
The Relative Importance of Human Disturbance, Environmental and Spatial Factors on the Community Composition of Wetland Birds




Abstract
1. Introduction
2. Materials and Methods
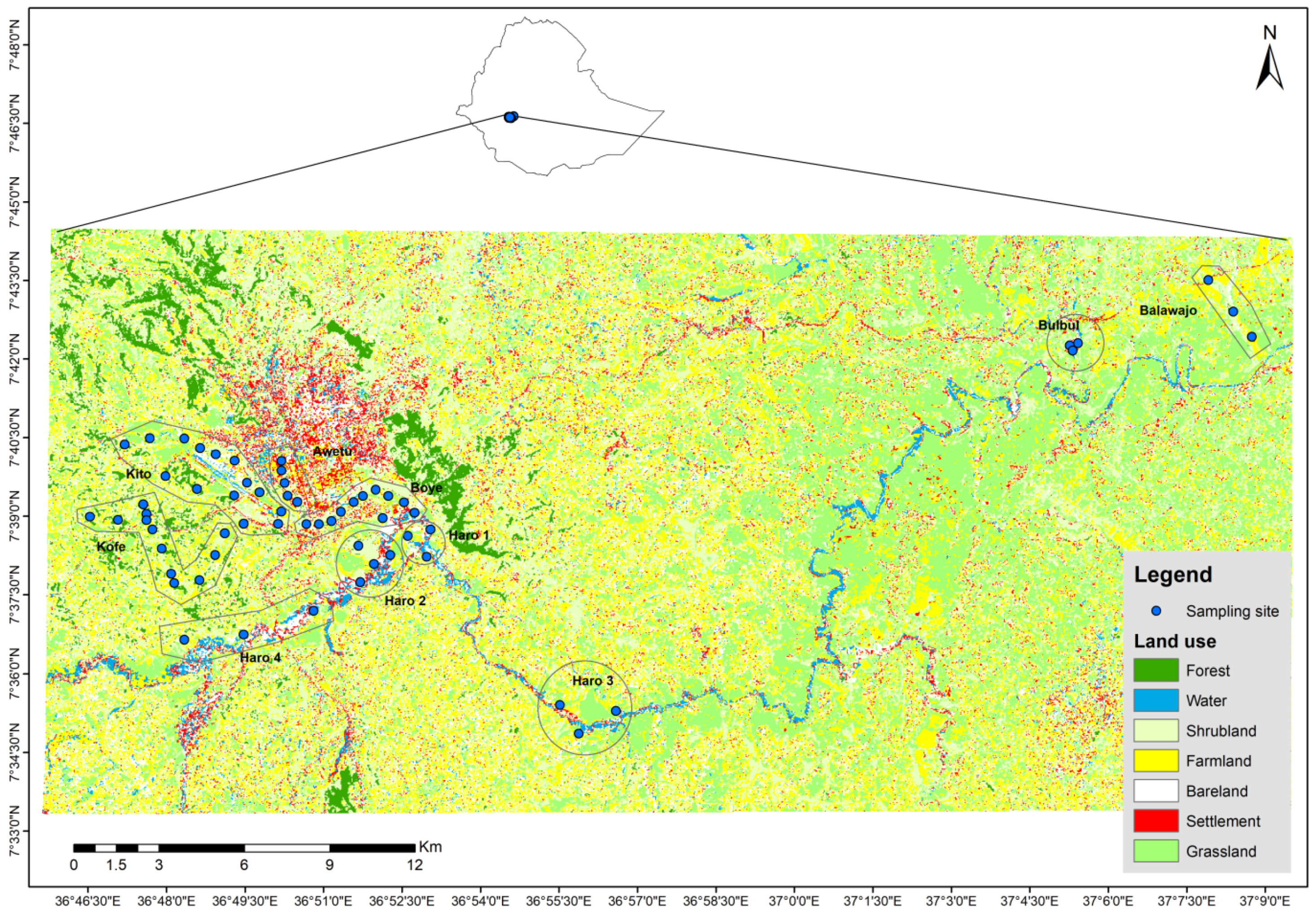
2.1. Study Area
2.2. Bird Surveys
2.3. Environmental Variables
2.4. Human Disturbance
2.5. Spatial Variables
2.6. Data Analyses
3. Results
3.1. Bird Surveys
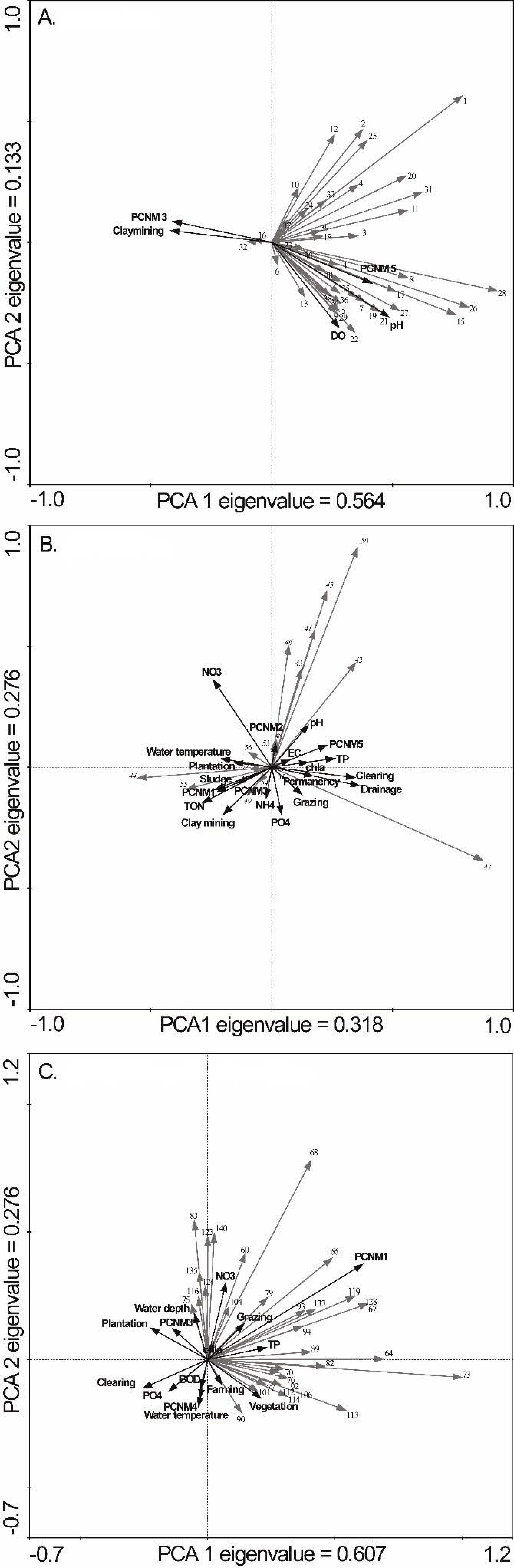
3.2. The Effect of Environment, Human Disturbance, Space on Bird Community Composition
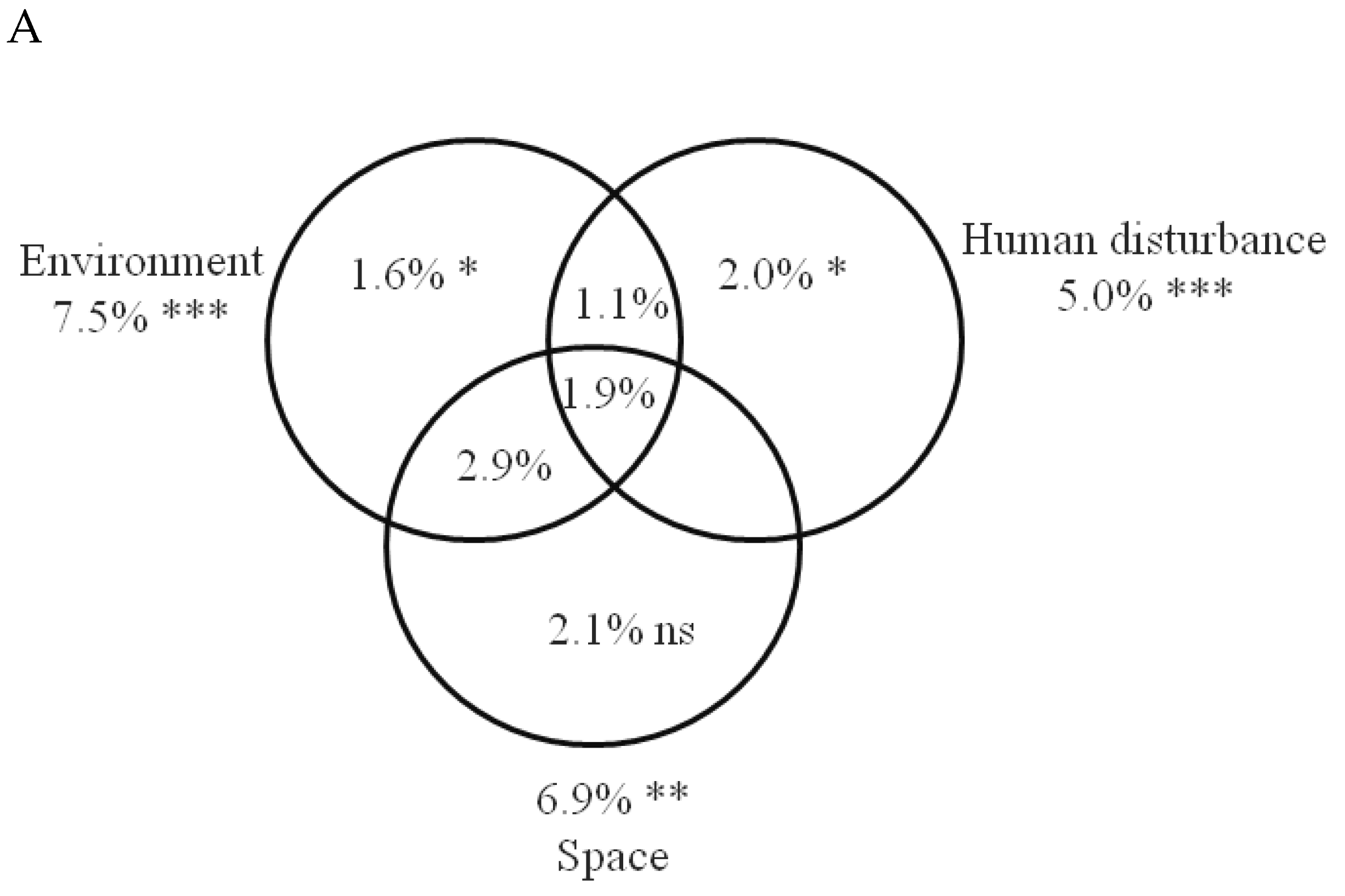
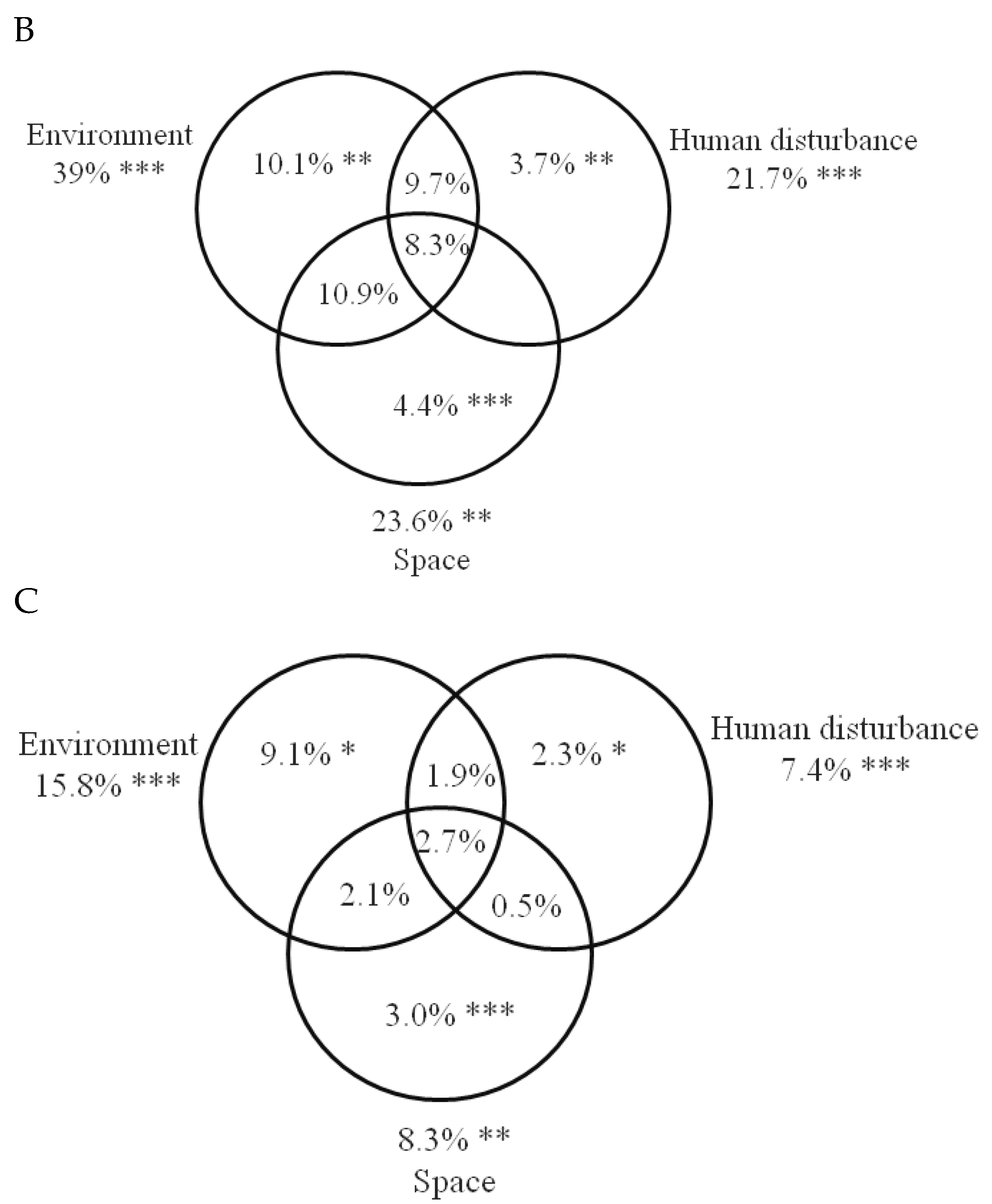
3.3. The Relative Importance of Local Environment, Human Disturbance, Space and Wetland Identity on Bird Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frohn, R.C.; Reif, M.; Lane, C.; Autrey, B. Satellite Remote Sensing of Isolated Wetlands Using object-Oriented Classification of Landsat-7 Data. Wetlands 2009, 29, 931–941. [Google Scholar] [CrossRef]
- Tiner, R.W. The primary indicators method: A practical approach to wetland recognition and delineation in the United States. Wetlands 1993, 13, 50–64. [Google Scholar] [CrossRef]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Lamsal, P.; Pant, K.P.; Kumar, L.; Atreya, K. Sustainable livelihoods through conservation of wetland resources: A case of economic benefits from Ghodaghodi Lake, western Nepal. Ecol. Soc. 2015, 20, 1. [Google Scholar] [CrossRef]
