Comprehensive Assessment Indicator of Ecosystem Resilience in Central Asia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
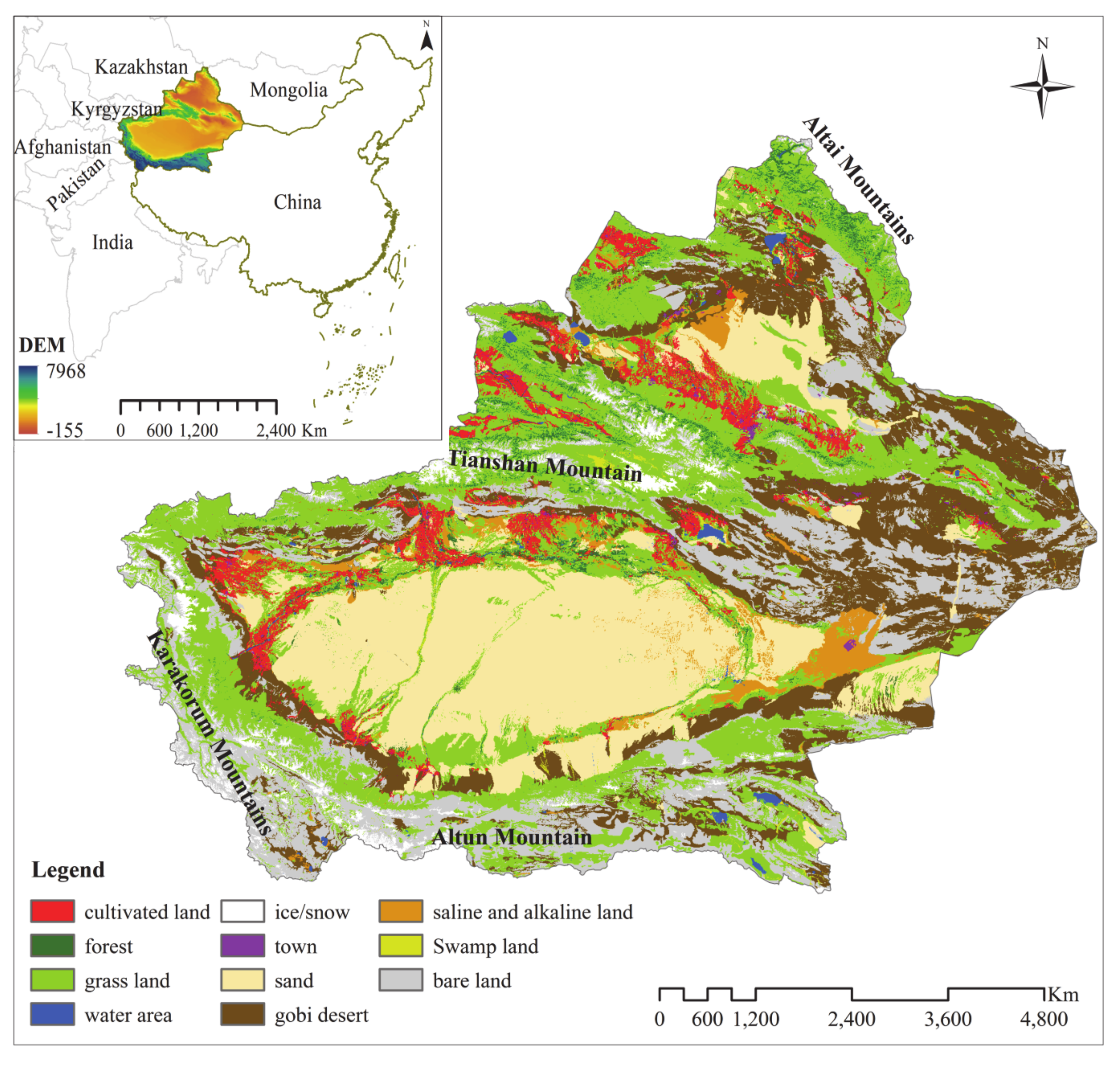
2.1. Study Area
2.2. Data
2.3. Methods
2.3.1. Latitude of Resilience
2.3.2. Resistance
2.3.3. Recovery Time Rate
2.3.4. Comprehensive Resilience Index
2.3.5. Grading of Resilience Indicators
3. Results
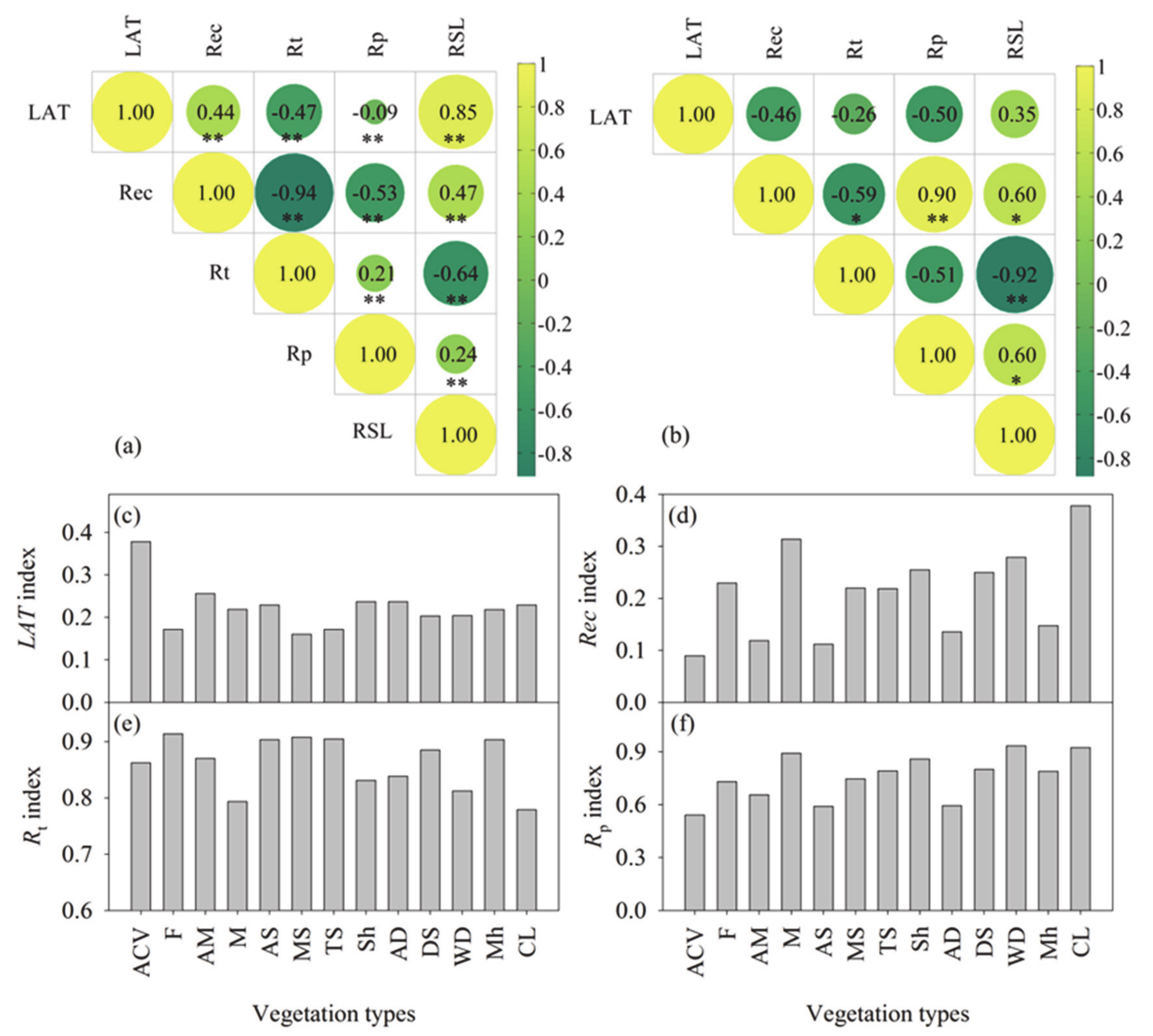
3.1. Multidimensional Characters of Resilience
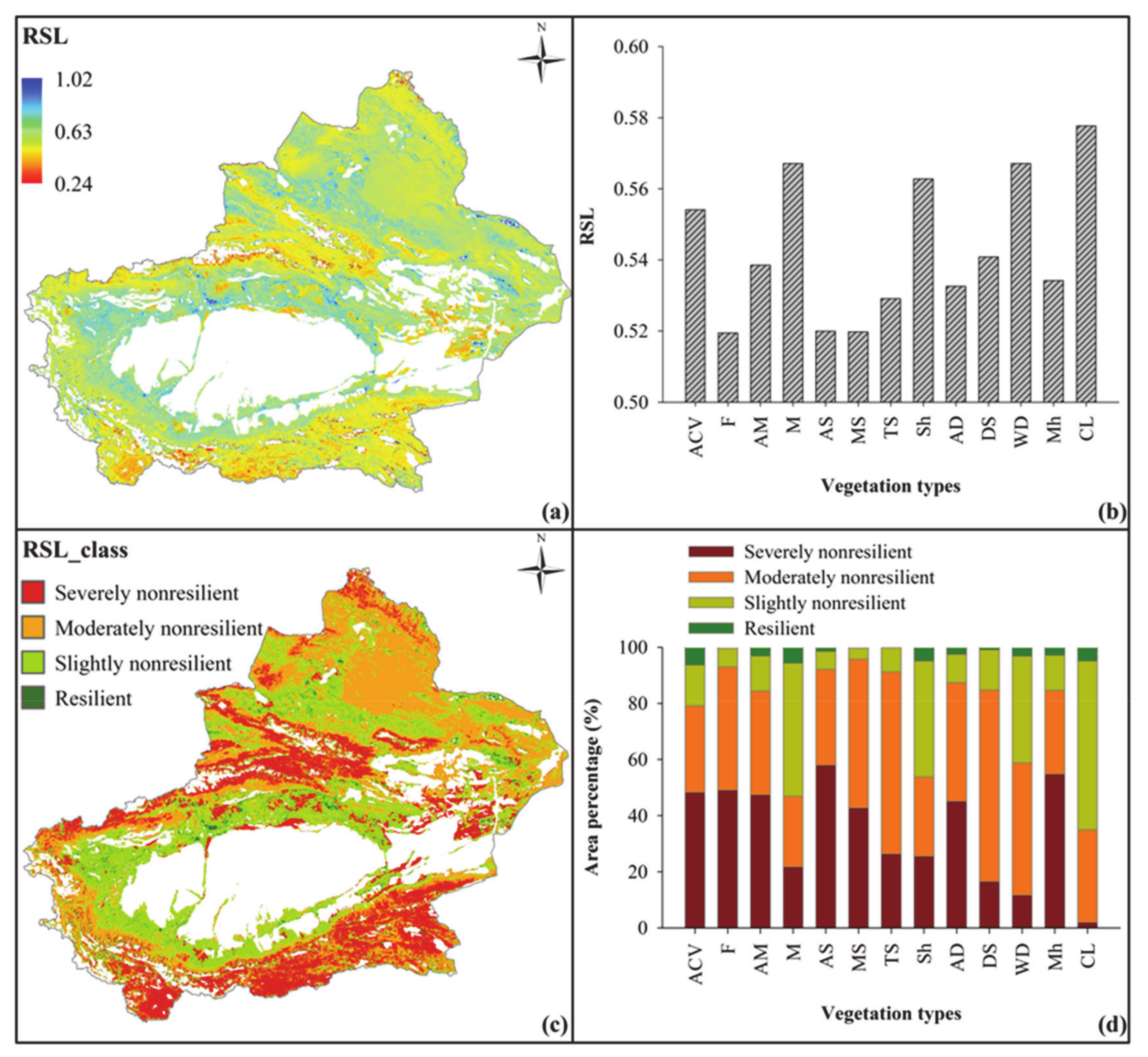
3.2. Comprehensive Resilience
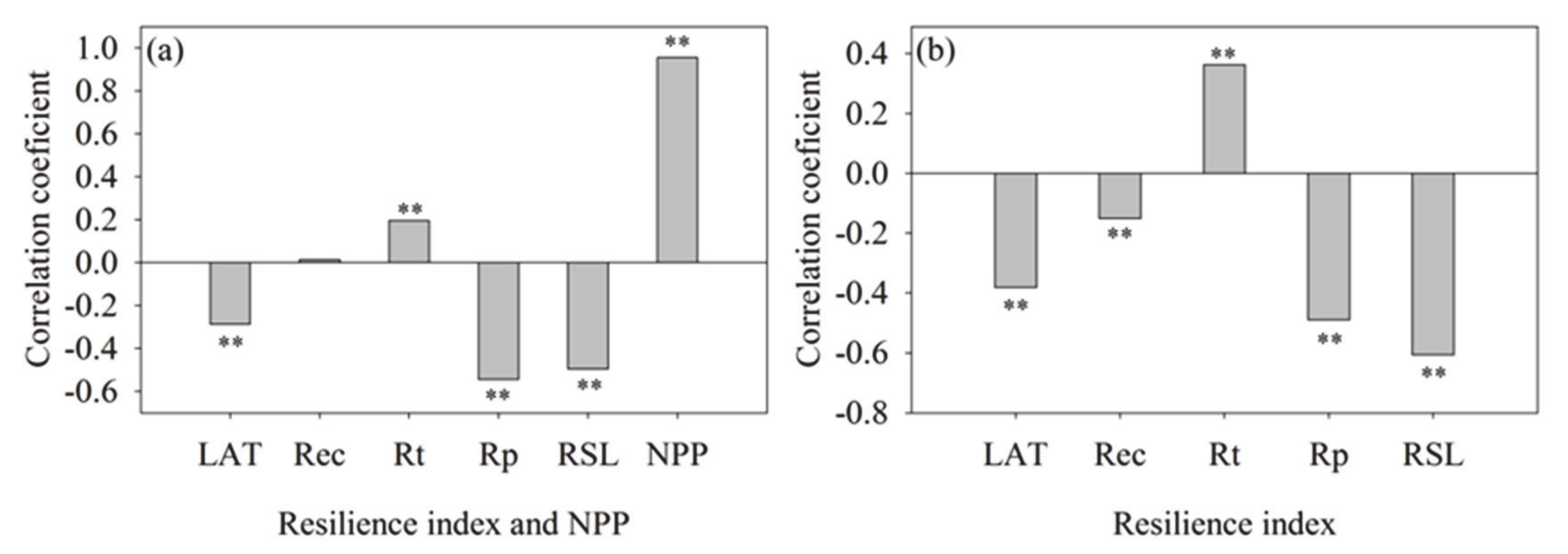
3.3. Coverage, NPP, and Resilience
4. Discussion
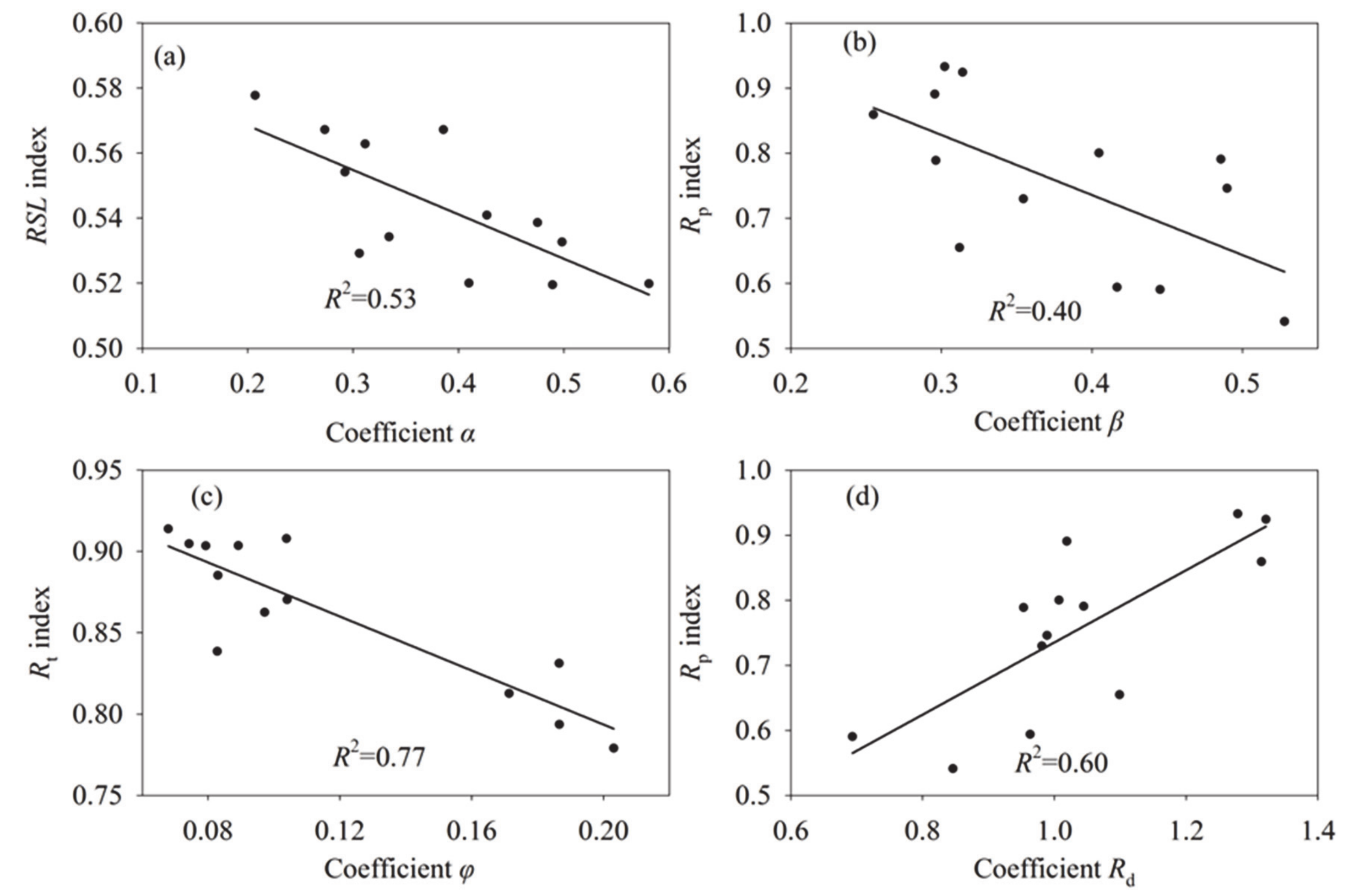
4.1. The Improvement and Reliability of the Method
4.2. Resistance, Resilience, and the Stability
4.3. Ecosystem Resilience and Ecosystem Management
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Resilience Index Based on the WUE
Appendix B. Resilience Index Based on the NDVI Anomaly
References
- Craine, J.M.; Ocheltree, T.W.; Nippert, J.B.; Towne, E.G.; Skibbe, A.M.; Kembel, S.W.; Fargione, J.E. Global diversity of drought tolerance and grassland climate-change resilience. Nat. Clim. Chang. 2012, 3, 63–67. [Google Scholar] [CrossRef]
- Marx, A.; Erhard, M.; Thober, S.; Kumar, R.; Shafer, D.; Samaniego, L.; Zink, M. Climate Change as Driver for Ecosystem Services Risk and Opportunities, in Atlas of Ecosystem Services: Drivers, Risks, and Societal Responses; Springer: Berlin/Heidelberg, Germany, 2019; pp. 173–178. [Google Scholar]
