, R. Relationship between the Energy Status of Daphnia magna and Its Sensitivity to Environmental Stress. Aquat. Toxicol. 2005, 73, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.R.; Graham, M.D.; Vinebrooke, R.D.; Findlay, D.L.; Paterson, M.J.; Turner, M.A. Multiple Anthropogenic Stressors Cause Ecological Surprises in Boreal Lakes. Glob. Chang. Biol. 2006, 12, 2316–2322. [Google Scholar] [CrossRef]
- Paerl, H.W.; Valdes, L.M.; Peierls, B.L.; Adolf, J.E.; Harding, L.J.W. Anthropogenic and Climatic Influences on the Eutrophication of Large Estuarine Ecosystems. Limnol. Oceanogr. 2006, 51, 448–462. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Sommer, U. Effects of Harmful Blooms of Large-Sized and Colonial Cyanobacteria on Aquatic Food Webs. Water 2020, 12, 1587. [Google Scholar] [CrossRef]
- Lampert, W.; Sommer, U. Limnoecology: The Ecology of Lakes and Streams, 2nd ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Lampert, W. Daphnia: Development of a Model Organism in Ecology and Evolution. Excell. Ecol. 2011, 21, 1–275. [Google Scholar]
- Tatarazako, N.; Oda, S. The Water Flea Daphnia magna (Crustacea, Cladocera) as a Test Species for Screening and Evaluation of Chemicals with Endocrine Disrupting Effects on Crustaceans. Ecotoxicology 2007, 16, 197–203. [Google Scholar] [CrossRef]
- Elser, J.J. The Pathway to Noxious Cyanobacteria Blooms in Lakes: The Food Web as the Final Turn. Freshw. Biol. 1999, 42, 537–543. [Google Scholar] [CrossRef]
- Lampert, W. Daphnia: Model Herbivore, Predator and Prey. Pol. J. Ecol. 2006, 54, 607–620. [Google Scholar]
- Miner, B.E.; De Meester, L.; Pfrender, M.E.; Lampert, W.; Hairston, N.G. Linking Genes to Communities and Ecosystems: Daphnia as an Ecogenomic Model. Proc. Biol. Sci. 2012, 279, 1873–1882. [Google Scholar] [CrossRef]
- Orsini, L.; Gilbert, D.; Podicheti, R.; Jansen, M.; Brown, J.B.; Solari, O.S.; Spanier, K.I.; Colbourne, J.K.; Rusch, D.B.; Decaestecker, E.; et al. Daphnia magna Transcriptome by RNA-Seq across 12 Environmental Stressors. Sci. Data 2016, 3, 160030. [Google Scholar] [CrossRef]
- Dalla Bona, M.; Di Leva, V.; De Liguoro, M. The Sensitivity of Daphnia magna and Daphnia curvirostris to 10 Veterinary Antibacterials and to Some of Their Binary Mixtures. Chemosphere 2014, 115, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Mikulski, A.; Grzesiuk, M.; Kloc, M.; Pijanowska, J. Heat Shock Proteins in Daphnia Detected Using Commercial Antibodies: Description and Responsiveness to Thermal Stress. Chemoecology 2009, 19, 69. [Google Scholar] [CrossRef]
- Coors, A.; Hammers-Wirtz, M.; Ratte, H.T. Adaptation to Environmental Stress in Daphnia magna Simultaneously Exposed to a Xenobiotic. Chemosphere 2004, 56, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Altshuler, I.; McLeod, A.M.; Colbourne, J.K.; Yan, N.D.; Cristescu, M.E. Synergistic Interactions of Biotic and Abiotic Environmental Stressors on Gene Expression. Genome 2015, 58, 99–109. [Google Scholar] [CrossRef]
- ISO 10706:2000. Water Quality—Determination of Long Term Toxicity of Substances to Daphnia Magna Straus (Cladocera, Crustacea); ISO: Genève, Switzerland, 2000. [Google Scholar]
- OECD 202: 2004. Guideline for Testing of Chemicals. Daphnia sp., Acute Immobilisation Test; OECD: Paris, France, 2004. [Google Scholar]
- ISO 6341:2012. Water Quality—Determination of the Inhibition of the Mobility of Daphnia magna Straus (Cladocera, Crustacea)—Acute Toxicity Test; ISO: Genève, Switzerland, 2012. [Google Scholar]
