
Benthic Nutrient Fluxes across Subtidal and Intertidal Habitats in Breton Sound in Response to River-Pulses of a Diversion in Mississippi River Delta

Abstract
:1. Introduction
2. Materials and Methods
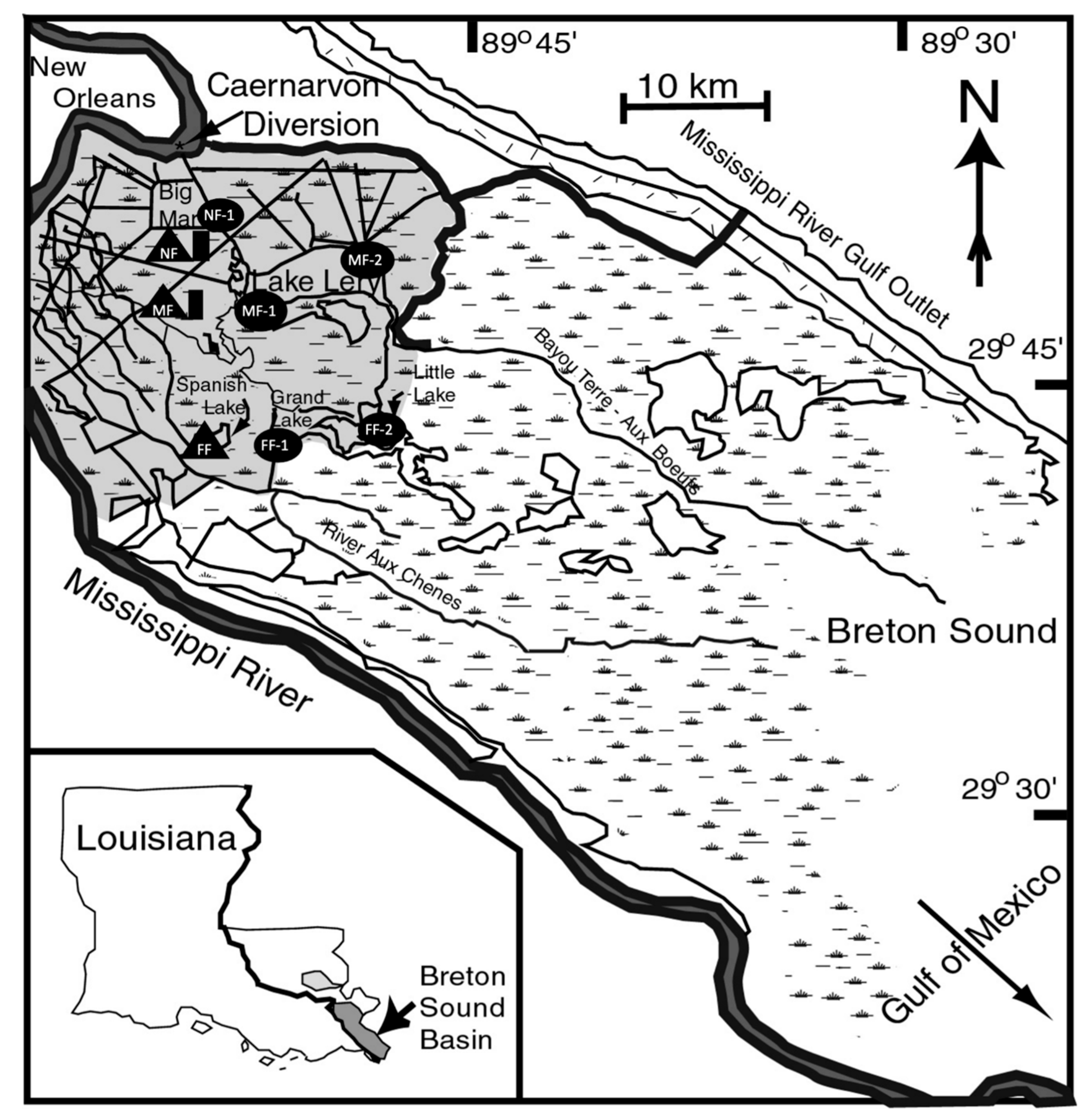
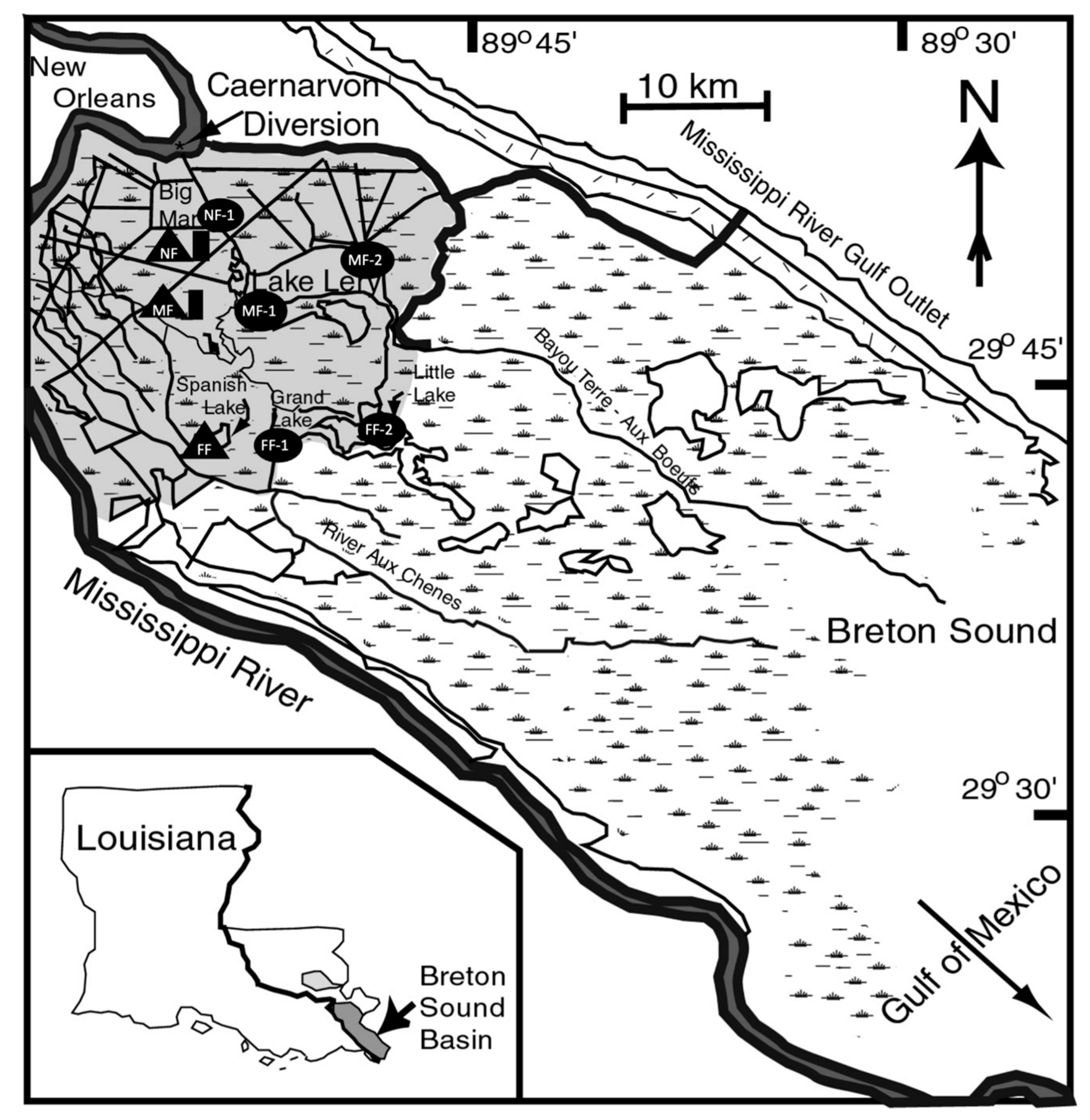
2.1. Study Site
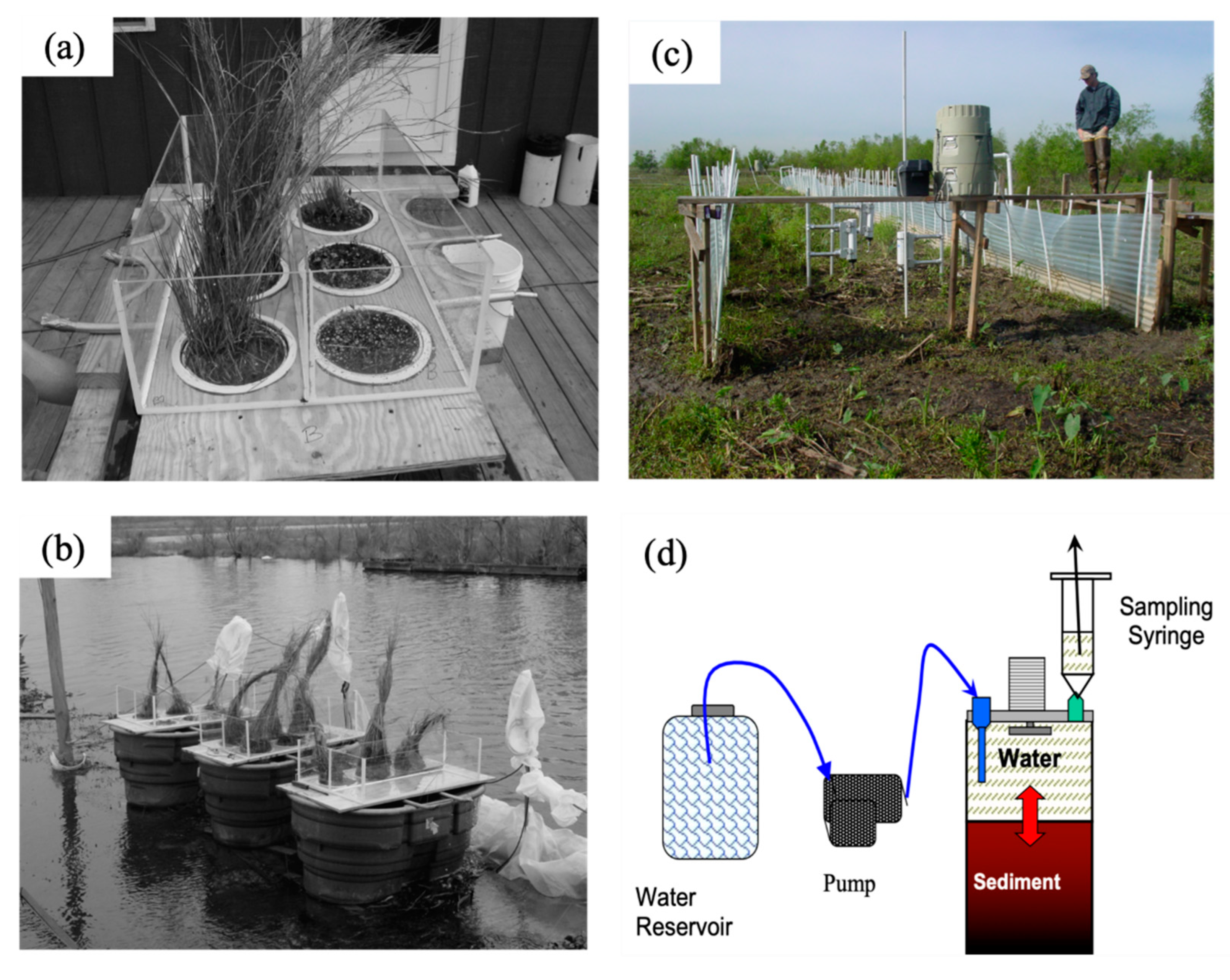
2.2. Subtidal Benthic Fluxes
2.3. Intertidal Benthic Fluxes
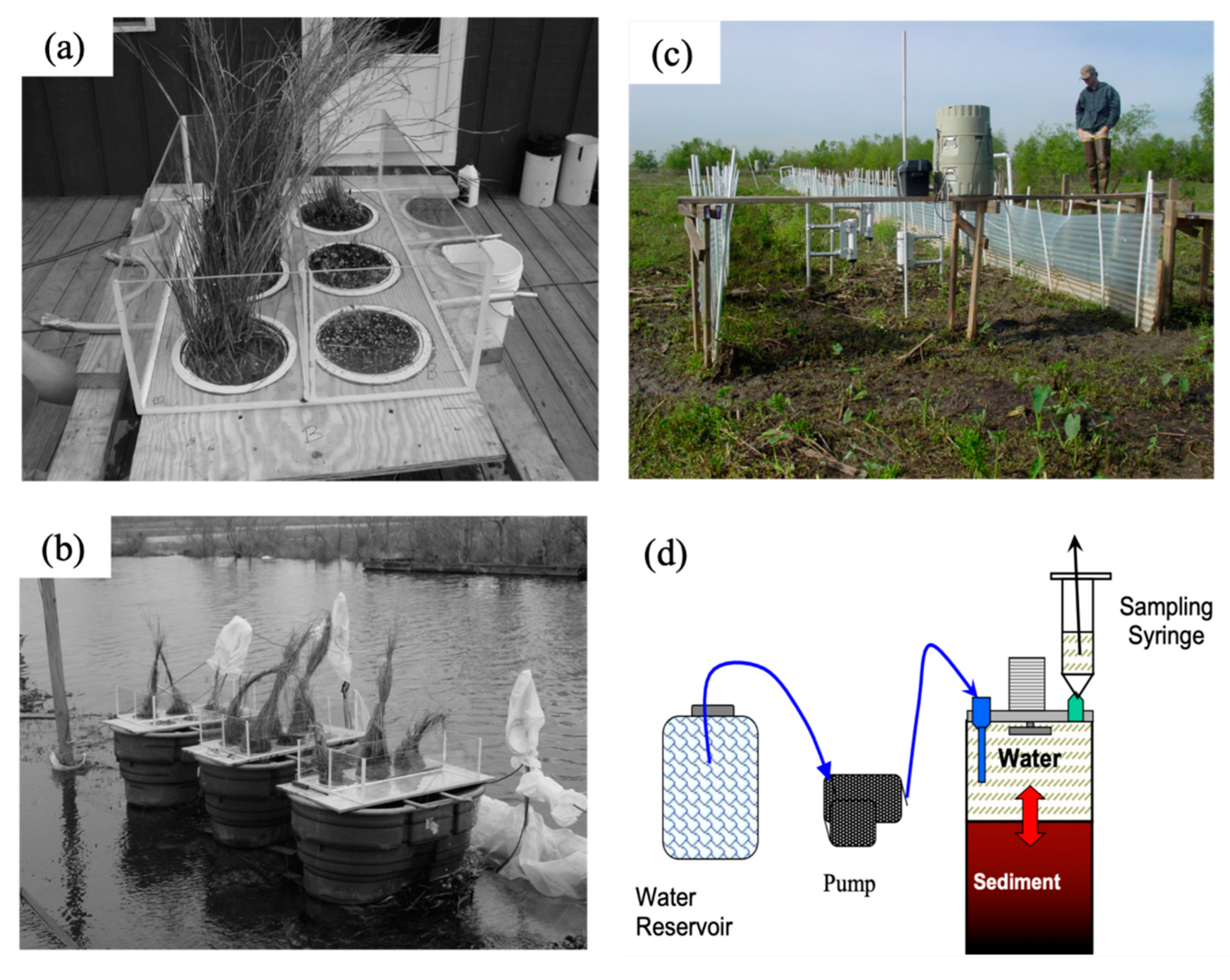
2.3.1. Flux Chambers
2.3.2. Flumes
2.4. Landscape Nutrient Surveys
2.5. Analytical Techniques
2.5.1. Sediment Samples
2.5.2. Water Samples
2.6. Statistical Analyses
2.6.1. Subtidal Benthic Fluxes
2.6.2. Intertidal Benthic Fluxes
2.6.3. Intertidal Flume Fluxes
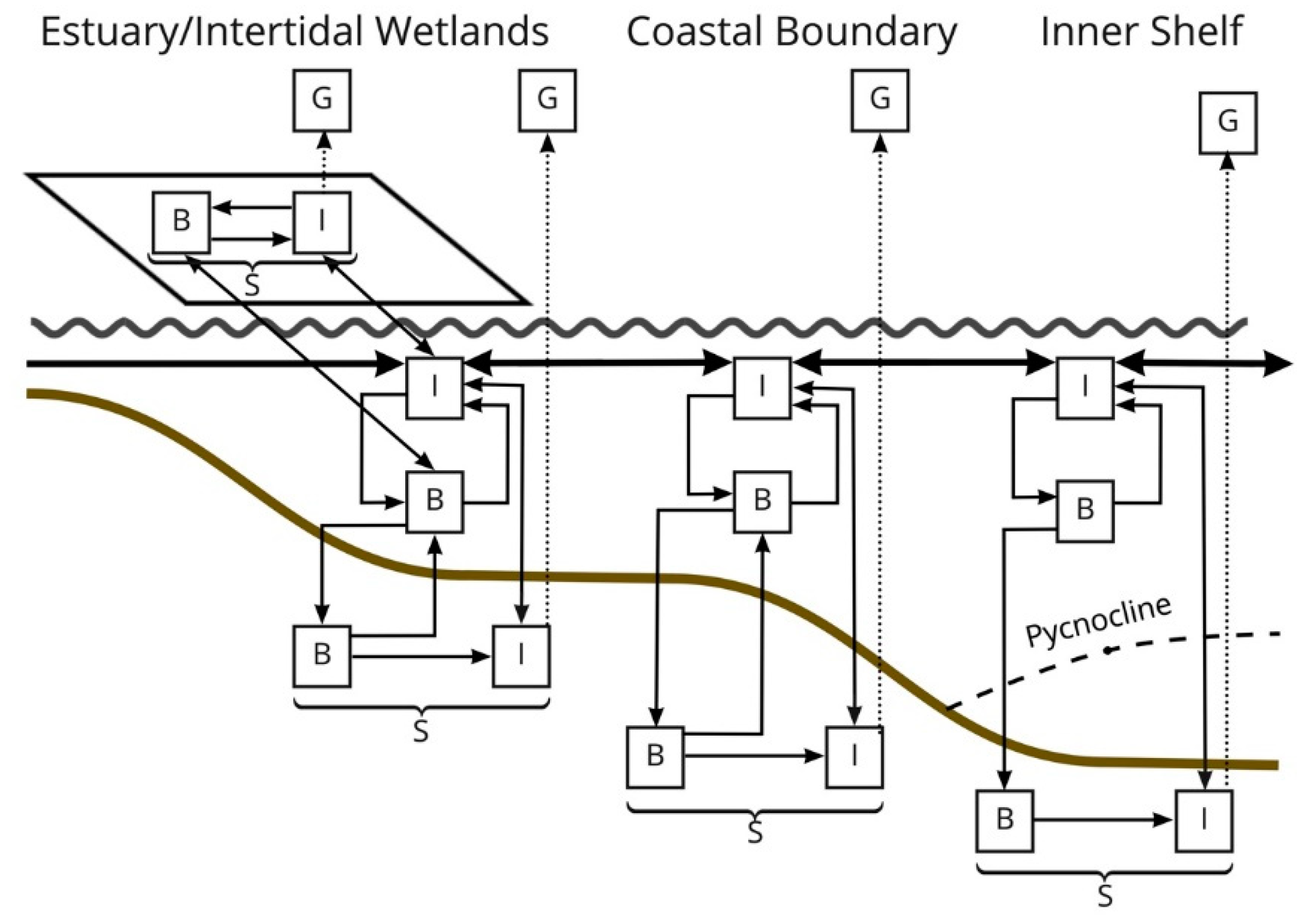
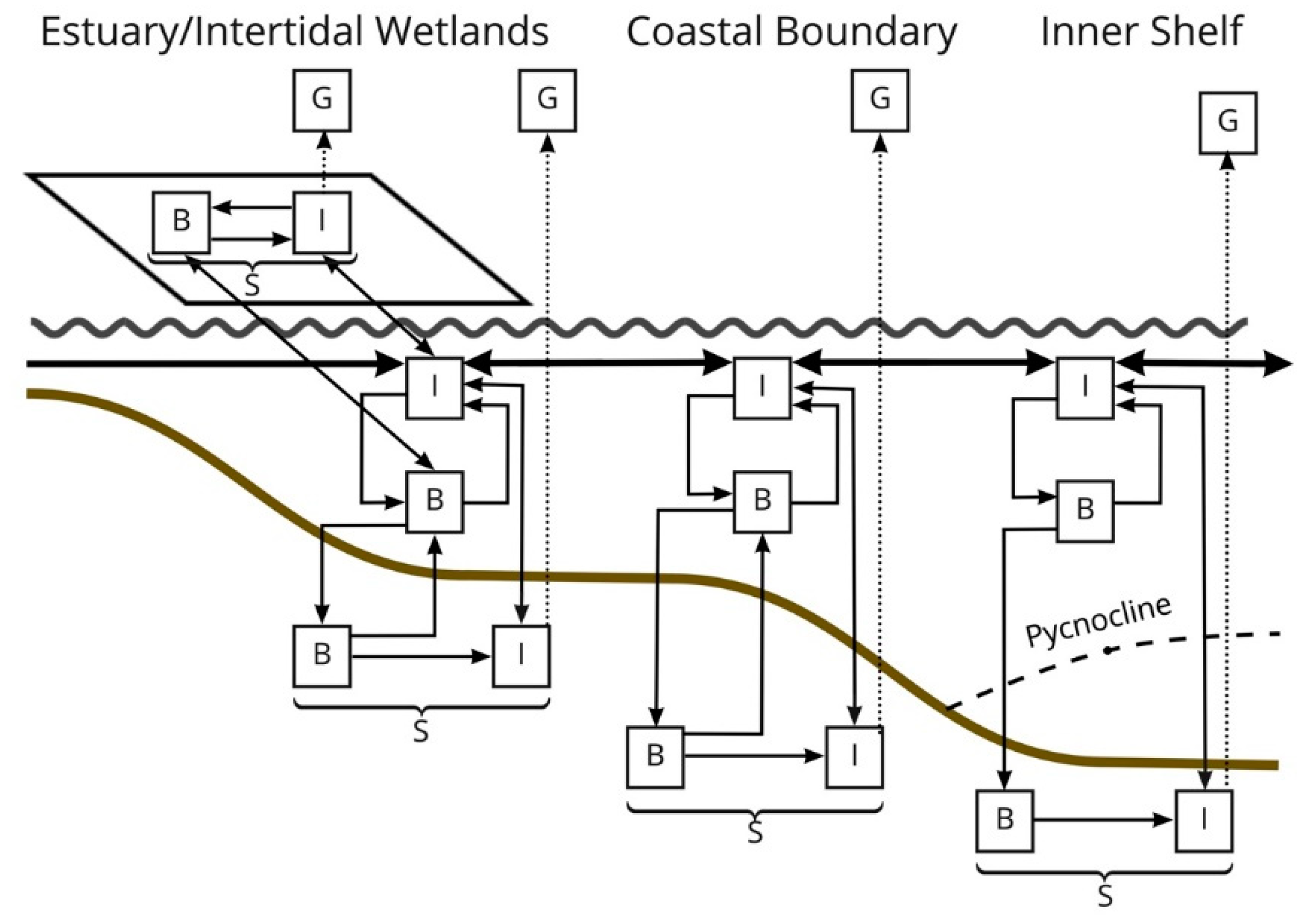
2.7. System Analyses
3. Results
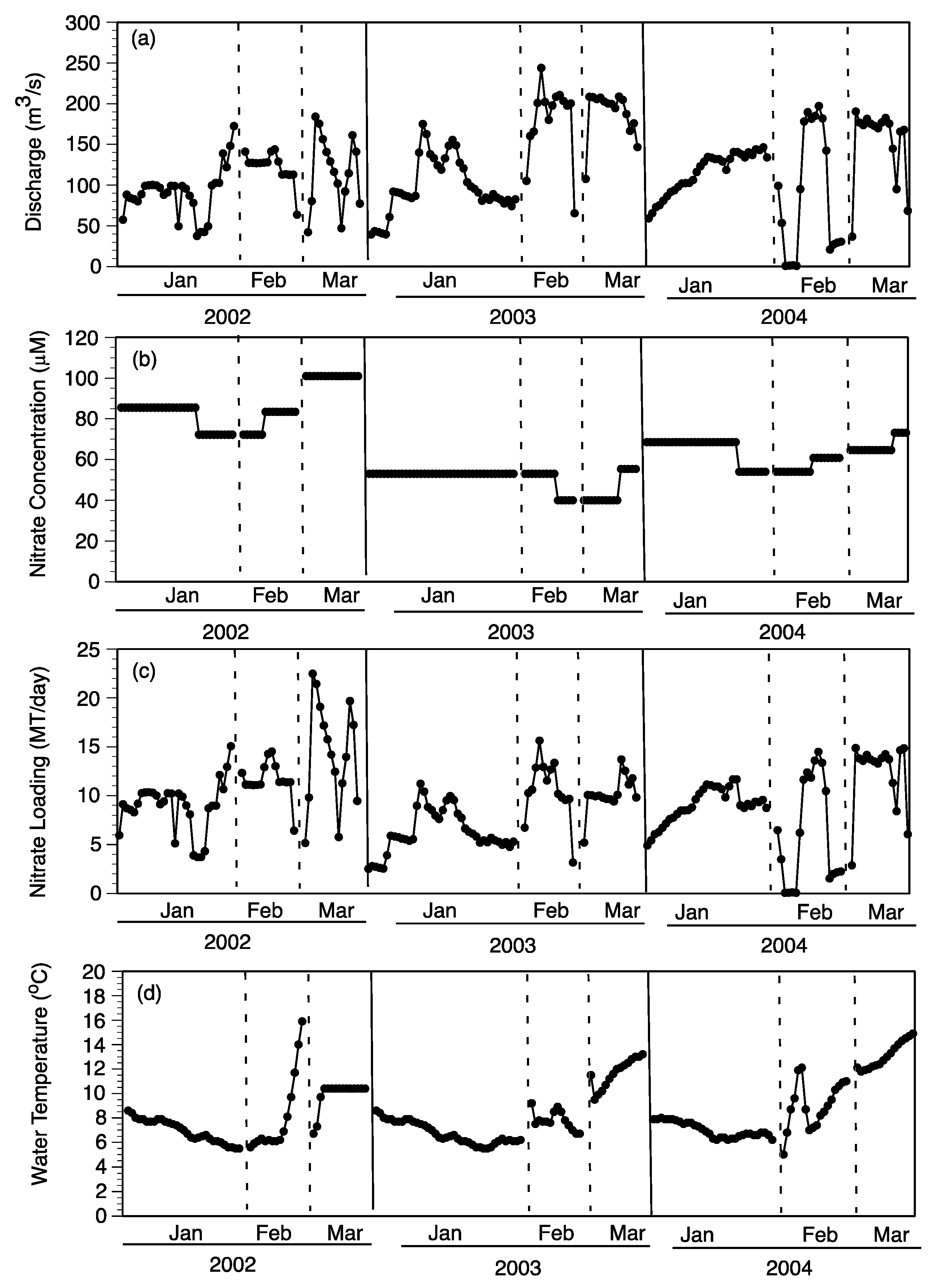
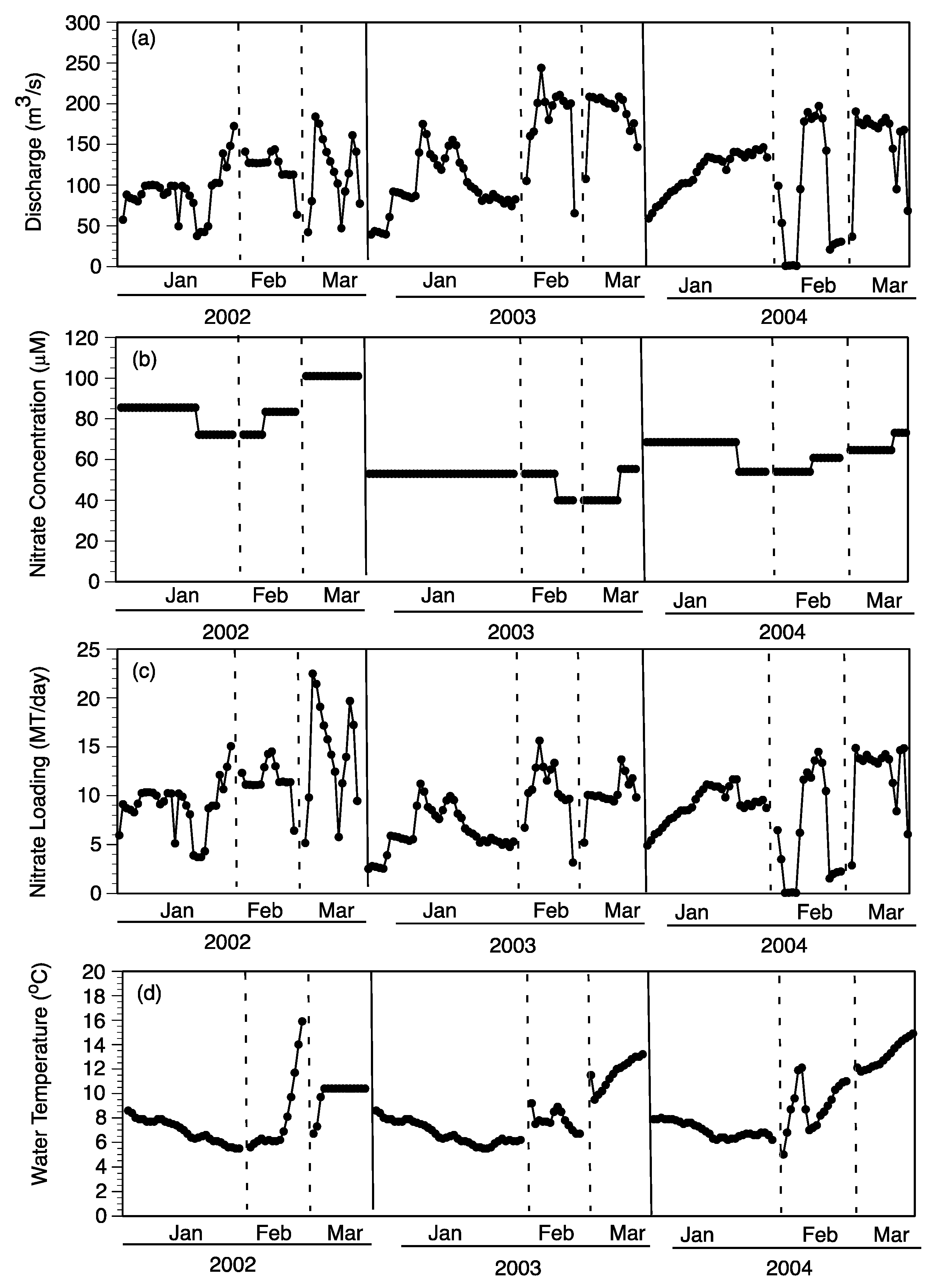
3.1. Experimental Field Conditions
3.1.1. Water Column
3.1.2. Sediments
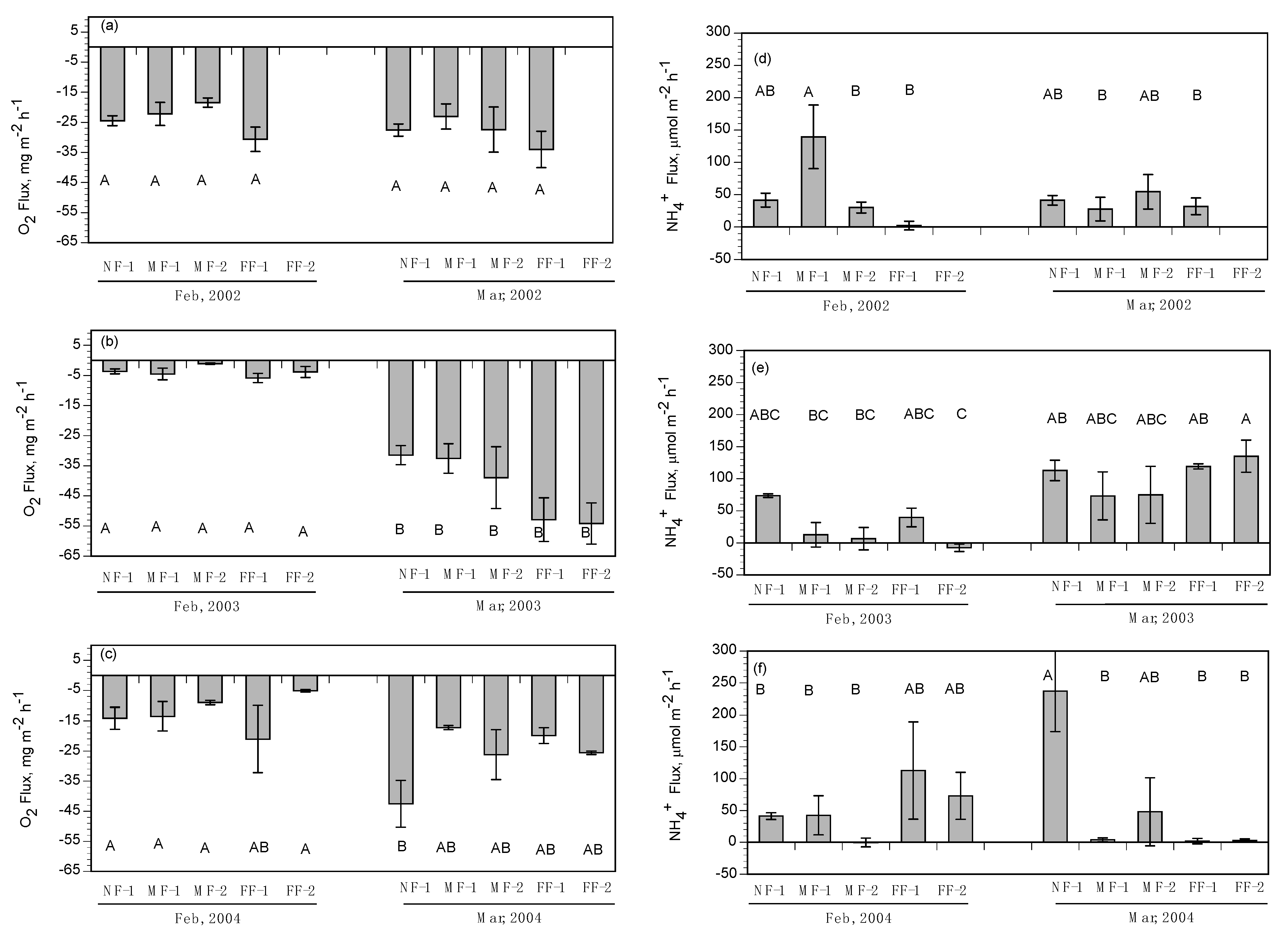
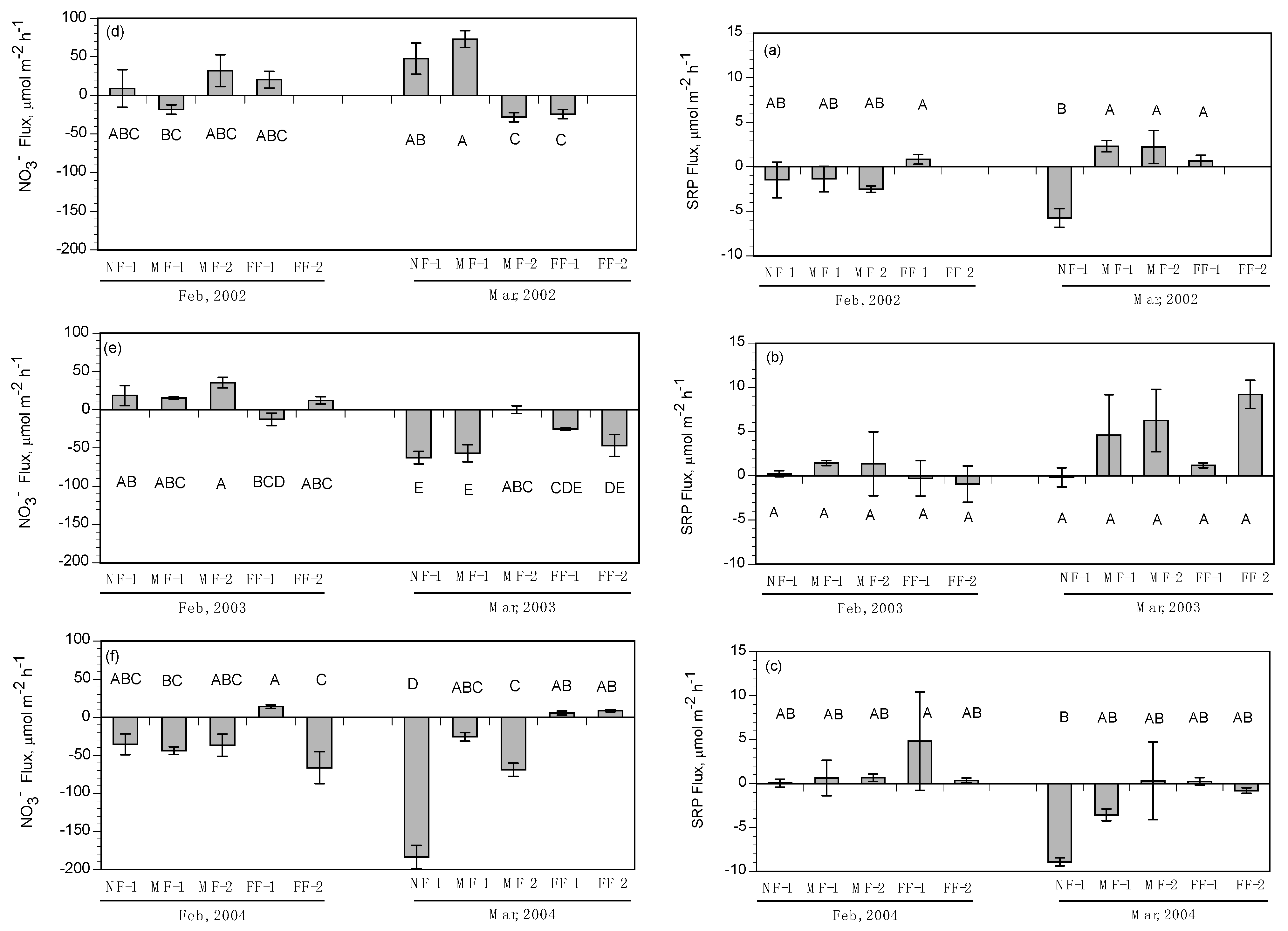
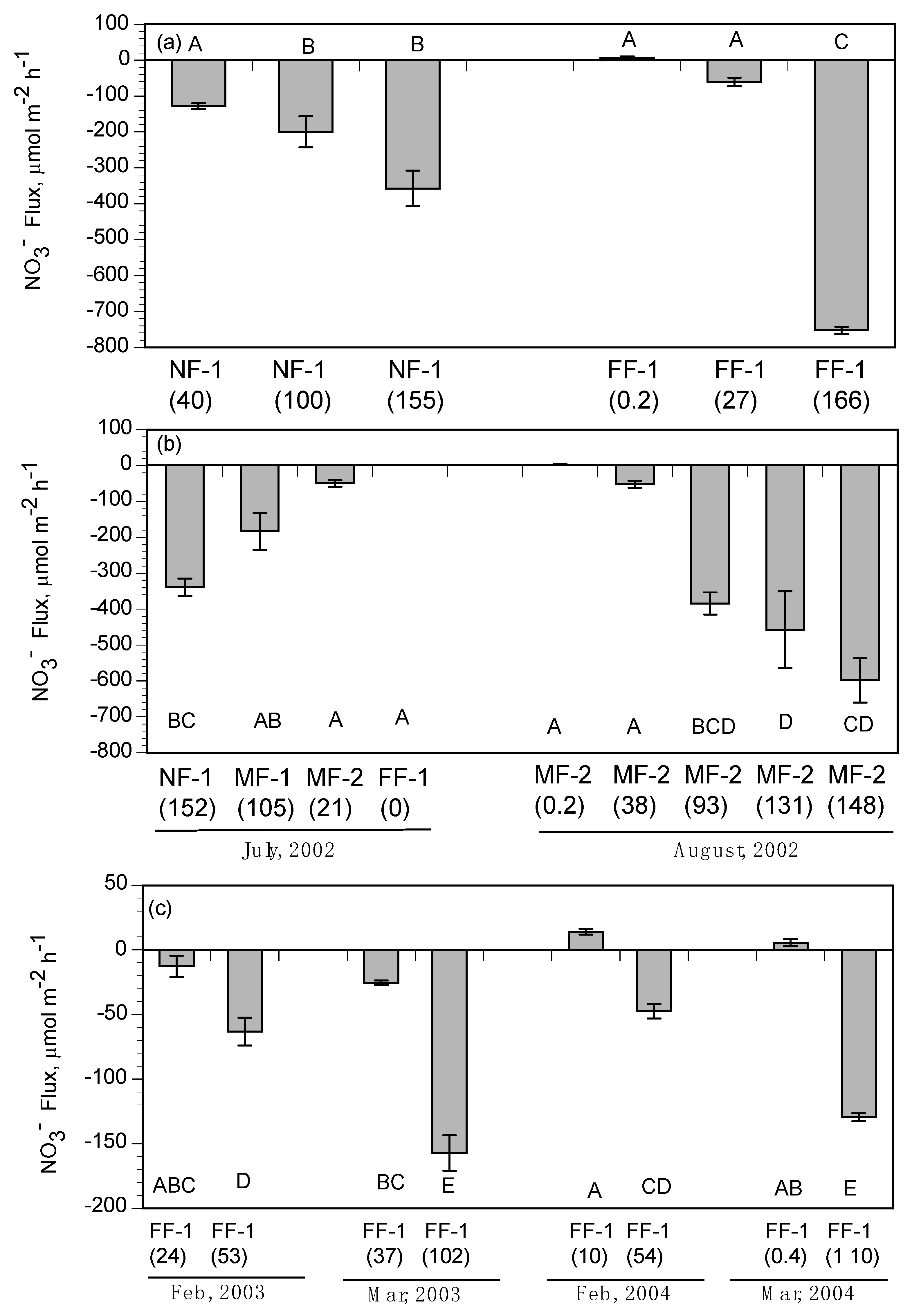
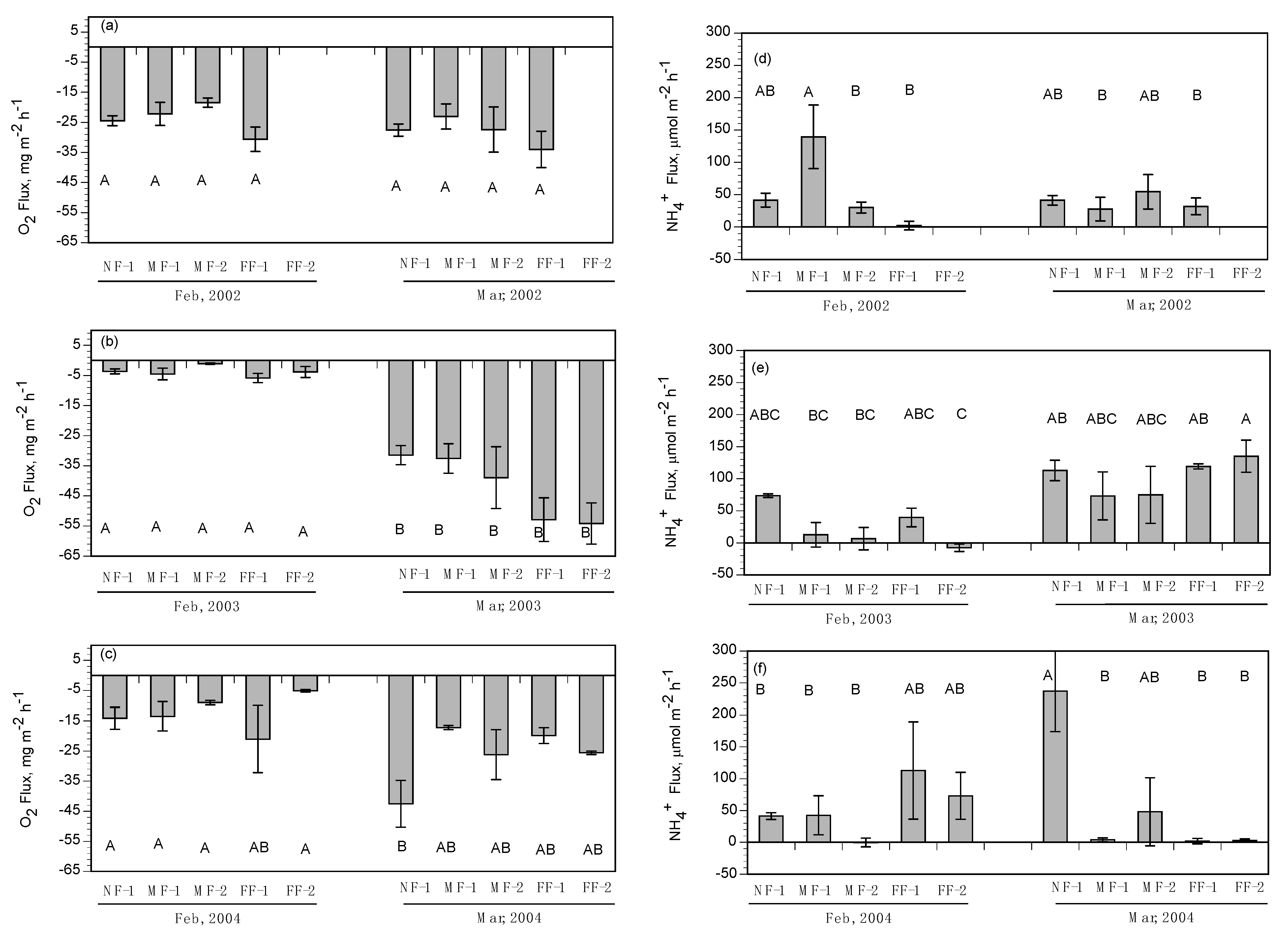
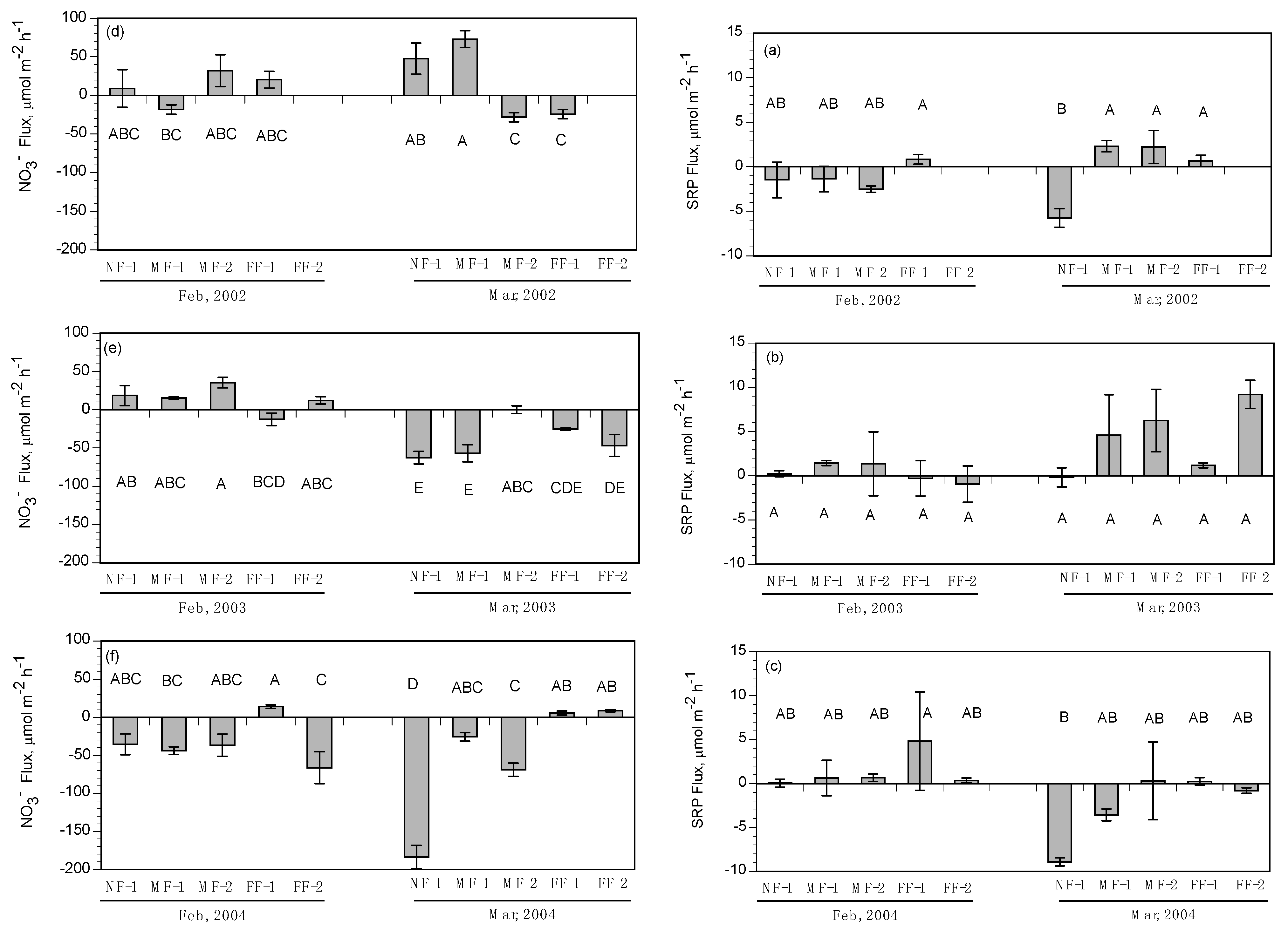
3.2. Subtidal Benthic Fluxes
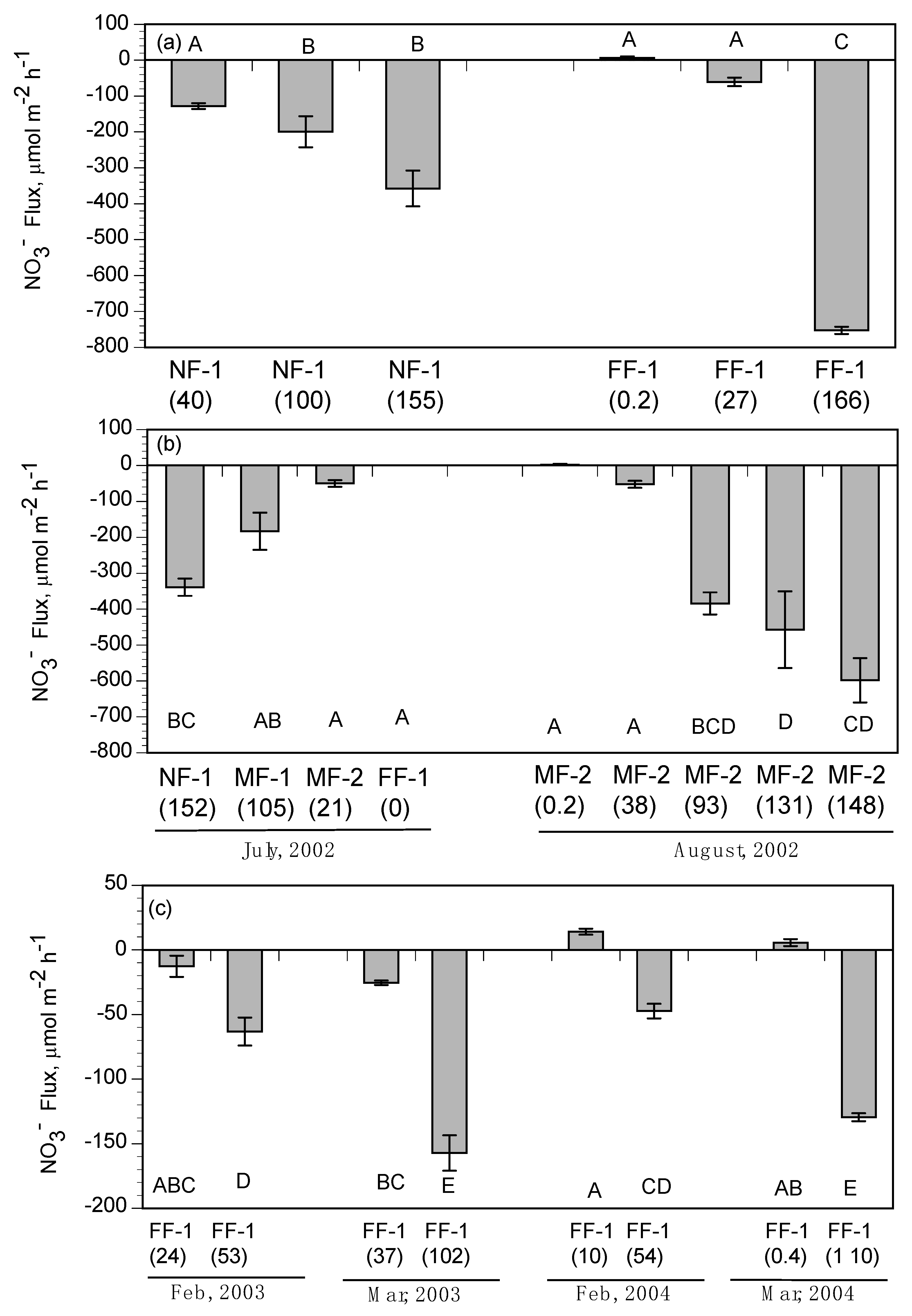
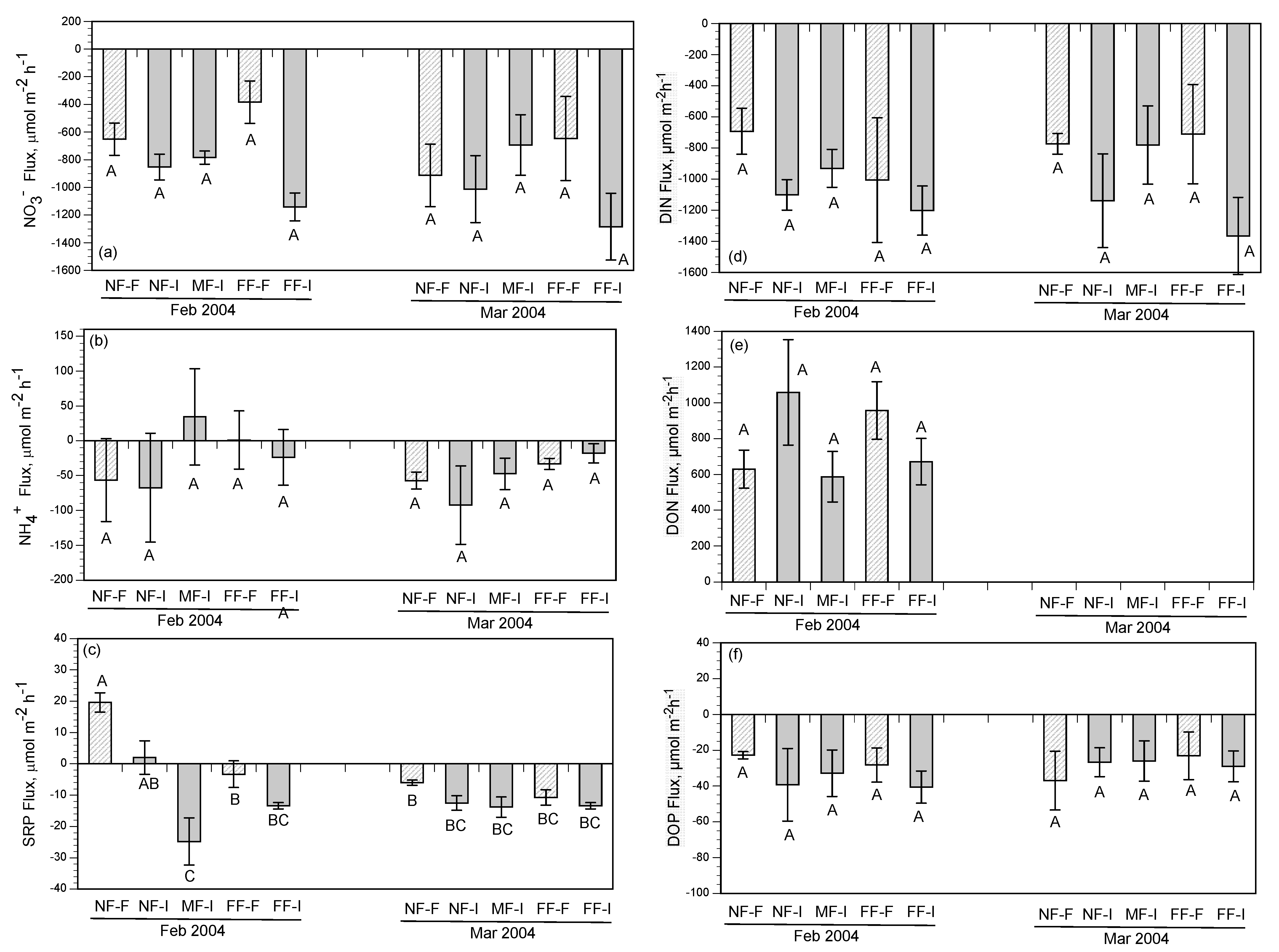
3.3. Intertidal Benthic Fluxes
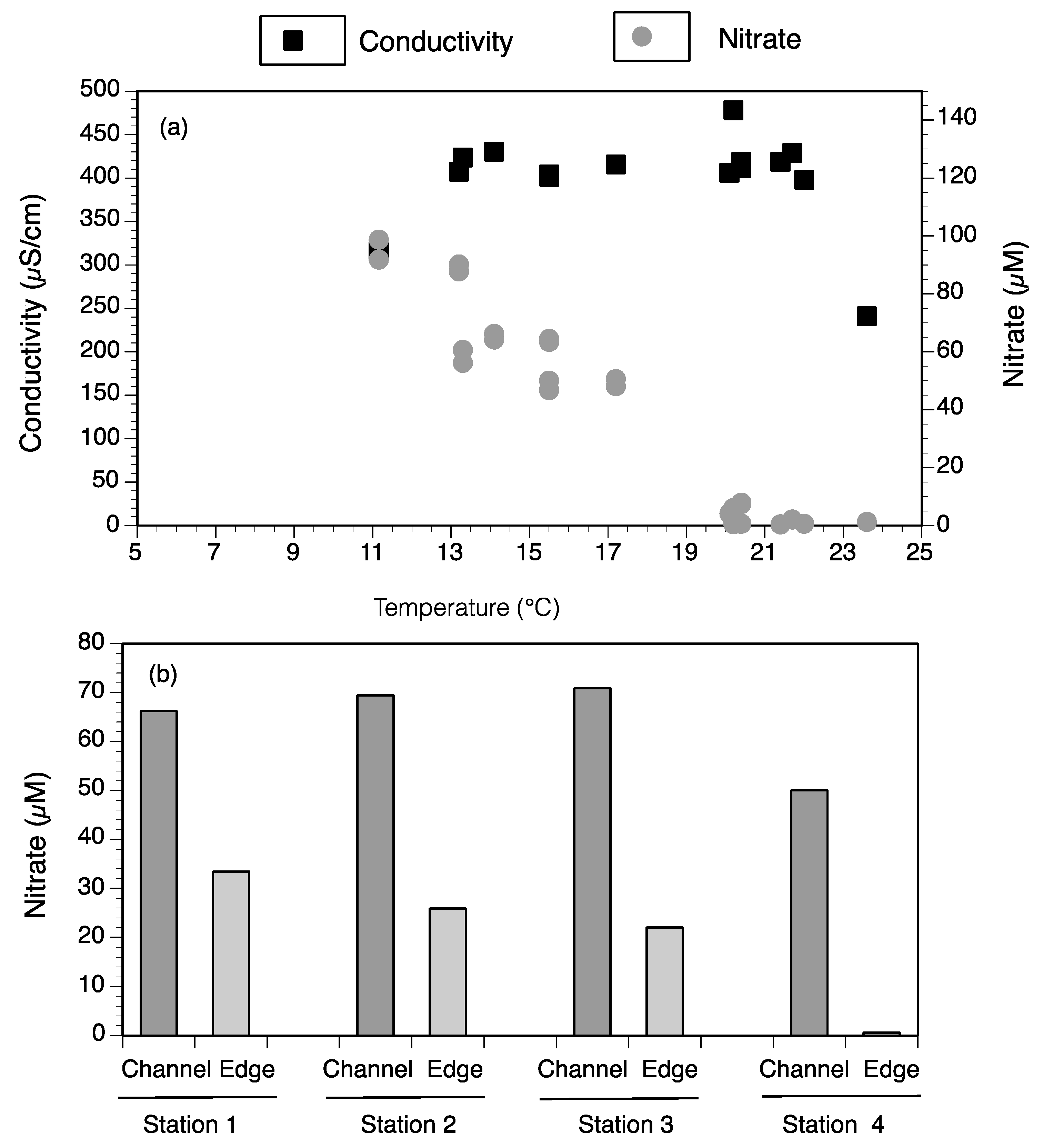
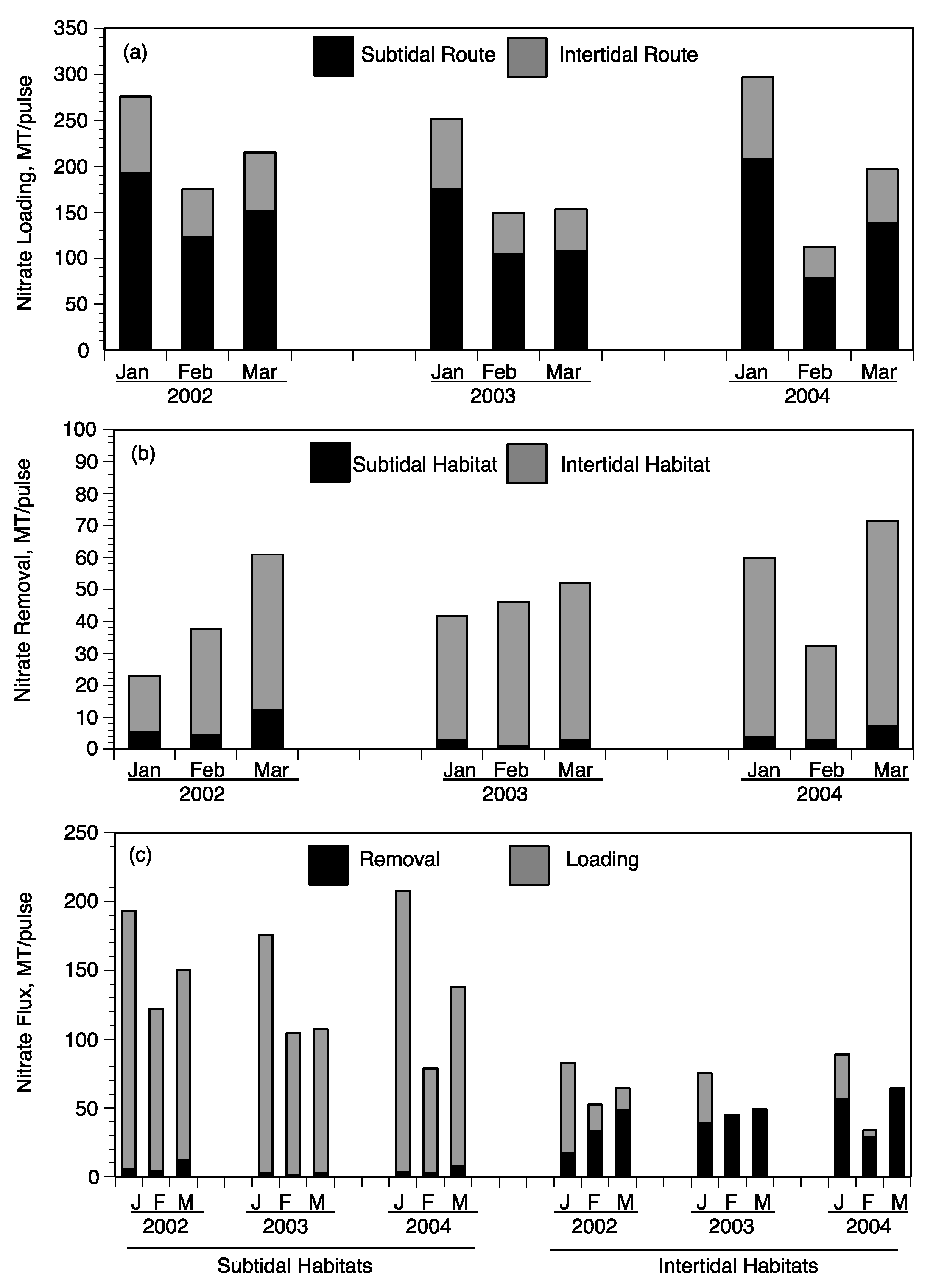
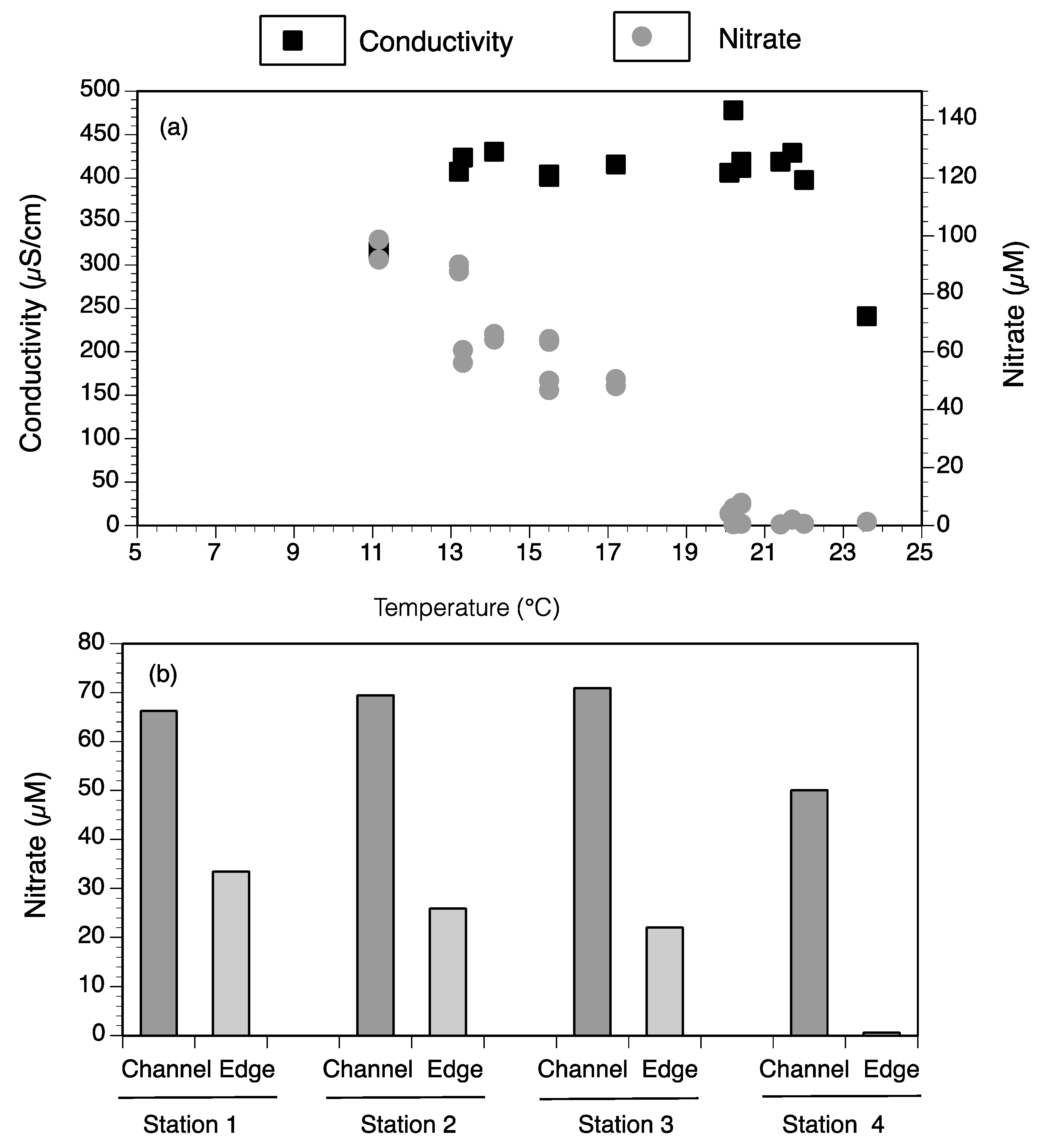
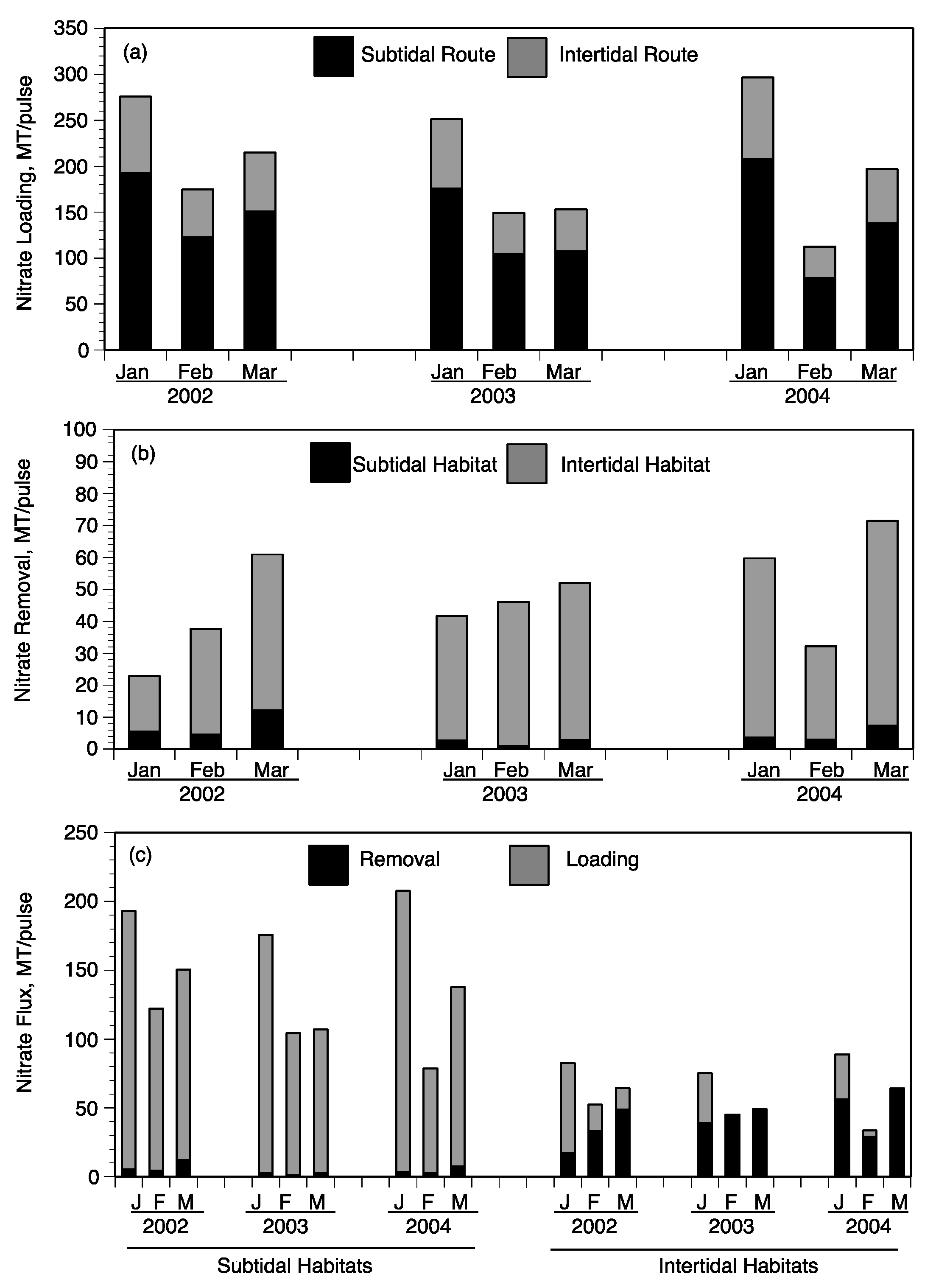
3.4. Landscape Nutrient Gradients
4. Discussion
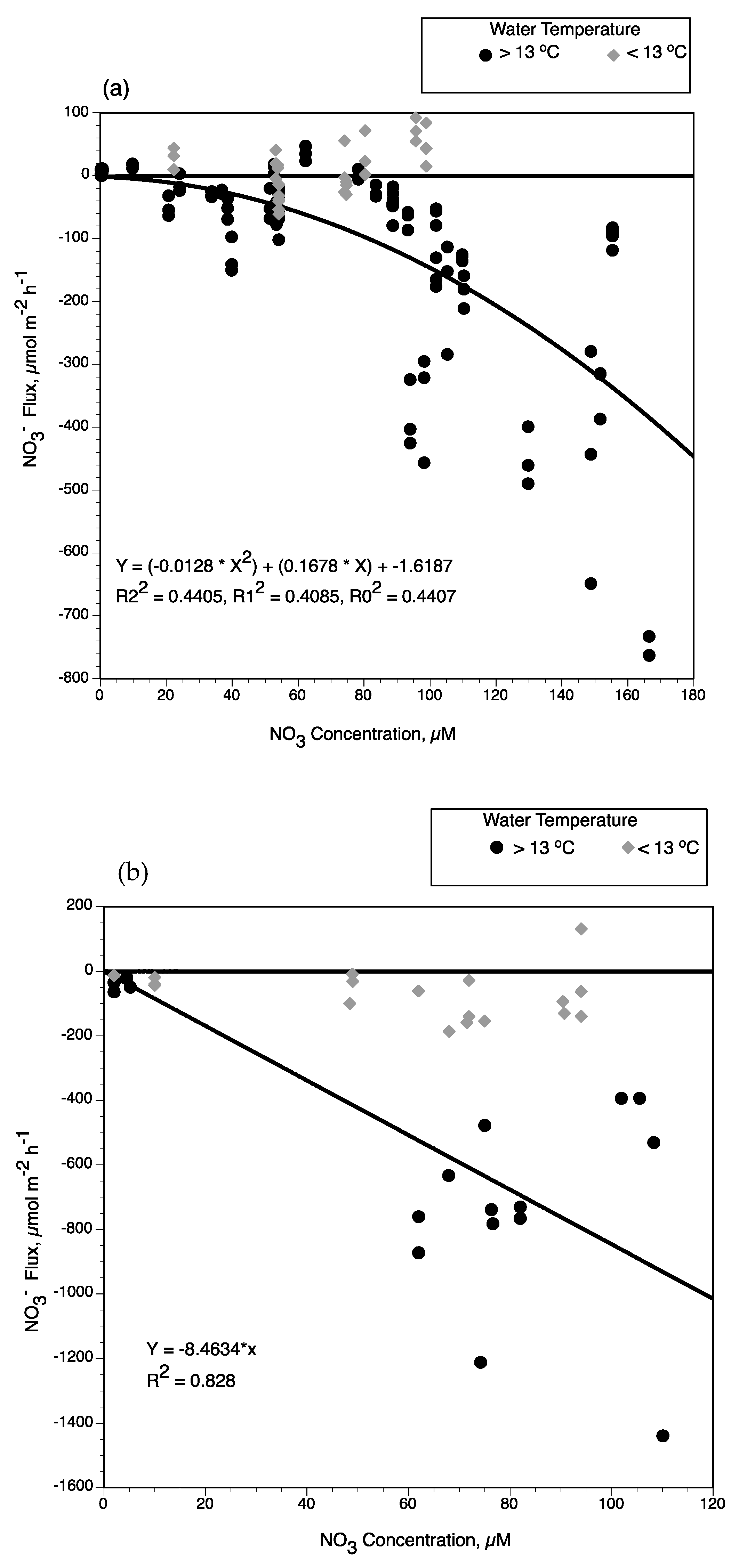
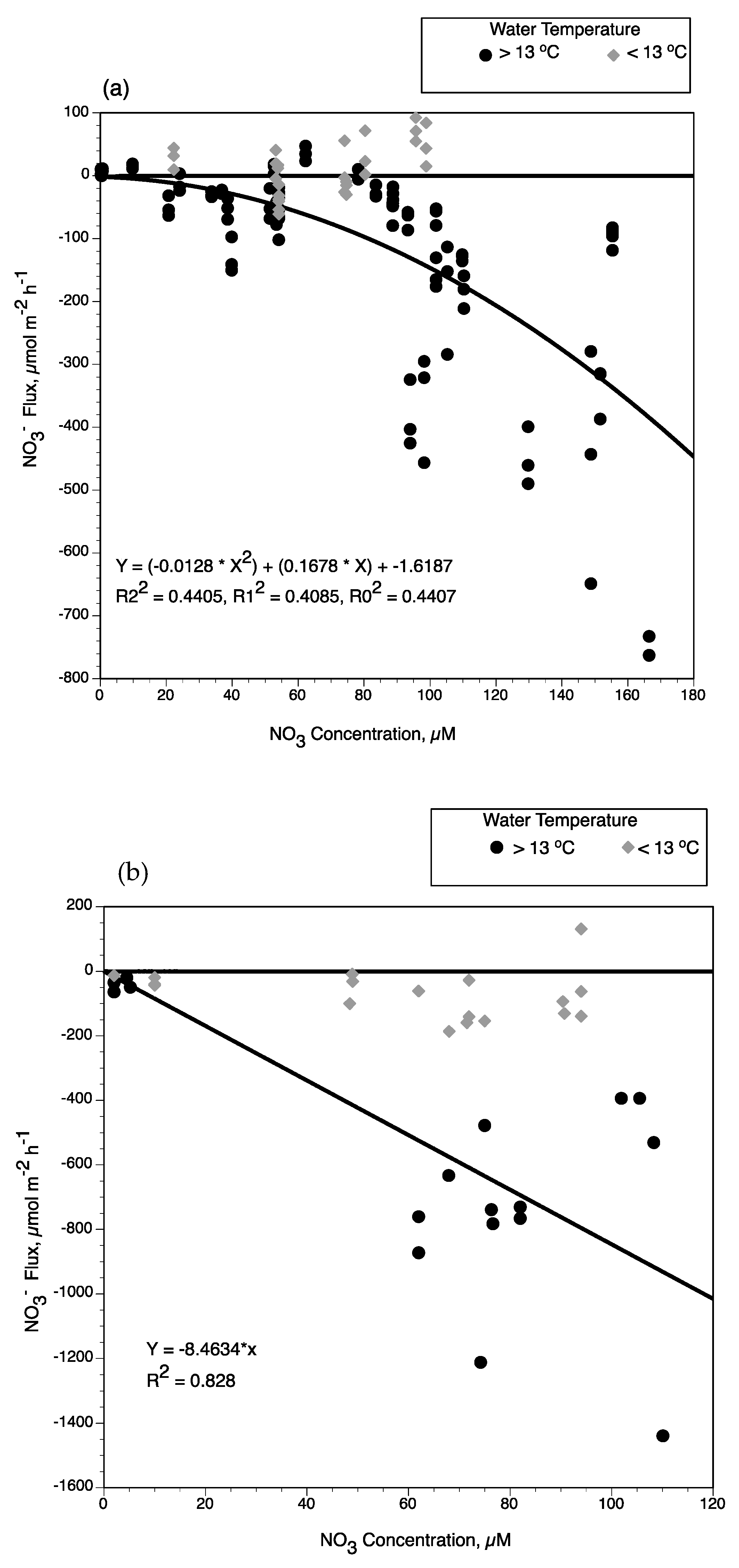
4.1. Factors Controlling Nitrogen Fluxes
4.2. Nitrate Removal
5. Conclusions-Ecosystem Design
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Day, J.W., Jr.; Boesch, D.F.F.; Clairain, E.J.J.; Kemp, G.P.P.; Laska, S.D.B.; Mitsch, W.J.J.; Orth, K.; Mashriqui, H.; Reed, D.J.J.; Shabman, L.; et al. Restoration of the Mississippi Delta: Lessons from Hurricanes Katrina and Rita. Science 2007, 315, 1679–1684. [Google Scholar] [CrossRef] [Green Version]
- Paola, C.; Twilley, R.R.; Edmonds, D.A.; Kim, W.; Mohrig, D.; Parker, G.; Viparelli, E.; Voller, V.R.V.R. Natural Processes in Delta Restoration: Application to the Mississippi Delta. Annu. Rev. Mar. Sci. 2011, 3, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Bayley, P.B. Understanding Large River: Floodplain Ecosystems. BioScience 1995, 45, 153–158. [Google Scholar] [CrossRef]
- Sparks, R.E. Need for ecosystem management of large rivers and their floodplains. BioScience 1995, 45, 168–182. [Google Scholar] [CrossRef]
- Twilley, R.R.; Bentley, S.J.; Chen, Q.; Edmonds, D.A.; Hagen, S.C.; Lam, N.S.N.; Willson, C.S.; Xu, K.; Braud, D.W.; Peele, H.R.; et al. Co-evolution of wetland landscapes, flooding, and human settlement in the Mississippi River Delta Plain. Sustain. Sci. 2016, 11, 711–731. [Google Scholar] [CrossRef] [Green Version]
