
Metal Bioaccumulation and Oxidative Stress in Ulva laetevirens in the Venice Lagoon: Early Warning Biomarker for Metal Bioaccumulation

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
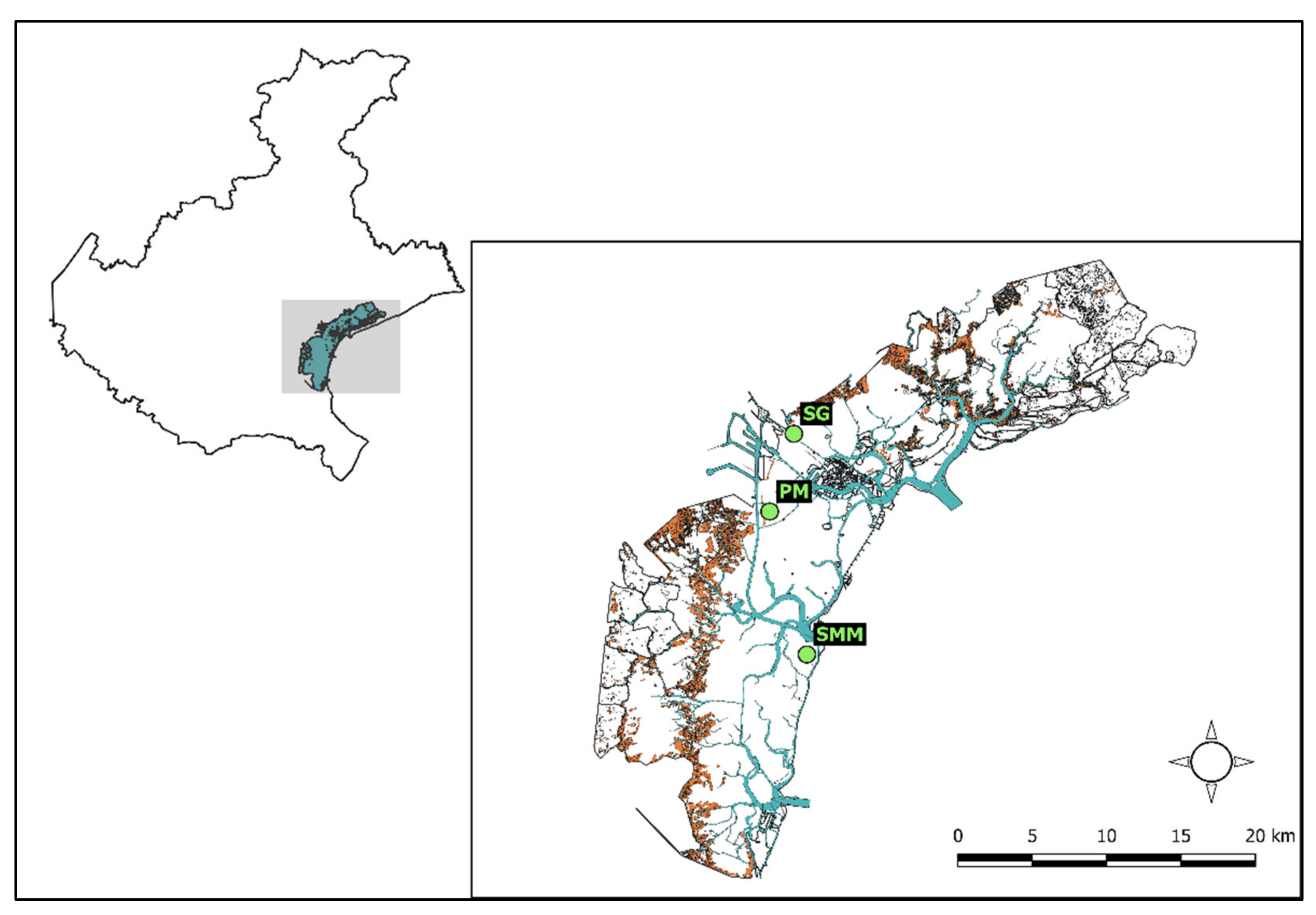
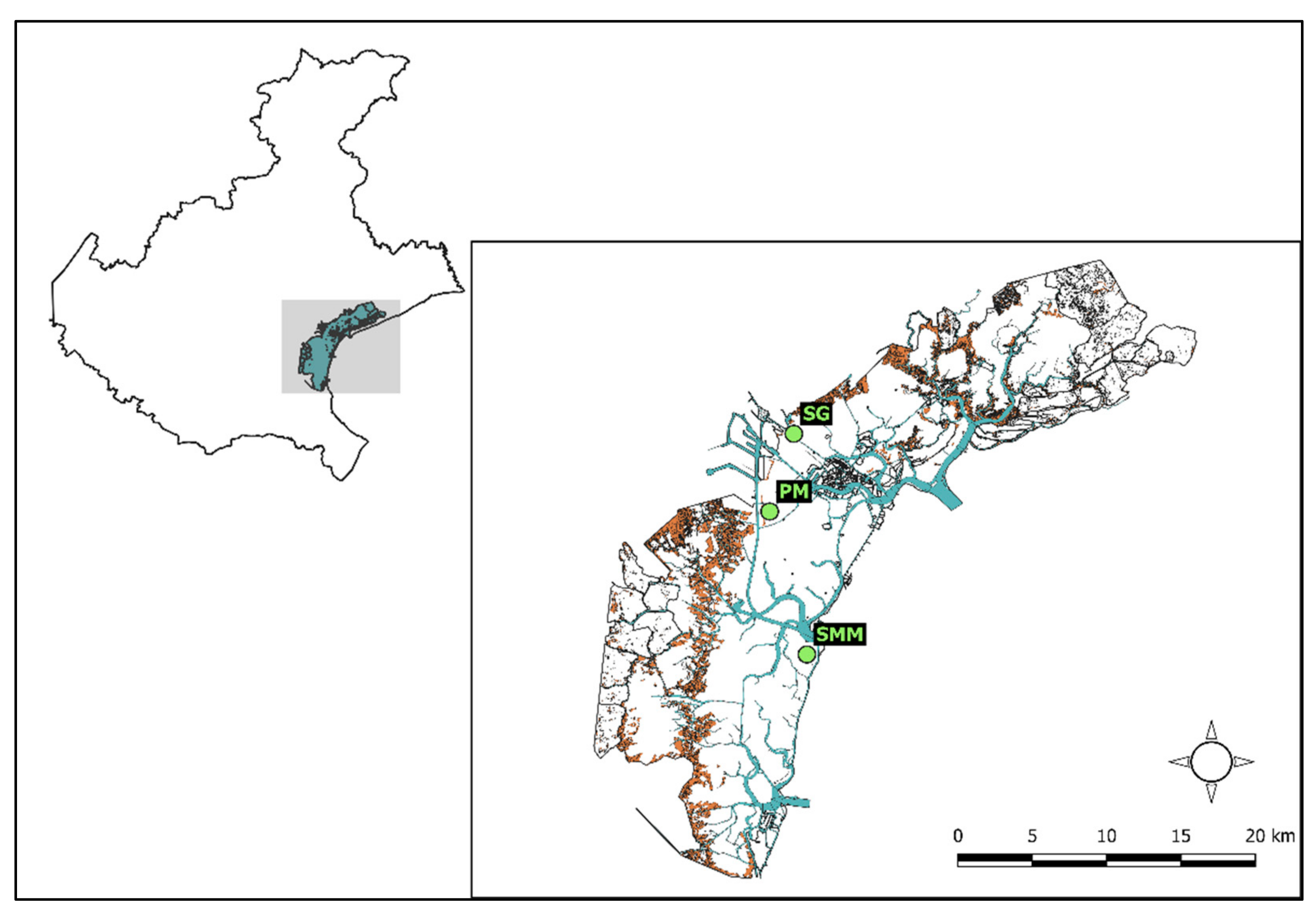
2.1. Study Area
2.2. Macroalgal Collection and Preparation
2.3. Water, Sediments, and SPM Collection and Preparation
2.4. Physicochemical Characteristics of the Sediment and SPM
2.5. Metal(Loid) Determination in Sediment, SPM, and Macroalgal Tissues
2.6. Biochemical Analysis in Macroalgal Tissues
2.7. Statistical Analyses
3. Results
3.1. Environmental Parameters for Water, Surface Sediments, and SPM
3.2. Metal(Loid)s in Sediments and SPM
3.3. Metals Accumulated in U. laetevirens
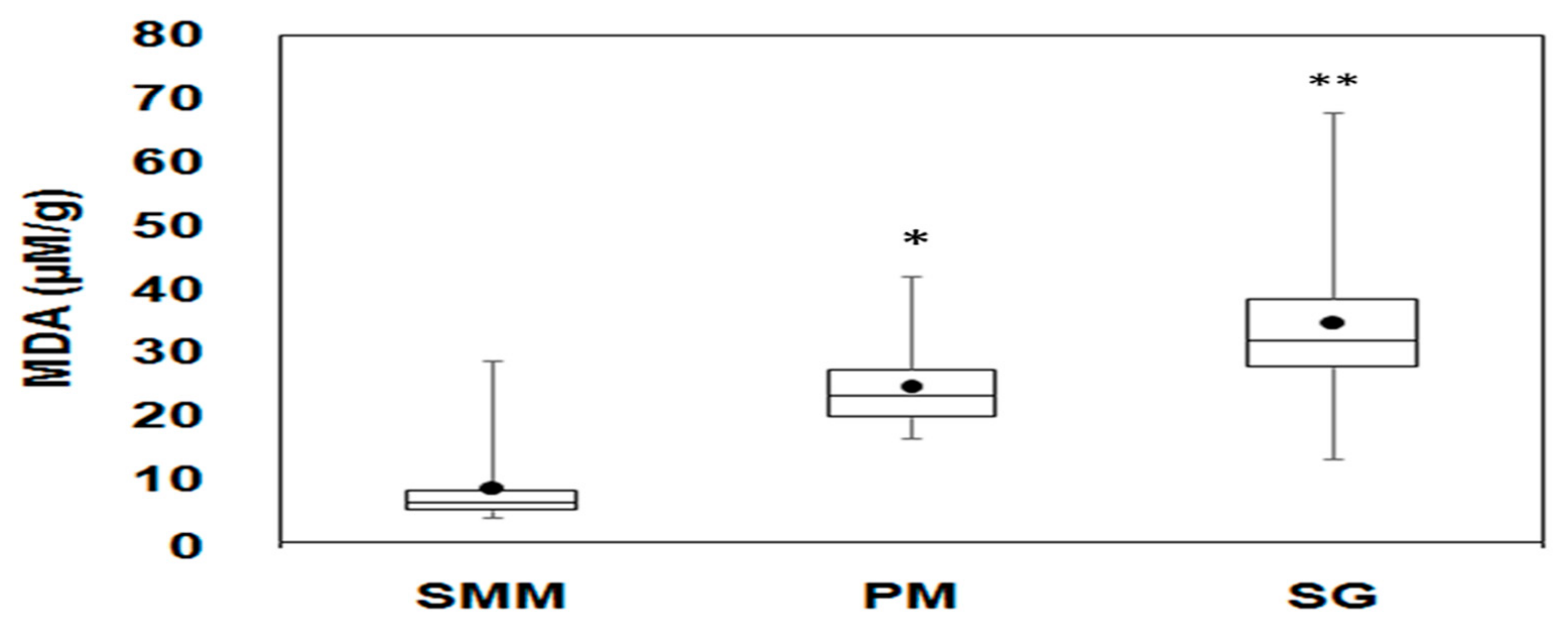
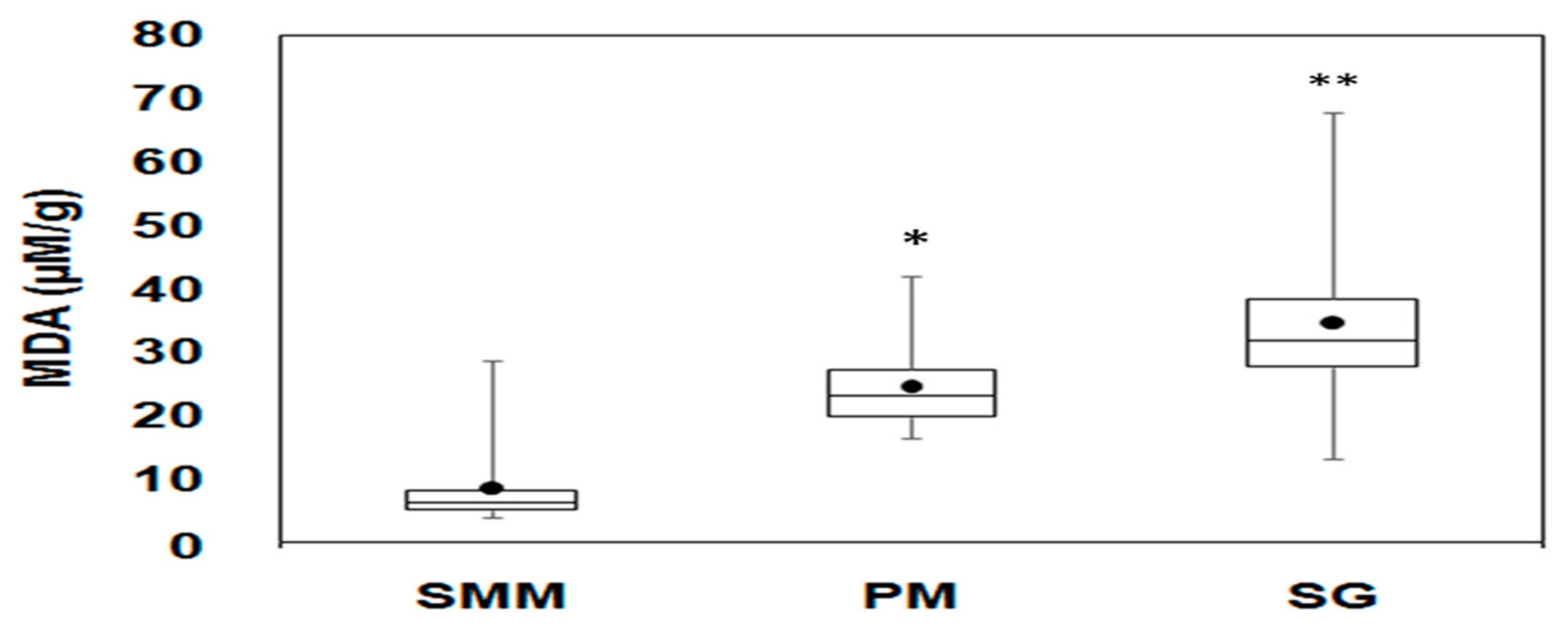
3.4. Oxidative Stress Biomarker (LPO)
4. Discussion
4.1. Metal Concentrations in Sediment and SPM
4.2. Metal Levels in U. laetevirens
4.3. Oxidative Stress and Metal Exposure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Le Guen, C.; Tecchio, S.; Dauvin, J.-C.; De Roton, G.; Lobry, J.; Lepage, M.; Morin, J.; Lassalle, G.; Raoux, A.; Niquil, N. Assessing the ecological status of an estuarine ecosystem: Linking biodiversity and food-web indicators. Estuar. Coast. Shelf Sci. 2019, 228, 106339. [Google Scholar] [CrossRef]
- Boerema, A.; Meire, P. Management for estuarine ecosystem services: A review. Ecol. Eng. 2017, 98, 172–182. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M.; Pérez-Marcos, M. Coastal lagoons: “Transitional ecosystems” between transitional and coastal waters. J. Coast. Conserv. 2011, 15, 369–392. [Google Scholar] [CrossRef]
- Wahsha, M.; Juhmani, A.-S.; Buosi, A.; Sfriso, A.; Sfriso, A. Assess the environmental health status of macrophyte ecosystems using an oxidative stress biomarker. Case studies: The Gulf of Aqaba and the Lagoon of Venice. Energy Procedia 2017, 125, 19–26. [Google Scholar] [CrossRef]
- Luoma, S.N.; Rainbow, P.S. Why Is Metal Bioaccumulation So Variable? Biodynamics as a Unifying Concept. Environ. Sci. Technol. 2005, 39, 1921–1931. [Google Scholar] [CrossRef] [PubMed]
- Falace, A.; Curiel, D.; Sfriso, A. Study of the macrophyte assemblages and application of phytobenthic indices to assess the Ecological Status of the Marano-Grado Lagoon (Italy). Mar. Ecol. 2009, 30, 480–494. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Ghetti, P.F. Rapid Quality Index (R-MaQI), based mainly on macrophyte associations, to assess the ecological status of Mediterranean transitional environments. Chem. Ecol. 2007, 23, 493–503. [Google Scholar] [CrossRef]
- Pereira, P.; de Pablo, H.; Dulce Subida, M.; Vale, C.; Pacheco, M. Biochemical responses of the shore crab (Carcinus maenas) in a eutrophic and metal-contaminated coastal system (Óbidos lagoon, Portugal). Ecotoxicol. Environ. Saf. 2009, 72, 1471–1480. [Google Scholar] [CrossRef]
