
Stress Resistance and Adaptation of the Aquatic Invasive Species Tubastraea Coccinea (Lesson, 1829) to Climate Change and Ocean Acidification


 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials, Study Organism, Collection, and Preparation
2.2. Experimental Design and Analysis
2.3. Tissue Loss Analysis
2.4. Western Blotting: Tissue Collection
2.4.1. Tissue Isolation
2.4.2. Protein Isolation
2.4.3. Gel Electrophoresis
2.5. Statistical Analysis
3. Results
3.1. Preliminary Analysis
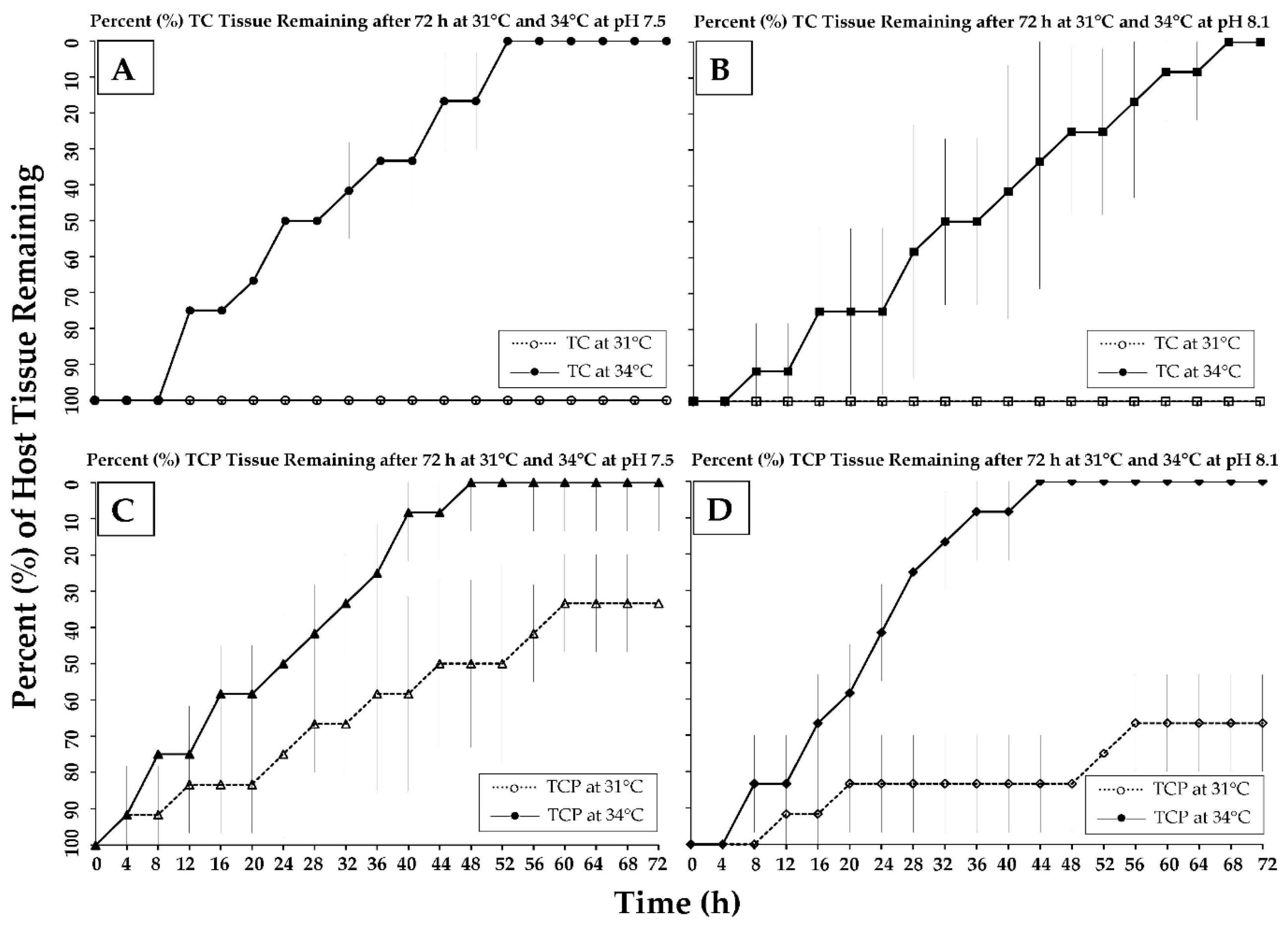
3.2. Tissue Loss Analysis
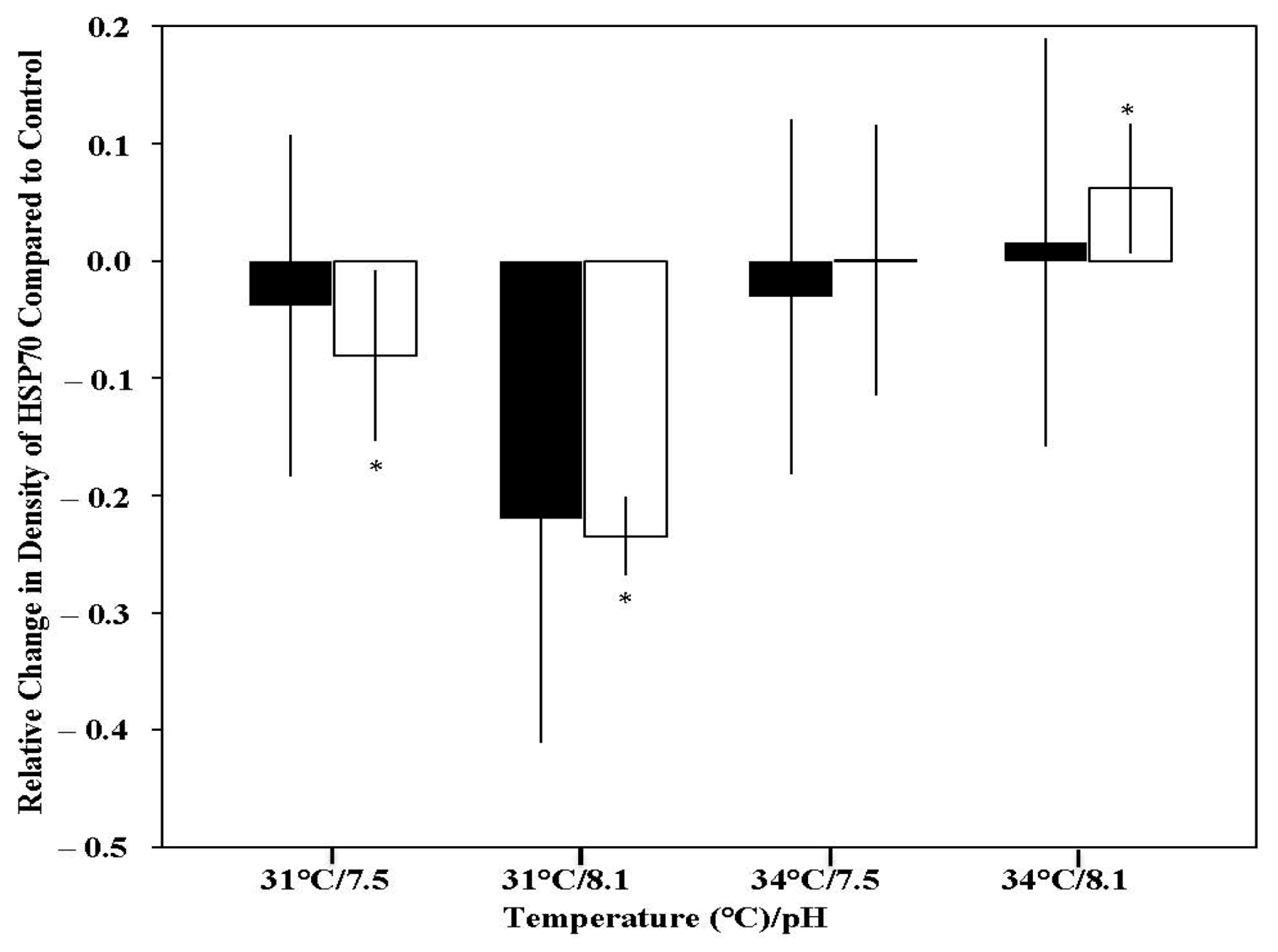
3.3. Western Blotting for HSP70
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strychar, K.B.; Sammarco, P.W. Temperate Marine and Brackish Ecosystems. In Climate Change and Non-Infectious Fish Disorders (CCNFD); Woo Patrick, T.K., Iwama, G.K., Eds.; CAB International: Wallingford, UK, 2020; pp. 1–24. ISBN 978-1786393982. [Google Scholar]
- Sammarco, P.W.; Strychar, K.B. Ecological and evolutionary considerations regarding corals in a rapidly changing environment. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 553–573. ISBN 978-3319313030. [Google Scholar] [CrossRef]
- European Environment Agency (EEA). Climate Change and Water-Warmer Oceans, Flooding and Droughts. 2018. Available online: https://www.eea.europa.eu/signals/signals-2018-content-list/articles/climate-change-and-water-2014 (accessed on 20 October 2021).