- Day, J.W.; Cable, J.E.; Cowan, J.H.; DeLaune, R.; de Mutsert, K.; Fry, B.; Mashriqui, H.; Justic, D.; Kemp, P.; Lane, R.R.; et al. The Impacts of Pulsed Reintroduction of River Water on a Mississippi Delta Coastal Basin. Coast. Res. 2009, 10054, 225–243. [Google Scholar] [CrossRef] [Green Version]
- Vanote, R.L.; Minshall, W.G.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Johnson, B.L.; Richardson, W.B.; Naimo, T.J. Past, present, and future concepts in large river ecology: How rivers function and how human activities influence river processes. BioScience 1995, 45, 134–141. [Google Scholar] [CrossRef]
- DeLaune, R.; Sasser, C.; Evers-Hebert, E.; White, J.; Roberts, H. Influence of the Wax Lake Delta sediment diversion on aboveground plant productivity and carbon storage in deltaic island and mainland coastal marshes. Estuar. Coast. Shelf Sci. 2016, 177, 83–89. [Google Scholar] [CrossRef]
- Elsey-Quirk, T.; Graham, S.A.; Mendelssohn, I.A.; Snedden, G.; Day, J.W.; Twilley, R.; Shaffer, G.; Sharp, L.; Pahl, J.; Lane, R. Mississippi river sediment diversions and coastal wetland sustainability: Synthesis of responses to freshwater, sediment, and nutrient inputs. Estuar. Coast. Shelf Sci. 2019, 221, 170–183. [Google Scholar] [CrossRef]
- Twilley, R.; Day, J.; Bevington, A.; Castañeda-Moya, E.; Christensen, A.; Holm, G.; Heffner, L.; Lane, R.; McCall, A.; Aarons, A. Ecogeomorphology of coastal deltaic floodplains and estuaries in an active delta: Insights from the Atchafalaya Coastal Basin. Coast. Shelf Sci. 2019, 227, 106341. [Google Scholar] [CrossRef]
- White, J.R.; DeLaune, R.D.; Justic, D.; Day, J.W.; Pahl, J.; Lane, R.R.; Boynton, W.R.; Twilley, R.R. Consequences of Mississippi River diversions on nutrient dynamics of coastal wetland soils and estuarine sediments: A review. Estuar. Coast. Shelf Sci. 2019, 224, 209–216. [Google Scholar] [CrossRef]
- Snedden, G.A.; Cable, J.E.; Swarzenski, C.; Swenson, E. Sediment discharge into a subsiding Louisiana seltaic estuary through a Mississippi River diversion. Estuar. Coast. Shelf Sci. 2007, 71, 181–193. [Google Scholar] [CrossRef]
- Ou, Y.; Xue, Z.G.; Li, C.; Xu, K.; White, J.R.; Bentley, S.J.; Zang, Z. A numerical investigation of salinity variations in the Barataria Estuary, Louisiana in connection with the Mississippi River and restoration activities. Estuar. Coast. Shelf Sci. 2020, 245, 107021. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rivera-Monroy, V.H. Sediment and nutrient trade-offs in restoring Mississippi River Delta: Restoration versus eutrophicaion. J. Contemp. Water Res. Educ. 2009, 141, 1–6. [Google Scholar] [CrossRef]
- Bargu, S.; Justic, D.; White, J.R.; Lane, R.; Day, J.; Paerl, H.; Raynie, R. Mississippi River diversions and phytoplankton dynamics in deltaic Gulf of Mexico estuaries: A review. Estuar. Coast. Shelf Sci. 2019, 221, 39–52. [Google Scholar] [CrossRef]