- Villares, R.; Puente, X.; Carballeira, A. Ulva and Enteromorpha as indicators of heavy metal pollution. Hydrobiologia 2001, 462, 221–232. [Google Scholar] [CrossRef]
- Pereira, P.; de Pablo, H.; Rosa-Santos, F.; Pacheco, M.; Vale, C. Metal accumulation and oxidative stress in Ulva sp. substantiated by response integration into a general stress index. Aquat. Toxicol. 2009, 91, 336–345. [Google Scholar] [CrossRef]
- Sfriso, A.; Francesco Ghetti, P. Seasonal variation in biomass, morphometric parameters and production of seagrasses in the lagoon of Venice. Aquat. Bot. 1998, 61, 207–223. [Google Scholar] [CrossRef]
- Elliott, M.; Quintino, V. The Estuarine Quality Paradox, Environmental Homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar. Pollut. Bull. 2007, 54, 640–645. [Google Scholar] [CrossRef]
- Ahmad, I.; Mohmood, I.; Coelho, J.P.; Pacheco, M.; Santos, M.A.; Duarte, A.C.; Pereira, E. Role of non-enzymatic antioxidants on the bivalves’ adaptation to environmental mercury: Organ-specificities and age effect in Scrobicularia plana inhabiting a contaminated lagoon. Environ. Pollut. 2012, 163, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Alves de Almeida, E.; Celso Dias Bainy, A.; Paula de Melo Loureiro, A.; Regina Martinez, G.; Miyamoto, S.; Onuki, J.; Fujita Barbosa, L.; Carrião Machado Garcia, C.; Manso Prado, F.; Eliza Ronsein, G.; et al. Oxidative stress in Perna perna and other bivalves as indicators of environmental stress in the Brazilian marine environment: Antioxidants, lipid peroxidation and DNA damage. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 588–600. [Google Scholar] [CrossRef]
- Freitas, R.; Costa, E.; Velez, C.; Santos, J.; Lima, A.; Oliveira, C.; Maria Rodrigues, A.; Quintino, V.; Figueira, E. Looking for suitable biomarkers in benthic macroinvertebrates inhabiting coastal areas with low metal contamination: Comparison between the bivalve Cerastoderma edule and the Polychaete Diopatra neapolitana. Ecotoxicol. Environ. Saf. 2012, 75, 109–118. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Wahsha, M.; Bini, C.; Fontana, S.; Wahsha, A.; Zilioli, D. Toxicity assessment of contaminated soils from a mining area in Northeast Italy by using lipid peroxidation assay. J. Geochem. Explor. 2012, 113, 112–117. [Google Scholar] [CrossRef]
- Malea, P.; Rijstenbil, J.W.; Haritonidis, S. Effects of cadmium, zinc and nitrogen status on non-protein thiols in the macroalgae Enteromorpha spp. from the Scheldt Estuary (SW Netherlands, Belgium) and Thermaikos Gulf (N Aegean Sea, Greece). Mar. Environ. Res. 2006, 62, 45–60. [Google Scholar] [CrossRef]
- Nimptsch, J.; Pflugmacher, S. Ammonia triggers the promotion of oxidative stress in the aquatic macrophyte Myriophyllum mattogrossense. Chemosphere 2007, 66, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Ratkevicius, N.; Correa, J.A.; Moenne, A. Copper accumulation, synthesis of ascorbate and activation of ascorbate peroxidase in Enteromorpha compressa (L.) Grev. (Chlorophyta) from heavy metal-enriched environments in northern Chile. Plant Cell Environ. 2003, 26, 1599–1608. [Google Scholar] [CrossRef]