- Moges, A.; Beyene, A.; Kelbessa, E.; Mereta, S.T.; Ambelu, A. Development of a multimetric plant-based index of biotic integrity for assessing the ecological state of forested, urban and agricultural natural wetlands of Jimma Highlands, Ethiopia. Ecol. Indic. 2016, 71, 208–217. [Google Scholar] [CrossRef]
- Acreman, M.; Harding, R.; Lloyd, C.; Mcneil, D. Evaporation characteristics of wetlands: Experience from a, wet grassland and a reedbed using eddy correlation measurements. Hydrol. Earth. Syst. Sci. 2003, 7, 11–21. [Google Scholar] [CrossRef]
- Gayen, J.G.; Khosh, P.K.; Datta, D. Avifauna habitability of wetlands in an intensive cultural landscape: Insights from the Ichhamati floodplains, India. Environ. Monit. Assess. 2020, 192, 329. [Google Scholar] [CrossRef] [PubMed]
- Rajpar, M.N.; Zakaria, M. Bird species abundance and their correlationship with microclimate and habitat variables at Natural Wetland Reserve, Peninsular Malaysia. Int. J. Zool. 2011, 2011, 1–17. [Google Scholar] [CrossRef]
- Aynalem, S.; Bekele, A. Species composition, relative abundance and distribution of bird fauna of riverine and wetland habitats of Infranz and Yiganda at southern tip of Lake Tana, Ethiopia. Trop. Ecol. 2008, 49, 199. [Google Scholar]
- Guareschi, S.; Abellan, P.; Laini, A.; Green, A.J.; Sanchez-Zapata, J.A.; Velasco, J.; Millan, A. Cross-taxon congruence in wetlands: Assessing the value of waterbirds as surrogates of macroinvertebrate biodiversity in Mediterranean Ramsar sites. Ecol. Indic. 2015, 49, 204–215. [Google Scholar] [CrossRef]
