
Intra-Body Variations of Stable Isotope Ratios (δ13C, δ15N) and Influence of Storage Methods in Aquatic and Post-Aquatic Stages of the Common Toad, Bufo bufo


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sampling
2.2. Stable Isotope Analysis
2.3. Data Analysis
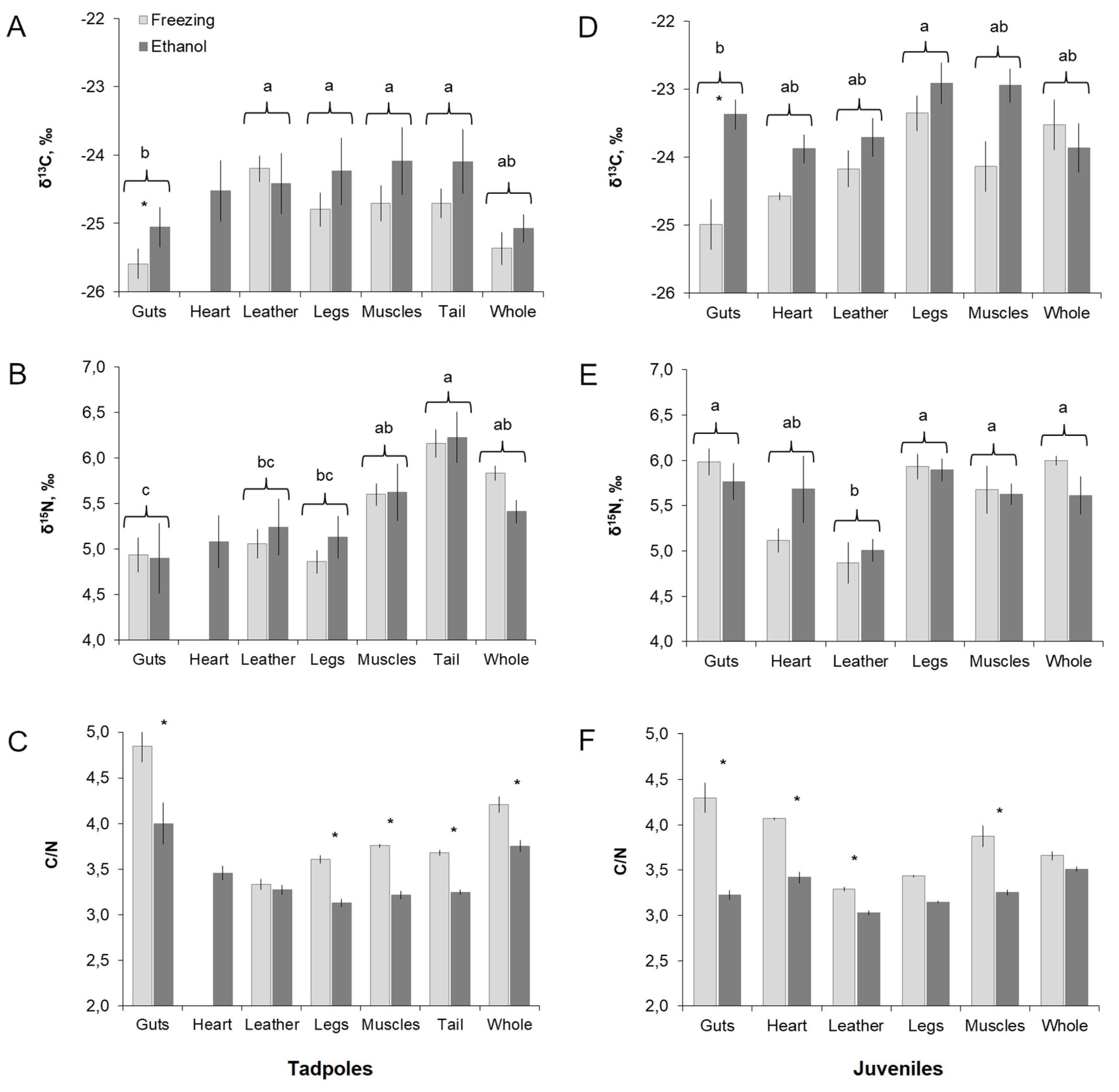
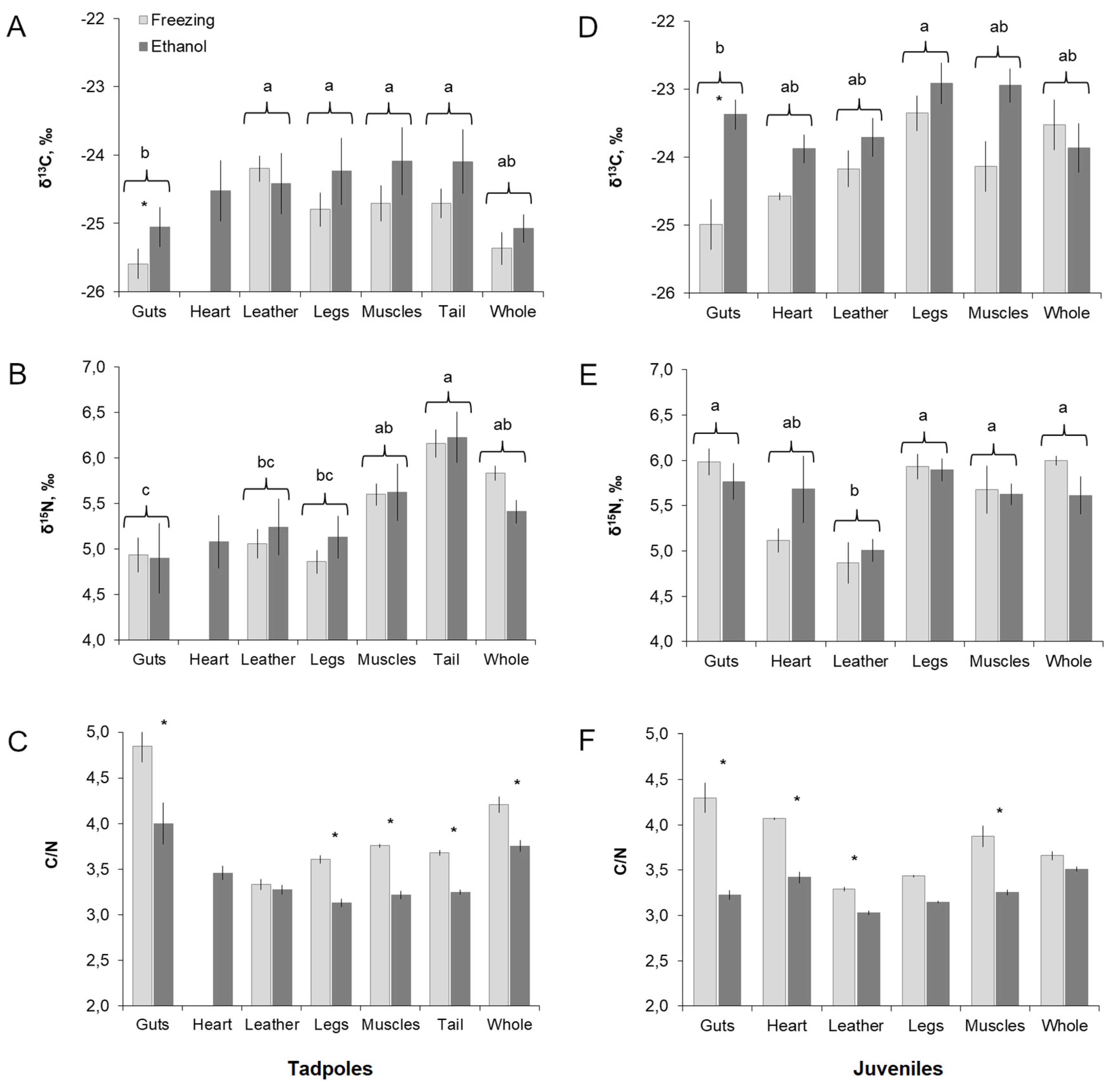
3. Results
3.1. “Tissue” and “Storage Method” Factors in Tadpoles
3.2. “Tissue” and “Storage Method” Factors in Juveniles
3.3. The Tissue Isotope Composition and Effect of Ethanol: Tadpoles vs. Juveniles
4. Discussion
4.1. Differences in Stable Isotope Signatures of Tissues/Organs and Influence of Storage Methods
4.1.1. Tissues/Organs
4.1.2. Storage Method
4.2. Practical Recommendations Concerning Choice of Tissues/Organs and Preservation Methods (Freezing vs. Ethanol) for Stable Isotope Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peterson, B.J.; Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Evol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Matich, P.; Rosenblatt, A.E.; Vaudo, J.J.; Yeager, L.A.; et al. Applying stable isotopes to examine food-web structure: An overview of analytical tools. Biol. Rev. 2011, 87, 545–562. [Google Scholar] [CrossRef]
- Boecklen, W.J.; Yarnes, C.T.; Cook, B.A.; James, A.C. On the use of stable isotopes in trophic ecology. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 411–440. [Google Scholar] [CrossRef] [Green Version]
- Araujo-Lima, C.; Forsberg, B.R.; Victoria, R.; Martinelli, L. Energy Sources for Detrivorous Fishes in the Amazon. Science 1986, 234, 1256–1258. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.B.; Polis, G.A. Marine subsidies of island communities in the Gulf of California: Evidence from stable carbon and nitrogen isotopes. Oikos 1998, 81, 75–80. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Hobson, K.A. Tracing origins and migration of wildlife using stable isotopes: A review. Oecologia 1999, 120, 314–326. [Google Scholar] [CrossRef]
- Newsome, S.D.; Yeakel, J.D.; Wheatley, P.V.; Tinker, M.T. Tools for quantifying isotopic niche space and dietary variation at the individual and population level. J. Mammal. 2012, 93, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, O.; Scrimgeour, C.M.; Curry, J.P. Carbon and nitrogen stable isotope ratios in body tissue and mucus of feeding and fasting earthworms (Lumbricus festivus). Oecologia 1999, 118, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.L.; Eldridge, P.M. Estimating the timing of diet shifts using stable isotopes. Oecologia 2006, 147, 195–203. [Google Scholar] [CrossRef]