- Australian Academy of Science. More than Just Temperature—Climate Change and Ocean Acidification. 2021. Available online: https://www.science.org.au/curious/earth-environment/ocean-acidification. (accessed on 20 October 2021).
- Skutnik, J.E.; Otieno, S.; Kean-Khoo, S.; Strychar, K.B. Examining the Effect of Heat Stress on Montastraea cavernosa (Linnaeus 1767) from a Mesophotic Coral Ecosystem (MCE). J. Water 2020, 12, 1303. [Google Scholar] [CrossRef]
- Doney, S.C.; Ruckelshaus, M.; Duffy, J.E.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate change impacts on marine ecosystems. Annu. Rev. Mar. Sci. 2011, 4, 11–37. [Google Scholar] [CrossRef]
- Allemand, D.; Osborn, D. Ocean acidification impacts on coral reefs: From sciences to solutions. Reg. Stud. Mar. Sci. 2019, 28, 100558. [Google Scholar] [CrossRef]
- Doo, S.S.; Edmunds, P.J.; Carpenter, R.C. Ocean acidification effects on in situ coral reef metabolism. Sci. Rep. 2019, 9, 12067. [Google Scholar] [CrossRef]
- Sammarco, P.W.; Porter, S.A.; Cairns, S.D. A new coral species introduced into the Atlantic Ocean—Tubastraea micranthus (Ehrenberg 1834) (Cnidaria, Anthozoa, Scleractinia): An invasive threat? Aquat. Invasions 2010, 5, 131–140. [Google Scholar] [CrossRef]