- Schirpke, U.; Kohler, M.; Leitinger, G.; Fontana, V.; Tasser, E.; Tappeiner, U. Future impacts of changing land-use and climate on ecosystem services of mountain grassland and their resil-ience. Ecosyst. Serv. 2017, 26, 79–94. [Google Scholar] [CrossRef]
- Holl, K.; Aide, T. When and where to actively restore ecosystems? For. Ecol. Manag. 2011, 261, 1558–1563. [Google Scholar] [CrossRef]
- Moritz, C.; Agudo, R. The Future of Species under Climate Change: Resilience or Decline? Science 2013, 341, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Valdecantos, A.; Baeza, M.J.; Vallejo, V.R. Vegetation Management for Promoting Ecosystem Resilience in Fire-Prone Med-iterranean Shrublands. Restor. Ecol. 2008, 17, 414–421. [Google Scholar] [CrossRef]
- Johnson, R.M.; Edwards, E.; Gardner, J.S.; Diduck, A.P. Community vulnerability and resilience in disaster risk reduction: An example from Phojal Nalla, Himachal Pradesh, India. Reg. Environ. Chang. 2018, 18, 2073–2087. [Google Scholar] [CrossRef]
- Peng, M.; Wen, Z.; Xie, L.; Cheng, J.; Jia, Z.; Shi, D.; Zeng, H.; Zhao, B.; Liang, Z.; Li, T.; et al. 3D Printing of Ultralight Biomimetic Hierarchical Graphene Materials with Exceptional Stiffness and Resilience. Adv. Mater. 2019, 31, e1902930. [Google Scholar] [CrossRef]
- Stork, N.; Coddington, J.A.; Colwell, R.K.; Chazdon, R.L.; Dick, C.W.; Peres, C.A.; Sloan, S.; Willis, K. Vulnerability and Resilience of Tropical Forest Species to Land-Use Change. Conserv. Biol. 2009, 23, 1438–1447. [Google Scholar] [CrossRef]
