
Modelling, Characterizing, and Monitoring Boreal Forest Wetland Bird Habitat with RADARSAT-2 and Landsat-8 Data

 ,
,  , ,
, , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
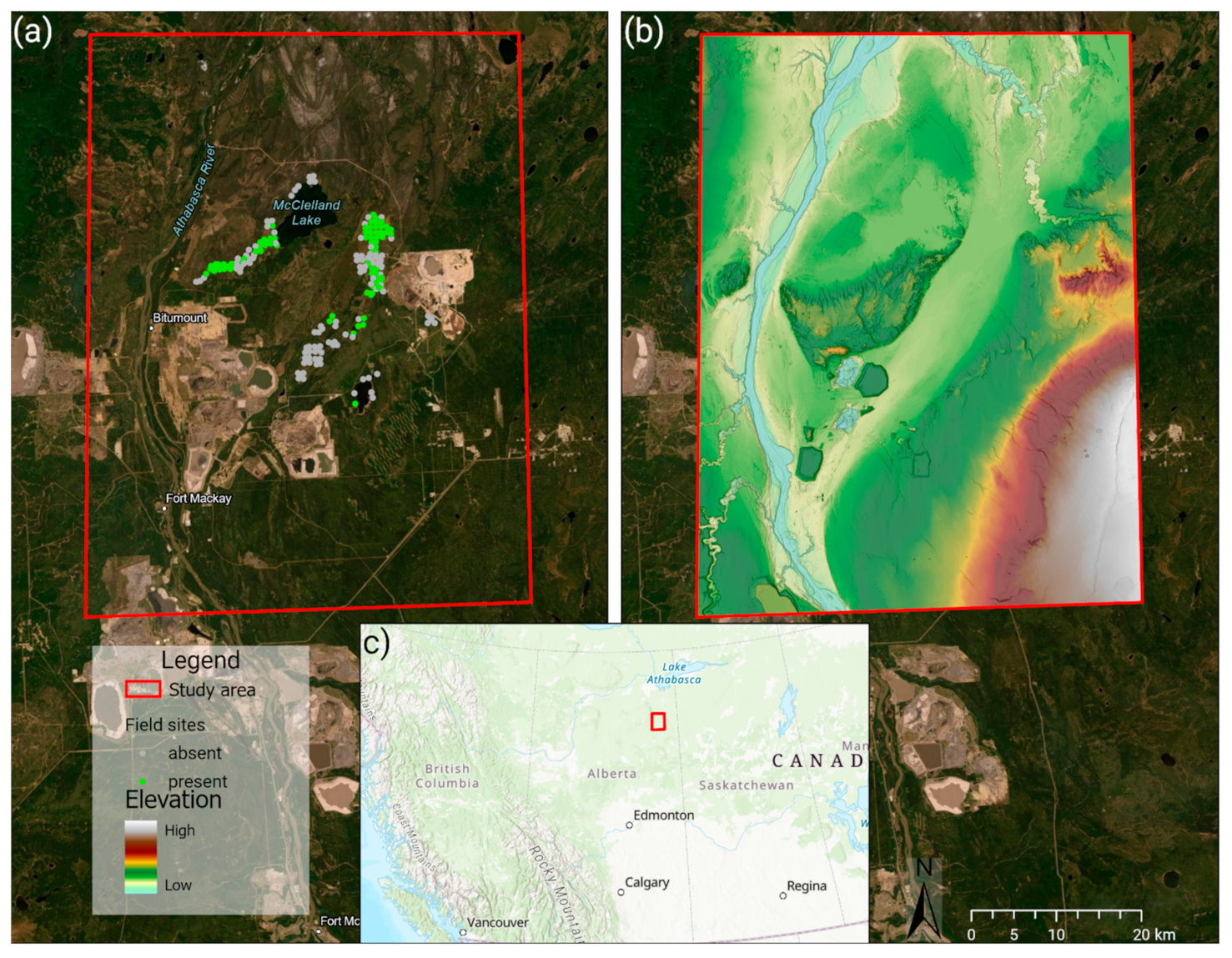
2.1. Study Area
2.2. Yellow Rail Abundance Data
2.3. Earth Observation Data
2.4. Habitat Modeling, Prediction, and Monitoring
3. Results
4. Discussion
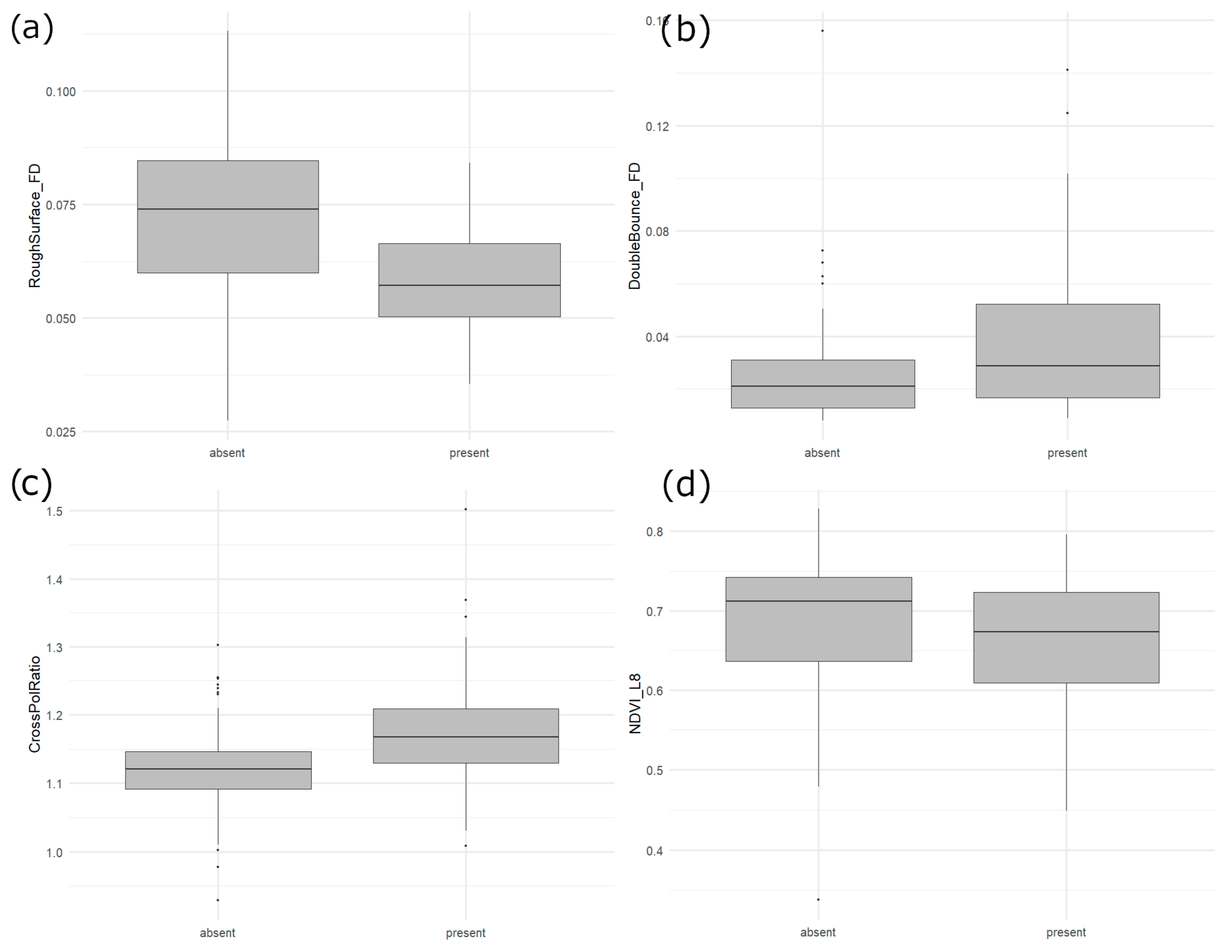
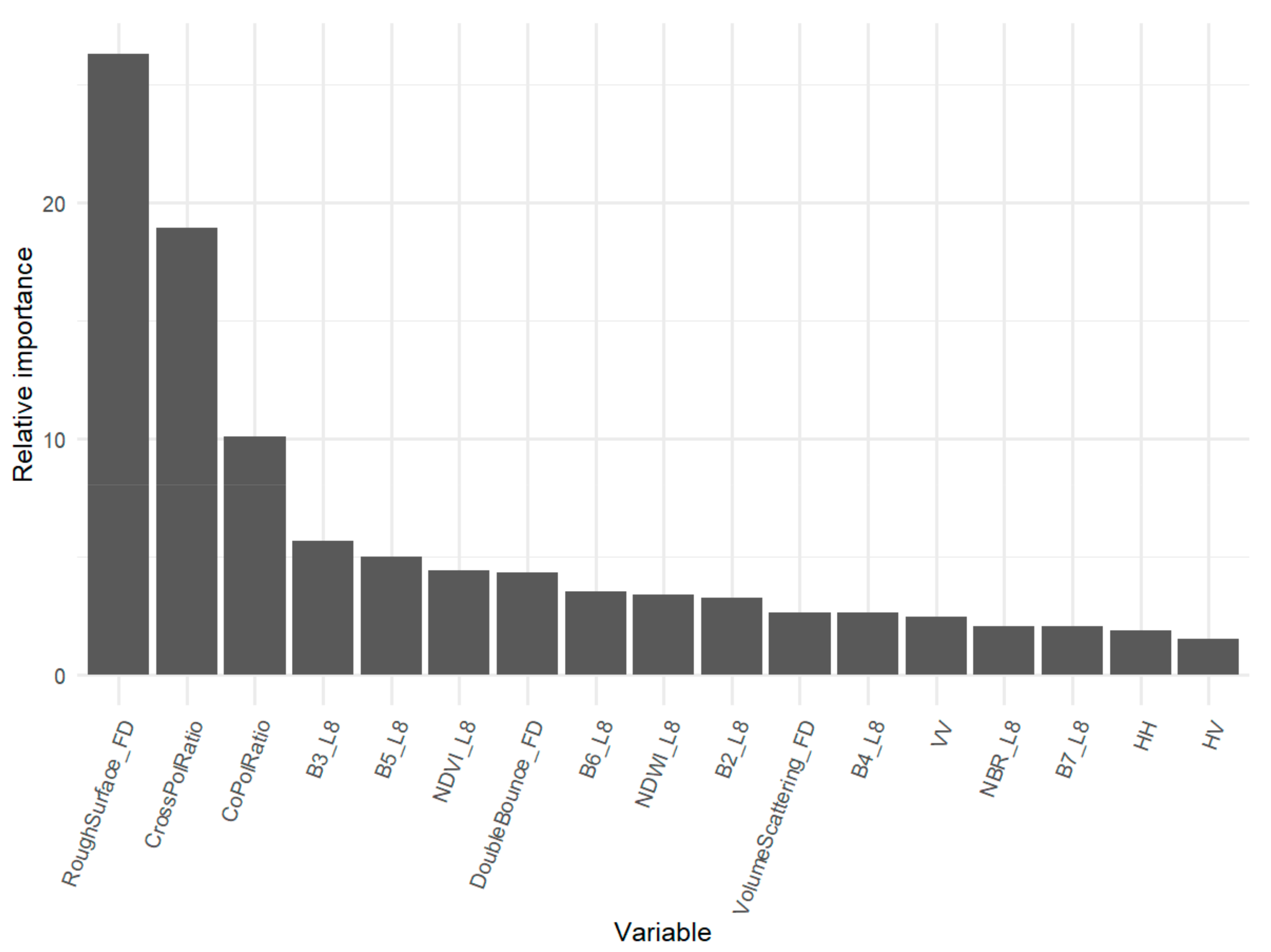
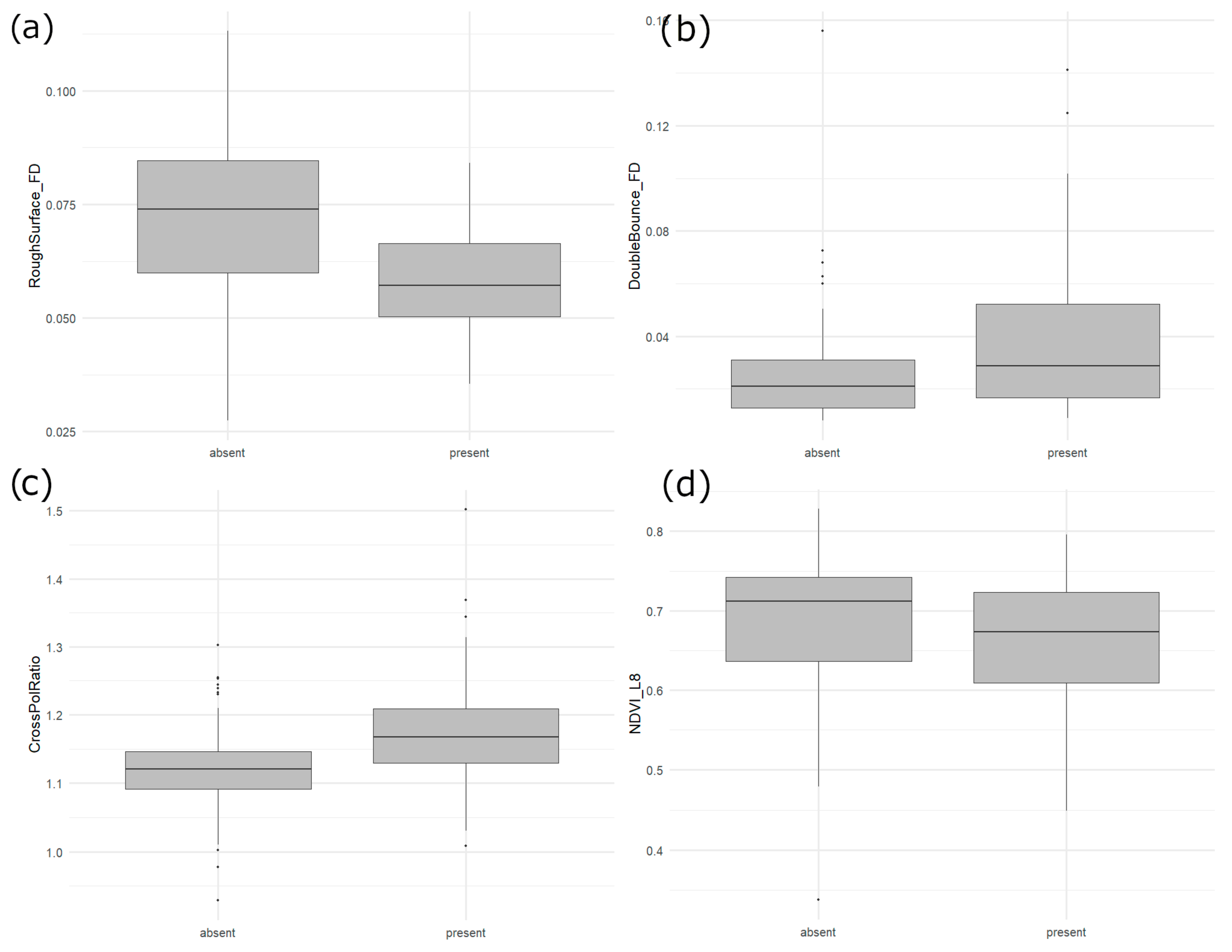
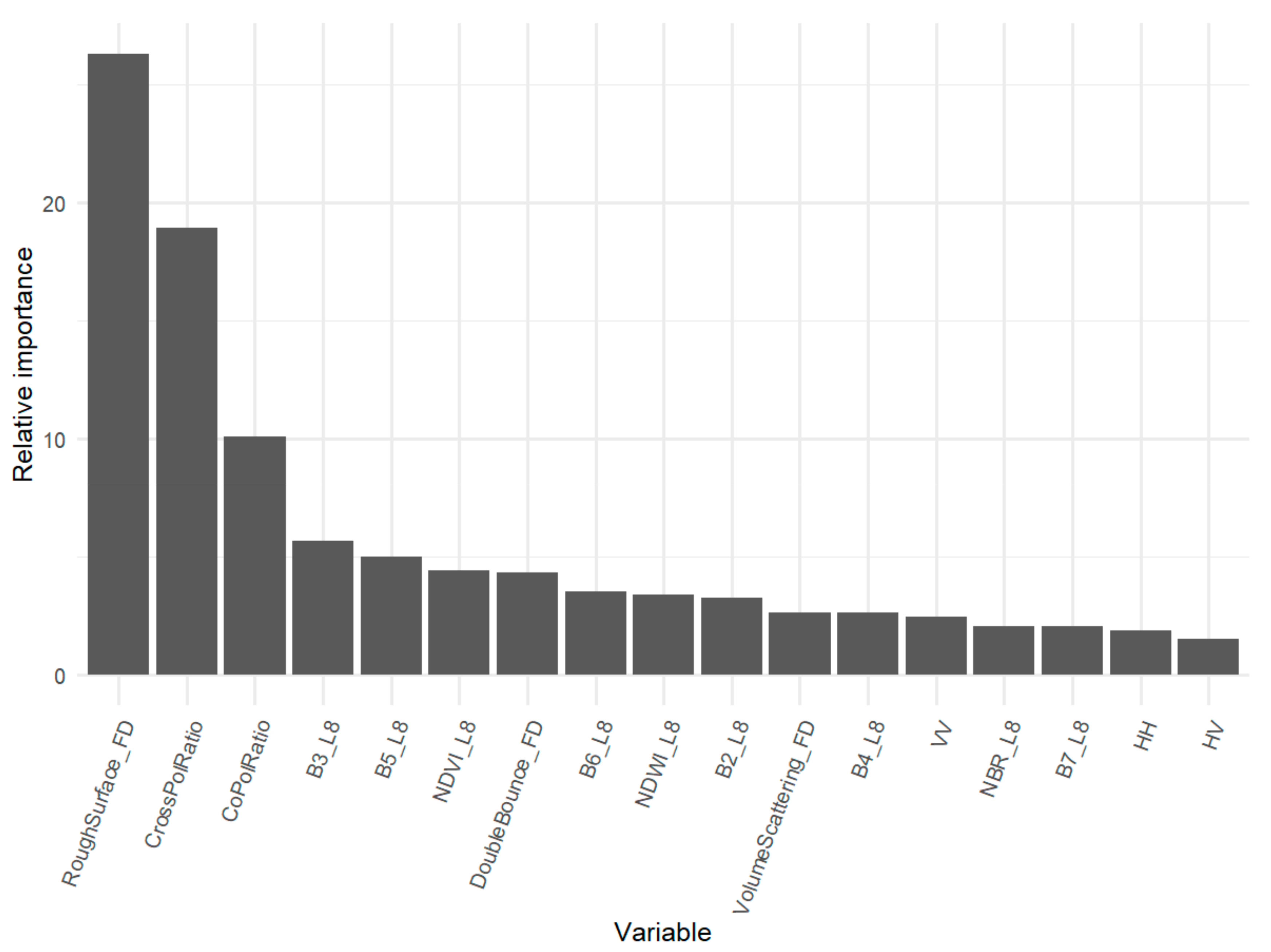
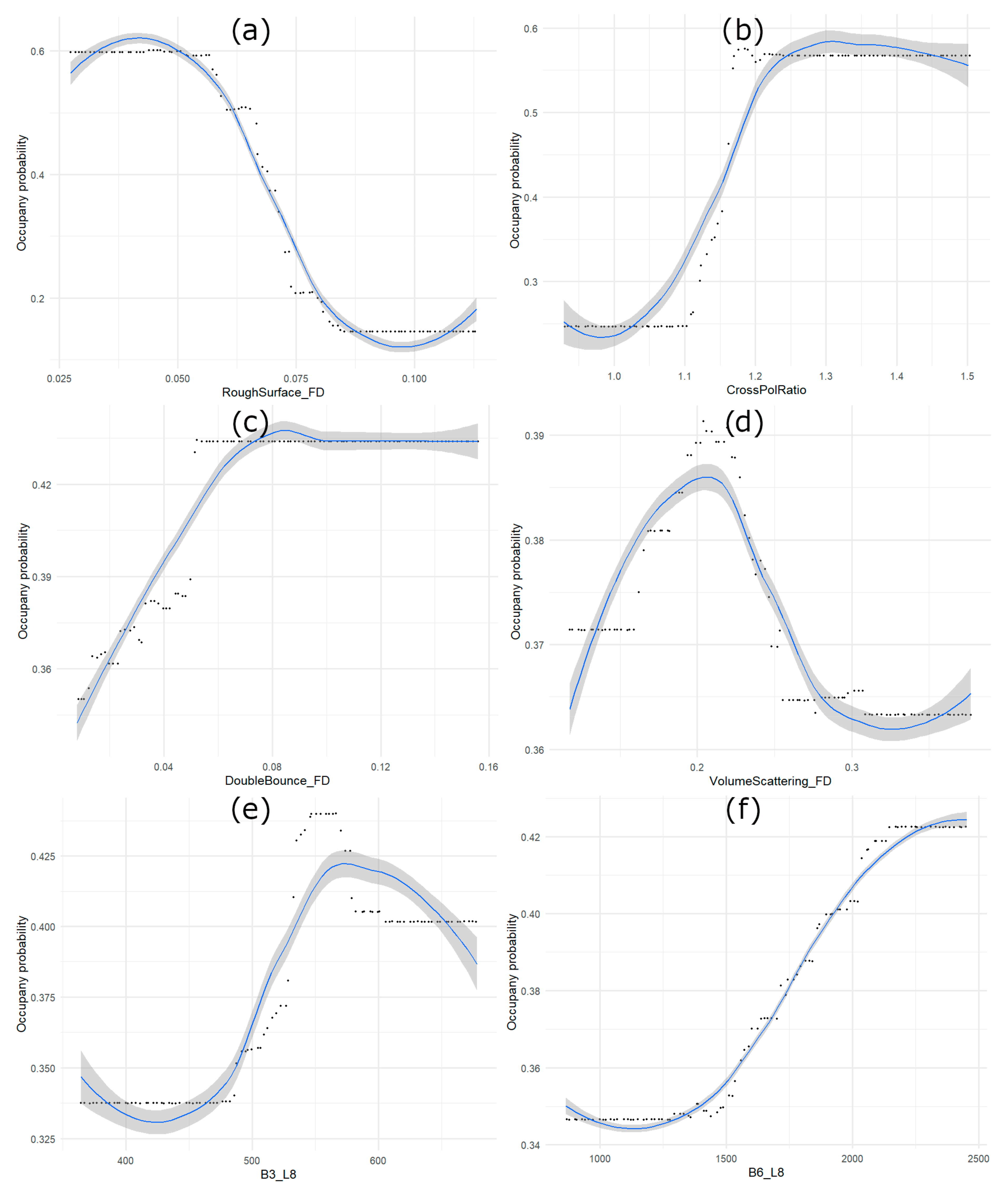
4.1. Relative Importance of EO Inputs
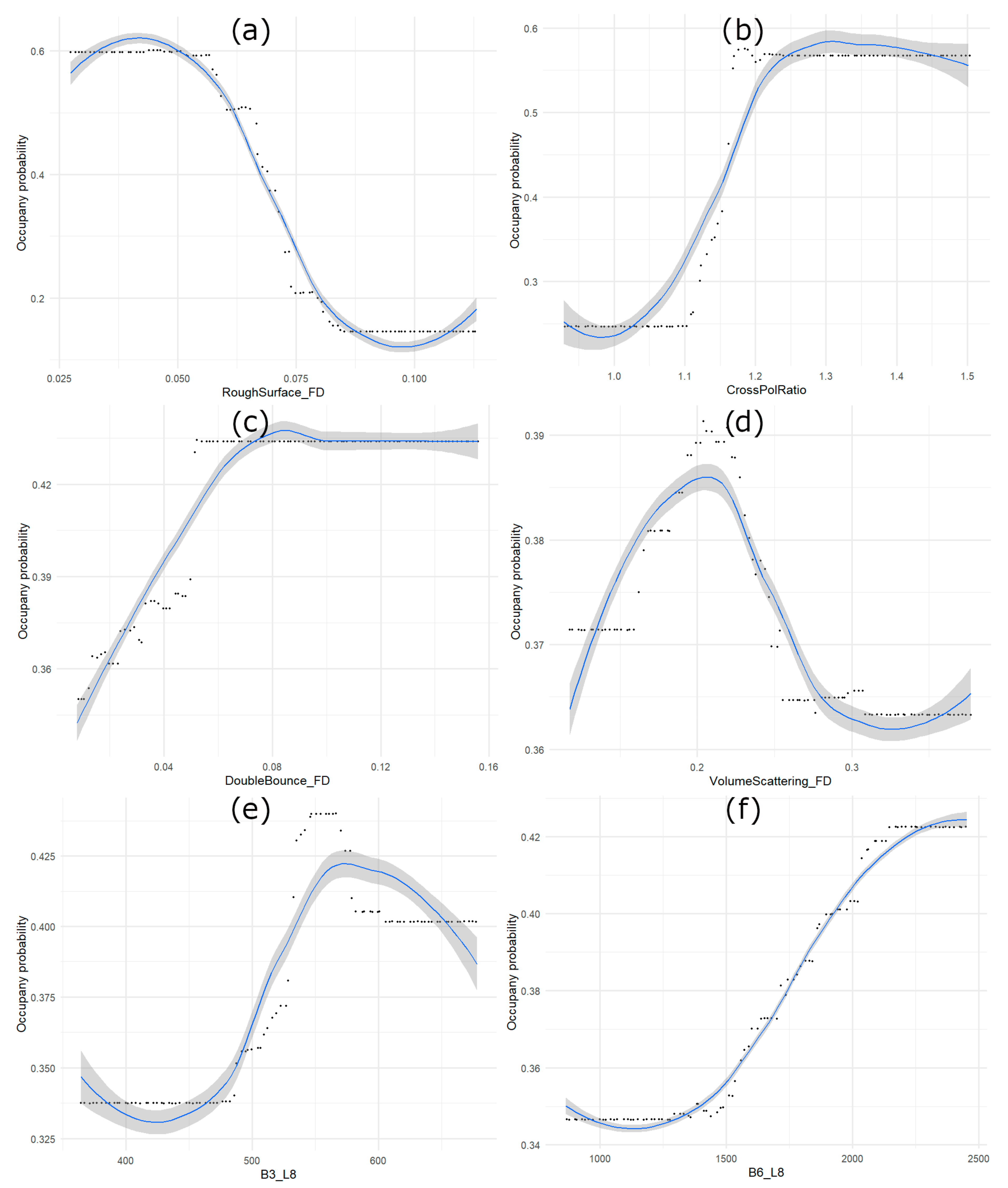
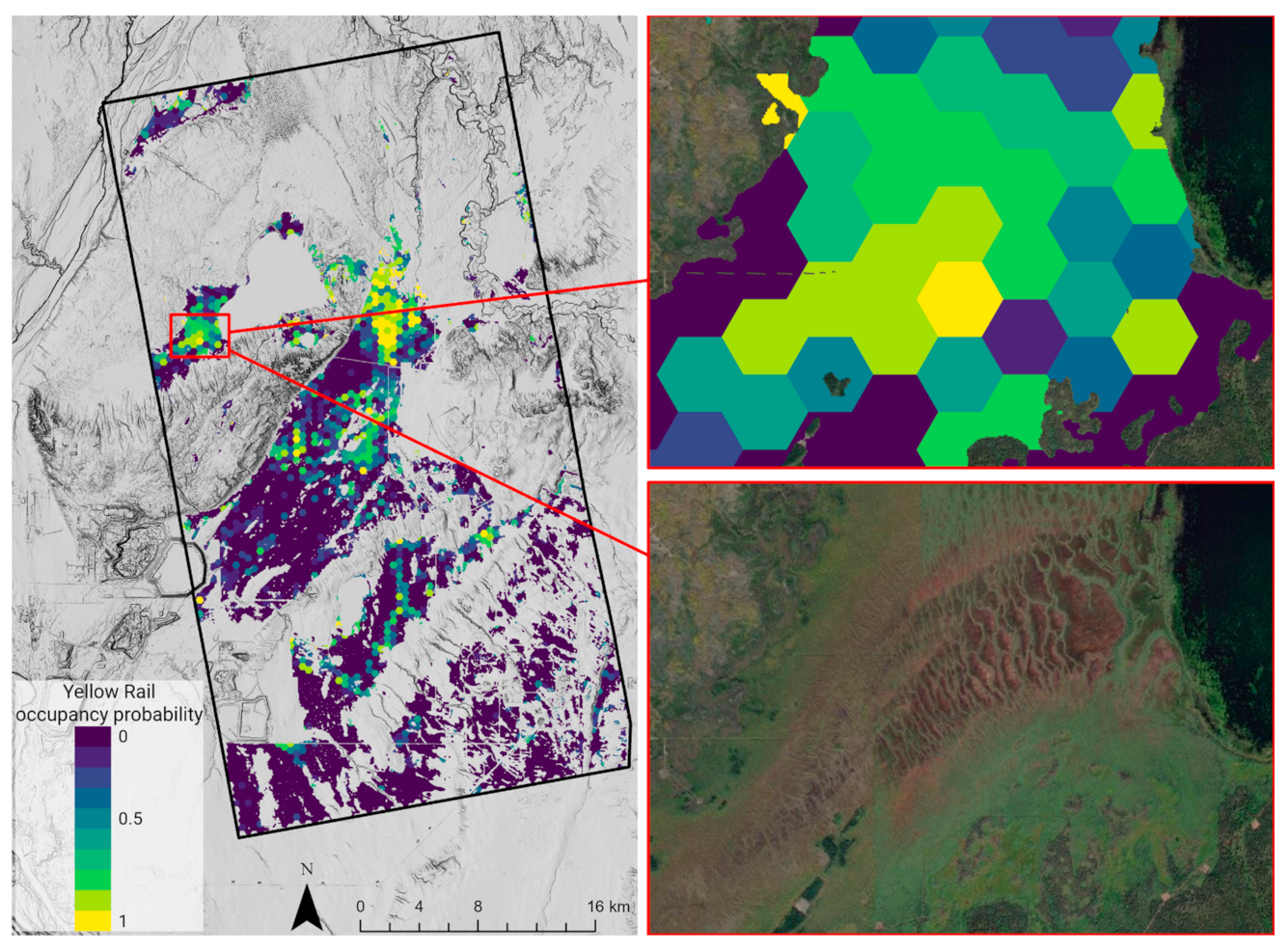
4.2. Yellow Rail Habitat
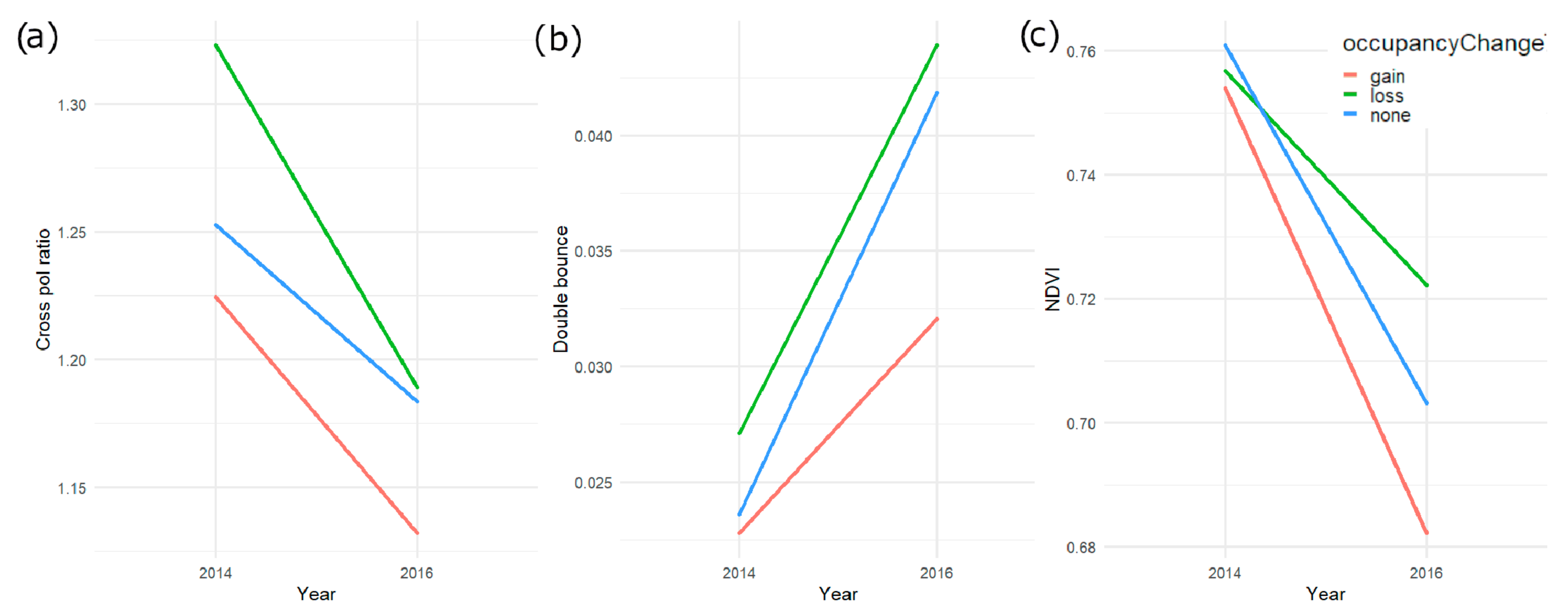
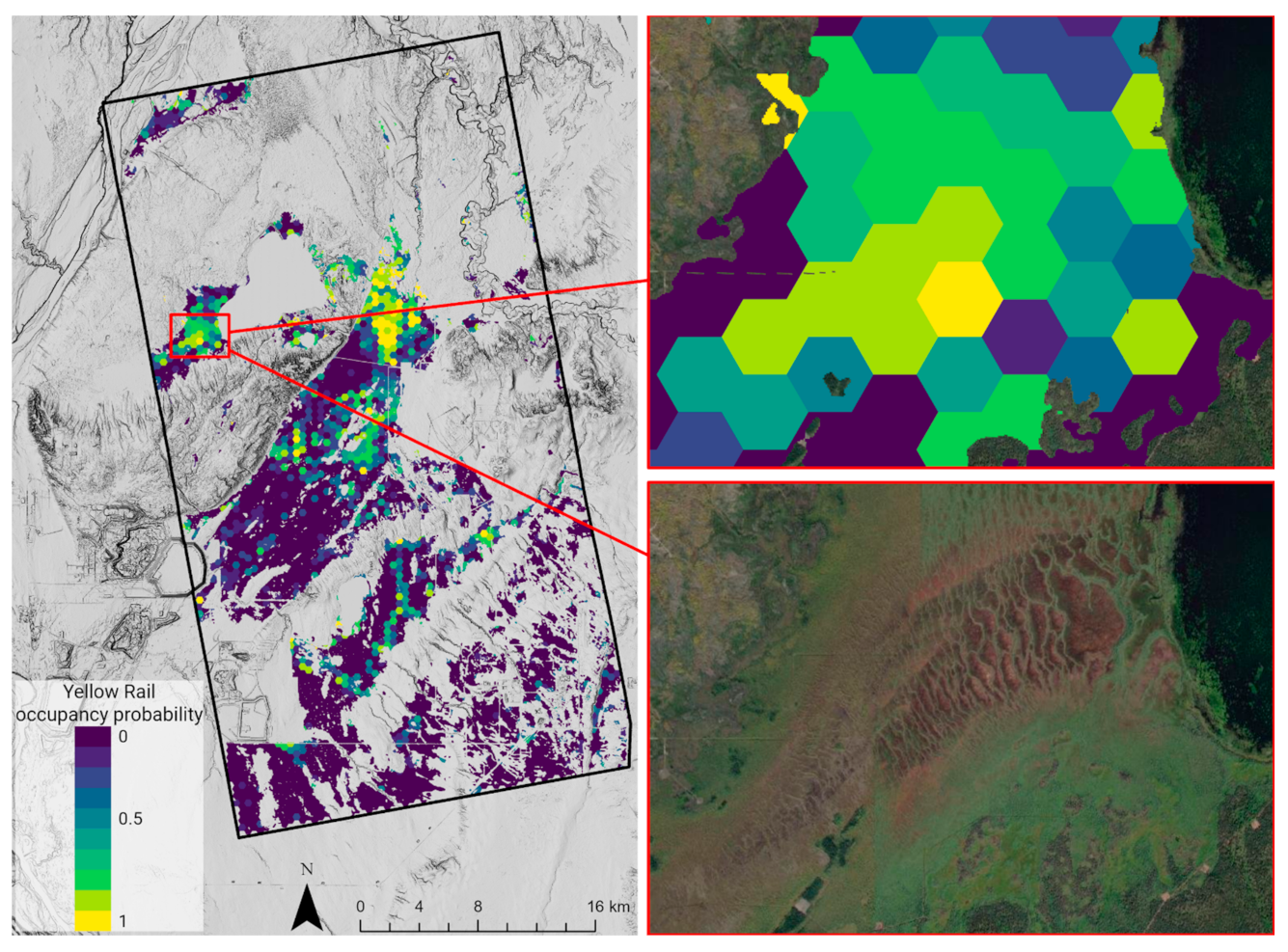
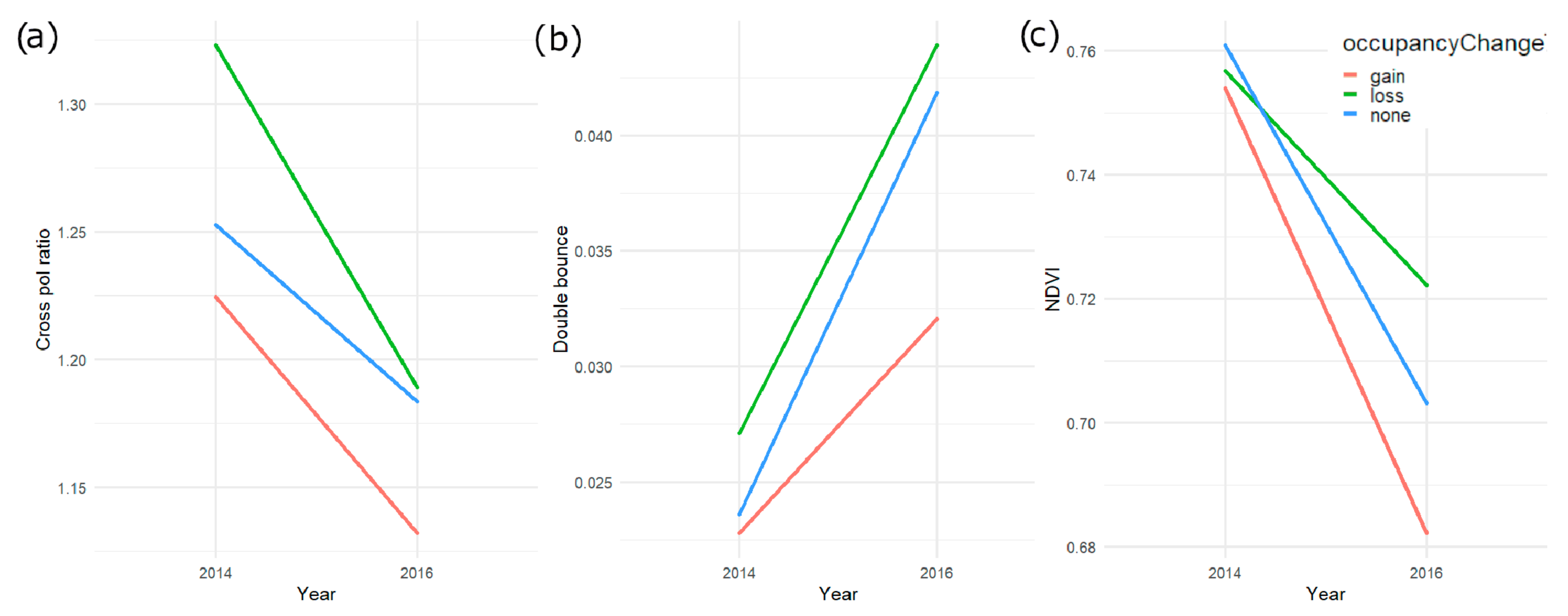
4.3. Monitoring Trends
4.4. Habitat Prediction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will remote sensing shape the next generation of species distribution models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Brisco, B. Mapping and monitoring surface water and wetlands with synthetic aperture radar. Remote Sens. Wetl. Appl. Adv. 2015, 119–136. [Google Scholar] [CrossRef]