- Matthews, T.J.; Cottee-Jones, H.E.; Whittaker, R.J. Habitat fragmentation and the species–area relationship: A focus on total species richness obscures the impact of habitat loss on habitat specialists. Divers. Distrib. 2014, 20, 1136–1146. [Google Scholar] [CrossRef]
- Sreekar, R.; Huang, G.; Zhao, J.B.; Pasion, B.O.; Yasuda, M.; Zhang, K.; Peabotuwage, I.; Wang, X.; Quan, R.C.; Slik, W.J.F.; et al. The use of species–area relationships to partition the effects of hunting and deforestation on bird extirpations in a fragmented landscape. Divers. Distrib. 2015, 21, 441–450. [Google Scholar] [CrossRef]
- Mereta, S.T.; Boets, P.; Bayih, A.A.; Malu, A.; Ephrem, Z.; Sisay, A.; Endale, H.; Yitbarek, M.; Jemal, A.; De Meester, L.; et al. Analysis of environmental factors determining the abundance and diversity of macroinvertebrate taxa in natural wetlands of southwest Ethiopia. Ecol. Inform. 2012, 7, 52–61. [Google Scholar] [CrossRef]
- Junk, W.J.; An, S.; Finlayson, C.; Gopal, B.; Květ, J.; Mitchell, S.A.; Mitsch, W.J.; Robarts, R.D. Current state of knowledge regarding the world’s wetlands and their future under global climate change: A synthesis. Aquat. Sci. 2013, 75, 151–167. [Google Scholar] [CrossRef]