- Riekenberg, J.; Bargu, S.; Twilley, R. Phytoplankton Community Shifts and Harmful Algae Presence in a Diversion Influenced Estuary. Estuaries Coasts 2014, 38, 2213–2226. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Turner, R.E.; Dortch, Q.; Justic, D.; Bierman, V.J., Jr.; Wiseman, W.J., Jr. Nutirent-enhanced productivity in the northern Gulf of Mexico: Past, present and future. Hydrobiologia 2002, 475, 39–63. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Turner, R.E.; Justić, D.; Dortch, Q.; Wiseman, W.J.; Gupta, B.K.S. Nutrient changes in the Mississippi River and system responses on the adjacent continental shelf. Estuaries 1996, 19, 386–407. [Google Scholar] [CrossRef]
- Broussard, W.; Turner, R.E. A century of changing land-use and water-quality relationships in the continental US. Front. Ecol. Environ. 2009, 7, 302–307. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Turner, R.E.; Scavia, D. Beyond Science into Policy: Gulf of Mexico Hypoxia and the Mississippi River: Nutrient policy development for the Mississippi River watershed reflects the accumulated scientific evidence that the increase in nitrogen loading is the primary factor in the worsening of hypoxia in the northern Gulf of Mexico. AIBS Bull. 2002, 52, 129–142. [Google Scholar]
- Lane, R.R.; Madden, C.J.; Day, J.W., Jr.; Solet, D.J. Hydrologic and nutrient dynamics of a coastal bay and wetland receiving discharge from the Atchafalaya River. Hydrobiologia 2011, 658, 55–66. [Google Scholar] [CrossRef]
- Roberts, B.J.; Doty, S.M. Spatial and temporal patterns of benthic respiration and net nutrient fluxes in the Atchafalaya River Delta Estuary. Estuaries Coasts 2015, 38, 1918–1936. [Google Scholar] [CrossRef]
- Christensen, A.; Twilley, R.R.; Willson, C.S.; Castañeda-Moya, E. Simulating hydrological connectivity and water age within a coastal deltaic floodplain of the Mississippi River Delta. Estuar. Coast. Shelf Sci. 2020, 245, 106995. [Google Scholar] [CrossRef]
- Li, S.; Christensen, A.; Twilley, R.R. Benthic fluxes of dissolved oxygen and nutrients across hydrogeomorphic zones in a coastal deltaic floodplain within the Mississippi River delta plain. Biogeochemistry 2020, 149, 115–140. [Google Scholar] [CrossRef]
- Lane, R.R.; Day, J.W., Jr. Water quality analysis of a freshwater diversion at Caernarvon, Louisiana. Estuaries 1999, 22, 327–336. [Google Scholar] [CrossRef]
- Smith, C.J.; DeLaune, R.D.; Patrick, W.H., Jr. Fate of riverine nitrate entering an estuary: I. Denitrification and nitrogen burial. Estuaries 1985, 8, 15–21. [Google Scholar] [CrossRef]
- Reddy, K.R.; Patrick, W.H., Jr. Effects of alternate aerobic and anaerobic conditions on redox potential, organic matter decomposition and nitrogen loss in a flooded soil. Soil Biol. Biochem. 1975, 7, 87–94. [Google Scholar] [CrossRef]
- Patrick, W.; Jugsujinda, A. Sequential reduction and oxidation of inorganic nitrogen, manganese, and iron in flooded soil. Soil Sci. Soc. Am. J. 1992, 56, 1071–1073. [Google Scholar] [CrossRef]
- Lane, R.R.; Day, J.W.; Justic, D.; Reyes, E.; Marx, B.; Day, J.N.; Hyfield, E. Changes in stoichiometric Si, N and P ratios of Mississippi River water diverted through coastal wetlands to the Gulf of Mexico. Estuar. Coast. Shelf Sci. 2004, 60, 1–10. [Google Scholar] [CrossRef]