- Sammarco, P.W.; Porter, S.A.; Sinclair, J.; Genazzio, M. Population expansion of a new invasive coral species, Tubastraea micranthus, in the northern Gulf of Mexico. Mar. Ecol. Prog. Ser. 2014, 495, 161–173. [Google Scholar] [CrossRef]
- Sammarco, P.W.; Brazeau, D.A.; McKoin, M.; Strychar, K.B. Tubastraea micranthus: Comments on the population genetics of a new invasive coral in the Western Atlantic and a possible secondary invasion. J. Exp. Mar. Biol. Ecol. 2017, 490, 56–63. [Google Scholar] [CrossRef]
- Capel, K.; Creed, J.C.; Kitahara, M.V. Invasive corals trigger seascape changes in the southwestern Atlantic. Bull. Mar. Sci. 2020, 96, 217–218. [Google Scholar] [CrossRef]
- Figueroa, D.F.; McClure, A.; Figueroa, N.J.; Hicks, D.W. Hiding in plain sight: Invasive coral Tubastraea tagusensis (Scleractinia:Hexacorallia) in the Gulf of Mexico. Coral Reefs 2019, 38, 395–403. [Google Scholar] [CrossRef]
- Vaughan, T.W.; Wells, J.W. (Eds.) Revision of the Suborders, Families, and Genera of the Scleractinia; Special Papers; Geological Society of America: Boulder, CO, USA, 1943; p. 44. [Google Scholar] [CrossRef]
- Lesson, R.P. Voyage Autour du Monde sur La Coquille, Pendant les Annees 1822, 1823, 1824, et 1825, Zoology, 2: Zoophytes; A. Bertrand: Paris, France, 1829; pp. 1–151, (In French). Available online: https://gallica.bnf.fr/ark:/12148/bpt6k850915v (accessed on 16 December 2021).