- OECD 211: 2012. Guidelines for the Testing of Chemicals. Daphnia magna Reproduction Test; OECD: Paris, France, 2012. [Google Scholar]
- Pijanowska, J.; Kloc, M. Daphnia Response to Predation Threat Involves Heat-Shock Proteins and the Actin and Tubulin Cytoskeleton. Genesis 2004, 38, 81–86. [Google Scholar] [CrossRef]
- Pauwels, K.; Stoks, R.; de Meester, L. Coping with Predator Stress: Interclonal Differences in Induction of Heat-Shock Proteins in the Water Flea Daphnia magna. J. Evol. Biol. 2005, 18, 867–872. [Google Scholar] [CrossRef]
- Mikulski, A.; Bernatowicz, P.; Grzesiuk, M.; Kloc, M.; Pijanowska, J. Differential Levels of Stress Proteins (HSPs) in Male and Female Daphnia magna in Response to Thermal Stress: A Consequence of Sex-Related Behavioral Differences? J. Chem. Ecol. 2011, 37, 670–676. [Google Scholar] [CrossRef]
- Mikulski, A.; Grzesiuk, M.; Rakowska, A.; Bernatowicz, P.; Pijanowska, J. Thermal Shock in Daphnia: Cost of Diel Vertical Migrations or Inhabiting Thermally-Unstable Waterbodies? Fundam. Appl. Limnol. 2017, 213–220. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The Evolutionary and Ecological Role of Heat Shock Proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Schumpert, C.A.; Anderson, C.; Dudycha, J.L.; Patel, R.C. Involvement of Daphnia pulicaria Sir2 in Regulating Stress Response and Lifespan. Aging 2016, 8, 402–417. [Google Scholar] [CrossRef][Green Version]
- Dahlhoff, E.P. Biochemical Indicators of Stress and Metabolism: Applications for Marine Ecological Studies. Annu. Rev. Physiol. 2004, 66, 183–207. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, J.; Duan, Y.; Chen, P.; Liu, P. The Roles of Heat Shock Proteins 70 and 90 in Exopalaemon carinicauda After WSSV and Vibrio anguillarum Challenges. J. Ocean Univ. China 2018, 17, 399–406. [Google Scholar] [CrossRef]
- Antonopoulou, E.; Kentepozidou, E.; Feidantsis, K.; Roufidou, C.; Despoti, S.; Chatzifotis, S. Starvation and Re-Feeding Affect Hsp Expression, MAPK Activation and Antioxidant Enzymes Activity of European Sea Bass (Dicentrarchus labrax). Comp. Biochem. Physiol. A 2013, 165, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-M.; Jeong, C.-B.; Kim, M.-S.; Lee, J.-S.; Zhou, J.; Lee, Y.H.; Kim, D.-H.; Moon, E.; Kweon, H.-S.; Lee, S.-J.; et al. The Role of the P38-Activated Protein Kinase Signaling Pathway-Mediated Autophagy in Cadmium-Exposed Monogonont Rotifer. Aquat. Toxicol. 2018, 194, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Widmann, C.; Gibson, S.; Jarpe, M.B.; Johnson, G.L. Mitogen-Activated Protein Kinase: Conservation of a Three-Kinase Module from Yeast to Human. Physiol. Rev. 1999, 79, 143–180. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, Y.; Jiao, Y.; Chen, Q.; Wu, D.; Yu, P.; Li, Y.; Cai, M.; Zhao, Y. Polystyrene Nanoplastic Induces ROS Production and Affects the MAPK-HIF-1/NFkB-Mediated Antioxidant System in Daphnia pulex. Aquat. Toxicol. 2020, 220, 105420. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M.; Avruch, J. Sounding the Alarm: Protein Kinase Cascades Activated by Stress and Inflammation. J. Biol. Chem. 1996, 271, 24313–24316. [Google Scholar] [CrossRef]
- Roux, P.P.; Blenis, J. ERK and P38 MAPK-Activated Protein Kinases: A Family of Protein Kinases with Diverse Biological Functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef]