- Gratton, C.; Forbes, A. Changes in δ13C stable isotopes in multiple tissues of insect predators fed isotopically distinct prey. Oecologia 2006, 147, 615–624. [Google Scholar] [CrossRef]
- Klarner, B.; Maraun, M.; Scheu, S. Trophic diversity and niche partitioning in a species rich predator guild—Natural variations in stable isotope ratios (13C/12C, 15N/14N) of mesostigmatid mites (Acari, Mesostigmata) from Central European beech forests. Soil Biol. Biochem. 2013, 57, 327–333. [Google Scholar] [CrossRef]
- Schilder, J.; Tellenbach, C.; Möst, M.; Spaak, P.; van Hardenbroek, M.; Wooller, M.J.; Heiri, O. The stable isotopic composition of Daphnia ephippia reflects changes in δ13C and δ18O values of food and water. Biogeosciences 2015, 12, 3819–3830. [Google Scholar] [CrossRef] [Green Version]
- Potapov, A.M.; Tiunov, A.V.; Scheu, S. Uncovering trophic positions and food resources of soil animals using bulk natural stable isotope composition. Biol. Rev. 2019, 94, 37–59. [Google Scholar] [CrossRef]
- Deniro, M.J.; Epstein, S. Influence of diet on distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- Webb, S.C.; Hedges, R.E.M.; Simpson, S.J. Diet quality influences the δ13C and δ15N of locusts and their biochemical components. J. Exp. Biol. 1998, 201, 2903–2911. [Google Scholar] [CrossRef]
- Shipley, O.N.; Matich, P. Studying animal niches using bulk stable isotope ratios: An updated synthesis. Oecologia 2020, 193, 27–51. [Google Scholar] [CrossRef] [PubMed]
- Semenyuk, I.I.; Tiunov, A.V. Intraspecific dispersion and age changes of isotopic composition (15N/14N, 13C/12C) of diplopod tissues (Myriapoda, Diplopoda). Izv. Penz. Gos. Pedagog. Univ. Im. V.G. Belinskogo 2011, 25, 428–436. (In Russian) [Google Scholar]
- Wehi, P.; Hicks, B. Isotopic fractionation in a large herbivorous insect, the Auckland tree weta. J. Insect Physiol. 2010, 56, 1877–1882. [Google Scholar] [CrossRef] [PubMed]
- Perkins, M.J.; McDonald, R.A.; van Veen, F.J.F.; Kelly, S.D.; Rees, G.; Bearhop, S. Important impacts of tissue selection and lipid extraction on ecological parameters derived from stable isotope ratios. Methods Ecol. Evol. 2013, 4, 944–953. [Google Scholar] [CrossRef]