- Tozer, D.C.; Nol, E.; Abraham, K.F. Effects of local and landscape-scale habitat variables on abundance and reproductive success of wetland birds. Wetl. Ecol. Manag. 2010, 18, 679–693. [Google Scholar] [CrossRef]
- Quesnelle, P.E.; Fahrig, L.; Lindsay, K.E. Effects of habitat loss, habitat configuration and matrix composition on declining wetland species. Biol. Conserv. 2013, 160, 200–208. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Virkkala, R.; Rainio, K. Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural–forest mosaic. J. Appl. Ecol. 2004, 41, 824–835. [Google Scholar] [CrossRef]
- Guadagnin, D.L.; Maltchik, L.; Fonseca, C.R. Species–area relationship of Neotropical waterbird assemblages in remnant wetlands: Looking at the mechanisms. Divers. Distrib. 2009, 15, 319–327. [Google Scholar] [CrossRef]
- Guadagnin, D.L.; Peter, A.S.; Carvalho, L.F.; Perello, L.F.C.; Maltchick, L. Spatial and temporal patterns of waterbird assemblages in fragmented wetlands of southern Brazil. Waterbirds 2005, 28, 261–404. [Google Scholar] [CrossRef]
- Gianuca, A.T.; Bastazini, V.A.G.; Dias, R.A.; Hernández, M.I.M. Independent and shared effects of environmental features and space driving avian community beta diversity across a coastal gradient in southern Brazil. Austral. J. Ecol. 2013, 38, 864–873. [Google Scholar] [CrossRef]