- Lin, W.; Jiang, R.; Hu, S.; Xiao, X.; Wu, J.; Wei, S.; Xiong, Y.; Ouyang, G. Investigating the Toxicities of Different Functionalized Polystyrene Nanoplastics on Daphnia magna. Ecotoxicol. Environ. Saf. 2019, 180, 509–516. [Google Scholar] [CrossRef]
- Haap, T.; Köhler, H.-R. Cadmium Tolerance in Seven Daphnia magna Clones Is Associated with Reduced Hsp70 Baseline Levels and Induction. Aquat. Toxicol. 2009, 94, 131–137. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Michaloudi, E.; Kormas, K.A.; Katsiapi, M.; Vardaka, E.; Genitsaris, S. Plankton Changes as Critical Processes for Restoration Plans of Lakes Kastoria and Koronia. Eur. Water 2012, 40, 43–51. [Google Scholar]
- Michaloudi, E.; Moustaka-Gouni, M.; Pantelidakis, K.; Katsiapi, M.; Genitsaris, S. Plankton Succession in the Temporary Lake Koronia after Intermittent Dry-Out. Int. Rev. Hydrobiol. 2012, 97, 405–419. [Google Scholar] [CrossRef]
- Michaloudi, E.; Moustaka-Gouni, M.; Gkelis, S.; Pantelidakis, K. Plankton Community Structure during an Ecosystem Disruptive Algal Bloom of Prymnesium parvum. J. Plankton Res. 2009, 31, 301–309. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Cook, C.M.; Gkelis, S.; Michaloudi, E.; Pantlelidakis, K.; Pyrovetsi, M.; Lanaras, T. The Coincidence of a Prymnesium parvum Bloom and the Mass Kill of Birds and Fish in Lake Koronia. Harmful Algae News 2004, 26, 1–2. [Google Scholar]
- Malamataris, D.; Kolokytha, E.; Mylopoulos, I.; Loukas, A. Critical Review of Adaptation Strategies for the Restoration of Lake Koronia. Eur. Water 2017, 58, 203–208. [Google Scholar]
- Kolokytha, E.; Malamataris, D. Integrated Water Management Approach for Adaptation to Climate Change in Highly Water Stressed Basins. Water Resour. Manag. 2020, 34, 1173–1197. [Google Scholar] [CrossRef]
- Bobori, D.C.; Economidis, P.S. The Effect of Size, Sex and Season on the Accumulation of Heavy Metals in Perch (Perca fluviatilis L., Pisces: Cyprinidae) in Lake Koronia (Macedonia, Greece). Toxicol. Environ. Chem. 1996, 57, 103–121. [Google Scholar] [CrossRef]
- Kaiserli, A.; Voutsa, D.; Samara, C. Phosphorus Fractionation in Lake Sediments--Lakes Volvi and Koronia, N. Greece. Chemosphere 2002, 46, 1147–1155. [Google Scholar] [CrossRef]
- Petaloti, C.; Voutsa, D.; Samara, C.; Sofoniou, M.; Stratis, I.; Kouimtzis, T. Nutrient Dynamics in Shallow Lakes of Northern Greece. Environ. Sci. Pollut. Res. 2004, 11, 11. [Google Scholar] [CrossRef]
- Papadaki, E.; Voutsa, D. Arsenic Cycling in Lakes Volvi and Koronia, Northern Greece. Assessment of Arsenic Mobility in Sediments. Fresenius Environ. Bull. 2007, 16, 421–427. [Google Scholar]
- Anthemidis, A.; Zachariadis, G.; Voutsa, D.; Kouras, A.; Samara, C. Variation of Heavy Metal and Other Toxic Element Concentrations in Surface Waters of Macedonia. In Proceedings of the 1st Environmental Conference of Macedonia, Thessaloniki, Greece, 1 March 2002; pp. 104–109. [Google Scholar]
- Kaloyianni, M.; Feidantsis, K.; Nteli, I.; Stergiou, P.; Tsoulia, T.; Dimitriadi, A.; Antonopoulou, E.; Bobori, D. Biochemical and Molecular Responses of Cyprinids in Two Mediterranean Lacustrine Ecosystems: Opportunities for Ecological Assessment and Biomonitoring. Aquat. Toxicol. 2019, 211, 105–115. [Google Scholar] [CrossRef] [PubMed]
- ISO 5667-3:2018. Water Quality–Sampling-Part 3: Preservation and Handling of Water Samples; ISO: Genève, Switzerland, 2018. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater; APHA: Washington, DC, USA, 2012. [Google Scholar]