- White, L.; Brisco, B.; Dabboor, M.; Schmitt, A.; Pratt, A. A collection of SAR methodologies for monitoring wetlands. Remote Sens. 2015, 7, 7615–7645. [Google Scholar] [CrossRef] [Green Version]
- Hird, J.N.; DeLancey, E.R.; McDermid, G.J.; Kariyeva, J. Google earth engine, open-access satellite data, and machine learning in support of large-area probabilistic wetland mapping. Remote Sens. 2017, 9, 1315. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, J.; Brisco, B.; Chasmer, L.; Devito, K.; Cobbaert, D.; Hopkinson, C. SAR and LiDAR temporal data fusion approaches to boreal wetland ecosystem monitoring. Remote Sens. 2019, 11, 161. [Google Scholar] [CrossRef] [Green Version]
- DeLancey, E.R.; Simms, J.F.; Mahdianpari, M.; Brisco, B.; Mahoney, C.; Kariyeva, J. Comparing deep learning and shallow learning for large-scale wetland classification in Alberta, Canada. Remote Sens. 2020, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.V. Boreal Birds of North America: A Hemispheric View of Their Conservation Links and Significance; University of California Press: Berkley, CA, USA, 2011. [Google Scholar]
- Wetlands International. Waterbird Population Estimates. Available online: http://wpe.wetlands.org/ (accessed on 10 January 2019).