- Rajaniemi, T.K.; Goldberg, D.E.; Turkington, R.; Dyer, A.R. Quantitative partitioning of regional and local processes shaping regional diversity patterns. Ecol. Lett. 2006, 9, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.P.; García, C.E.; Estay, S.A.; Barbosa, O. Bird richness and abundance in response to urban form in a Latin American city: Valdivia, Chile as a case study. PLoS ONE 2015, 10, e0138120. [Google Scholar] [CrossRef]
- Wilson, J.W.; Van Rensburg, B.J.; Ferguson, J.W.H.; Keith, M. The relative importance of environment, human activity and space in explaining species richness of South African bird orders. J. Biogeogr. 2008, 35, 342–352. [Google Scholar] [CrossRef]
- Hájek, M.; Roleček, J.; Cottenie, K.; Kintrová, K.; Horsák, M.; Poulíčková, A.; Hájková, P.; Fránková, M.; Dítě, D. Environmental and spatial controls of biotic assemblages in a discrete semi-terrestrial habitat: Comparison of organisms with different dispersal abilities sampled in the same plots. J. Biogeogr. 2011, 38, 1683–1693. [Google Scholar] [CrossRef]
- Birdlife International. Important Bird Areas and Potential Ramsar Sites in Africa; BirdLife International: Cambridge, UK, 2002. [Google Scholar]
- Kassa, Y.A.; Teshome, T.T. The Impact of wetland degradation and conversion on socioeconomic values: The case of Amhara national regional state Tekuma Wetland, Lake Tana Sub-Basin, Ethiopia. Int. J. Agric. Syst. 2015, 3, 1–14. [Google Scholar]