- Connell, S.D.; Ghedini, G. Resisting regime-shifts: The stabilising effect of compensatory processes. Trends Ecol. Evol. 2015, 30, 513–515. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and Stability of Ecological Systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Hodgson, D.; McDonald, J.L.; Hosken, D.J. What do you mean, ‘resilient’? Trends Ecol. Evol. 2015, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Standish, R.J.; Hobbs, R.J.; Mayfield, M.M.; Bestelmeyer, B.T.; Suding, K.N.; Battaglia, L.L.; Eviner, V.T.; Hawkes, C.V.; Temperton, V.M.; Cramer, V.A.; et al. Resilience in ecology: Abstraction, distraction, or where the action is? Biol. Conserv. 2014, 177, 43–51. [Google Scholar] [CrossRef]
- Yeung, A.C.; Richardson, J.S. Some Conceptual and Operational Considerations when Measuring ‘Resilience’: A Response to Hodgson et al. Trends Ecol. Evol. 2016, 31, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.; Scherer-Lorenzen, M.; Weigelt, A. Grassland Resistance and Resilience after Drought Depends on Management Intensity and Species Richness. PLoS ONE 2012, 7, e36992. [Google Scholar] [CrossRef]
- Pimm, S.L. The complexity and stability of ecosystems. Nat. Cell Biol. 1984, 307, 321–326. [Google Scholar] [CrossRef]
- Sasaki, T.; Furukawa, T.; Iwasaki, Y.; Seto, M.; Mori, A.S. Perspectives for ecosystem management based on ecosystem resilience and ecological thresholds against multiple and stochastic disturbances. Ecol. Indic. 2015, 57, 395–408. [Google Scholar] [CrossRef]