- Okubo, A.; Levin, S.A. Diffusion and Ecological Problems: Modern Perspectives; Springer: New York, NY, USA, 2001. [Google Scholar]
- Schlichting, H.; Gersten, K. Boundary-Layer Theory; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Hiatt, M.; Castañeda-Moya, E.; Twilley, R.; Hodges, B.R.; Passalacqua, P. Channel-Island Connectivity Affects Water Exposure Time Distributions in a Coastal River Delta. Water Resour. Res. 2018, 54, 2212–2232. [Google Scholar] [CrossRef]
- Hiatt, M.; Passalacqua, P. Hydrological connectivity in river deltas: The first-order importance of channel-island exchange. Water Resour. Res. 2015, 51, 2264–2282. [Google Scholar] [CrossRef] [Green Version]
- Visser, J.M.; Sasser, C.E.; Chabreck, R.H.; Linscombe, R. Marsh vegetation types of the Mississippi River deltaic plain. Estuaries 1998, 21, 818–828. [Google Scholar] [CrossRef]
- Day, J.; Lane, R.; Moerschbaecher, M.; DeLaune, R.; Mendelssohn, I.; Baustian, J.; Twilley, R. Vegetation and Soil Dynamics of a Louisiana Estuary Receiving Pulsed Mississippi River Water Following Hurricane Katrina. Estuaries Coasts 2013, 36, 665–682. [Google Scholar] [CrossRef]
- Miller-Way, T.; Boland, G.S.; Rowe, G.T.; Twilley, R.R. Sediment Oxygen Consumption and Benthic Fluxes on the Louisiana Nutrient Shelf: A Continental Methodological Comparison. Estuaries 1994, 17, 809–815. [Google Scholar] [CrossRef]
- Miller-Way, T.; Twilley, R.R. Theory and operation of continuous flow systems for the study of benthic-pelagic coupling. Mar. Ecol. Prog. Ser. 1996, 140, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Cowan, J.L.; Pennock, J.R.; Boynton, W.R. Seasonal and interannual patterns of sedimentwater nutrient and oxygen fluxes in Mobile Bay, Alabama (USA): Regulating factors and ecological significance. Mar. Ecol. Prog. Ser. 1996, 141, 229–245. [Google Scholar] [CrossRef] [Green Version]
- Asmus, R.; Jensen, M.; Jensen, K.; Kristensen, E.; Asmus, H.; Wille, A. The role of water movement and spatial scaling for measurement of dissolved inorganic nitrogen fluxes in intertidal sediments. Estuar. Coast. Shelf Sci. 1998, 46, 221–232. [Google Scholar] [CrossRef]
- Childers, D.L.; Davis, S.E.; Twilley, R.; Rivera-Monroy, T.S. Wetland-water column interactions and the biogeochemistry of estuary-watershed coupling around the gulf of Mexico. Biochem. Gulf. Mex. Estuaries 1999, 73, 211–235. [Google Scholar]
- Childers, D.L.; Day, J.W., Jr. A flow-through flume technique for quantifying nutrient and materials fluxes in microtidal estuaries. Estuar. Coast. Shelf Sci. 1988, 27, 483–494. [Google Scholar] [CrossRef]
- Childers, D.L.; Day, J.W., Jr. Marsh-water column interactions in two Louisiana estuaries. II. Nutrient dynamics. Estuaries 1990, 13, 404–417. [Google Scholar] [CrossRef]