- Leston, L.B.T. Yellow Rail (Coturnicops Noveboracensis), version 2.0; Cornell Lab of Ornithology: Ithaca, NY, USA, 2015. [Google Scholar]
- Robert, M.; Laporte, P. Numbers and movements of yellow rails along the St. Lawrence River, Quebec. Condor 1999, 101, 667–671. [Google Scholar] [CrossRef]
- Amani, M.; Mahdavi, S.; Afshar, M.; Brisco, B.; Huang, W.; Mohammad Javad Mirzadeh, S.; White, L.; Banks, S.; Montgomery, J.; Hopkinson, C. Canadian wetland inventory using google earth engine: The first map and preliminary results. Remote Sens. 2019, 11, 842. [Google Scholar] [CrossRef] [Green Version]
- Pouliot, D.; Latifovic, R.; Pasher, J.; Duffe, J. Assessment of convolution neural networks for wetland mapping with landsat in the central Canadian boreal forest region. Remote Sens. 2019, 11, 772. [Google Scholar] [CrossRef] [Green Version]
- Mao, D.; Wang, Z.; Du, B.; Li, L.; Tian, Y.; Jia, M.; Zeng, Y.; Song, K.; Jiang, M.; Wang, Y. National wetland mapping in China: A new product resulting from object-based and hierarchical classification of Landsat 8 OLI images. ISPRS J. Photogramm. Remote Sens. 2020, 164, 11–25. [Google Scholar] [CrossRef]
- Millard, K.; Richardson, M. Wetland mapping with LiDAR derivatives, SAR polarimetric decompositions, and LiDAR–SAR fusion using a random forest classifier. Can. J. Remote Sens. 2013, 39, 290–307. [Google Scholar] [CrossRef]