- Côté, I.M.; Darling, E.S. Rethinking Ecosystem Resilience in the Face of Climate Change. PLoS Biol. 2010, 8, e1000438. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Donner, S.D.; Maynard, J.A.; MacNeil, M.A.; Graham, N.A.; Maina, J.; Baker, A.C.; Beger, A.C.; Beger, M.; Campbell, S.J.; et al. Prioritizing Key Resilience Indicators to Support Coral Reef Management in a Changing Climate. PLoS ONE 2012, 7, e42884. [Google Scholar] [CrossRef]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.; Roy, D.B.; Procter, D.A.; Eigenbrod, F.; Freckleton, R.P.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. Biodiversity and Resilience of Ecosystem Functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef]
- Walker, B.; Holling, C.S.; Carpenter, S.R.; Kinzig, A.P. Resilience, Adaptability and Transformability in Social-ecological Systems. Ecol. Soc. 2004, 9, 3438–3447. [Google Scholar] [CrossRef]
- Mouillot, D.; Graham, N.A.J.; Villéger, S.; Mason, N.W.H.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Briske, D.D.; Fuhlendorf, S.D.; Smeins, F.E. State-and-Transition Models, Thresholds, and Rangeland Health: A Synthesis of Ecological Concepts and Perspectives. Rangel. Ecol. Manag. 2005, 58, 1–10. [Google Scholar] [CrossRef]
- Meyer, K.; Hoyer-Leitzel, A.; Iams, S.; Klasky, I.; Lee, V.; Ligtenberg, S.; Bussmann, E.; Zeeman, M.L. Quantifying resilience to recurrent ecosystem disturbances using flow–kick dynamics. Nat. Sustain. 2018, 1, 671–678. [Google Scholar] [CrossRef]