- Beyers, R.J.; Odum, H.T. Ecological Microcosms; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbooks of Sea-Water Analysis; Fisheries of Research Board of Canada: Ottawa, ON, Canada, 1972. [Google Scholar]
- Federation, W.E.; Association, A. Standard Methods for the Examination of Water and Wastewater; American Public Health Association (APHA): Washington, DC, USA, 2005. [Google Scholar]
- Institute, S. Base SAS 9.4 Procedures Guide; SAS Institute: Cary, NC, USA, 2015. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. SAS System for Mixed Models; SAS Institute: Cary, NC, USA, 1996. [Google Scholar]
- Megonigal, J.P.; Neubauer, S.C. Biogeochemistry of Tidal Freshwater Wetlands; Coastal Wetlands; Elsevier: Amsterdam, The Netherlands, 2019; pp. 641–683. [Google Scholar]
- Tobias, C.; Neubauer, S. Salt Marsh Biogeochemistry—An Overview. In COASTAL WETLANDS: An Integrated Ecosystem Approach; Elsevier: Cambridge, MA, USA, 2009; Volume 76, pp. 445–492. [Google Scholar] [CrossRef]
- Boynton, W.R.; Hagy, J.D.; Cornwell, J.C.; Kemp, W.M.; Greene, S.M.; Owens, M.S.; Baker, J.E.; Larsen, R.K. Nutrient budgets and management actions in the Patuxent River estuary, Maryland. Estuaries Coasts 2008, 31, 623–651. [Google Scholar] [CrossRef]
- DeLaune, R.; Jugsujinda, A. Denitrification potential in a Louisiana wetland receiving diverted Mississippi River water. Chem. Ecol. 2003, 19, 411–418. [Google Scholar] [CrossRef]
- Li, S.; Twilley, R.R. Nitrogen dynamics of inundated sediments in an emerging coastal deltaic floodplain in mississippi river delta using isotope pairing technique to test response to nitrate enrichment and sediment organic matter. Estuaries Coasts 2021, 1–17. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Day, J.W.; Zhang, L.; Lane, R.R. Nitrate-nitrogen retention in wetlands in the Mississippi River Basin. Ecol. Eng. 2005, 24, 267–278. [Google Scholar] [CrossRef]
- Henry, K.M.; Twilley, R.R. Nutrient biogeochemistry during the early stages of delta development in the Mississippi River deltaic plain. Ecosystems 2014, 17, 327–343. [Google Scholar] [CrossRef]
- Bevington, A.E.; Twilley, R.R. Island Edge Morphodynamics along a Chronosequence in a Prograding Deltaic Floodplain Wetland. J. Coast. Res. 2018, 34, 806–817. [Google Scholar] [CrossRef]
- Cornwell, J.C.; Kemp, W.M.; Kana, T.M. Denitrification in coastal ecosysems: Methods, environmental controlsl, and ecosystem level controls, a review. Aquat. Ecol. 1999, 33, 41–54. [Google Scholar] [CrossRef]
- Eyre, B.; France, L. Importance of marine imputs to the sediment and nutrient load of coastal-plain estuaries: A case study of Pumicestone Passage, south-eastern Queensland, Australia. Mar. Freshw. Res. 1997, 48, 277–286. [Google Scholar] [CrossRef]
- Giblin, A.E.; Hopkinson, C.S.; Tucker, J. Benthic metabolism and nutrient cycling in Boston Harbor, Massachusetts. Estuaries 1997, 20, 346–364. [Google Scholar] [CrossRef]
- Scaroni, A.E.; Nyman, J.A.; Lindau, C.W. Comparison of denitrification characteristics among three habitat types of a large river floodplain: Atchafalaya River Basin, Louisiana. Hydrobiologia 2011, 658, 17–25. [Google Scholar] [CrossRef]