- Ravera, O. The Lagoon of Venice: The result of both natural factors and human influence. J. Limnol. 2000, 59, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Sfriso, A.; Facca, C.; Ceoldo, S.; Silvestri, S.; Ghetti, P.F. Role of macroalgal biomass and clam fishing on spatial and temporal changes in N and P sedimentary pools in the central part of the Venice lagoon. Oceanol. Acta 2003, 26, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Sfriso, A.; Facca, C.; Raccanelli, S. PCDD/F and dioxin-like PCB bioaccumulation by Manila clam from polluted areas of Venice lagoon (Italy). Environ. Pollut. 2014, 184, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Sfriso, A.; Argese, E.; Bettiol, C.; Facca, C. Tapes philippinarum seed exposure to metals in polluted areas of the Venice lagoon. Estuar. Coast. Shelf Sci. 2008, 79, 581–590. [Google Scholar] [CrossRef]
- Adriano, S.; Facca, C.; Marcomini, A. Sedimentation rates and erosion processes in the lagoon of Venice. Environ. Int. 2005, 31, 983–992. [Google Scholar] [CrossRef]
- Bellucci, L.G.; Frignani, M.; Paolucci, D.; Ravanelli, M. Distribution of heavy metals in sediments of the Venice Lagoon: The role of the industrial area. Sci. Total Environ. 2002, 295, 35–49. [Google Scholar] [CrossRef]
- Secco, T.; Pellizzato, F.; Sfriso, A.; Pavoni, B. The changing state of contamination in the Lagoon of Venice. Part 1: Organic pollutants. Chemosphere 2005, 58, 279–290. [Google Scholar] [CrossRef]
- Rigollet, V.; Sfriso, A.; Marcomini, A.; De Casabianca, M.L. Seasonal evolution of heavy metal concentrations in the surface sediments of two Mediterranean Zostera marina L. beds at Thau lagoon (France) and Venice lagoon (Italy). Bioresour. Technol. 2004, 95, 159–167. [Google Scholar] [CrossRef]
- Zonta, R.; Botter, M.; Cassin, D.; Pini, R.; Scattolin, M.; Zaggia, L. Sediment chemical contamination of a shallow water area close to the industrial zone of Porto Marghera (Venice Lagoon, Italy). Mar. Pollut. Bull. 2007, 55, 529–542. [Google Scholar] [CrossRef]
- Raccanelli, S.; Favotto, M.; Vio, P. Estimation of PCDD/F and PCB detoxification rates in contaminated Tapes philippinarum in the Lagoon of Venice. Chemosphere 2008, 73, S166–S170. [Google Scholar] [CrossRef]
- Masiol, M.; Facca, C.; Visin, F.; Sfriso, A.; Pavoni, B. Interannual heavy element and nutrient concentration trends in the top sediments of Venice Lagoon (Italy). Mar. Pollut. Bull. 2014, 89, 49–58. [Google Scholar] [CrossRef]
- Guerzoni, S.; Tagliapietra, D. Atlante della Laguna; Venezia: Venice, Italy, 2006. [Google Scholar]
- Guerzoni, S.; Rossini, P.; Molinaroli, E.; Rampazzo, G.; Raccanelli, S. Measurement of atmospheric deposition of polychlorinated dibenzo-p-dioxins and dibenzofurans in the Lagoon of Venice, Italy. Chemosphere 2004, 54, 1309–1317. [Google Scholar] [CrossRef] [Green Version]