- Fenner, B.; Banks, K. Orange cup coral Tubastraea cocciena invades Florida and the Flower Garden Banks, Northwestern Gulf of Mexico. Coral Reefs 2004, 23, 505–507. [Google Scholar] [CrossRef]
- de Paula, A.F.; Pires, D.; Creed, J. Reproductive strategies of two invasive sun corals (Tubastraea spp.) in the southwestern Atlantic. J. Mar. Biol. Assoc. UK 2014, 94, 481–492. [Google Scholar] [CrossRef]
- Vermeij, M.J.A. A novel growth strategy allows Tubastraea coccinea to escape small-scale adverse conditions and start over again. Coral Reefs 2005, 24, 442. [Google Scholar] [CrossRef]
- Sentoku, A.; Ezaki, Y. Regularity in budding mode and resultant growth morphology of the azooxanthellate colonial scleractinian Tubastraea coccinea. Coral Reefs 2012, 31, 67–74. [Google Scholar] [CrossRef]
- Riul, P.; Targino, C.H.; Júnior, L.A.C.; Creed, J.C.; Horta, P.A.; Costa, G.C. Invasive potential of the coral Tubastraea coccinea in the southwest Atlantic. Mar. Ecol. Prog. Ser. 2013, 480, 73–81. [Google Scholar] [CrossRef]
- Creed, J.C.; Fenner, D.; Sammarco, P.W.; Cairns, S.; Capel, K.; Junqueira, A.O.R.; Cruz, I.; Miranda, R.J.; Carlos-Junior, L.; Mantelatto, M.C.; et al. The invasion of the azooxanthellate coral Tubastraea (Scleractinia: Dendrophylliidae) throughout the world: History, pathways and vectors. Biol. Invasions 2016, 19, 283–308. [Google Scholar] [CrossRef]
- Luz, B.L.P.; Domenico, M.D.; Migotto, A.E.; Kitahara, M.V. Life-history traits of Tubastraea coccinea: Reproduction, development, and larval competence. Ecol. Evol. 2020, 10, 6223–6238. [Google Scholar] [CrossRef] [PubMed]
- Engelen, A.H.; Aires, T.; Vermeij, M.J.A.; Herndl, G.J.; Serrão, E.A.; Frade, P.R. Host differentiation and compartmentalization of microbial communities in the azooxanthellate cupcorals Tubastrea coccinea and Rhizopsammia goesi in the Caribbean. Front. Mar. Sci. 2018, 5, 391. [Google Scholar] [CrossRef]
- Moreira, T.S.G.; Creed, J.C. Invasive, non-indigenous corals in a tropical rocky shore environment: No evidence for generalist predation. J. Exp. Mar. Biol. Ecol. 2012, 438, 7–13. [Google Scholar] [CrossRef]
- Sammarco, P.W.; (Louisiana Universities Marine Consortium, Chauvin, LA, USA). Personal Communication, 2021.