- Mereta, S.T.; Boets, P.; De Meester, L.; Goethals, P.L.M. Development of a multimetric index based on benthic macroinvertebrates for the assessment of natural wetlands in southwest Ethiopia. Ecol. Indic. 2013, 29, 510–521. [Google Scholar] [CrossRef]
- Hennings, L.A.; Edge, W.D. Riparian bird community structure in Portland, Oregon: Habitat, urbanization, and spatial scale patterns. Condor 2003, 105, 288–302. [Google Scholar] [CrossRef]
- Bonthoux, S.; Balent, G. Point count duration: Five minutes are usually sufficient to model the distribution of bird species and to study the structure of communities for a French landscape. J. Ornithol. 2012, 153, 491–504. [Google Scholar] [CrossRef]
- Sibley, C.G.; Monroe, B.L. Distribution and Taxonomy of Birds of the World; Yale University Press: New Haven, CT, USA, 1990. [Google Scholar]
- Almaw, R.A. Checklist of the Birds of the Abijata-Shalla Lakes National Park (Central Rift-Valley, Ethiopia); Ethiopian Wildlife Conservation Authority: Addis Ababa, Ethiopia, 2012. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 19th ed.; APHA American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Parsons, M.; Thoms, M.; Norris, R. AUSRIVAS (Australian River Assessment System) Physical Assessment Protocol; Cooperative Research Centre for Freshwater Ecology, University of Canberra: Bruce, Australia, 2001; Available online: https://ausrivas.ewater.org.au/protocol/Download/protocol-1.pdf (accessed on 20 June 2021).