- Bottrell, H.H.; Duncan, A.; Gliwicz, Z.M.; Grygierek, E.; Herzig, A.; Hillbright-Ilkowska, A.; Kurasawa, H.; Larsson, P.; Weglenska, T.A. A Review of Some Problems in Zooplankton Production Studies. Nor. J. Zool. 1976, 24, 419–456. [Google Scholar]
- Downing, J.A.; Rigler, F.H. A Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters; Blackwell Scientific Publications: Oxford, UK, 1984. [Google Scholar]
- Taggart, C.T. Hypolimnetic Aeration and Zooplankton Distribution: A Possible Limitation to the Restoration of Cold-Water Fish Production. Can. J. Fish. Aquat. Sci. 1984, 41, 191–198. [Google Scholar] [CrossRef]
- Herzig, A. Some Population Characteristics of Planktonic Crustaceans in Neusiedler See. Oecologia 1974, 15, 127–141. [Google Scholar] [CrossRef]
- Dumont, H.J.; Van de Velde, I.; Dumont, S. The Dry Weight Estimate of Biomass in a Selection of Cladocera, Copepoda and Rotifera from the Plankton, Periphyton and Benthos of Continental Waters. Oecologia 1975, 19, 75–97. [Google Scholar] [CrossRef]
- Zarfdjian, M.H. Seasonal Variations and Spatial Distribution of Planktic Invertebrates in Lake Volvi (Macedonia Greece). Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1989. [Google Scholar]
- Michaloudi, E. Dry Weights of the Zooplankton of Lake Mikri Prespa (Macedonia, Greece). Belg. J. Zool. 2005, 135, 223–227. [Google Scholar]
- Romero-Calvo, I.; Ocón, B.; Martínez-Moya, P.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; de Medina, F.S. Reversible Ponceau Staining as a Loading Control Alternative to Actin in Western Blots. Anal. Biochem. 2010, 401, 318–320. [Google Scholar] [CrossRef]
- Liu, X.; Dur, G.; Ban, S.; Sakai, Y.; Ohmae, S.; Morita, T. Planktivorous Fish Predation Masks Anthropogenic Disturbances on Decadal Trends in Zooplankton Biomass and Body Size Structure in Lake Biwa, Japan. Limnol. Oceanogr. 2020, 65, 667–682. [Google Scholar] [CrossRef]
- Choi, J.-Y.; Kim, S.-K.; Chang, K.-H.; Kim, M.-C.; La, G.-H.; Joo, G.-J.; Jeong, K.-S. Population Growth of the Cladoceran, Daphnia magna: A Quantitative Analysis of the Effects of Different Algal Food. PLoS ONE 2014, 9, e95591. [Google Scholar] [CrossRef][Green Version]
- Michaloudi, E.; Kostecka, M. Zooplankton of Lake Koroneia (Macedonia, Greece). Biologia 2004, 59, 165–172. [Google Scholar]
- Gilbert, J.J. Suppression of Rotifer Populations by Daphnia: A Review of the Evidence, the Mechanisms, and the Effects on Zooplankton Community Structure1. Limnol. Oceanogr. 1988, 33, 1286–1303. [Google Scholar] [CrossRef]
- Ebert, D. Ecology, Epidemiology, and Evolution of Parasitism in Daphnia; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2005. [Google Scholar]
- Decaestecker, E.; De Meester, L.; Mergeay, J. Cyclical Parthenogenesis in Daphnia: Sexual Versus Asexual Reproduction. In Lost Sex: The Evolutionary Biology of Parthenogenesis; Schön, I., Martens, K., Dijk, P., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 295–316. [Google Scholar] [CrossRef]
- Pijanowska, J.; Stolpe, G. Summer Diapause in Daphnia as a Reaction to the Presence of Fish. J. Plankton Res. 1996, 18, 1407–1412. [Google Scholar] [CrossRef][Green Version]
- Oda, S.; Tatarazako, N.; Watanabe, H.; Morita, M.; Iguchi, T. Production of Male Neonates in Daphnia magna (Cladocera, Crustacea) Exposed to Juvenile Hormones and Their Analogs. Chemosphere 2005, 61, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, E.; Søndergaard, M.; Jensen, J.P.; Mortensen, E.; Hansen, A.-M.; Jørgensen, T. Cascading Trophic Interactions from Fish to Bacteria and Nutrients after Reduced Sewage Loading: An 18-Year Study of a Shallow Hypertrophic Lake. Ecosystems 1998, 1, 250–267. [Google Scholar] [CrossRef]