- Pinnegar, J.K.; Polunin, N.V.C. Differential fractionation of δ13C and δ15N among fish tissues: Implications for the study of trophic interactions. Funct. Ecol. 1999, 13, 225–231. [Google Scholar] [CrossRef]
- Murray, I.W.; Wolf, B.O. Diet and growth influence carbon incorporation rates and discrimination factors (Δ13C) in desert box turtles, Terrapene ornata luteola. Herpetol. Conserv. Biol. 2013, 8, 149–162. [Google Scholar]
- Warne, R.W.; Wolf, B.O. Nitrogen stable isotope turnover and discrimination in lizards. Rapid Commun. Mass Spectrom. 2021, 35, e9030. [Google Scholar] [CrossRef]
- Hobson, K.A.; Clark, R.G. Assessing avian diets using stable isotopes 2. Factors influencing diet-tissue fractionation. Condor 1992, 94, 189–197. [Google Scholar] [CrossRef]
- Lesage, V.; Hammill, M.O.; Kovacs, K.M. Diet-tissue fractionation of stable carbon and nitrogen isotopes in phocid seals. Mar. Mam. Sci. 2002, 18, 182–193. [Google Scholar] [CrossRef]
- Ehrich, D.; Tarroux, A.; Stien, J.; Lecomte, N.; Killengreen, S.; Berteaux, D.; Yoccoz, N.G. Stable isotope analysis: Modelling lipid normalization for muscle and eggs from arctic mammals and birds. Methods Ecol. Evol. 2011, 2, 66–76. [Google Scholar] [CrossRef]
- Javornik, J.; Hopkins, J.B.; Zavadlav, S.; Levanič, T.; Lojen, S.; Polak, T.; Jerina, K. Effects of ethanol storage and lipids on stable isotope values in a large mammalian omnivore. J. Mammal. 2019, 100, 150–157. [Google Scholar] [CrossRef]
- Caut, S.; Angulo, E.; Courchamp, F. Variation in discrimination factors (δ15N and δ13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 2009, 46, 443–453. [Google Scholar] [CrossRef]
- Cloyed, C.S.; Newsome, S.D.; Eason, P.K. Trophic discrimination factors and incorporation rates of carbon- and nitrogen-stable isotopes in adult green frogs, Lithobates clamitans. Physiol. Biochem. Zool. 2015, 88, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bélouard, N.; Petit, E.J.; Huteau, D.; Oger, A.; Paillisson, J.-M. Fins are relevant non-lethal surrogates for muscle to measure stable isotopes in amphibians. Knowl. Manag. Aquat. Ecosyst. 2019, 420, 2–8. [Google Scholar] [CrossRef]
- Martinez del Rio, C.; Wolf, N.; Carleton, S.A.; Gannes, L.Z. Isotopic ecology ten years after a call for more laboratory experiments. Biol. Rev. 2009, 84, 91–111. [Google Scholar] [CrossRef]
- Smith, R.L.; Beard, K.H.; Shiels, A.B. Different prey resources suggest little competition between non-native frogs and insectivorous birds despite isotopic niche overlap. Biol. Invasions 2017, 19, 1001–1013. [Google Scholar] [CrossRef] [Green Version]