- Childers, D.L. Fifteen Years of Marsh Flumes: A Review of Marsh-Water Column Interactions in Southeastern USA Estuaries; Global Wetlands: Old World and New; Elsevier Science: New York, NY, USA, 1994; pp. 277–293. [Google Scholar]
- Lundberg, C.J.; Lane, R.R.; Day, J.W., Jr. Spatial and temporal variations in nutrients and water-quality parameters in the Mississippi River-influenced Breton Sound Estuary. J. Coast. Res. 2014, 30, 328–336. [Google Scholar] [CrossRef]
- Reddy, K.; DeLaune, R.D. Biogeochemistry of Wetlands: Science and Applications; Handbook of Soil Science: Boca Roton, FL, USA, 2008. [Google Scholar] [CrossRef]
- Hyfield, E.C.; Day, J.W.; Cable, J.E.; Justic, D. The impacts of re-introducing Mississippi River water on the hydrologic budget and nutrient inputs of a deltaic estuary. Ecol. Eng. 2008, 32, 347–359. [Google Scholar] [CrossRef]
- Ross, M.R.V.; Bernhardt, E.S.; Doyle, M.W.; Heffernan, J.B. Designer Ecosystems: Incorporating Design Approaches into Applied Ecology. Annu. Rev. Environ. Resour. 2015, 40, 419–443. [Google Scholar] [CrossRef]
- Cahoon, D.R.; White, D.A.; Lynch, J.C. Sediment infilling and wetland formation dynamics in an active crevasse splay of the Mississippi River delta. Geomorphology 2011, 131, 57–68. [Google Scholar] [CrossRef]
- Bianchi, T.S.; Allison, M.A. Large-river delta-front estuaries as natural “recorders” of global environmental change. Proc. Natl. Acad. Sci. USA 2009, 106, 8085–8092. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subtidal Stations * | Subtidal Stations * | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Date | NF-1 | MF-1 | MF-2 | FF-1 | FF-2 | Date | NF-1 | MF-1 | MF-2 | FF-1 | FF-2 |
| Water Temperature (°C) | NO3− Concentration (µM) | ||||||||||
| Feb 2002 | 9.0 | 8.7 | 10.4 | 12.0 | nd ** | Feb 2002 | 74.1 | 74.7 | 80.3 | 22.3 | nd |
| Mar 2002 | 10.4 | 12.3 | 14.3 | 16.6 | nd | Mar 2002 | 98.8 | 95.7 | 88.8 | 33.8 | nd |
| Feb 2003 | 9.4 | 10.6 | 13.1 | 15.5 | 15.2 | Feb 2003 | 53.2 | 53.8 | 62.3 | 24 | 52.7 |
| Mar 2003 | 12.8 | 15.6 | 21.5 | 21.9 | 21.0 | Mar 2003 | 101.8 | 88.7 | 78.3 | 36.8 | 51.4 |
| Feb, 2004 | 7.6 | 8.4 | 13.2 | 13.7 | 13.0 | Feb, 2004 | 55.2 | 54.1 | 54.1 | 9.8 | 54.1 |
| Mar, 2004 | 13.1 | 17.5 | 18.9 | 20.1 | 21.6 | Mar, 2004 | 109.8 | 83.6 | 93.3 | 0.41 | 109.8 |
| Conductivity (µS/cm) | NH4+ Concentration (µM) | ||||||||||
| Feb 2002 | 327 | 339 | 408 | 716 | nd | Feb 2002 | 5.49 | 5.84 | 6.57 | 3.49 | nd |
| Mar 2002 | 473 | 456 | 435 | 794 | nd | Mar 2002 | 5.84 | 5.02 | 4.77 | 2.66 | nd |
| Feb 2003 | 333 | 363 | 418 | 665 | 482 | Feb 2003 | 5.67 | 5.78 | 7.21 | 1.45 | 5.26 |
| Mar 2003 | 319 | 322 | 326 | 630 | 393 | Mar 2003 | 1.12 | 0.92 | 0.62 | 0.33 | 0.23 |
| Feb, 2004 | 264 | 264 | 264 | 601 | 264 | Feb, 2004 | 4.35 | 4.55 | 4.55 | 0.6 | 4.55 |
| Mar, 2004 | 335 | 352 | 367 | 759 | 1605 | Mar, 2004 | 1.6 | 1.78 | 1.84 | 1.79 | 1.56 |