- Ghazy, M.M.E.-D.; Habashy, M.M.; Mohammady, E.Y. Effects of PH on Survival, Growth and Reproduction Rates of the Crustacean. Aust. J. Basic Appl. Sci. 2011, 5, 1–10. [Google Scholar]
- Roelke, D.L.; Errera, R.M.; Riesling, R.; Brooks, B.W.; Grover, J.P.; Schwierzke, L.; Urena-Boeck, F.; Baker, J.; Pinckney, J.L. Effects of Nutrient Enrichment on Prymnesium parvum Population Dynamics and Toxicity: Results from Field Experiments, Lake Possum Kingdom, USA. Aquat. Microb. Ecol. 2007, 46, 16. [Google Scholar] [CrossRef]
- Luu, P.T.; Yen, T.T.H.; Thai, T.T.; Quang, N.X. Effects of Non-Toxic Filamentous Cyanobacteria Isolated from Tri an Reservoir on Daphnia. Acad. J. Biol. 2020, 42. [Google Scholar] [CrossRef]
- Brooks, J.L.; Dodson, S.I. Predation, Body Size, and Composition of Plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef]
- Pirow, R.; Bäumer, C.; Paul, R.J. Benefits of Haemoglobin in the Cladoceran Crustacean Daphnia magna. J. Exp. Biol. 2001, 204, 3425–3441. [Google Scholar]
- Gerke, P.; Börding, C.; Zeis, B.; Paul, R.J. Adaptive Haemoglobin Gene Control in Daphnia pulex at Different Oxygen and Temperature Conditions. Comp. Biochem. Physiol. A 2011, 159, 56–65. [Google Scholar] [CrossRef]
- Cambronero, M.C.; Zeis, B.; Orsini, L. Haemoglobin-mediated Response to Hyper-thermal Stress in the Keystone Species Daphnia magna. Evol. Appl. 2017, 11, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Gonoi, H. Horizontal Movement of Pale and Red Daphnia magna in Low Oxygen Concentration. Physiol. Zool. 1985, 58, 190–196. [Google Scholar] [CrossRef]
- Schumpert, C.; Handy, I.; Dudycha, J.L.; Patel, R.C. Relationship between Heat Shock Protein 70 Expression and Life Span in Daphnia. Mech. Ageing Dev. 2014, 139, 1–10. [Google Scholar] [CrossRef][Green Version]
- Lyu, K.; Wang, Q.; Li, Z.; Chen, R.; Zhu, C.; Liu, J.; Yang, Z. Age-Dependent Survival and Selected Gene Expression in Daphnia magna after Short-Term Exposure to Low Dissolved Oxygen. J. Plankton Res. 2015, 37, 66–74. [Google Scholar] [CrossRef][Green Version]
- Haap, T.; Schwarz, S.; Köhler, H.-R. Metallothionein and Hsp70 Trade-off against One Another in Daphnia magna Cross-Tolerance to Cadmium and Heat Stress. Aquat. Toxicol. 2016, 170, 112–119. [Google Scholar] [CrossRef]
- Wilczynski, W.; Dynak, P.; Babkiewicz, E.; Bernatowicz, P.; Leniowski, K.; Maszczyk, P. The Combined Effects of Hypoxia and Fish Kairomones on Several Physiological and Life History Traits of Daphnia. Freshw. Biol. 2019, 64, 2204–2220. [Google Scholar] [CrossRef]
- Pauwels, K.; Stoks, R.; Meester, L.D. Enhanced Anti-Predator Defence in the Presence of Food Stress in the Water Flea Daphnia magna. Funct. Ecol. 2010, 24, 322–329. [Google Scholar] [CrossRef]
- Effertz, C.; von Elert, E. Light Intensity Controls Anti-Predator Defences in Daphnia: The Suppression of Life-History Changes. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133250. [Google Scholar] [CrossRef]
- Klumpen, E.; Hoffschröer, N.; Zeis, B.; Gigengack, U.; Dohmen, E.; Paul, R.J. Reactive Oxygen Species (ROS) and the Heat Stress Response of Daphnia pulex: ROS-Mediated Activation of Hypoxia-Inducible Factor 1 (HIF-1) and Heat Shock Factor 1 (HSF-1) and the Clustered Expression of Stress Genes. Biol. Cell 2017, 109, 39–64. [Google Scholar] [CrossRef]