- Krab, E.J.; Van Logtestijn, R.S.P.; Cornelissen, J.H.C.; Berg, M.P. Reservations about preservations: Storage methods affect delta C-13 signatures differently even in closely related soil fauna. Methods Ecol. Evol. 2012, 3, 138–144. [Google Scholar] [CrossRef]
- Barrow, L.M.; Bjorndal, K.A.; Reich, K.J. Effects of preservation method on stable carbon and nitrogen isotope values. Physiol. Biochem. Zool. 2008, 81, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Fleming, N.E.C.; Houghton, J.D.R.; Magill, C.L.; Harrod, C. Preservation methods alter stable isotope values in gelatinous zooplankton: Implications for interpreting trophic ecology. Mar. Biol. 2011, 158, 2141–2146. [Google Scholar] [CrossRef] [Green Version]
- Le Bourg, B.; Lepoint, G.; Michel, L.N. Effects of preservation methodology on stable isotope compositions of sea stars. Rapid Commun. Mass Spectrom. 2020, 34, e8589. [Google Scholar] [CrossRef] [PubMed]
- Bissattini, A.M.; Buono, V.; Vignoli, L. Disentangling the trophic interactions between American bullfrogs and native anurans: Complications resulting from post metamorphic ontogenetic niche shifts. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 270–281. [Google Scholar] [CrossRef]
- Jesus, F.M.; Pereira, M.R.; Rosa, C.S.; Moreira, M.Z.; Sperber, C.F. Preservation methods alter carbon and nitrogen stable isotope values in crickets (Orthoptera: Grylloidea). PLoS ONE 2015, 10, e0137650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carabel, S.; Verísimo, P.; Freire, J. Effects of preservatives on stable isotope analyses of four marine species. Estuar. Coast. Shelf. Sci. 2009, 82, 348–350. [Google Scholar] [CrossRef]
- Vanderklift, M.A.; Ponsard, S. Sources of variation in consumer-diet δ15N enrichment: A meta-analysis. Oecologia 2003, 136, 169–182. [Google Scholar] [CrossRef]
- Butler, O.M.; Rashti, M.R.; Chen, C. Influence of storage and drying methods on invertebrate elemental and isotopic measurements. Comm. Soil Sci. Plant Anal. 2018, 49, 1–7. [Google Scholar] [CrossRef]
- Bugoni, L.; Mcgill, R.A.R.; Furness, R.W. Effects of preservation methods on stable isotope signatures in bird tissues. Rapid Commun. Mass Spectrom. 2008, 22, 2457–2462. [Google Scholar] [CrossRef] [PubMed]