- Campos, G.E.; Moran, S.M.; Huete, A.; Zhang, Y.; Bresloff, S.; Huxman, T.E.; Eamus, E.; Bosch, D.D.; Buda, A.R.; Gunter, S.A.; et al. Ecosystem resilience despite large-scale altered hydroclimatic conditions. Nature 2013, 494, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Goyal, M.K. Assessment of ecosystem resilience to hydroclimatic disturbances in India. Glob. Chang. Biol. 2018, 24, e432–e441. [Google Scholar] [CrossRef]
- De Keersmaecker, W.; Lhermitte, S.; Tits, L.; Honnay, O.; Somers, B.; Coppin, P. A model quantifying global vegetation resistance and resilience to short-term climate anomalies and their relationship with vegetation cover. Glob. Ecol. Biogeogr. 2015, 24, 539–548. [Google Scholar] [CrossRef]
- MODIS. Available online: http://modis.gsfc.nasa.gov/ (accessed on 1 November 2019).
- USGS. Available online: http://tahoe.usgs.gov/DEM.html (accessed on 1 November 2019).
- CMDC. Available online: http://data.cma.cn/data/cdcdetail/dataCode/SURF_CLI_CHN_MUL_DAY_V3.0.html (accessed on 1 November 2019).
- REDCP, CAS. Available online: http://www.resdc.cn (accessed on 1 November 2019).
- Sannigrahi, S. Modeling terrestrial ecosystem productivity of an estuarine ecosystem in the Sundarban Biosphere Region, India using seven ecosystem models. Ecol. Model. 2017, 356, 73–90. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Zhu, B.; Tan, K.; Tao, S. Changes in vegetation net primary productivity from 1982 to 1999 in China. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Hao, X.; Ma, H.; Hua, D.; Qin, J.; Zhang, Y. Response of ecosystem water use efficiency to climate change in the Tianshan Mountains, Central Asia. Environ. Monit. Assess. 2019, 191, 561. [Google Scholar] [CrossRef]