- Gliwicz, Z.M. Filtering Rates, Food Size Selection, and Feeding Rates in Cladocerans—Another Aspect of Interspecific Competition in Filter-Feeding Zooplankton. In Evolution and Ecology of Zooplankton Communities; Kerfoot, W.C., Ed.; The University Press of New England: Hanover, NH, USA, 1980; pp. 282–291. [Google Scholar]
- Sarpe, D.; de Senerpont Domis, L.N.; Declerck, S.A.J.; van Donk, E.; Ibelings, B.W. Food Quality Dominates the Impact of Food Quantity on Daphnia Life History: Possible Implications for Re-Oligotrophication. Inland Waters 2014, 4, 363–368. [Google Scholar] [CrossRef]
- Coors, A.; Meester, L.D. Synergistic, Antagonistic and Additive Effects of Multiple Stressors: Predation Threat, Parasitism and Pesticide Exposure in Daphnia magna. J. Appl. Ecol. 2008, 45, 1820–1828. [Google Scholar] [CrossRef]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP Kinase Signalling Pathways in Cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.-B.; Won, E.-J.; Kang, H.-M.; Lee, M.-C.; Hwang, D.-S.; Hwang, U.-K.; Zhou, B.; Souissi, S.; Lee, S.-J.; Lee, J.-S. Microplastic Size-Dependent Toxicity, Oxidative Stress Induction, and p-JNK and p-P38 Activation in the Monogonont Rotifer (Brachionus koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-M.; Rhee, J.-S.; Lee, K.-W.; Kim, M.-J.; Shin, K.-H.; Lee, S.-J.; Lee, Y.-M.; Lee, J.-S. UV-B Radiation-Induced Oxidative Stress and P38 Signaling Pathway Involvement in the Benthic Copepod Tigriopus japonicus. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2015, 167, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-H.; Puthumana, J.; Kang, H.-M.; Lee, M.-C.; Jeong, C.-B.; Han, J.; Hwang, D.-S.; Kim, I.-C.; Lee, J.W.; Lee, J.-S. Adverse Effects of MWCNTs on Life Parameters, Antioxidant Systems, and Activation of MAPK Signaling Pathways in the Copepod Paracyclopina nana. Aquat. Toxicol. 2016, 179, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-C.; Puthumana, J.; Lee, S.-H.; Kang, H.-M.; Park, J.C.; Jeong, C.-B.; Han, J.; Hwang, D.-S.; Seo, J.S.; Park, H.G.; et al. BDE-47 Induces Oxidative Stress, Activates MAPK Signaling Pathway, and Elevates de Novo Lipogenesis in the Copepod Paracyclopina nana. Aquat. Toxicol. 2016, 181, 104–112. [Google Scholar] [CrossRef]
- Jeong, C.-B.; Kang, H.-M.; Lee, M.-C.; Kim, D.-H.; Han, J.; Hwang, D.-S.; Souissi, S.; Lee, S.-J.; Shin, K.-H.; Park, H.G.; et al. Adverse Effects of Microplastics and Oxidative Stress-Induced MAPK/Nrf2 Pathway-Mediated Defense Mechanisms in the Marine Copepod Paracyclopina nana. Sci. Rep. 2017, 7, 41323. [Google Scholar] [CrossRef]
- Uehara, T.; Kaneko, M.; Tanaka, S.; Okuma, Y.; Nomura, Y. Possible Involvement of P38 MAP Kinase in HSP70 Expression Induced by Hypoxia in Rat Primary Astrocytes. Brain Res. 1999, 823, 226–230. [Google Scholar] [CrossRef]
- Rafiee, P.; Theriot, M.E.; Nelson, V.M.; Heidemann, J.; Kanaa, Y.; Horowitz, S.A.; Rogaczewski, A.; Johnson, C.P.; Ali, I.; Shaker, R.; et al. Human Esophageal Microvascular Endothelial Cells Respond to Acidic PH Stress by PI3K/AKT and P38 MAPK-Regulated Induction of Hsp70 and Hsp27. Am. J. Physiol. Cell Physiol. 2006, 291, C931–C945. [Google Scholar] [CrossRef]
- Bironaite, D.; Brunk, U.; Venalis, A. Protective Induction of Hsp70 in Heat-Stressed Primary Myoblasts: Involvement of MAPKs. J. Cell. Biochem. 2013, 114, 2024–2031. [Google Scholar] [CrossRef]