- Silva, A.G.; Lima, R.P.; Gomes, A.N.; Fleury, B.G.; Creed, J.C. Expansion of the invasive corals Tubastraea coccinea and Tubastraea tagusensis into the Tamoios Ecological Station Marine Protected Area, Brazil. Aquat. Invasion 2011, 6, S105–S110. [Google Scholar] [CrossRef]
- Rodríguez-Villalobos, J.C.; Ayala-Bocos, A.; Hernández, L. Predation by Epidendrium billeeanum on Tubastrea coccinea: Use of the denuded skeleton for laying eggs. Coral Reefs 2016, 35, 271. [Google Scholar] [CrossRef]
- Paz-García, D.A.; Aldana-Moreno, A.; Cabral-Tena, R.A.; Balart, E.F. High predation by the corallivore sea snail Jenneria pustulata in a high-latitude reef in the Gulf of California. Mar. Biodivers. Rec. 2012, 5, e94. [Google Scholar] [CrossRef]
- Castro, C.B.; Pires, D. Brazilian coral reefs: What we already know and what is still missing. Bull. Mar. Sci. 2001, 69, 357–371. [Google Scholar]
- Figueira de Paula, A.F.; Creed, J.C. Two species of the coral Tubastraea (Cnidaria, Scleractinia) in Brazil: A case study of accidental introduction. Bull. Mar. Sci. 2004, 74, 175–183. [Google Scholar]
- Sampaio, C.L.S.; Miranda, R.J.; Maia-Nogueira, R.; José de Anchieta, C.C. NunesNew occurrences of the nonindigenous orange cup corals Tubastraea coccinea and T. tagusensis (Scleractinia: Dendrophylliidae) in Southwestern Atlantic. Check List 2012, 8, 528–530. [Google Scholar] [CrossRef]
- Bianco, E.M.; de Oliveira, S.Q.; da Rosa Guimarães, T.; de Souza Pessoa, L.G.; Santos, M.E.A.; Dal-Pizzol, F.; Schenkel, E.P.; Reginatto, F.H. Tubastraea coccinea: A non-indigenous coral (Cnidaria, Scleractinia) collected at Arvoredo Island, South of Brazil with potential MRSA and VRE antimicrobial activity. Open J. Mar. Sci. 2016, 6, 334–340. [Google Scholar] [CrossRef]
- Fenner, D. Biogeography of three Caribbean corals (Scleractinia) and the invasion of Tubastraea coccinea into the Gulf of Mexico. Bull. Mar. Scil. 2001, 69, 1175–1189. [Google Scholar]
- Sammarco, P.W.; Porter, S.A.; Sinclair, J.; Genazzio, M. Depth distribution of a new invasive coral (Gulf of Mexico)—Tubastraea micranthus, comparisons with T. coccinea, and implications for control. Manag. Biol. Invasions 2013, 4, 291–303. [Google Scholar] [CrossRef]
- Crivellaro, M.S.; Silveira, T.C.L.; Custódio, F.Y.; Battaglin, L.C.; Dechoum, M.d.S.; Fonseca, A.C.; Bárbara, S.; Riul, P.; Targino, C.H.; Junior, L.A.C.; et al. Fighting on the edge: Reproductive effort and population structure of the invasive coral Tubastraea coccinea in its southern Atlantic limit of distribution following control activities. Biol. Invasions 2020, 23, 811–823. [Google Scholar] [CrossRef]
- Capel, K.C.C.; Toonen, R.J.; Rachid, C.T.C.C.; Creed, J.C.; Kitahara, M.V.; Forsman, Z.; Zilberberg, C. Clone wars: Asexual reproduction dominates in the invasive range of Tubastraea spp. (Anthozoa: Scleractinia) in the South-Atlantic Ocean. PeerJ 2017, e3873. [Google Scholar] [CrossRef]
- Wellington, G.M.; Trench, R.K. Persistence and coexistence of a nonsymbiotic coral in open reef environments. Proc. Natl. Acad. Sci. USA 1984, 82, 2432–2436. [Google Scholar] [CrossRef] [PubMed]