- Sfriso, A.; CFacca, H.; Ceoldo, S.; Marcomini, A. Recording the occurrence of trophic level changes in the lagoon of Venice over the ’90s. Environ. Int. 2005, 31, 993–1001. [Google Scholar] [CrossRef]
- Oxner, M.; Knudsen, M. The Determination of Chlorinity by the Knudsen Method; GM Manufacturing Company: New York, NY, USA, 1962; 63p. [Google Scholar]
- Byers, S.C.; Mills, E.L.; Stewart, P.L. A comparison of methods of determining organic carbon in marine sediments, with suggestions for a standard method. Hydrobiologia 1978, 58, 43–47. [Google Scholar] [CrossRef]
- Aspila, K.I.; Agemian, H.; Chau, A.S.Y. A semi-automated method for the determination of inorganic, organic and total phosphate in sediments. Analyst 1976, 101, 187–197. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A.; Tomio, Y.; Juhmani, A.-S.; Chiesa, S.; Greco, M.; Gazzola, C.; Mistri, M.; Munari, C.; Sfriso, A.A. Sediment carbon variations in the venice lagoon and other transitional water systems of the northern adriatic sea. Water 2020, 12, 3430. (In Switzerland) [Google Scholar] [CrossRef]
- Figueira, E.; Lima, A.; Branco, D.; Quintino, V.; Rodrigues, A.M.; Freitas, R. Health concerns of consuming cockles (Cerastoderma edule L.) from a low contaminated coastal system. Environ. Int. 2011, 37, 965–972. [Google Scholar] [CrossRef]
- Usero, J.; González-Regalado, E.; Gracia, I. Trace metals in the bivalve molluscs Ruditapes decussatus and Ruditapes philippinarum from the atlantic coast of Southern Spain. Environ. Int. 1997, 23, 291–298. [Google Scholar] [CrossRef]
- Velez, C.; Figueira, E.; Soares, A.; Freitas, R. Spatial distribution and bioaccumulation patterns in three clam populations from a low contaminated ecosystem. Estuar. Coast. Shelf Sci. 2015, 155, 114–125. [Google Scholar] [CrossRef]
- Ministero per l’Ambiente. Criteri di Sicurezza Ambientale per lo Scavo, Trasporto e Riutilizzo di Sedimenti dei Canali di Venezia; Ministero per l’Ambiente: Rome, Italy, 1993; 23p.
- Picone, M.; Bergamin, M.; Losso, C.; Delaney, E.; Arizzi Novelli, A.; Ghirardini, A.V. Assessment of sediment toxicity in the Lagoon of Venice (Italy) using a multi-species set of bioassays. Ecotoxicol. Environ. Saf. 2016, 123, 32–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moschino, V.; Delaney, E.; Meneghetti, F.; Ros, L. Da Biomonitoring approach with mussel Mytilus galloprovincialis (Lmk) and clam Ruditapes philippinarum (Adams and Reeve, 1850) in the Lagoon of Venice. Environ. Monit. Assess. 2011, 177, 649–663. [Google Scholar] [CrossRef]
- Breda, S.; Chiesa, S.; Freitas, R.; Figueira, E.; Becherini, F.; Gobbo, L.; Soares, A.M.V.M.; Argese, E. Biogeochemical dynamics and bioaccumulation processes in Manila clam: Implications for biodiversity and ecosystem services in the Ria de Aveiro Lagoon. Estuar. Coast. Shelf Sci. 2018, 209, 136–148. [Google Scholar] [CrossRef]
- Sfriso, A.A.; Chiesa, S.; Sfriso, A.; Buosi, A.; Gobbo, L.; Boscolo Gnolo, A.; Argese, E. Spatial distribution, bioaccumulation profiles and risk for consumption of edible bivalves: A comparison among razor clam, Manila clam and cockles in the Venice Lagoon. Sci. Total Environ. 2018, 643, 579–591. [Google Scholar] [CrossRef]