- McLeod, L.J.; DeLancey, E.R.; Bayne, E.M. Spatially Explicit Abundance Modelling of a Highly Specialized Wetland Bird using Sentinel-1 and Sentinel-2. Candian J. Remote Sens. 2021, in press. [Google Scholar]
- Schmitt, A.; Brisco, B. Wetland monitoring using the curvelet-based change detection method on polarimetric SAR imagery. Water 2013, 5, 1036–1051. [Google Scholar] [CrossRef]
- Brisco, B.; Schmitt, A.; Murnaghan, K.; Kaya, S.; Roth, A. SAR polarimetric change detection for flooded vegetation. Int. J. Digit. Earth 2013, 6, 103–114. [Google Scholar] [CrossRef]
- Freeman, A.; Durden, S.L. A three-component scattering model for polarimetric SAR data. IEEE Trans. Geosci. Remote Sens. 1998, 36, 963–973. [Google Scholar] [CrossRef] [Green Version]
- Wellmann, T.; Lausch, A.; Scheuer, S.; Haase, D. Earth observation based indication for avian species distribution models using the spectral trait concept and machine learning in an urban setting. Ecol. Indic. 2020, 111, 106029. [Google Scholar] [CrossRef]
- Bino, G.; Levin, N.; Darawshi, S.; Van Der Hal, N.; Reich-Solomon, A.; Kark, S. Accurate prediction of bird species richness patterns in an urban environment using Landsat-derived NDVI and spectral unmixing. Int. J. Remote Sens. 2008, 29, 3675–3700. [Google Scholar] [CrossRef]