| Dissolved Oxygen (mg/L) | SRP Concentration (µM) | ||||||||||
| Feb 2002 | 11.17 | 10.76 | 11.05 | 10.96 | nd | Feb 2002 | 1.29 | 1.15 | 1.23 | 0.71 | nd |
| Mar 2002 | 9.67 | 9.89 | 9.85 | 10.26 | nd | Mar 2002 | 1.69 | 1.09 | 1.45 | 0.85 | nd |
| Feb 2003 | 9.19 | 9.53 | 9.35 | 9.56 | 9.38 | Feb 2003 | 0.75 | 0.84 | 1.20 | 0.63 | 1.06 |
| Mar 2003 | 9.34 | 9.21 | 9.54 | 9.36 | 9.14 | Mar 2003 | 1.36 | 1.38 | 1.22 | 0.36 | 0.61 |
| Feb, 2004 | nd | 10.43 | 10.43 | 10.88 | 10.43 | Feb, 2004 | 1.01 | 0.95 | 0.95 | 0.85 | 0.95 |
| Mar, 2004 | 8.59 | 9.06 | 8.87 | 8.78 | 8.62 | Mar, 2004 | 1.7 | 1.58 | 1.44 | 0.6 | 1.66 |
| Habitat Type | Total | Near-Field | Mid-Field | Far-Field |
|---|---|---|---|---|
| Area (ha) | Area (ha) | Area (ha) | Area (ha) | |
| Outfall Region | 33,288 | 4813 | 14,495 | 13,980 |
| Subtidal | 6593 | 752 | 2517 | 3324 |
| Intertidal | 26,695 | 4061 | 11,978 | 10,656 |
| Subtidal Cores * | Intertidal Cores ** | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NF-1 | MF-1 | MF-2 | FF-1 | FF-2 | NF-F | NF-I | MF-I | FF-F | FF-I | |
| Latitude | 29°82.94.2 N | 29°79.31.4 N | 29°79.29.8 N | 29°71.14.9 N | 29°71.14.3 N | 29°50.10.68 N | 29°46.59.54 N | 29°42.17.85 N | ||
| Longitude | 89°89.55.3 W | 89°86.32.3 W | 89°80.13.7 W | 89°83.94.3 W | 89°80.41.2 W | 89°54.38.16 W | 89°56.50.51 W | 89°54.10.60 W | ||
| Water Temperature (°C) | ||||||||||
| Feb, 2004 | 7.6 | 8.4 | 13.2 | 13.7 | 13.0 | 24.2 | 24.0 | 23.9 | 23.7 | 23.8 |
| Mar, 2004 | 13.1 | 17.5 | 18.9 | 20.1 | 21.6 | 22.8 | 22.8 | 22.6 | 22.8 | 22.4 |
| Conductivity (µS/cm) | ||||||||||
| Feb, 2004 | 264 | 264 | 264 | 601 | 264 | nd *** | nd | nd | nd | nd |
| Mar, 2004 | 335 | 352 | 367 | 759 | 1605 | nd | nd | nd | nd | nd |
| NO3− Concentration (µM) | ||||||||||
| Feb, 2004 | 55.2 | 54.1 | 54.1 | 9.8 | 54.1 | 27.0 | 46.1 | 48.2 | 42.9 | 45.6 |
| Mar, 2004 | 109.8 | 83.6 | 93.3 | 0.41 | 109.8 | 54.7 | 55.4 | 55.8 | 59.6 | 53.7 |
| NH4+ Concentration (µM) | ||||||||||
| Feb, 2004 | 4.35 | 4.55 | 4.55 | 0.6 | 4.55 | 3.73 | 5.16 | 4.7 | 4.3 | 4.71 |
| Mar, 2004 | 1.6 | 1.78 | 1.84 | 1.79 | 1.56 | 2.81 | 5.22 | 3.62 | 2.7 | 2.74 |
| SRP Concentration (µM) | ||||||||||
| Feb, 2004 | 1.01 | 0.95 | 0.95 | 0.85 | 0.95 | 0.62 | 0.64 | 0.73 | 0.61 | 0.72 |
| Mar, 2004 | 1.7 | 1.58 | 1.44 | 0.6 | 1.66 | 0.77 | 0.87 | 0.74 | 0.58 | 0.71 |
| Near-Field Flume | Mid-Field Flume | ||
|---|---|---|---|
| Parameter | February 2004 Pulse | March 2004 Pulse | March 2004 Pulse |
| Physical Parameters | |||
| Temperature (°C) | 11.3 | 15.0 | 20.8 |
| Water Depth (m) | 0.31 | 0.34 | 0.30 |
| Flow (cm/s) | 3.03 | 2.33 | 0.22 |
| Conductivity (µS/cm) | 360 | 990 | 780 |
| Reynolds Number | 6627 | 7847 | 601 |
| Residence Time (h) | 0.92 | 1.22 | 19.7 |
| Nutrient Concentrations | |||
| 52.99 | 106.07 | 0.20 | |
| (SE) | (2.30) | (1.62) | (0.00) |
| 0.77 | 1.02 | 0.21 | |
| (SE) | (0.04) | (0.03) | (0.01) |
| 3.19 | 1.97 | 1.49 | |
| (SE) | (0.10) | (0.10) | (0.12) |
| 59.36 | 108.97 | 1.58 | |