- Saroar, M.; Rahman, M. Ecosystem-Based Adaptation: Opportunities and Challenges in Coastal Bangladesh: Policy Strategies for Adaptation and Resilience. In The Anthropocene: Politik Economics Society Science; Springer: Cham, Switzerland, 2019; Volume 28. [Google Scholar] [CrossRef]
- Lam, V.Y.Y.; Doropoulos, C.; Mumby, P.J. The influence of resilience-based management on coral reef monitoring: A systematic review. PLoS ONE 2017, 12, e0172064. [Google Scholar] [CrossRef]
- Fatemeh, H.; Reza, J.; Hossein, B.; Mostafa, T.; Kenneth, C.D. Estimation of spatial and temporal changes in net primary production based on Carnegie Ames Stanford Ap-proach(CASA) model in semi-arid rangelands of Semirom County, Iran. J. Arid Land 2019, 477–494. [Google Scholar]
- Jiang, Y.A.; Chen, Y.; Zhao, Y.Z.; Chen, P.X.; Yu, X.J.; Fan, J.; Bai, S.Q. Analysis on Changes of Basic Climatic Elements and Extreme Events in Xinjiang, China during 1961–2010. Adv. Clim. Chang. Res. 2013, 4, 20–29. [Google Scholar] [CrossRef]
- Xu, C.; Li, J.; Zhao, J.; Gao, S.; Chen, Y. Climate variations in northern Xinjiang of China over the past 50 years under global warming. Quat. Int. 2015, 358, 83–92. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.; Chen, Z. Spatial distribution and temporal trends of mean precipitation and extremes in the arid region, northwest of China, during 1960–2010. Hydrol. Process. 2012, 27, 1807–1818. [Google Scholar] [CrossRef]