- St-Louis, V.; Pidgeon, A.M.; Kuemmerle, T.; Sonnenschein, R.; Radeloff, V.C.; Clayton, M.K.; Locke, B.A.; Bash, D.; Hostert, P. Modelling avian biodiversity using raw, unclassified satellite imagery. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buermann, W.; Saatchi, S.; Smith, T.B.; Zutta, B.R.; Chaves, J.A.; Milá, B.; Graham, C.H. Predicting species distributions across the Amazonian and Andean regions using remote sensing data. J. Biogeogr. 2008, 35, 1160–1176. [Google Scholar] [CrossRef]
- Farrell, S.; Collier, B.; Skow, K.; Long, A.; Campomizzi, A.; Morrison, M.; Hays, K.; Wilkins, R. Using LiDAR-derived vegetation metrics for high-resolution, species distribution models for conservation planning. Ecosphere 2013, 4, 1–18. [Google Scholar] [CrossRef]
- Chen, F.; Guo, H.; Ishwaran, N.; Zhou, W.; Yang, R.; Jing, L.; Chen, F.; Zeng, H. Synthetic aperture radar (SAR) interferometry for assessing Wenchuan earthquake (2008) deforestation in the Sichuan giant panda site. Remote Sens. 2014, 6, 6283–6299. [Google Scholar] [CrossRef] [Green Version]