- Carvalho, S.; Moura, A.; Gaspar, M.B.; Pereira, P.; Cancela da Fonseca, L.; Falcão, M.; Drago, T.; Leitão, F.; Regala, J. Spatial and inter-annual variability of the macrobenthic communities within a coastal lagoon (Óbidos lagoon) and its relationship with environmental parameters. Acta Oecologica 2005, 27, 143–159. [Google Scholar] [CrossRef]
- Carvalho, S.; Barata, M.; Pereira, F.; Gaspar, M.B.; Cancela da Fonseca, L.; Pousão-Ferreira, P. Distribution patterns of macrobenthic species in relation to organic enrichment within aquaculture earthen ponds. Mar. Pollut. Bull. 2006, 52, 1573–1584. [Google Scholar] [CrossRef]
- Cecchi, R.; Ghermandi, G.; Zonta, R. Geochemical processes in Venice Lagoon by PIXE technique: An overview. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 1996, 109–110, 407–414. [Google Scholar] [CrossRef]
- Hornberger, M.I.; Luoma, S.N.; van Geen, A.; Fuller, C.; Anima, R. Historical trends of metals in the sediments of San Francisco Bay, California. Mar. Chem. 1999, 64, 39–55. [Google Scholar] [CrossRef]
- Zonta, R.; Zaggia, L.; Argese, E. Heavy metal and grain-size distributions in estuarine shallow water sediments of the Cona Marsh (Venice Lagoon, Italy). Sci. Total Environ. 1994, 151, 19–28. [Google Scholar] [CrossRef]
- Casado-Martinez, M.C.; Smith, B.D.; DelValls, T.A.; Luoma, S.N.; Rainbow, P.S. Biodynamic modelling and the prediction of accumulated trace metal concentrations in the polychaete Arenicola marina. Environ. Pollut. 2009, 157, 2743–2750. [Google Scholar] [CrossRef]
- Rainbow, P.S.; Luoma, S.N. Metal toxicity, uptake and bioaccumulation in aquatic invertebrates—Modelling zinc in crustaceans. Aquat. Toxicol. 2011, 105, 455–465. [Google Scholar] [CrossRef]
- Molinaroli, E.; Guerzoni, S.; De Falco, G.; Sarretta, A.; Cucco, A.; Como, S.; Simeone, S.; Perilli, A.; Magni, P. Relationships between hydrodynamic parameters and grain size in two contrasting transitional environments: The Lagoons of Venice and Cabras, Italy. Sediment. Geol. 2009, 219, 196–207. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, M.T.S.D.; Leal, M.F.C. Antagonistic interactions of Pb and Cd on Cu uptake, growth inhibition and chelator release in the marine algae Emiliania huxleyi. Mar. Chem. 2001, 75, 123–139. [Google Scholar] [CrossRef]
- Caliceti, M.; Argese, E.; Sfriso, A.; Pavoni, B. Heavy metal contamination in the seaweeds of the Venice lagoon. Chemosphere 2002, 47, 443–454. [Google Scholar] [CrossRef]
- Fritioff, Å.; Kautsky, L.; Greger, M. Influence of temperature and salinity on heavy metal uptake by submersed plants. Environ. Pollut. 2005, 133, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Malea, P.; Haritonidis, S. Use of the green alga Ulva rigida C. Agardh as an indicator species to reassess metal pollution in the Thermaikos Gulf, Greece, after 13 years. J. Appl. Phycol. 2000, 12, 169–176. [Google Scholar] [CrossRef]