- Oczkowski, A.; Thornber, C.S.; Markham, E.E.; Rossi, R.; Ziegler, A.; Rinehart, S. Testing sample stability using four storage methods and the macroalgae Ulva and Gracilaria. Limnol. Oceanogr. Methods 2015, 13, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Fanelli, E.; Cartes, J.E.; Papiol, V.; Rumolo, P.; Sprovieri, M. Effects of preservation on the δ13C and δ15N values of deep sea macrofauna. J. Exp. Mar. Biol. Ecol. 2010, 395, 93–97. [Google Scholar] [CrossRef]
- Durante, L.M.; Sabadel, A.J.M.; Frew, R.D.; Ingram, T.; Wing, S.R. Effects of fixatives on stable isotopes of fish muscle tissue: Implications for trophic studies on preserved specimens. Ecol. Appl. 2020, 30, e02080. [Google Scholar] [CrossRef] [PubMed]
- Kiszka, J.; Lesage, V.; Ridoux, V. Effect of ethanol preservation on stable carbon and nitrogen isotope values in cetacean epidermis: Implication for using archived biopsy samples. Mar. Mamm. Sci. 2014, 30, 788–795. [Google Scholar] [CrossRef]
- Gibbons, J.W.; Winne, C.T.; Scott, D.E.; Willson, J.D.; Glaudas, X.; Andrews, K.M.; Todd, B.D.; Fedewa, L.A.; Wilkinson, L.; Tsaliagos, R.N.; et al. Remarkable amphibian biomass and abundance in an isolated wetland: Implications for wetland conservation. Conserv. Biol. 2006, 20, 1457. [Google Scholar] [CrossRef]
- Wells, K.D. The Ecology and Behavior of Amphibians; The University of Chicago Press: Chicago, IL, USA; London, UK, 2007. [Google Scholar]
- Glos, J.; Ruthsatz, K.; Schroder, D.; Riemann, J.C. Food source determines stable isotope discrimination factor dN and dC in tadpoles. Amphib. Reptil. 2020, 41, 501–507. [Google Scholar] [CrossRef]
- Reshetnikov, A.N.; Korobushkin, D.I.; Gongalskiy, K.B.; Korotkevich, A.Y.; Selskaya, A.N.; Kotov, A.A.; Tiunov, A.V. Trophic positions and niche segregation of two anuran species in the ecosystem of a forest lake. Hydrobiologia 2021. [Google Scholar] [CrossRef]
- Smirnov, N.N. Lake Glubokoe (Moscow region, Eastern Europe), general characteristics. Hydrobiologia 1986, 141, 1–6. [Google Scholar] [CrossRef]
- Annenskaya, G.N.; Zhichkova, V.K.; Kalinina, V.P.; Mamay, I.I.; Nizovtsev, V.A.; Khrustaleva, M.A.; Tseselchuk, Y.N. Landscapes of Moscow Region and Their Current Condition; SGU: Smolensk, Russia, 1997; 296p. (In Russian) [Google Scholar]
- Miller, J.; Haden, P. Statistical Analysis with the General Linear Model; Department of Psychology, University of Otago: Dunedin, New Zealand, 2006; Available online: https://www.otago.ac.nz/psychology/otago039309.pdf (accessed on 1 July 2021).