- Evans, T.L. Habitat Mapping of the Brazilian Pantanal Using Synthetic Aperture Radar Imagery and Object Based Image Analysis; University of Victoria: Victoria, BC, Canada, 2013. [Google Scholar]
- Battaglia, M.J.; Banks, S.; Behnamian, A.; Bourgeau-Chavez, L.; Brisco, B.; Corcoran, J.; Chen, Z.; Huberty, B.; Klassen, J.; Knight, J. Multi-source eo for dynamic wetland mapping and monitoring in the great lakes basin. Remote Sens. 2021, 13, 599. [Google Scholar] [CrossRef]
- Hedley, R.; McLeod, L.; Yip, D.; Farr, D.; Knaga, P.; Drake, K.; Bayne, E. Modeling the occurrence of the Yellow Rail (Coturnicops noveboracensis) in the context of ongoing resource development in the oil sands region of Alberta. Avian Conserv. Ecol. 2020, 15, 10. [Google Scholar] [CrossRef]
- Natural Regions Committee. Natural Regions and Subregions of Alberta. Compiled by DJ Downing and WW Pettapiece. Government of Alberta; Government of Alberta Pub: Edmonton, AB, Canada, 2006. [Google Scholar]
- Yip, D.; Leston, L.; Bayne, E.; Sólymos, P.; Grover, A. Experimentally derived detection distances from audio recordings and human observers enable integrated analysis of point count data. Avian Conserv. Ecol. 2017, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Brotons, L.; Araújo, M.B.; Lavorel, S. Effects of restricting environmental range of data to project current and future species distributions. Ecography 2004, 27, 165–172. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Bailey, L.L.; Hines, J.E.; Nichols, J.D. An integrated model of habitat and species occurrence dynamics. Methods Ecol. Evol. 2011, 2, 612–622. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Satellite | Date | Resolution (m) | Data Type |
|---|---|---|---|
| RADARSAT-2 | 06 Aug 2014 | 8 | Fine Quad-Pol |
| RADARSAT-2 | 28 June 2016 | 8 | Fine Quad-Pol |
| Landsat-8 | 02 Aug 2014 | 30 | Surface reflectance |
| Landsat-8 | 29 June 2016 | 30 | Surface reflectance |
| Variable | Satellite | Description |
|---|---|---|
| HH | RADARSAT-2 | Horizontal-Horizontal backscatter |
| HV | RADARSAT-2 | Horizontal-Vertical backscatter |
| VV | RADARSAT-2 | Vertical-Vertical backscatter |
| CrossPolRatio | RADARSAT-2 | HH/VV |
| CoPolRatio | RADARSAT-2 | HH/HV |
| DoubleBounce_FD | RADARSAT-2 | Double bounce component of the Freeman–Durden decomposition |
| RoughSurface_FD | RADARSAT-2 | Rough surface component of the Freeman–Durden decomposition |
| VolumeScattering_FD | RADARSAT-2 | Volume scattering component of the Freeman–Durden decomposition |
| B2–7_ | Landsat-8 | Bands 2–7 from Landsat 8 |
| NBR_ | Landsat-8 | |