- Villares, R.; Puente, X.; Carballeira, A. Seasonal variation and background levels of heavy metals in two green seaweeds. Environ. Pollut. 2002, 119, 79–90. [Google Scholar] [CrossRef]
- Jarvis, T.A.; Bielmyer-Fraser, G.K. Accumulation and effects of metal mixtures in two seaweed species. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 171, 28–33. [Google Scholar] [CrossRef]
- Lee, W.Y.; Wang, W.X. Metal accumulation in the green macroalga Ulva fasciata: Effects of nitrate, ammonium and phosphate. Sci. Total Environ. 2001, 278, 11–22. [Google Scholar] [CrossRef]
- Bibak, M.; Tahmasebi, S.; Sattari, M.; Kafaei, R.; Ramavandi, B. Empirical cumulative entropy as a new trace elements indicator to determine the relationship between algae-sediment pollution in the Persian Gulf, southern Iran. Environ. Sci. Pollut. Res. Int. 2021, 28, 4634–4644. [Google Scholar] [CrossRef]
- Alahverdi, M.; Savabieasfahani, M. Metal pollution in seaweed and related sediment of the Persian Gulf, Iran. Bull. Environ. Contam. Toxicol. 2012, 88, 939–945. [Google Scholar] [CrossRef]
- Argese, E.; Bettiol, C. Heavy metal partitioning in sediments from the lagoon of Venice (Italy). Toxicol. Environ. Chem. 2001, 79, 157–170. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C. Annual growth and environmental relationships of the invasive species Sargassum muticum and Undaria pinnatifida in the lagoon of Venice. Estuar. Coast. Shelf Sci. 2013, 129, 162–172. [Google Scholar] [CrossRef]
- Escobar, J.A.; Rubio, M.A.; Lissi, E.A. SOD and catalase inactivation by singlet oxygen and peroxyl radicals. Free Radic. Biol. Med. 1996, 20, 285–290. [Google Scholar] [CrossRef]
- Pereira, P.; Raimundo, J.; Araújo, O.; Canário, J.; Almeida, A.; Pacheco, M. Fish eyes and brain as primary targets for mercury accumulation—A new insight on environmental risk assessment. Sci. Total Environ. 2014, 494–495, 290–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasiotis, K.M.; Emmanouil, C.; Anastasiadou, P.; Papadi-Psyllou, A.; Papadopoulos, A.; Okay, O.; Machera, K. Organic pollution and its effects in the marine mussel Mytilus galloprovincialis in Eastern Mediterranean coasts. Chemosphere 2015, 119, S145–S152. [Google Scholar] [CrossRef]
- Emmanouil, C.; Green, R.M.; Willey, F.R.; Chipman, J.K. Oxidative damage in gill of Mytilus edulis from Merseyside, UK, and reversibility after depuration. Environ. Pollut. 2008, 151, 663–668. [Google Scholar] [CrossRef]
- Vidal, M.-L.; Bassères, A.; Narbonne, J.-F. Seasonal variations of pollution biomarkers in two populations of Corbicula fluminea (Müller). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 131, 133–151. [Google Scholar] [CrossRef]
- Lin, H.; Sun, T.; Zhou, Y.; Zhang, X. Anti-oxidative feedback and biomarkers in the intertidal seagrass Zostera japonica induced by exposure to copper, lead and cadmium. Mar. Pollut. Bull. 2016, 109, 325–333. [Google Scholar] [CrossRef]