- Kundzewicz, Z.W. Climate change impacts on the hydrological cycle. Ecohydrol. Hydrobiol. 2008, 8, 195–203. [Google Scholar] [CrossRef]
- Sun, S.; Sun, G.; Cohen, E.; McNulty, S.G.; Caldwell, P.V.; Duan, K.; Zhang, Y. Projecting water yield and ecosystem productivity across the United States by linking an ecohydrological model to WRF dynamically downscaled climate data. Hydrol. Earth Syst. Sci. 2016, 20, 935–952. [Google Scholar] [CrossRef]
- Sorg, A.; Bolch, T.; Stoffel, M.; Solomina, O.; Beniston, M. Climate change impacts on glaciers and runoff in Tien Shan (Central Asia). Nat. Clim. Chang. 2012, 2, 725–731. [Google Scholar] [CrossRef]
- Etemadi, H.; Samadi, S.; Sharifikia, M.; Smoak, J.M. Assessment of climate change downscaling and non-stationarity on the spatial pattern of a mangrove ecosystem in an arid coastal region of southern Iran. Theor. Appl. Clim. 2016, 126, 35–49. [Google Scholar] [CrossRef]
- Li, W.; Li, X.; Zhao, Y.; Zheng, S.; Bai, Y. Ecosystem structure, functioning and stability under climate change and grazing in grasslands: Current status and future prospects. Curr. Opin. Environ. Sustain. 2018, 33, 124–135. [Google Scholar] [CrossRef]
- Crabtree, R.; Potter, C.; Mullen, R.; Sheldon, J.; Huang, S.; Harmsen, J.; Rodman, A.; Jean, C. A modeling and spatio-temporal analysis framework for monitoring environmental change using NPP as an ecosystem indicator. Remote Sens. Environ. 2009, 113, 1486–1496. [Google Scholar] [CrossRef]