- Lages, B.G.; Fleury, B.G.; Menegola, C.; Creed, J.C. Change in tropical rocky shore communities due to an alien coral invasion. Mar. Ecol. Prog. Ser. 2011, 438, 85–96. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Hiemstra, A.-F.; Vermeij, M.J.A. The rise of a native sun coral species on southern Caribbean coral reefs. Ecosphere 2019, 10, e02942. [Google Scholar] [CrossRef]
- Sammarco, P.W.; Porter, S.A.; Genazzio, M.; Sinclair, J. Success in competition for space in two invasive coral species in the western Atlantic—Tubastraea micranthus and T. coccinea. PLoS ONE 2015, 10, e0144581. [Google Scholar] [CrossRef]
- Kolian, S.R.; Sammarco, P.W. Densities of reef-associated fish and corals on offshore platforms in the Gulf of Mexico. Bull. Mar. Sci. 2019, 95, 393–407. [Google Scholar] [CrossRef]
- Cairns, S. Stratigraphic distribution of Neogene azooxanthellate corals (Scleractinia, Stylasteridae). Bull. Am. Paleontol. 1999, 357, 109–118. [Google Scholar]
- Capel, K.C.C.; Creed, J.; Kitahara, M.V.; Chen, C.A.; Zilberberg, C. Multiple introductions and secondary dispersion of Tubastraea spp. in the Southwestern Atlantic. Sci. Rep. 2019, 9, 13978. [Google Scholar] [CrossRef] [PubMed]
- Zanotti, A.A.; Gregoracci, G.B.; Capel, K.C.C.; Kitahara, M.V. Microbiome of the Southwestern Atlantic invasive scleractinian coral, Tubastraea tagusensis. Anim. Microbiome 2020, 2, 29. [Google Scholar] [CrossRef]
- Luz, B.L.P.; Zilberberg, C.; Flores, A.A.V.; Migotto, A.E.; Kitahara, M.V. A polyp from nothing: The extreme regeneration capacity of the Atlantic invasive sun corals Tubastraea coccinea and T. tagusensis (Anthozoa, Scleractinia). J. Exp. Mar. Biol. Ecol. 2018, 503, 60–65. [Google Scholar] [CrossRef]
- de Mello Carpes, R.; de Assis Alves, M.; Creed, J.C.; da Silva, C.A.; Hamerski, L.; Garden, S.J.; Fleury, B.G.; Felzenszwalb, I. Mutagenic, genotoxic and cytotoxic studies of invasive corals Tubastraea coccinea and Tubastraea tagusensis. J. Appl. Toxicol. 2020, 40, 373–387. [Google Scholar] [CrossRef] [PubMed]
- de Mello Carpes, R.; Fernandes, D.C.; Coelho, M.G.P.; Creed, J.C.; Fleury, B.G.; Garden, S.J.; Felzenszwalb, I. Anti-inflammatory potential of invasive sun corals (Scleractinia: Tubastraea spp.) from Brazil: Alternative use for management? J. Pharm. Pharmacol. 2020, 72, 633–647. [Google Scholar] [CrossRef]
- Soares-Souza, G.B.; Amaral, D.; Batista, D.; Torres, A.Q.; Serra, A.C.S.; Uliano-Silva, M.; Leomil, L.; Reis, A.C.; Monteiro de Matos, E.; Calderon, E.; et al. The genomes of invasive coral Tubastraea spp. (Dendrophylliidae) as tool for the development of biotechnological solutions. bioRxiv 2020. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef]
- Strychar, K.B.; Sammarco, P.W. Effects of heat stress on phytopigments of zooxanthellae (Symbiodinium spp.) symbiotic with the corals Acropora hyacinthus, Porites solida, and Favites complanata. Int. J. Biol. 2012, 4, 3–19. [Google Scholar] [CrossRef][Green Version]
- Hayes, R.L.; King, C.M. Induction of 70-kD heat shock protein in scleractinian corals by elevated temperature: Significance for coral bleaching. Mol. Mar. Bio. Biotechnol. 1995, 4, 36–42. [Google Scholar] [PubMed]
- Agostini, S.; Harvey, B.P.; Wada, S.; Kon, K.; Milazzo, M.; Inaba, K.; Hall-Spencer, J.M. Ocean acidification drives community shifts towards simplified non-calcified habitats in a subtropical−temperate transition zone. Sci. Rep. 2018, 8, 11354. [Google Scholar] [CrossRef] [PubMed]