- Marques, A.; Piló, D.; Carvalho, S.; Araújo, O.; Guilherme, S.; Santos, M.A.; Vale, C.; Pereira, F.; Pacheco, M.; Pereira, P. Metal bioaccumulation and oxidative stress profiles in Ruditapes philippinarum—insights towards its suitability as bioindicator of estuarine metal contamination. Ecol. Indic. 2018, 95, 1087–1099. [Google Scholar] [CrossRef]
- Kournoutou, G.G.; Giannopoulou, P.C.; Sazakli, E.; Leotsinidis, M.; Kalpaxis, D.L.; Dinos, G.P. Oxidative Damage of Mussels Living in Seawater Enriched with Trace Metals, from the Viewpoint of Proteins Expression and Modification. Toxics 2020, 8, 89. [Google Scholar] [CrossRef]
- Kournoutou, G.G.; Giannopoulou, P.C.; Sazakli, E.; Leotsinidis, M.; Kalpaxis, D.L. Oxidative damage of 18S and 5S ribosomal RNA in digestive gland of mussels exposed to trace metals. Aquat. Toxicol. 2017, 192, 136–147. [Google Scholar] [CrossRef]
- Elbaz, A.; Wei, Y.Y.; Meng, Q.; Zheng, Q.; Yang, Z.M. Mercury-induced oxidative stress and impact on antioxidant enzymes in Chlamydomonas reinhardtii. Ecotoxicology 2010, 19, 1285–1293. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Wang, S.J.; Yang, Z.M. Biological detection and analysis of mercury toxicity to alfalfa (Medicago sativa) plants. Chemosphere 2008, 70, 1500–1509. [Google Scholar] [CrossRef]
- Chen, J.; Shiyab, S.; Han, F.X.; Monts, D.L.; Waggoner, C.A.; Yang, Z.; Su, Y. Bioaccumulation and physiological effects of mercury in Pteris vittata and Nephrolepis exaltata. Ecotoxicology 2008, 18, 110. [Google Scholar] [CrossRef]
- Ratte, H.T. Bioaccumulation and toxicity of silver compounds: A review. Environ. Toxicol. Chem. 1999, 18, 89–108. [Google Scholar] [CrossRef]
- Bloom, N.S.; Lasorsa, B.K. Changes in mercury speciation and the release of methyl mercury as a result of marine sediment dredging activities. Sci. Total Environ. 1999, 237–238, 379–385. [Google Scholar] [CrossRef]
- Bloom, N.S.; Moretto, L.M.; Ugo, P. A comparison of the speciation and fate of mercury in two contaminated coastal marine ecosystems: The Venice Lagoon (Italy) and Lavaca Bay (Texas). Limnol. Oceanogr. 2004, 49, 367–375. [Google Scholar] [CrossRef]
- Favero, N.; Cattalini, F.; Bertaggia, D.; Albergoni, V. Metal accumulation in a biological indicator (Ulva rigida) from the lagoon of Venice (Italy). Arch. Environ. Contam. Toxicol. 1996, 31, 9–18. [Google Scholar] [CrossRef]
- Sfriso, A.; Marcomini, A.; Pavoni, B. Annual nutrient exchanges between the central lagoon of Venice and the northern Adriatic Sea. Sci. Total Environ. 1994, 156, 77–92. [Google Scholar] [CrossRef]
- Sfriso, A. Coexistence of Ulva rigida and Ulva laetevirens (Ulvales, Chlorophyta) in Venice Lagoon and other Italian transitional and marine environments. Bot. Mar. 2010, 53, 9–18. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
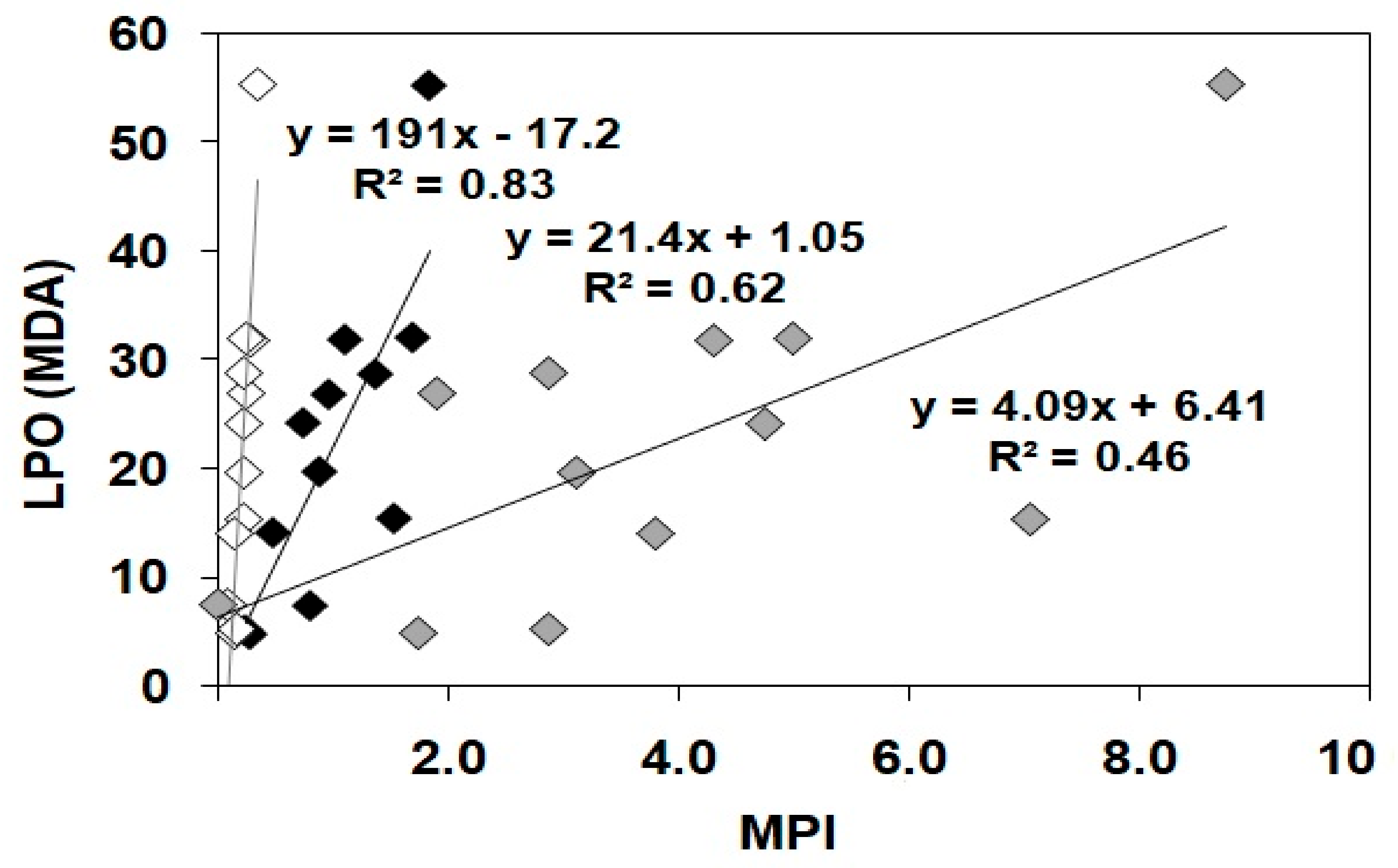{kind=link}
| Site | As | V | Ni | Mn | Zn | Cu | Pb | Hg | MPI | |
|---|---|---|---|---|---|---|---|---|---|---|