- Yang, H.; Mu, S.; Li, J. Effects of ecological restoration projects on land use and land cover change and its influences on territorial NPP in Xinjiang, China. Catena 2014, 115, 85–95. [Google Scholar] [CrossRef]
- Allen, R.G.; Irmak, A.; Trezza, R.; Hendrickx, J.M.H.; Bastiaanssen, W.G.M.; Kjaersgaard, J. Satellite-based ET estimation in agriculture using SEBAL and METRIC. Hydrol. Process. 2011, 25, 4011–4027. [Google Scholar] [CrossRef]
- Tegos, A.; Malamos, N.; Koutsoyiannis, D. A parsimonious regional parametric evapotranspiration model based on a sim-plification of the Penman–Monteith formula. J. Hydrol. 2015, 524, 708–717. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Hao, X.; Hao, H.; Zhang, J.; Li, Y. Comprehensive Assessment Indicator of Ecosystem Resilience in Central Asia. Water 2021, 13, 124. https://doi.org/10.3390/w13020124
Fan X, Hao X, Hao H, Zhang J, Li Y. Comprehensive Assessment Indicator of Ecosystem Resilience in Central Asia. Water. 2021; 13(2):124. https://doi.org/10.3390/w13020124
Chicago/Turabian StyleFan, Xue, Xingming Hao, Haichao Hao, Jingjing Zhang, and Yuanhang Li. 2021. "Comprehensive Assessment Indicator of Ecosystem Resilience in Central Asia" Water 13, no. 2: 124. https://doi.org/10.3390/w13020124
APA StyleFan, X., Hao, X., Hao, H., Zhang, J., & Li, Y. (2021). Comprehensive Assessment Indicator of Ecosystem Resilience in Central Asia. Water, 13(2), 124. https://doi.org/10.3390/w13020124