- Mollica, N.R.; Guo, W.; Cohen, A.L.; Huang, K.-F.; Foster, G.L.; Donald, H.K.; Solow, A.R. Ocean acidification affects coral growth by reducing skeletal density. Proc. Natl. Acad. Sci. USA 2018, 115, 1754–1759. [Google Scholar] [CrossRef] [PubMed]
- Kohler, K.E.; Gill, S.M. Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 23, 1259–1269. [Google Scholar] [CrossRef]
- Winters, G.; Holzman, R.; Blekhman, A.; Beer, S.; Loya, Y. Photographic assessment of coral chlorophyll contents: Implications for ecophysiological studies and coral monitoring. J. Exp. Mar. Biol. Ecol. 2009, 380, 25–35. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; Estrada-Saldívar, N.; Pérez-Cervantes, E.; Molina-Hernández, A.; González-Barrios, F.J. A rapid spread of the stony coral tissue loss disease outbreak in the Mexican Caribbean. PeerJ 2019, 7, e8069. [Google Scholar] [CrossRef]
- Meiling, S.; Muller, E.M.; Smith, T.B.; Brandt, M.E. 3D Photogrammetry reveals dynamics of stony coral tissue loss disease (SCTLD) lesion progression across a thermal stress event. Front. Mar. Sci. 2020, 7, 597643. [Google Scholar] [CrossRef]
- Louis, Y.D.; Bhagooli, R.; Seveso, D.; Maggioni, D.; Galli, P.; Vai, M.; Dyall, S.D. Local acclimatisation-driven differential gene and protein expression patterns of Hsp70 in Acropora muricata: Implications for coral tolerance to bleaching. Mol. Ecol. 2020, 29, 4382–4394. [Google Scholar] [CrossRef]
- Seveso, D.; Arrigoni, R.; Montano, S.; Maggioni, D.; Orlandi, I.; Berumen, M.L.; Galli, P.; Vai, M. Investigating the heat shock protein response involved in coral bleaching across scleractinian species in the central Red Sea. Coral Reefs 2020, 39, 85–98. [Google Scholar] [CrossRef]
- Tom, M.; Douek, J.; Yankelevich, I.; Bosch, T.C.; Rinkevich, B. Molecular characterization of the first heat shock protein 70 from a reef coral. Biochem. Biophys. Res. Commun. 1999, 262, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Yetsko, K.L. Estimating the Heritability of Thermal Tolerance in Acropora cervicornis and the Physiological Basis of Adaptation that Correlates to Survival at Elevated Temperatures. Master’s Thesis, University of North Florida, Jacksonville, FL, USA, 2018. [Google Scholar]
- Ghosh, S.; Gepstein, S.; Heikkila, J.J.; Dumbroff, B.G. Use of a scanning densitometer or an ELISA plate reader for measurement of nanogram amounts of protein in crude extracts from biological tissue. Anal. Biochem. 1988, 169, 227–233. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 16 December 2021).
- Marshall, A.T.; Clode, P. Calcification rate and the effect of temperature in a zooxanthellate and azooxanthellate scleractinian reef coral. Coral Reefs 2004, 23, 218–224. [Google Scholar] [CrossRef]
- US Navy. Atlantic Ocean Facts. Archived from the Original on 2 March 2001. Available online: http://oceanographer.navy.mil/atlantic.html (accessed on 15 November 2021).
- Anonymous. Temperature of the Water of the Gulf of Mexico. 2021. Available online: https://www.seatemperatu.re/seas-and-oceans/gulf-of-mexico/ (accessed on 15 November 2021).