| SMM | Sediment | 1.00 ± 0.14 | 14.0 ± 0.21 | 8.65 ± 1.63 | 258 ± 1.77 | 17.3 ± 1.13 | 6.40 ± 0.84 | 4.75 ± 0.07 | <d.l. | 10.7 |
| PM | 4.89 ± 0.21 | 32.5 ± 1.27 | 22.0 ± 0.85 | 242 ± 1.34 | 327 ± 3.39 | 28.4 ± 1.06 | 68.1 ± 1.83 | 1.65 ± 0.05 | 31.1 | |
| SG | 11.2 ± 0.89 | 22.9 ± 0.63 | 15.8 ± 1.27 | 368 ± 28.9 | 428 ± 12.5 | 45.3 ± 2.76 | 56.1 ± 0.98 | 0.85 ± 0.02 | 32.9 | |
| SMM | SPM | 13.6 ± 3.39 | 19.1 ± 0.47 | 14.6 ± 0.85 | 365 ± 0.70 | 46.7 ± 0.07 | 9.17 ± 0.22 | 13.3 ± 0.78 | 0.29 ± 0.03 | 14.8 |
| PM | 16.9 ± 0.49 | 53.6 ± 1.41 | 22.1 ± 0.35 | 485 ± 3.53 | 659 ± 2.12 | 49.1 ± 0.85 | 72.8 ± 0.21 | 1.09 ± 0.04 | 47.3 | |
| SG | 17.8 ± 3.89 | 39.5 ± 0.85 | 23.5 ± 0.21 | 606 ± 2.83 | 325 ± 2.83 | 49.2 ± 0.01 | 45.4 ± 0.78 | 0.87 ± 0.05 | 39.9 |
| Site | As | V | Ni | Mn | Zn | Cu | Pb | Hg | MPI |
|---|---|---|---|---|---|---|---|---|---|
| SMM | 1.6 ± 0.7 | 10 ± 10.6 | 2.4 ± 1.0 | 38 ± 20.9 | 14.7 ± 11.1 | 8.6 ± 2.8 | 0.3 ± 0.3 | 0.1 ± 0.1 | 2.99 |
| PM | 9.9 ± 3.7 | 2.3 ± 2.0 | 1.4 ± 0.5 | 31.3 ± 11.6 | 33.3 ± 24.1 | 28.8 ± 30.5 | 0.6 ± 0.5 | 0.2 ± 0.1 | 4.37 |
| SG | 4.8 ± 1.2 | 1.5 ± 1.6 | 1.9 ± 1.0 | 164.8 ± 159.4 | 56.2 ± 35.3 | 29.2 ± 22.0 | 2.7 ± 1.3 | 0.3 ± 0.1 | 6.33 |
| Ulva/Sediment | As | V | Ni | Mn | Zn | Cu | Pb | Hg |
|---|---|---|---|---|---|---|---|---|
| SMM | 1.62 | 0.72 | 0.28 | 0.15 | 0.85 | 1.35 | 0.05 | 0 |
| PM | 2.03 | 0.07 | 0.06 | 0.13 | 0.10 | 1.02 | 0.01 | 0.14 |
| SG | 0.43 | 0.06 | 0.12 | 0.45 | 0.13 | 0.64 | 0.05 | 0.30 |
| Average | 1.36 | 0.28 | 0.16 | 0.24 | 0.36 | 1.00 | 0.04 | 0.15 |
| Ulva/SPM | As | V | Ni | Mn | Zn | Cu | Pb | Hg |
| SMM | 0.12 | 0.53 | 0.17 | 0.10 | 0.31 | 0.94 | 0.02 | 0.44 |
| PM | 0.59 | 0.04 | 0.06 | 0.06 | 0.05 | 0.59 | 0.01 | 0.21 |
| SG | 0.27 | 0.04 | 0.08 | 0.27 | 0.17 | 0.59 | 0.06 | 0.30 |
| Average | 0.33 | 0.20 | 0.10 | 0.15 | 0.18 | 0.71 | 0.03 | 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juhmani, A.-S.; Sfriso, A.A.; Wahsha, M.; Buosi, A.; Tomio, Y.; Sfriso, A. Metal Bioaccumulation and Oxidative Stress in Ulva laetevirens in the Venice Lagoon: Early Warning Biomarker for Metal Bioaccumulation. Water 2021, 13, 2626. https://doi.org/10.3390/w13192626
Juhmani A-S, Sfriso AA, Wahsha M, Buosi A, Tomio Y, Sfriso A. Metal Bioaccumulation and Oxidative Stress in Ulva laetevirens in the Venice Lagoon: Early Warning Biomarker for Metal Bioaccumulation. Water. 2021; 13(19):2626. https://doi.org/10.3390/w13192626
Chicago/Turabian StyleJuhmani, Abdul-Salam, Andrea A. Sfriso, Mohammad Wahsha, Alessandro Buosi, Yari Tomio, and Adriano Sfriso. 2021. "Metal Bioaccumulation and Oxidative Stress in Ulva laetevirens in the Venice Lagoon: Early Warning Biomarker for Metal Bioaccumulation" Water 13, no. 19: 2626. https://doi.org/10.3390/w13192626
APA StyleJuhmani, A.-S., Sfriso, A. A., Wahsha, M., Buosi, A., Tomio, Y., & Sfriso, A. (2021). Metal Bioaccumulation and Oxidative Stress in Ulva laetevirens in the Venice Lagoon: Early Warning Biomarker for Metal Bioaccumulation. Water, 13(19), 2626. https://doi.org/10.3390/w13192626