- Feidantsis, K.; Pörtner, H.O.; Markou, T.; Lazou, A.; Michaelidis, B. Involvement of P38 MAPK in the Induction of Hsp70 during Acute Thermal Stress in Red Blood Cells of the Gilthead Sea Bream, Sparus aurata. J. Exp. Zool. Part Ecol. Genet. Physiol. 2012, 317, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Kefaloyianni, E.; Gourgou, E.; Ferle, V.; Kotsakis, E.; Gaitanaki, C.; Beis, I. Acute Thermal Stress and Various Heavy Metals Induce Tissue-Specific pro- or Anti-Apoptotic Events via the P38-MAPK Signal Transduction Pathway in Mytilus galloprovincialis (Lam.). J. Exp. Biol. 2005, 208 Pt 23, 4427–4436. [Google Scholar] [CrossRef] [PubMed]
- Sheikh-Hamad, D.; Di Mari, J.; Suki, W.N.; Safirstein, R.; Watts, B.A.; Rouse, D. P38 Kinase Activity Is Essential for Osmotic Induction of MRNAs for HSP70 and Transporter for Organic Solute Betaine in Madin-Darby Canine Kidney Cells. J. Biol. Chem. 1998, 273, 1832–1837. [Google Scholar] [CrossRef]
- Antonopoulou, E.; Chouri, E.; Feidantsis, K.; Lazou, A.; Chatzifotis, S. Effects of Partial Dietary Supplementation of Fish Meal with Soymeal on the Stress and Apoptosis Response in the Digestive System of Common Dentex (Dentex dentex). J. Biol. Res. Thessalon. 2017, 24, 14. [Google Scholar] [CrossRef] [PubMed]
- Escobar, M.D.C.; Souza, V.; Bucio, L.; Hernández, E.; Gómez-Quiroz, L.E.; Ruiz, M.C.G. MAPK Activation Is Involved in Cadmium-Induced Hsp70 Expression in HepG2 Cells. Toxicol. Mech. Methods 2009, 19, 503–509. [Google Scholar] [CrossRef]
- Goodman, R.; Lin-Ye, A.; Geddis, M.S.; Wickramaratne, P.J.; Hodge, S.E.; Pantazatos, S.P.; Blank, M.; Ambron, R.T. Extremely Low Frequency Electromagnetic Fields Activate the ERK Cascade, Increase Hsp70 Protein Levels and Promote Regeneration in Planaria. Int. J. Radiat. Biol. 2009, 85, 851–859. [Google Scholar] [CrossRef]
- Lim, S.; Kim, D.G.; Kim, S. ERK-Dependent Phosphorylation of the Linker and Substrate-Binding Domain of HSP70 Increases Folding Activity and Cell Proliferation. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef]
- Korpelainen, H. The Effects of Temperature and Photoperiod on Life History Parameters of Daphnia magna (Crustacea: Cladocera). Freshw. Biol. 1986, 16, 615–620. [Google Scholar] [CrossRef]
- Williams, P.J.; Dick, K.B.; Yampolsky, L.Y. Heat Tolerance, Temperature Acclimation, Acute Oxidative Damage and Canalization of Haemoglobin Expression in Daphnia. Evol. Ecol. 2012, 26, 591–609. [Google Scholar] [CrossRef]
- Al-Seria, M.H.M. Salinity Effects on Survival and Life History of Daphnia magna. Int. J. Aquat. Sci. 2019, 10, 19–26. [Google Scholar]
- Jeyasingh, P.D.; Weider, L.J. Phosphorus Availability Mediates Plasticity in Life-History Traits and Predator–Prey Interactions in Daphnia. Ecol. Lett. 2005, 8, 1021–1028. [Google Scholar] [CrossRef]
- Lyu, K.; Cao, H.; Chen, R.; Wang, Q.; Yang, Z. Combined Effects of Hypoxia and Ammonia to Daphnia similis Estimated with Life-History Traits. Environ. Sci. Pollut. Res. 2013, 20, 5379–5387. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, K.; Stoks, R.; Decaestecker, E.; De Meester, L. Evolution of Heat Shock Protein Expression in a Natural Population of Daphnia magna. Am. Nat. 2007, 170, 800–805. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| March | April | May | June | July | September | |
|---|---|---|---|---|---|---|
| Parameters | (Mar) | (Apr) | (May) | (Jun) | (Jul) | (Sep) |
| pH | 9.69 | 10.15 | 9.75 | 9.3 | 8.77 | 9.82 |
| Temp (°C) | 12 | 17.6 | 20.3 | 27.6 | 27.8 | 25.8 |
| Cond (μS cm−1) | 6220 | 6889 | 7200 | 8663 | 8800 | 12160 |
| Sal (ppt) | 4 | 4.1 | 4.4 | 4.5 | 4.6 | 7.1 |