- Kaur, A.; Kumar, P.; Weller, E.; Young, I.R. Positive relationship between seasonal Indo-Pacific Ocean wave power and SST. Sci. Rep. 2021, 11, 17419. [Google Scholar] [CrossRef]
- Hang, H.; Fox, M.H. Low pH suppresses synthesis of heat-shock proteins and thermotolerance. Radiat. Res. 1994, 140, 24–30. Available online: http://www.jstor.org/stable/3578564 (accessed on 16 December 2021). [CrossRef] [PubMed]
- Gullian, K.M.; Terrats, P.M. Effect of pH on temperature controlled degradation of reactive oxygen species, heat shock protein expression, and mucosal immunity in the sea cucumber Isostichopus badionotus. PLoS ONE 2017, 12, e0175812. [Google Scholar] [CrossRef]
- Cornwall, C.E.; Comeau, S.; Kornder, N.A.; Perry, C.T.; van Hooidonk, R.; DeCarlo, T.M.; Pratchett, M.S.; Anderson, K.D.; Browne, N.; Carpenter, R.; et al. Global declines in coral reef calcium carbonate production under ocean acidification and warming. Proc. Natl. Acad. Sci. USA 2021, 118, e2015265118. [Google Scholar] [CrossRef] [PubMed]
- Feely, R.; Doney, S.C.; Cooley, S.R. Ocean acidification: Present conditions and future changes in a high-CO2 world. Oceanography 2009, 22, 36–47. [Google Scholar] [CrossRef]
- Garcia-Soto, C.; Cheng, L.; Caesar, L.; Schmidtko, S.; Jewett, E.B.; Cheripka, A.; Rigor, I.; Caballero, A.; Chiba, S.; Báez, J.C.; et al. An overview of ocean climate change indicators: Sea surface temperature, ocean heat content, ocean pH, dissolved oxygen concentration, arctic sea ice extent, thickness and volume, sea level and strength of the AMOC (Atlantic Meridional Overturning Circulation). Front. Mar. Sci. 2021, 8, 642372. [Google Scholar] [CrossRef]
- Winner, C. Small drop in pH Means Big Change in Acidity. Oceans: The Socioeconomic Costs of Ocean Acidification. 2010. Available online: https://www.whoi.edu/oceanus/feature/small-drop-in-ph-means-big-change-in-acidity (accessed on 20 October 2021).
- Hendrick, J.P.; Hartl, F.U. Molecular chaperone functions of heat-shock proteins. Annu. Rev. Biochem. 1993, 62, 349–384. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.E.; Downs, C.A.; Dunne, R.P.; Gibb, S.W. Exploring the basis of thermotolerance in the reef coral Goniastrea aspera. Mar. Ecol. Prog. Ser. 2002, 242, 119–129. [Google Scholar] [CrossRef]
- Bradshaw, A.D. Evolutionary significance of phenotypic plasticity in plants. Adv. Genet. 1965, 13, 115–155. [Google Scholar] [CrossRef]
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strychar, K.B.; Hauff-Salas, B.; Haslun, J.A.; DeBoer, J.; Cryer, K.; Keith, S.; Wooten, S. Stress Resistance and Adaptation of the Aquatic Invasive Species Tubastraea Coccinea (Lesson, 1829) to Climate Change and Ocean Acidification. Water 2021, 13, 3645. https://doi.org/10.3390/w13243645
Strychar KB, Hauff-Salas B, Haslun JA, DeBoer J, Cryer K, Keith S, Wooten S. Stress Resistance and Adaptation of the Aquatic Invasive Species Tubastraea Coccinea (Lesson, 1829) to Climate Change and Ocean Acidification. Water. 2021; 13(24):3645. https://doi.org/10.3390/w13243645
Chicago/Turabian StyleStrychar, Kevin B., Briana Hauff-Salas, Joshua A. Haslun, Jessica DeBoer, Katherine Cryer, Scott Keith, and Sam Wooten. 2021. "Stress Resistance and Adaptation of the Aquatic Invasive Species Tubastraea Coccinea (Lesson, 1829) to Climate Change and Ocean Acidification" Water 13, no. 24: 3645. https://doi.org/10.3390/w13243645
APA StyleStrychar, K. B., Hauff-Salas, B., Haslun, J. A., DeBoer, J., Cryer, K., Keith, S., & Wooten, S. (2021). Stress Resistance and Adaptation of the Aquatic Invasive Species Tubastraea Coccinea (Lesson, 1829) to Climate Change and Ocean Acidification. Water, 13(24), 3645. https://doi.org/10.3390/w13243645