- Trakimas, G.; Jardine, T.D.; Barisevičiūtė, R.; Garbaras, A.; Skipitytė, R.; Remeikis, V. Ontogenetic dietary shifts in European common frog (Rana temporaria) revealed by stable isotopes. Hydrobiologia 2011, 675, 87–95. [Google Scholar] [CrossRef]
- Kadye, W.T.; Redelinghuys, S.; Parnell, P.S.; Booth, A.J. Exploring source differences on diet-tissue discrimination factors in the analysis of stable isotope mixing models. Sci. Rep. 2020, 10, 15816. [Google Scholar] [CrossRef] [PubMed]
- Beebee, T.J.C.; Wong, L.-C. Leucine uptake by enterobacterial and algal members of larval anuran gut flora. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1992, 101B, 527–530. [Google Scholar] [CrossRef]
- Pryor, G.S.; Bjorndal, K.A. Effects of the nematode Gyrinicola batrachiensis on development, gut morphology, and fermentation in bullfrog tadpoles (Rana catesbeiana): A novel mutualism. J. Exp. Zool. 2005, 303A, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Pryor, G.S.; Bjorndal, K.A. Symbiotic fermentation, digestal passage, and gastrointestinal morphology in bullfrog tadpoles (Rana catesbeiana). Physiol. Biochem. Zool. 2005, 78, 201–215. [Google Scholar] [CrossRef]
- Vences, M.; Lyra, M.L.; Kueneman, J.G.; Bletz, M.C.; Archer, H.M.; Canitz, J.; Handreck, S.; Randrianiaina, R.-D.; Struck, U.; Bhuju, S.; et al. Gut bacterial communities across tadpole ecomorphs in two diverse tropical anuran faunas. Sci. Nat. 2016, 103, 25. [Google Scholar] [CrossRef] [Green Version]
- Korotkevich, A.Y.; Kuznetsova, N.A.; Tiunov, A.V. Changes in Carbon and Nitrogen Isotope Ratios (13C/12C and 15N/14N) in Springtails during Long-term Storage of Soil Samples. Russ. J. Ecol. 2016, 47, 572–574. [Google Scholar] [CrossRef]
- Terentiev, P.V. Frog; Sovetskaya Nauka: Moscow, Russia, 1950; 345p. [Google Scholar]
- Caut, S.; Angulo, E.; Díaz-Paniagua, C.; Gomez-Mestre, I. Plastic changes in tadpole trophic ecology revealed by stable isotope analysis. Oecologia 2013, 173, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Arribas, R.; Díaz-Paniagua, C.; Caut, S.; Gomez-Mestre, I. Stable isotopes reveal trophic partitioning and trophic plasticity of a larval amphibian guild. PLoS ONE 2015, 10, e0130897. [Google Scholar] [CrossRef] [Green Version]
- Vassilieva, A.B.; Sinev, A.Y.; Tiunov, A.V. Trophic segregation of anuran larvae in two temporary tropical ponds in southern Vietnam. Herpetol. J. 2017, 27, 217–229. [Google Scholar]
- San Sebastián, O.; Navarro, J.; Llorente, G.A.; Richter-Boix, Á. Trophic strategies of a non-native and a native amphibian species in shared ponds. PLoS ONE 2015, 10, e0130549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmin, S.L. The Amphibians of the Former Soviet Union; Pensoft: Moscow, Russia; Sofia, Bulgaria, 2013; 383p. [Google Scholar]