| Max depth (m) | 1.25 | 1.3 | 1.25 | 1.1 | 1 | 0.3 |
| Phyto ab (cell mL−1 × 103) | 320.96 | 1956.74 | 965.69 | 2265.30 | 1557.90 | 1137.34 |
| Harmful phyto (cell mL−1 × 103) | 9.42 | 99.32 | 82.79 | 0 | 0 | 9.59 |
| Oscil cyan (cell mL−1 × 103) | 191.43 | 467.20 | 220.33 | 16.79 | 0.26 | 2.79 |
| Transp (Secchi m) | 0.4 | 0.3 | 0.25 | 0.25 | 0.3 | 0.05 |
| N-NO2− (mg N L−1) | 0.002 | 0.005 | 0.010 | 0.011 | 0.017 | 0.012 |
| N-NO3− (mg N L−1) | 0.204 | 0.256 | 0.363 | 0.256 | 0.210 | 0.102 |
| N-NH4+ (mg N L−1) | 0.007 | 0.028 | 0.034 | 1.00 | 1.95 | 0.25 |
| TN (mg N L−1) | 4.51 | 6.72 | 3.10 | 6.15 | 10.01 | 5.31 |
| TON (mg N L−1) | 4.293 | 6.21 | 2.70 | 5.14 | 7.84 | 4.95 |
| SRP (mg P L−1) | 0.003 | <dl | 0.008 | <dl | 0.288 | <dl |
| TP (mg P L−1) | 0.098 | 0.125 | 0.128 | 0.296 | 0.338 | 0.192 |
| Size (μm) | 701–900 | 901–1100 | 1101–1300 | 1301–1500 | 1501–1700 | 1701–1901 | 1901–2100 | 2101–2300 | 2301–2500 | 2501–2700 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sex | F | F | F | M | F | M | F | F | F | F | F | F |
| March | 0 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| June | 1.7 | 1.8 | 16.6 | 1.3 | 6.9 | 3.5 | 7.1 | 0.0 | 11.7 | 23.2 | 5.7 | 20.6 |
| July | 0 | 0 | 0 | 0 | 9.1 | 0 | 10.7 | 13.8 | 26.4 | 24.5 | 12.9 | 0 |
| HSP70 | HSP90 | Phospho p38 | Phospho p44/42 | |
|---|---|---|---|---|
| HSP70 | 1.000 | |||
| HSP90 | −1.000 ** | 1.000 | ||
| phospho p38 | −0.800 | 0.800 | 1.000 | |
| phospho p44/42 | 0.800 | −0.800 | −1.000 | 1.000 |
| pH | −0.600 | 0.600 | 0.800 | −0.800 |
| Temp | 0.800 | −0.800 | −1.000 ** | 1.000 ** |
| Cond | 0.800 | −0.800 | −1.000 ** | 1.000 ** |
| Sal | 0.800 | −0.800 | −1.000 ** | 1.000 ** |
| Max depth | −0.738 | 0.738 | 0.949 | −0.949 |
| Phyto ab | 1.000 ** | −1.000 ** | −0.800 | 0.800 |
| Harmful phyto | −0.738 | 0.738 | 0.738 | −0.738 |
| Oscil cyan | −0.600 | 0.600 | −0.800 | 0.800 |
| Transparency | −0.632 | 0.632 | 0.316 | −0.316 |
| N-NO2− | 0.800 | −0.800 | −1.000 ** | 1.000 ** |
| N-NO3− | 0.400 | −0.400 | −0.200 | 0.200 |
| N-NH4+ | 0.800 | −0.800 | −1.000 ** | 1.000 ** |
| TN | 0.600 | −0.600 | −0.800 | 0.800 |
| TON | 0.600 | −0.600 | −0.800 | 0.800 |
| SRP | −0.200 | 0.200 | −0.400 | 0.400 |
| TP | 0.800 | −0.800 | −1.000 ** | 1.000 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demertzioglou, M.; Antonopoulou, E.; Voutsa, D.; Kozari, A.; Moustaka-Gouni, M.; Michaloudi, E. MAPKs and HSPs’ Activation of a Natural Daphnia magna Population in a Man-Perturbed Lake: Implications of Ecological Significance. Water 2021, 13, 283. https://doi.org/10.3390/w13030283
Demertzioglou M, Antonopoulou E, Voutsa D, Kozari A, Moustaka-Gouni M, Michaloudi E. MAPKs and HSPs’ Activation of a Natural Daphnia magna Population in a Man-Perturbed Lake: Implications of Ecological Significance. Water. 2021; 13(3):283. https://doi.org/10.3390/w13030283
Chicago/Turabian StyleDemertzioglou, Maria, Efthimia Antonopoulou, Dimitra Voutsa, Argyri Kozari, Maria Moustaka-Gouni, and Evangelia Michaloudi. 2021. "MAPKs and HSPs’ Activation of a Natural Daphnia magna Population in a Man-Perturbed Lake: Implications of Ecological Significance" Water 13, no. 3: 283. https://doi.org/10.3390/w13030283
APA StyleDemertzioglou, M., Antonopoulou, E., Voutsa, D., Kozari, A., Moustaka-Gouni, M., & Michaloudi, E. (2021). MAPKs and HSPs’ Activation of a Natural Daphnia magna Population in a Man-Perturbed Lake: Implications of Ecological Significance. Water, 13(3), 283. https://doi.org/10.3390/w13030283