| (SE) | (1.91) | (1.61) | (0.13) |
| 83.43 | 133.45 | 36.26 | |
| (SE) | (0.25) | (1.92) | (0.08) |
| 86.21 | 107.65 | 43.05 | |
| (SE) | (2.40) | (3.84) | (1.41) |
| 0.50 | 0.70 | 0.31 | |
| (SE) | (0.03) | (0.02) | (0.02) |
| 2.03 | 2.25 | 0.84 | |
| (SE) | (0.03) | (0.05) | (0.10) |
| 2.42 | 2.85 | 1.20 | |
| (SE) | (0.05) | (0.02) | (0.04) |
| 75.39 | 88.43 | 18.50 | |
| (SE) | (1.44) | (1.05) | (0.81) |
| Intertidal Cores * | |||||
|---|---|---|---|---|---|
| NF-F | NF-I | MF-I | FF-F | FF-I | |
| Sediment Characteristics | |||||
| Bulk Density (g/cm3) | |||||
| X | 0.429 | 0.311 | 0.141 | 0.136 | 0.173 |
| (SE) | (0.026) | (0.022) | (0.009) | (0.004) | (0.013) |
| Carbon Content (mg/gdm) | |||||
| X | 85.8 | 117.8 | 338.5 | 351.8 | 258.3 |
| (SE) | (2.6) | (8.7) | (31.6) | (9.9) | (12.9) |
| Nitrogen Content (mg/gdm) | |||||
| X | 7.7 | 7.4 | 24.7 | 22.4 | 16.2 |
| (SE) | (0.2) | (0.3) | (3.5) | (0.6) | (0.4) |
| C:N Ratio | 11.1 | 16.0 | 13.7 | 15.7 | 16.0 |
| Month | Total Input | Subtidal Input (Channel Flow) | Intertidal Input (Residual Flow) | Subtidal Removal | Intertidal Removal | Total NO3− Removal |
|---|---|---|---|---|---|---|
| MT/Pulse | MT/Pulse | MT/Pulse | % of Input | % of Input | % of Total Input | |
| 2002 | ||||||
| Pulse Jan | 275.61 | 192.93 | 82.68 | −2.0% | −6.3% | −8.3% |
| Pulse Feb | 174.67 | 122.27 | 52.40 | −3.7% | −63.4% | −21.6% |
| Pulse Mar | 214.97 | 150.48 | 64.49 | −8.1% | −75.6% | −28.3% |
| 2003 | ||||||
| Pulse Jan | 251.18 | 175.83 | 75.36 | −1.1% | −15.5% | −16.6% |
| Pulse Feb | 149.08 | 104.36 | 44.73 | −0.9% | −101.0% | −30.9% |
| Pulse Mar | 152.97 | 107.08 | 45.89 | −2.7% | −107.3% | −34.1% |
| 2004 | ||||||
| Pulse Jan | 296.68 | 207.68 | 89.00 | −1.2% | −18.9% | −20.1% |
| Pulse Feb | 112.25 | 78.58 | 33.68 | −3.7% | −86.9% | −28.7% |
| Pulse Mar | 196.94 | 137.86 | 59.08 | −5.4% | −108.5% | −36.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Twilley, R.R.; Rick, S.; Bond, D.C.; Baker, J. Benthic Nutrient Fluxes across Subtidal and Intertidal Habitats in Breton Sound in Response to River-Pulses of a Diversion in Mississippi River Delta. Water 2021, 13, 2323. https://doi.org/10.3390/w13172323
Twilley RR, Rick S, Bond DC, Baker J. Benthic Nutrient Fluxes across Subtidal and Intertidal Habitats in Breton Sound in Response to River-Pulses of a Diversion in Mississippi River Delta. Water. 2021; 13(17):2323. https://doi.org/10.3390/w13172323
Chicago/Turabian StyleTwilley, Robert R., Silke Rick, Daniel C. Bond, and Justin Baker. 2021. "Benthic Nutrient Fluxes across Subtidal and Intertidal Habitats in Breton Sound in Response to River-Pulses of a Diversion in Mississippi River Delta" Water 13, no. 17: 2323. https://doi.org/10.3390/w13172323
APA StyleTwilley, R. R., Rick, S., Bond, D. C., & Baker, J. (2021). Benthic Nutrient Fluxes across Subtidal and Intertidal Habitats in Breton Sound in Response to River-Pulses of a Diversion in Mississippi River Delta. Water, 13(17), 2323. https://doi.org/10.3390/w13172323