- Houlahan, J.E.; Findlay, C.S.; Schmidt, B.R.; Meyer, A.H.; Sergius, L.; Kuzmin, S.L. Quantitative evidence for global amphibian population declines. Nature 2000, 404, 752–755. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Factors | δ13C | δ15N | C/N | |||
|---|---|---|---|---|---|---|
| SS | p | SS | p | SS | p | |
| Tadpoles | ||||||
| Storage method | 2.42 | 0.037 | n.s. | 3.28 | <0.0001 | |
| Tissue | 10.3 | 0.0044 | 11.78 | <0.0001 | 9.68 | <0.0001 |
| Storage method × Tissue | n.s. | n.s. | 0.80 | 0.0084 | ||
| Error | 28.05 | 12.8 | 2.16 | |||
| Model R2 | 0.31 | 0.479 | 0.864 | |||
| Model p | 0.0022 | <0.0001 | <0.0001 | |||
| Juveniles | ||||||
| Storage method | 6.99 | 0.0004 | n.s. | 3.2 | <0.0001 | |
| Tissue | 7.48 | 0.016 | 6.3 | 0.0001 | 2.67 | <0.0001 |
| Storage method × Tissue | n.s. | n.s. | 1.37 | <0.0001 | ||
| Error | 22.5 | 1.01 | ||||
| Model R2 | 0.386 | 9.2 | 0.878 | |||
| Model p | 0.00057 | 0.406 | 0.0001 | <0.0001 | ||
| Factor | Freezing | Ethanol | ||||||
|---|---|---|---|---|---|---|---|---|
| δ13C | δ15N | δ13C | δ15N | |||||
| SS | p | SS | p | SS | p | SS | p | |
| Stage | 9.26 | 0.000015 | 2.165 | 0.000293 | 18.49 | 0.000001 | 1.292 | 0.044 |
| Tissue | 8.96 | 0.000807 | 4.378 | 0.000087 | 6.79 | 0.030980 | n.s. | |
| Stage × Tissue | 4.68 | 0.025843 | 3.258 | 0.000784 | n.s. | n.s. | ||
| Error | 13.84 | 5.011 | 25.44 | 14.54 | ||||
| Model R2 | 0.61 | 0.66 | 0.498 | 0.082 | ||||
| Model p | 0.00002 | 0.000002 | 0.000008 | 0.044 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reshetnikov, A.N.; Korobushkin, D.I. Intra-Body Variations of Stable Isotope Ratios (δ13C, δ15N) and Influence of Storage Methods in Aquatic and Post-Aquatic Stages of the Common Toad, Bufo bufo. Water 2021, 13, 2544. https://doi.org/10.3390/w13182544
Reshetnikov AN, Korobushkin DI. Intra-Body Variations of Stable Isotope Ratios (δ13C, δ15N) and Influence of Storage Methods in Aquatic and Post-Aquatic Stages of the Common Toad, Bufo bufo. Water. 2021; 13(18):2544. https://doi.org/10.3390/w13182544
Chicago/Turabian StyleReshetnikov, Andrey N., and Daniil I. Korobushkin. 2021. "Intra-Body Variations of Stable Isotope Ratios (δ13C, δ15N) and Influence of Storage Methods in Aquatic and Post-Aquatic Stages of the Common Toad, Bufo bufo" Water 13, no. 18: 2544. https://doi.org/10.3390/w13182544
APA StyleReshetnikov, A. N., & Korobushkin, D. I. (2021). Intra-Body Variations of Stable Isotope Ratios (δ13C, δ15N) and Influence of Storage Methods in Aquatic and Post-Aquatic Stages of the Common Toad, Bufo bufo. Water, 13(18), 2544. https://doi.org/10.3390/w13182544