- Borcard, D.; Legendre, P. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol. Modell. 2002, 153, 51–68. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; Gallagher, E. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.; Kindt, R.; Legendre, P.; O’hara, R.; Simpson, G.; Solymos, P.; Stevens, M.; Wagner, H. vegan Community Ecology Package: Ordination Methods, Diversity Analysis and Other Functions for Community and Vegetation Ecologists. R Package Version 2.5-1. 2018. Available online: https://cran.r-project.org/ (accessed on 20 June 2021).
- Lehman, P.W.; Sommer, T.; Rivard, L. The influence of floodplain habitat on the quantity and quality of riverine phytoplankton carbon produced during the flood season in San Francisco Estuary. Aquat. Ecol. 2008, 42, 363–378. [Google Scholar] [CrossRef]
- Paszkowski, C.A.; Tonn, W.M. Community concordance between the fish and aquatic birds of lakes in northern Alberta, Canada: The relative importance of environmental and biotic factors. Freshw. Biol. 2000, 43, 421–437. [Google Scholar] [CrossRef]
- Dures, S.G.; Cumming, G.S. The confounding influence of homogenising invasive species in a globally endangered and largely urban biome: Does habitat quality dominate avian biodiversity? Biol. Conserv. 2010, 143, 768–777. [Google Scholar] [CrossRef]
- Subasinghe, K.; Sumanapala, A.P.; Weerawardhena, S.R. The impact of forest conversion on bird communities in the northern flank of the Knuckles Mountain Forest Range, Sri Lanka. J. Asia-Pac. Biodivers. 2014, 7, 367–373. [Google Scholar] [CrossRef][Green Version]
- Robinson, S.K.; Thompson, F.R.; Donovan, T.M.; Whitehead, D.R.; Faaborg, J. Regional forest fragmentation and the nesting success of migratory birds. Science 1995, 267, 1987–1990. [Google Scholar] [CrossRef]
- Boulinier, T.; Nichols, J.D.; Hines, J.E.; Sauer, J.R.; Flather, C.H.; Pollock, K.H. Forest fragmentation and bird community dynamics: Inference at regional scales. Ecology 2001, 82, 1159–1169. [Google Scholar] [CrossRef]
- Sekercioglu, C.H. Effects of forestry practices on vegetation structure and bird community of Kibale National Park, Uganda. Biol. Conserv. 2002, 107, 229–240. [Google Scholar] [CrossRef]
- Raman, T.S. Effect of slash and burn shifting cultivation on rainforest birds in Mizoram, northeast India. Conserv. Biol. 2001, 15, 685–698. [Google Scholar] [CrossRef]
- Brouat, C.; Chevallier, H.; Meusnier, S.; Noblecourt, T.; Rasplus, J.Y. Specialization and habitat: Spatial and environmental effects on abundance and genetic diversity of forest generalist and specialist Carabus species. Mol. Ecol. 2004, 13, 1815–1826. [Google Scholar] [CrossRef] [PubMed]
- Devictor, V.; Julliard, R.; Jiguet, F. Distribution of specialist and generalist species along spatial gradients of habitat disturbance and fragmentation. Oikos 2008, 117, 507–514. [Google Scholar] [CrossRef]
- De Troyer, N.; Mereta, S.T.; Goethals, P.L.; Boets, P. Water quality assessment of streams and wetlands in a fast growing east African City. Water 2016, 8, 123. [Google Scholar] [CrossRef]
- Soka, G.E.; Munishi, P.K.; Thomas, M.B. Species diversity and abundance of Avifauna in and around Hombolo Wetland in Central Tanzania. Int. J. Biodivers. Conserv. 2013, 5, 782–790. [Google Scholar] [CrossRef]