| NDVI_ | Landsat-8 | |
| NDWI_ | Landsat-8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeLancey, E.R.; Brisco, B.; McLeod, L.J.T.; Hedley, R.; Bayne, E.M.; Murnaghan, K.; Gregory, F.; Kariyeva, J. Modelling, Characterizing, and Monitoring Boreal Forest Wetland Bird Habitat with RADARSAT-2 and Landsat-8 Data. Water 2021, 13, 2327. https://doi.org/10.3390/w13172327
DeLancey ER, Brisco B, McLeod LJT, Hedley R, Bayne EM, Murnaghan K, Gregory F, Kariyeva J. Modelling, Characterizing, and Monitoring Boreal Forest Wetland Bird Habitat with RADARSAT-2 and Landsat-8 Data. Water. 2021; 13(17):2327. https://doi.org/10.3390/w13172327
Chicago/Turabian StyleDeLancey, Evan R., Brian Brisco, Logan J. T. McLeod, Richard Hedley, Erin M. Bayne, Kevin Murnaghan, Fiona Gregory, and Jahan Kariyeva. 2021. "Modelling, Characterizing, and Monitoring Boreal Forest Wetland Bird Habitat with RADARSAT-2 and Landsat-8 Data" Water 13, no. 17: 2327. https://doi.org/10.3390/w13172327
APA StyleDeLancey, E. R., Brisco, B., McLeod, L. J. T., Hedley, R., Bayne, E. M., Murnaghan, K., Gregory, F., & Kariyeva, J. (2021). Modelling, Characterizing, and Monitoring Boreal Forest Wetland Bird Habitat with RADARSAT-2 and Landsat-8 Data. Water, 13(17), 2327. https://doi.org/10.3390/w13172327