- Kour, D.N.; Sahi, D. Studies on the community ecology of cattle egrets Bubulcus ibis coromandus (Boddaert) in Jammu (Jammu and Kashmir), India. Int. J. Biodivers. Conser. 2012, 4, 439–445. [Google Scholar]
- Kioko, J.; Boyd, E.; Schaeffer, E.; Tareen, S.; Kiffner, C. Cattle egret Bubulcus ibis interactions with large mammals in the Tarangire-Manyara ecosystem, Northern Tanzania. J. East Afr. Ornithol. 2016, 36, 15–20. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| df | Variance | F | R2adj | P | Selected Variables * | ||
|---|---|---|---|---|---|---|---|
| specialists (n = 40) | environment | 16 | 0.199 | 1.36 | 0.085 | 0.003 | pH, oxygen concentration |
| land-use | 9 | 0.136 | 1.645 | 0.086 | 0.001 | clay mining | |
| space | 6 | 0.105 | 1.897 | 0.080 | 0.001 | PCNM3, PCNM5 | |
| wetland ID | 10 | 0.153 | 1.697 | 0.101 | 0.001 | ||
| generalist (n = 17) | environment | 16 | 0.249 | 3.557 | 0.398 | 0.001 | pH, sludge, NO3, TON, chla, temperature, PO4, TP, NH4, EC, permanency |
| land-use | 9 | 0.153 | 3.056 | 0.223 | 0.001 | plantation, clearing, grazing, clay mining, drainage | |
| space | 6 | 0.141 | 4.256 | 0.234 | 0.001 | PCNM 1, PCNM2, PCNM3, PCNM5 | |
| wetland ID | 10 | 0.256 | 6.994 | 0.492 | 0.001 | ||
| associated (n = 83) | environment | 16 | 0.194 | 1.905 | 0.190 | 0.001 | TP, vegetation, NO3, temperature, PO4, BOD, chla, water depth |
| land-use | 9 | 0.104 | 1.608 | 0.081 | 0.001 | plantation, farming, grazing, clearing | |
| space | 6 | 0.089 | 2.076 | 0.094 | 0.001 | PCNM1, PCNM3, PCNM4 | |
| wetland ID | 10 | 0.172 | 2.843 | 0.230 | 0.001 |
| Variables | df | Variance | F | R2adj | p | Selected Variables * |
|---|---|---|---|---|---|---|
| environment | 19 | 0.264 | 2.240 | 0.275 | 0.001 | water depth, sludge, temperature, TON, NO3, TP, morphology, permanency, chla, vegetation cover |
| land use | 9 | 0.146 | 2.236 | 0.152 | 0.001 | drainage, grazing, clearing, plantation, farming, clay mining, waste dumping |
| space | 6 | 0.124 | 2.837 | 0.151 | 0.001 | PCNM1, PCNM3, PCNM4, PCNM5 |
| wetland ID | 10 | 0.220 | 3.669 | 0.301 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mereta, S.T.; Lemmens, P.; De Meester, L.; Goethals, P.L.M.; Boets, P. The Relative Importance of Human Disturbance, Environmental and Spatial Factors on the Community Composition of Wetland Birds. Water 2021, 13, 3448. https://doi.org/10.3390/w13233448
Mereta ST, Lemmens P, De Meester L, Goethals PLM, Boets P. The Relative Importance of Human Disturbance, Environmental and Spatial Factors on the Community Composition of Wetland Birds. Water. 2021; 13(23):3448. https://doi.org/10.3390/w13233448
Chicago/Turabian StyleMereta, Seid Tiku, Pieter Lemmens, Luc De Meester, Peter L. M. Goethals, and Pieter Boets. 2021. "The Relative Importance of Human Disturbance, Environmental and Spatial Factors on the Community Composition of Wetland Birds" Water 13, no. 23: 3448. https://doi.org/10.3390/w13233448
APA StyleMereta, S. T., Lemmens, P., De Meester, L., Goethals, P. L. M., & Boets, P. (2021). The Relative Importance of Human Disturbance, Environmental and Spatial Factors on the Community Composition of Wetland Birds. Water, 13(23), 3448. https://doi.org/10.3390/w13233448